
Vasodilator Responses of Perivascular Adipose Tissue-Derived Hydrogen Sulfide Stimulated with L-Cysteine in Pregnancy Hypertension-Induced Endothelial Dysfunction in Rats
 and
and
Abstract
:
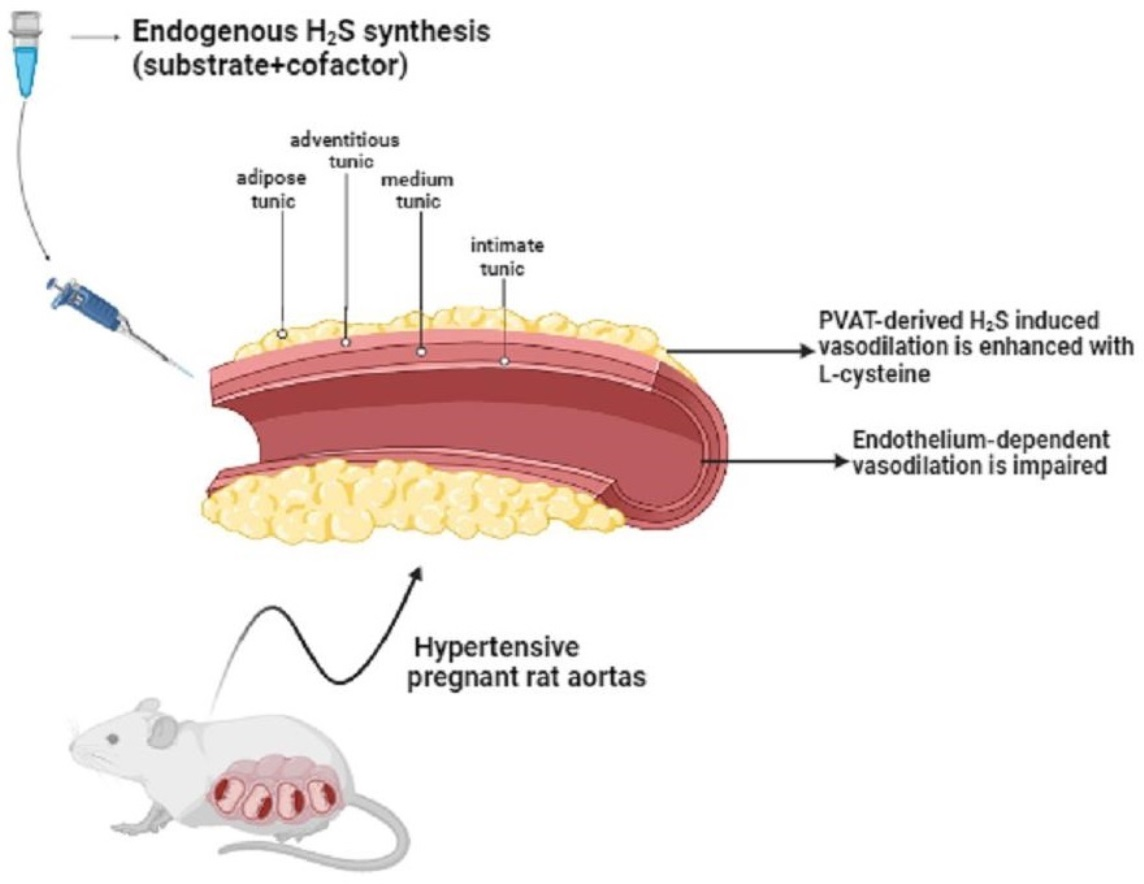{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Euthanasia, Blood Sampling, and Fetal and Placental Parameters
2.3. Systolic Blood Pressure Measurements
2.4. Vascular Reactivity Experiments
2.5. Determination of Fms-like Tyrosine Kinase 1 (sFlt-1) and Placental Growth Factor (PLGF) Levels in Plasma
2.6. Determination of H2S in Plasma
2.7. Determination of NO Metabolites in Plasma
2.8. Determinants of Oxidative Stress in Plasma
2.9. Determination of H2S Levels in PVAT, The Aorta without PVAT, and the Placenta
2.10. Statistical Analysis
2.11. Drugs and Chemicals
3. Results
3.1. Systolic Blood Pressure Measurements in Non-Pregnant and Pregnant Rats Submitted (or Not) to the DOCA-Salt Protocol
3.2. Placental and Fetal Parameters of Pregnant Rats Submitted (or Not) to the DOCA-Salt Protocol
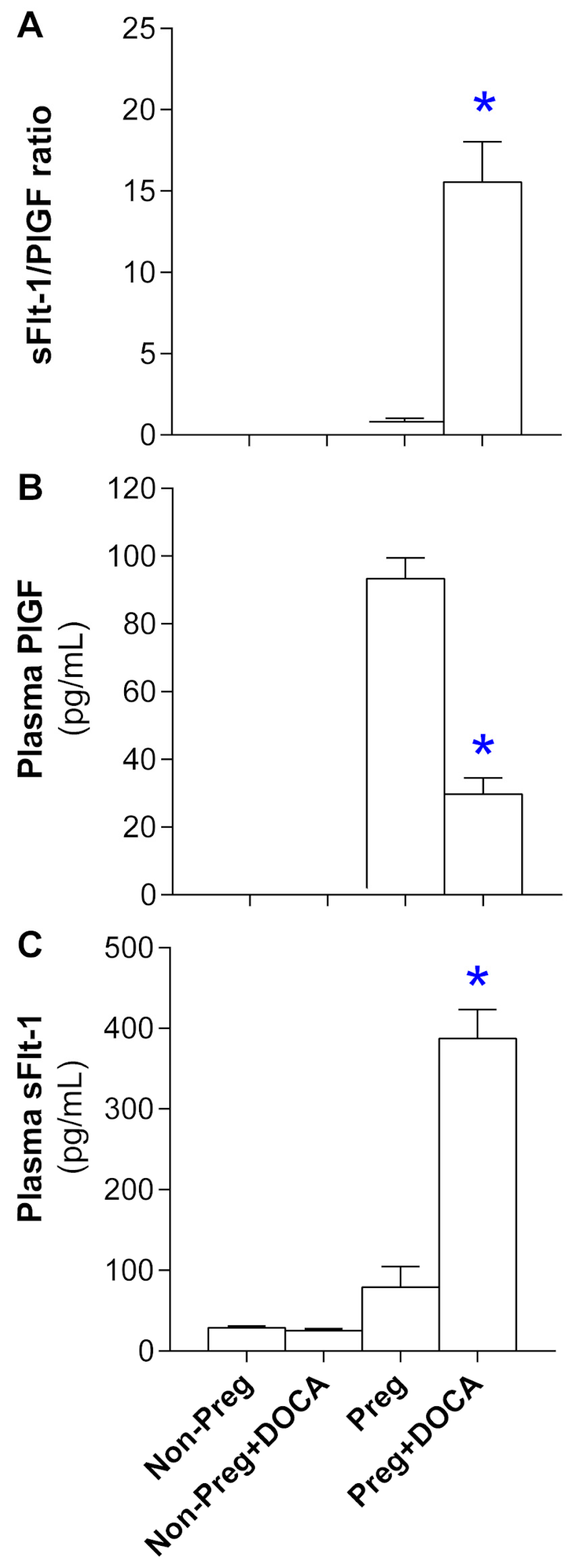
3.3. Angiogenesis Biomarker Levels in Plasma from Non-Pregnant and Pregnant Rats Submitted (or Not) to the DOCA-Salt Protocol
3.4. Plasma Level Determinants of H2S, NO, and Oxidative Stress in Non-Pregnant and Pregnant Rats Submitted (or Not) to the DOCA-Salt Protocol
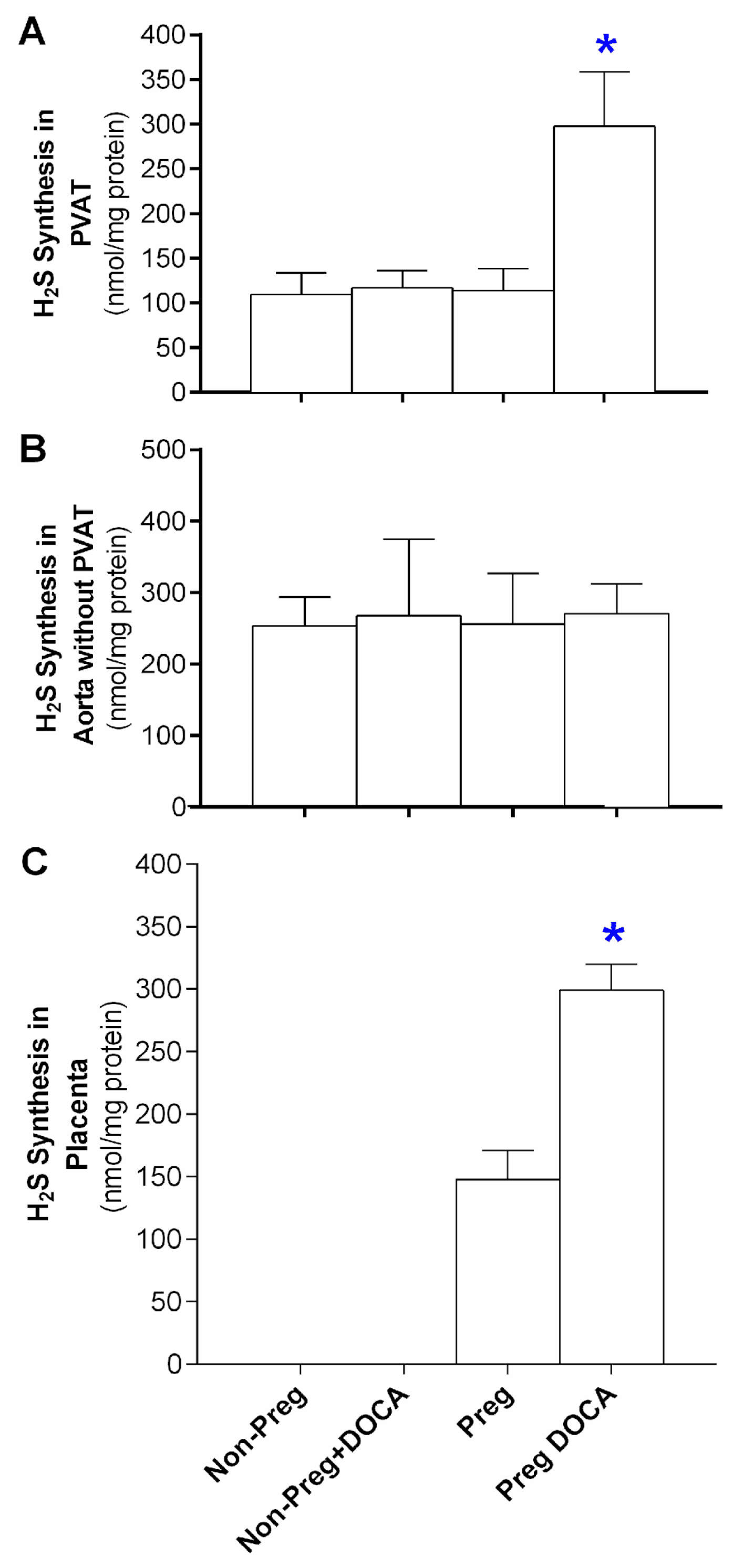
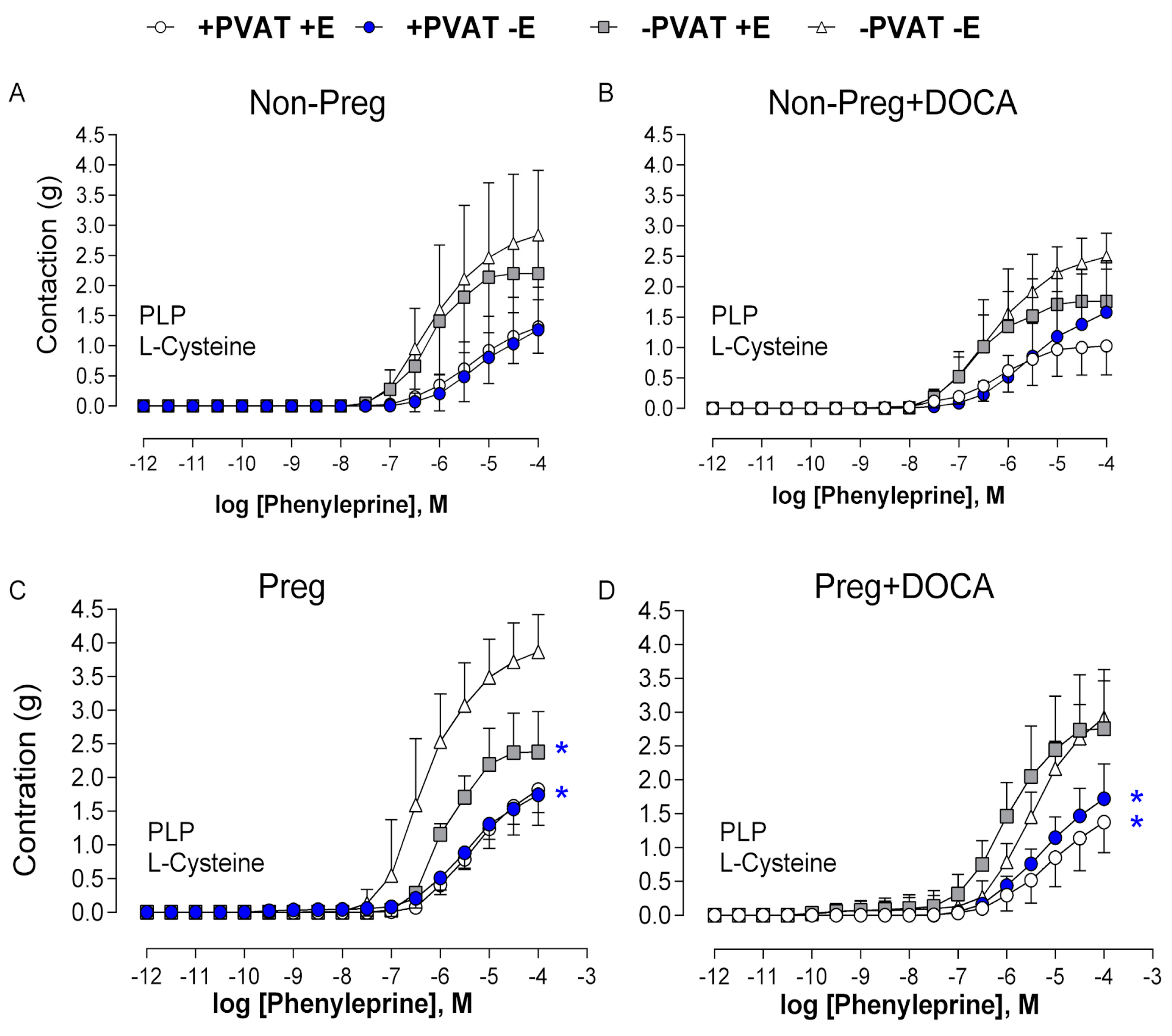
3.5. Stimulated H2S Synthesis in the PVAT, the Aorta without PVAT, and the Placenta
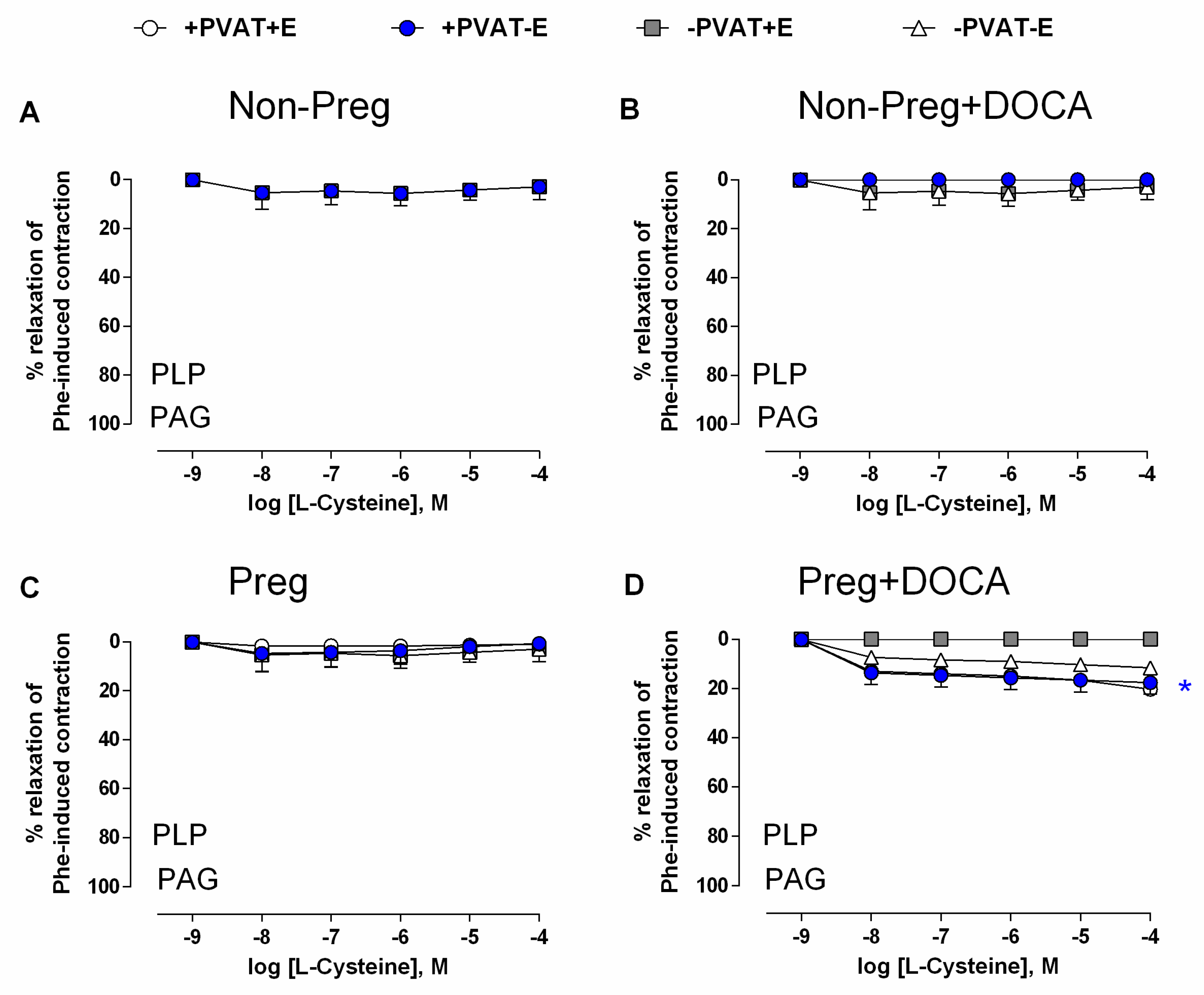
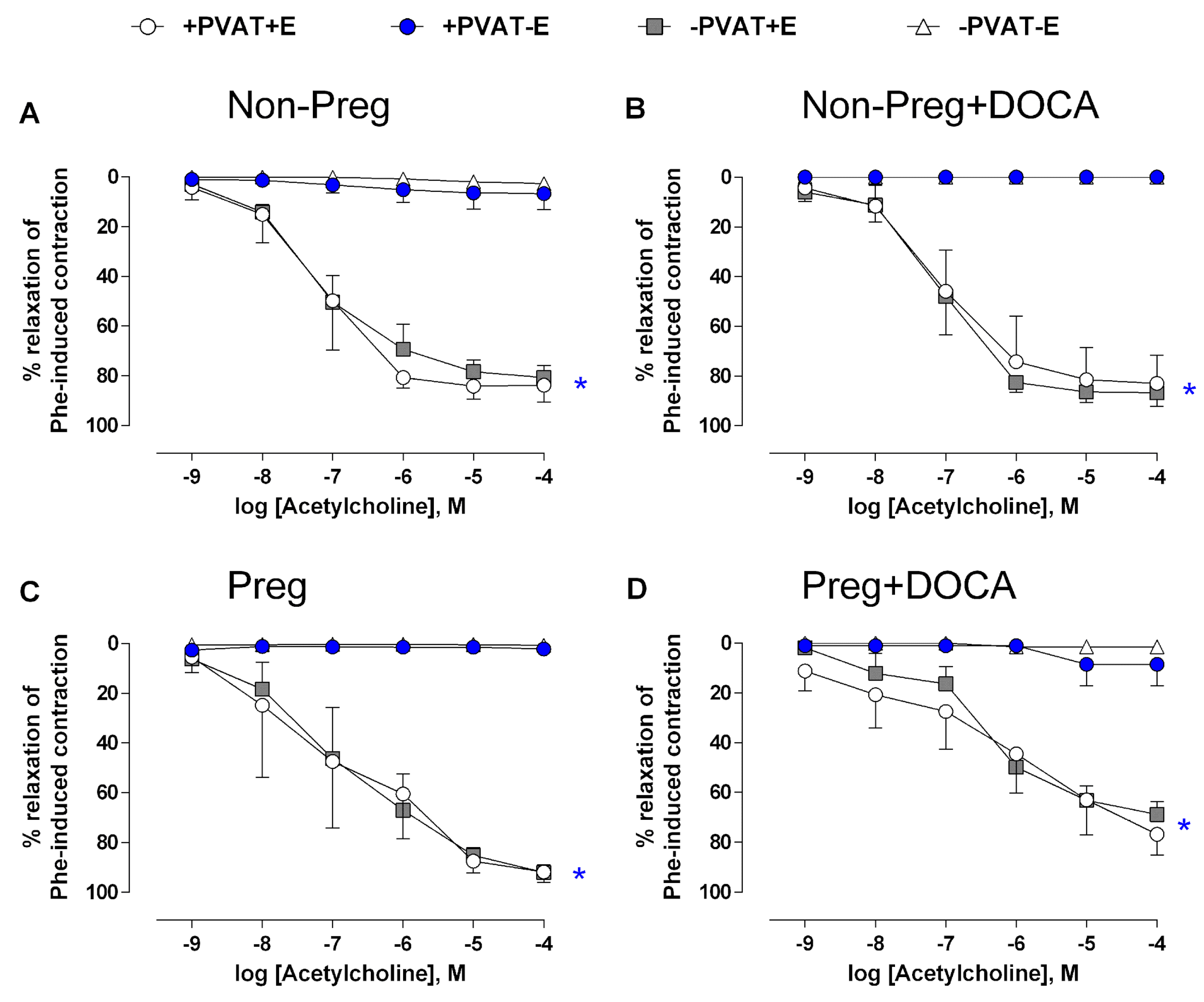
3.6. Vasodilation Responses via Endogenous H2S Stimulation in Aortas with Intact (or Removed) PVAT and/or Endothelium from Non-Pregnant and Pregnant Rats Submitted (or Not) to the DOCA-Salt Protocol
3.7. Vascular Responses to L-Cysteine in the Presence of H2S Synthesis Inhibitor in Aortas with Intact (or Removed) PVAT and/or Endothelium from Non-Pregnant and Pregnant Rats Submitted (or Not) to the DOCA-Salt Protocol
3.8. Vasodilation Responses via Endogenous NO Stimulation in Aortas with Intact (or Removed) PVAT and/or Endothelium from Non-Pregnant and Pregnant Rats Submitted (or Not) to the DOCA-Salt Protocol
3.9. Vascular Responses to Acetylcholine in The Presence of NO Synthesis Inhibitor in Aortas with Intact (or Removed) PVAT and/or Endothelium from Non-Pregnant and Pregnant Rats Submitted (or Not) to the DOCA-Salt Protocol
3.10. Anticontractile Responses Induced by PVAT and Endothelium in Aortas from Non-Pregnant and Pregnant Rats Submitted (or Not) to the DOCA-Salt Protocol
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mann, S.E.; Maille, N.; Clas, D.; Osol, G. Perivascular adipose tissue: A novel regulator of vascular tone in the rat pregnancy. Reprod. Sci. 2015, 22, 802–807. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Lu, C.; Su, L.; Sharma, A.M.; Lee, R.M.K.W. Modulation of vascular function by perivascular adipose tissue: The role of endothelium and hydrogen peroxide. Br. J. Pharmacol. 2007, 151, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Saxton, S.N.; Clark, B.J.; Withers, S.B.; Eringa, E.C.; Heagerty, A.M. Mechanistic Links Between Obesity, Diabetes, and Blood Pressure: Role of Perivascular Adipose Tissue. Physiol. Rev. 2019, 99, 1701–1763. [Google Scholar] [CrossRef] [PubMed]
- Löhn, M.; Dubrovska, G.; Lauterbach, B.; Luft, F.C.; Gollasch, M.; Sharma, A.M. Periadventitial fat releases a vascular relaxing factor. FASEB J. 2002, 16, 1057–1063. [Google Scholar] [CrossRef]
- Osikoya, O.; Ahmed, H.; Panahi, S.; Bourque, S.L.; Goulopoulou, S. Uterine perivascular adipose tissue is a novel mediator of uterine artery blood flow and reactivity in rat pregnancy. J. Physiol. 2019, 597, 3833–3852. [Google Scholar] [CrossRef] [PubMed]
- Zeigler, M.B.; Fay, E.E.; Moreni, S.L.; Mao, J.; Totah, R.A.; Hebert, M.F. Plasma hydrogen sulfide, nitric oxide, and thiocyanate levels are lower during pregnancy compared to postpartum in a cohort of women from the Pacific northwest of the United States. Life Sci. 2023, 322, 121625. [Google Scholar] [CrossRef]
- Boeldt, D.S.; Bird, I.M. Vascular adaptation in pregnancy and endothelial dysfunction in preeclampsia. J. Endocrinol. 2017, 232, R27–R44. [Google Scholar] [CrossRef]
- Baltieri, N.; Guizoni, D.M.; Victorio, J.A.; Davel, A.P. Protective Role of Perivascular Adipose Tissue in Endothelial Dysfunction and Insulin-Induced Vasodilatation of Hypercholesterolemic LDL Receptor-Deficient Mice. Front. Physiol. 2018, 9, 229. [Google Scholar] [CrossRef]
- Beltowski, J.; Jamroz-Wisniewska, A. Hydrogen sulfide in the adipose tissue-physiology, pathology and a target for pharmacotherapy. Molecules 2016, 22, 63. [Google Scholar] [CrossRef]
- Polhemus, D.J.; Calvert, J.W.; Butler, J.; Lefer, D.J. The cardioprotective actions of hydrogen sulfide in acute myocardial infarction and heart failure. Scientifica 2014, 2014, 768607. [Google Scholar] [CrossRef]
- Wang, Y.-Z.; Ngowi, E.E.; Wang, D.; Qi, H.-W.; Jing, M.-R.; Zhang, Y.-X.; Cai, C.-B.; He, Q.-L.; Khattak, S.; Khan, N.H.; et al. The Potential of Hydrogen Sulfide Donors in Treating Cardiovascular Diseases. Int. J. Mol. Sci. 2021, 22, 2194. [Google Scholar] [CrossRef]
- Golas, S.; Berenyiova, A.; Majzunova, M.; Drobna, M.; Tuorkey, M.J.; Cacanyiova, S. The Vasoactive Effect of Perivascular Adipose Tissue and Hydrogen Sulfide in Thoracic Aortas of Normotensive and Spontaneously Hypertensive Rats. Biomolecules 2022, 12, 457. [Google Scholar] [CrossRef]
- Zhao, W.; Zhang, J.; Lu, Y.; Wang, R. The vasorelaxant effect of H2S as a novel endogenous gaseous KATP channel opener. EMBO J. 2001, 20, 6008–6016. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.K.; Sikka, G.; Gazi, S.K.; Steppan, J.; Jung, S.M.; Bhunia, A.K.; Barodka, V.M.; Gazi, F.K.; Barrow, R.K.; Wang, R.; et al. Hydrogen sulfide as endothelium-derived hyperpolarizing factor sulfhydrates potassium channels. Circ. Res. 2011, 109, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Zhao, J.; Chen, Y.; Ma, T.; Xu, G.; Tang, C.; Liu, X.; Geng, B. Hydrogen sulfide derived from periadventitial adipose tissue is a vasodilator. J. Hypertens. 2009, 27, 2174–2185. [Google Scholar] [CrossRef] [PubMed]
- Köhn, C.; Schleifenbaum, J.; Szijártó, I.A.; Markó, L.; Dubrovska, G.; Huang, Y.; Gollasch, M. Differential effects of cystathionine-g-lyase–dependent vasodilatory H2S in periadventitial vasoregulation of rat and mouse aortas. PLoS ONE 2012, 7, e41951. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.; Yang, G.; Jiang, B.; Ju, Y.; Wu, L.; Wang, R. H₂S is an endothelium-derived hyperpolarizing factor. Antioxid. Redox Signal. 2013, 19, 1634–1646. [Google Scholar] [CrossRef] [PubMed]
- Kanagy, N.L.; Szabo, C.; Papapetropoulos, A. Vascular biology of hydrogen sulfide. Am. J. Physiol. Physiol. 2017, 312, C537–C549. [Google Scholar] [CrossRef]
- Cirino, G.; Szabo, C.; Papapetropoulos, A. Physiological roles of hydrogen sulfide in mammalian cells, tissues, and organs. Physiol. Rev. 2023, 103, 31–276. [Google Scholar] [CrossRef]
- Kabil, O.; Vitvitsky, V.; Xie, P.; Banerjee, R. The quantitative significance of the transsulfuration enzymes for H2S production in murine tissues. Antioxid. Redox Signal. 2011, 15, 363–372. [Google Scholar] [CrossRef]
- Powell, C.R.; Dillon, K.M.; Matson, J.B. A review of hydrogen sulfide (H2S) donors: Chemistry and potential therapeutic applications. Biochem. Pharmacol. 2018, 149, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Asimakopoulou, A.; Panopoulos, P.; Chasapis, C.T.; Coletta, C.; Zhou, Z.; Cirino, G.; Giannis, A.; Szabo, C.; Spyroulias, G.A.; Papapetropoulos, A. Selectivity of commonly used pharmacological inhibitors for cystathionine b synthase (CBS) and cystathionine g lyase (CSE). Br. J. Pharmacol. 2013, 169, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Chen, Y.; Zhao, J.; Tang, C.; Jiang, Z.; Geng, B. Hydrogen sulfide from adipose tissue is a novel insulin resistance regulator. Biochem. Biophys. Res. Commun. 2009, 380, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Gheibi, S.; Jeddi, S.; Kashfi, K.; Ghasemi, A. Regulation of vascular tone homeostasis by NO and H2S: Implications in hypertension. Biochem. Pharmacol. 2018, 149, 42–59. [Google Scholar] [CrossRef]
- Selovic, A.; Sarac, J.; Missoni, S. Changes in adipose tissue distribution during pregnancy estimated by ultrasonography. J. Matern. Neonatal Med. 2016, 29, 2131–2137. [Google Scholar] [CrossRef]
- Trivett, C.; Lees, Z.J.; Freeman, D.J. Adipose tissue function in healthy pregnancy, gestational diabetes mellitus and pre-eclampsia. Eur. J. Clin. Nutr. 2021, 75, 1745–1756. [Google Scholar] [CrossRef]
- Souza-Paula, E.P.D.; Polonio, L.C.P.D.M.; Zochio, G.P.P.; da Silva, K.P.; Kushima, H.; Dias-Junior, C.A.P.D. Anticontractile Effect of Perivascular Adipose Tissue but Not of Endothelium Is Enhanced by Hydrogen Sulfide Stimulation in Hypertensive Pregnant Rat Aortae. J. Cardiovasc. Pharmacol. 2020, 76, 715–729. [Google Scholar] [CrossRef]
- Ianosi-Irimie, M.; Vu, H.V.; Whitbred, J.M.; Pridjian, C.A.; Nadig, J.D.; Williams, M.Y.; Wrenn, D.C.; Pridjian, G.; Puschett, J.B. A rat model of preeclampsia. Clin. Exp. Hypertens. 2005, 27, 605–617. [Google Scholar] [CrossRef]
- Mitchell, B.M.; Cook, L.G.; Danchuk, S.; Puschett, J.B. Uncoupled endothelial nitric oxide synthase and oxidative stress in a rat model of pregnancy-induced hypertension. Am. J. Hypertens. 2007, 20, 1297–1304. [Google Scholar] [CrossRef]
- Gonzaga, N.A.; Awata, W.M.; Vale, G.T.D.; Marchi, K.C.; Muniz, J.J.; Tanus-Santos, J.E.; Tirapelli, C.R. Perivascular adipose tissue protects against the vascular dysfunction induced by acute ethanol intake: Role of hydrogen peroxide. Vasc. Pharmacol. 2018, 111, 44–53. [Google Scholar] [CrossRef]
- Tirapelli, C.R.; Al-Khoury, J.; Bkaily, G.; D’Orléans-Juste, P.; Lanchote, V.L.; Uyemura, S.A.; de Oliveira, A.M. Chronic ethanol consumption enhances phenylephrine-induced contraction in the isolated rat aorta. Experiment 2006, 316, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Ahmad, S.; Cai, M.; Rennie, J.; Fujisawa, T.; Crispi, F.; Baily, J.; Miller, M.R.; Cudmore, M.; Hadoke, P.W.; et al. Dysregulation of hydrogen sulfide producing enzyme cystathionine γ-lyase contributes to maternal hypertension and placental abnormalities in preeclampsia. Circulation 2013, 127, 2514–2522. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Lei, F.; Zhang, S.; Yan, X.; Wang, W.; Zheng, Y. Hydrogen sulfide alleviates placental injury induced by maternal cigarette smoke exposure during pregnancy in rats. Nitric Oxide 2018, 74, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Toghi, C.J.; Martins, L.Z.; Pacheco, L.L.; Caetano, E.S.P.; Mattos, B.R.; Rizzi, E.; Dias-Junior, C.A. Pravastatin Prevents Increases in Activity of Metalloproteinase-2 and Oxidative Stress, and Enhances Endothelium-Derived Nitric Oxide-Dependent Vasodilation in Gestational Hypertension. Antioxidants 2023, 12, 939. [Google Scholar] [CrossRef]
- Chimini, J.S.; Possomato-Vieira, J.S.; da Silva, M.L.S.; Dias-Junior, C.A. Placental nitric oxide formation and endothelium-dependent vasodilation underlie pravastatin effects against angiogenic imbalance, hypertension in pregnancy and intrauterine growth restriction. Basic Clin. Pharmacol. Toxicol. 2019, 124, 385–393. [Google Scholar] [CrossRef]
- Rodrigues, L.; Ekundi-Valentim, E.; Florenzano, J.; Cerqueira, A.; Soares, A.; Schmidt, T.; Santos, K.; Teixeira, S.; Ribela, M.; Rodrigues, S.; et al. Protective effects of exogenous and endogenous hydrogen sulfide in mast cell-mediated pruritus and cutaneous acute inflammation in mice. Pharmacol. Res. 2017, 115, 255–266. [Google Scholar] [CrossRef]
- Uren, J.R.; Ragin, R.; Chaykovsky, M. Modulation of Cysteine Metabolism in Mice–Effects of Propargylglycine and L-Cyst(e)ine-degrading Enzymes. Biochem. Pharmacol. 1978, 27, 2807–2814. [Google Scholar] [CrossRef]
- Perusquía, M.; Hanson, A.E.; Meza, C.M.; Kubli, C.; Herrera, N.; Stallone, J.N. Antihypertensive responses of vasoactive androgens in an in vivo experimental model of preeclampsia. J. Steroid Biochem. Mol. Biol. 2018, 178, 65–72. [Google Scholar] [CrossRef]
- Al-Jarallah, A.; Oriowo, M.A. Loss of Anticontractile Effect of Perivascular Adipose Tissue on Pregnant Rats: A Potential Role of Tumor Necrosis Factor-α. J. Cardiovasc. Pharmacol. 2016, 67, 145–151. [Google Scholar] [CrossRef]
- Osikoya, O.; Cushen, S.C.; Ricci, C.A.; Goulopoulou, S. Cyclooxygenase-dependent mechanisms mediate in part the anti-dilatory effects of perivascular adipose tissue in uterine arteries from pregnant rats. Pharmacol. Res. 2021, 171, 105788. [Google Scholar] [CrossRef]
- Watts, S.W.; Flood, E.D.; Garver, H.; Fink, G.D.; Roccabianca, S. A New Function for Perivascular Adipose Tissue (PVAT): Assistance of Arterial Stress Relaxation. Sci. Rep. 2020, 10, 1807. [Google Scholar] [CrossRef]
- Donovan, J.; Wong, P.S.; Garle, M.J.; Alexander, S.P.H.; Dunn, W.R.; Ralevic, V. Coronary artery hypoxic vasorelaxation is augmented by perivascular adipose tissue through a mechanism involving hydrogen sulphide and cystathionine-β-synthase. Acta Physiol. 2018, 224, e13126. [Google Scholar] [CrossRef]
- Hart, J.L. Vasorelaxation elicited by endogenous and exogenous hydrogen sulfide in mouse mesenteric arteries. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.G.; O’Malley, E.J.; Ho, W.S.V. Pro-contractile effects of perivascular fat in health and disease. Br. J. Pharmacol. 2017, 174, 3482–3495. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.-T.; Chang, C.-K.; Tsao, C.-W.; Hsu, Y.-M.; Hsu, C.-T.; Cheng, J.-T. Activation of muscarinic M-3 receptor may decrease glucose uptake and lipolysis in adipose tissue of rats. Neurosci. Lett. 2009, 451, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Kameshima, S.; Yamada, K.; Morita, T.; Okada, M.; Yamawaki, H. Visceral adipose tissue-derived serine protease inhibitor augments acetylcholine-induced relaxation via the inhibition of acetylcholine esterase activity in rat isolated mesenteric artery. Acta Physiol. 2016, 216, 203–210. [Google Scholar] [CrossRef]
- Possomato-Vieira, J.S.; Khalil, R.A. Mechanisms of Endothelial Dysfunction in Hypertensive Pregnancy and Preeclampsia. Adv. Pharmacol. 2016, 77, 361–431. [Google Scholar]
- Xia, N.; Horke, S.; Habermeier, A.; Closs, E.I.; Reifenberg, G.; Gericke, A.; Mikhed, Y.; Münzel, T.; Daiber, A.; Förstermann, U.; et al. Uncoupling of Endothelial Nitric Oxide Synthase in Perivascular Adipose Tissue of Diet-Induced Obese Mice. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 78–85. [Google Scholar] [CrossRef]
- Gil-Ortega, M.; Condezo-Hoyos, L.; García-Prieto, C.F.; Arribas, S.M.; González, M.C.; Aranguez, I.; Ruiz-Gayo, M.; Somoza, B.; Fernández-Alfonso, M.S. Imbalance between pro and anti-oxidant mechanisms in perivascular adipose tissue aggravates long-term high-fat diet-derived endothelial dysfunction. PLoS ONE 2014, 9, e95312. [Google Scholar] [CrossRef]
- Bartha, J.L.; Comino-Delgado, R.; Bedoya, F.J.; Barahona, M.; Lubian, D.; Garcia-Benasach, F. Maternal serum nitric oxide levels associated with biochemical and clinical parameters in hypertension in pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 1999, 82, 201–207. [Google Scholar] [CrossRef]
- Possomato-Vieira, J.S.; Palei, A.C.; Pinto-Souza, C.C.; Cavalli, R.; Dias-Junior, C.A.; Sandrim, V. Circulating levels of hydrogen sulphide negatively correlate to nitrite levels in gestational hypertensive and preeclamptic pregnant women. Clin. Exp. Pharmacol. Physiol. 2021, 48, 1224–1230. [Google Scholar] [CrossRef] [PubMed]
- Socha, M.W.; Stankiewicz, M.; Żołnieżewicz, K.; Puk, O.; Wartęga, M. Decrease in Nitric Oxide Production as a Key Mediator in the Pathogenesis of Preeclampsia and a Potential Therapeutic Target: A Case-Control Study. Biomedicines 2022, 10, 2653. [Google Scholar] [CrossRef] [PubMed]
- Tashie, W.; Fondjo, L.A.; Owiredu, W.K.B.A.; Ephraim, R.K.D.; Asare, L.; Adu-Gyamfi, E.A.; Seidu, L. Altered Bioavailability of Nitric Oxide and L-Arginine Is a Key Determinant of Endothelial Dysfunction in Preeclampsia. BioMed Res. Int. 2020, 2020, 3251956. [Google Scholar] [CrossRef] [PubMed]
- Sandrim, V.C.; Palei, A.C.; Metzger, I.F.; Gomes, V.A.; Cavalli, R.C.; Tanus-Santos, J.E. Nitric oxide formation is inversely related to serum levels of antiangiogenic factors soluble FMS-like tyrosine kinase-1 and soluble endogline in preeclampsia. Hypertension 2008, 52, 402–407. [Google Scholar] [CrossRef]
- Palei, A.C.; Granger, J.P.; Spradley, F.T. Placental Ischemia Says “NO” to Proper NOS-Mediated Control of Vascular Tone and Blood Pressure in Preeclampsia. Int. J. Mol. Sci. 2021, 22, 11261. [Google Scholar] [CrossRef]
- Goulopoulou, S.; Davidge, S.T. Molecular mechanisms of maternal vascular dysfunction in preeclampsia. Trends Mol. Med. 2015, 21, 88–97. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E. Oxidative stress. Best Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 287–299. [Google Scholar] [CrossRef]
- Opichka, M.A.; Rappelt, M.W.; Gutterman, D.D.; Grobe, J.L.; McIntosh, J.J. Vascular Dysfunction in Preeclampsia. Cells 2021, 10, 3055. [Google Scholar] [CrossRef]
- Redman, C.; Sargent, I. Placental stress and pre-eclampsia: A revised view. Placenta 2009, 30, 38–42. [Google Scholar] [CrossRef]
- Al-Sheikh, Y.A.; Al-Zahrani, K.Y. The Status of Biochemical and Molecular Markers of Oxidative Stress in Preeclamptic Saudi Patients. Curr. Mol. Med. 2018, 18, 475–485. [Google Scholar] [CrossRef]
- Serdar, Z.; Gür, E.; Develioğlu, O. Serum iron and copper status and oxidative stress in severe and mild preeclampsia. Cell. Biochem. Funct. 2006, 24, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K.; Matsumura, Y.; Miyazaki, Y.; Takaoka, M.; Morimoto, S. Effects of the endothelin ETA-receptor antagonist FR139317 on development of hypertension and cardiovascular hypertrophy in deoxycorticosterone acetate-salt hypertensive rats. Jpn. J. Pharmacol. 1996, 70, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Agafonova, I.G.; Kotelnikov, V.N.; Geltser, B.I. Structural and Functional Changes of the Thoracic Aorta in Experimental Arterial Hypertension. Bull. Exp. Biol. Med. 2023, 174, 308–311. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Kita, S.; Morimoto, S.; Akimoto, K.; Furuya, M.; Oka, N.; Tanaka, T. Antihypertensive effect of sesamin. I. Protection against deoxycorticosterone acetate-salt-induced hypertension and cardiovascular hypertrophy. Biol. Pharm. Bull. 1995, 18, 1016–1019. [Google Scholar] [CrossRef]
- Piecha, G.; Koleganova, N.; Ritz, E.; Müller, A.; Fedorova, O.V.; Bagrov, A.Y.; Lutz, D.; Schirmacher, P.; Gross-Weissmann, M.-L. High salt intake causes adverse fetal programming—Vascular effects beyond blood pressure. Nephrol. Dial. Transplant. 2012, 27, 3464–3476. [Google Scholar] [CrossRef]
- Uddin, M.N.; McLean, L.B.; Hunter, F.A.; Horvat, D.; Severson, J.; Tharakan, B.; Childs, E.W.; Puschett, J.B. Vascular Leak in a Rat Model of Preeclampsia. Am. J. Nephrol. 2009, 30, 26–33. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, P.B.d.; Zochio, G.P.; Caetano, E.S.P.; da Silva, M.L.S.; Dias-Junior, C.A. Vasodilator Responses of Perivascular Adipose Tissue-Derived Hydrogen Sulfide Stimulated with L-Cysteine in Pregnancy Hypertension-Induced Endothelial Dysfunction in Rats. Antioxidants 2023, 12, 1919. https://doi.org/10.3390/antiox12111919
Oliveira PBd, Zochio GP, Caetano ESP, da Silva MLS, Dias-Junior CA. Vasodilator Responses of Perivascular Adipose Tissue-Derived Hydrogen Sulfide Stimulated with L-Cysteine in Pregnancy Hypertension-Induced Endothelial Dysfunction in Rats. Antioxidants. 2023; 12(11):1919. https://doi.org/10.3390/antiox12111919
Chicago/Turabian StyleOliveira, Priscilla Bianca de, Gabriela Palma Zochio, Edileia Souza Paula Caetano, Maria Luiza Santos da Silva, and Carlos Alan Dias-Junior. 2023. "Vasodilator Responses of Perivascular Adipose Tissue-Derived Hydrogen Sulfide Stimulated with L-Cysteine in Pregnancy Hypertension-Induced Endothelial Dysfunction in Rats" Antioxidants 12, no. 11: 1919. https://doi.org/10.3390/antiox12111919