
Melatonin-Mediated Suppression of mtROS-JNK-FOXO1 Pathway Alleviates Hypoxia-Induced Apoptosis in Porcine Granulosa Cells

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Sample Collection and Cell Culture
2.3. Experimental Design and Cell Treatment
2.4. Mitochondrial ROS Detection
2.5. Western Blotting
2.6. Immunofluorescence
2.7. TUNEL Assay
2.8. Quantitative Real-Time PCR (qRT-PCR)
2.9. RNA Interference and Gene Overexpression
- Scrambled siRNA:
- Sense (5′-UUCUCCGAACGUGUCACGUTT-3′)
- Antisense (5′-ACGUGACACGUUCGGAGAATT-3′)
- siFOXO1:
- Sense (5′-GCAUGUUCAUUGAGCGCUUTT-3′)
- Antisense (5′-AAGCGCUCAAUGAACAUGCTT-3′)
- siSOD2:
- Sense (5′-GGCCACAUCAAUCAUAGCATT-3′)
- Antisense (5′-UGCUAUGAUUGAUGUGGCC-3′)
2.10. Statistics Analysis
3. Results
3.1. Hypoxia Promotes GC Apoptosis by Increasing the Accumulation of mtROS
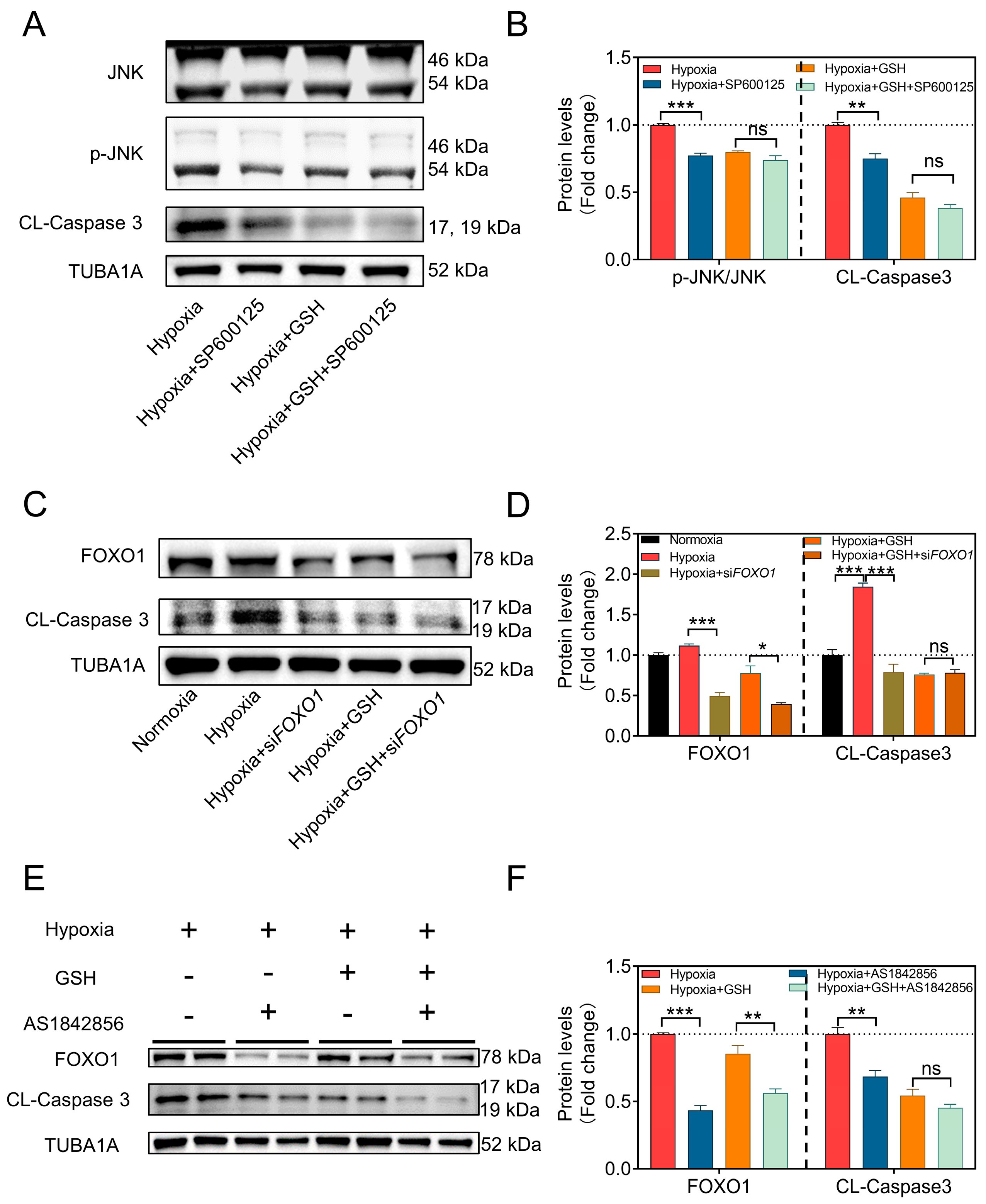
3.2. Hypoxia Induces GC Apoptosis by Activating the mtROS-JNK-FOXO1 Pathway
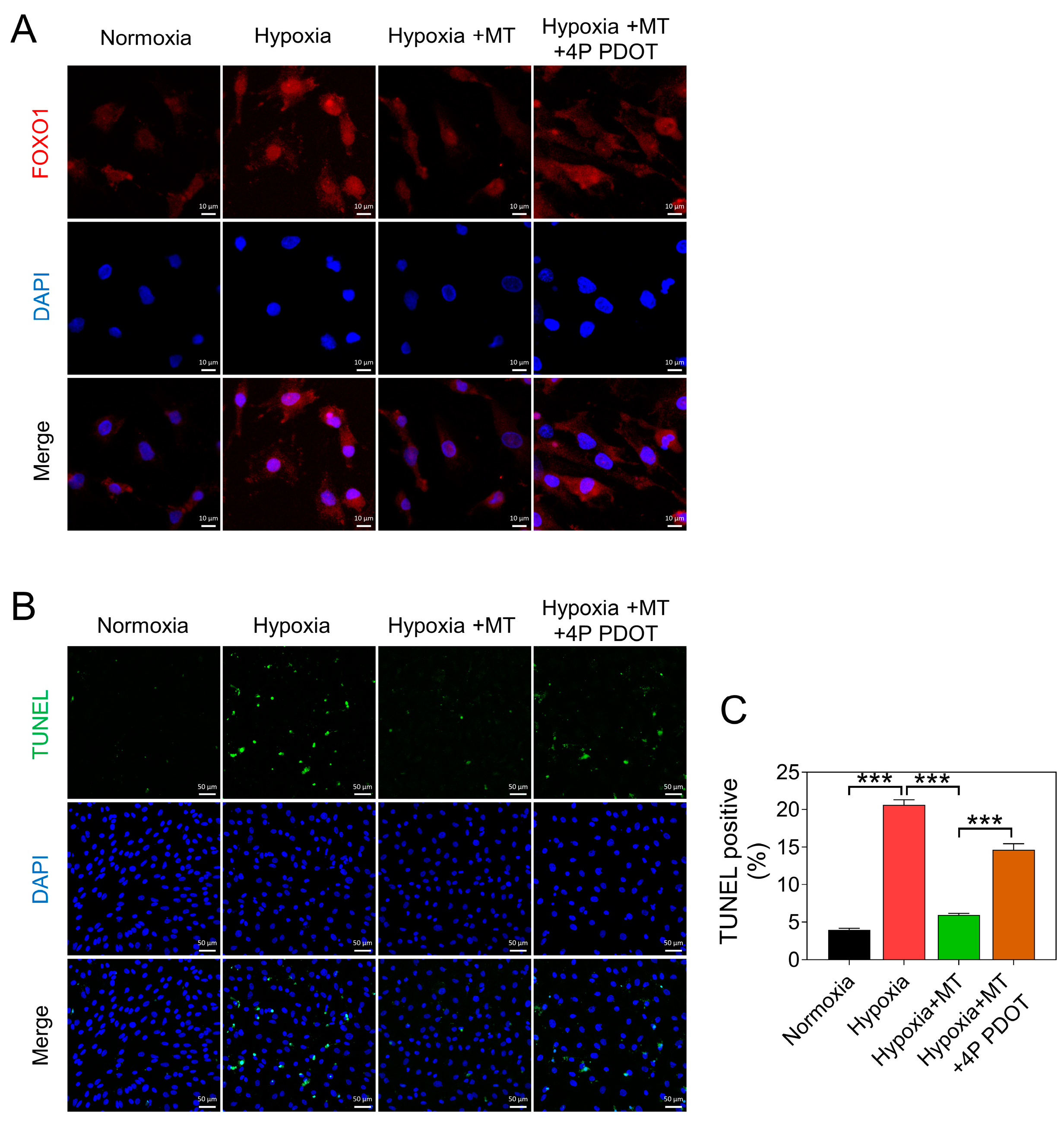
3.3. Melatonin Alleviates Hypoxia-Induced Apoptosis of GCs by Reducing mtROS Levels
3.4. Melatonin Inhibits Hypoxia-Induced mtROS Generation and GC Apoptosis through the MTNR1B Receptor
3.5. SOD2 Contributes to the Inhibition of Hypoxia-Induced mtROS Accumulation and GC Apoptosis by Melatonin
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, X.; Chen, Y.; Li, H.; Chen, B.; Liu, Z.; Wu, G.; Li, C.; Li, R.; Cao, Y.; Zhou, J. Sulforaphane Acts Through NFE2L2 to Prevent Hypoxia-Induced Apoptosis in Porcine Granulosa Cells via Activating Antioxidant Defenses and Mitophagy. J. Agric. Food Chem. 2022, 70, 8097–8110. [Google Scholar] [CrossRef]
- Li, L.; Huang, N.; Qi, Y.; Li, Y.; Wang, L. Effect of Wenshentiaojing Decoction on Hormone Level and Follicular Number in Patients with Menstrual Disorder of Polycystic Ovary Syndrome; Hindawi Ltd.: London, UK, 2021. [Google Scholar]
- Sun, Z.; Zhang, H.; Wang, X.; Wang, Q.C.; Zhang, C.; Wang, J.Q.; Wang, Y.-H.; An, C.-Q.; Yang, K.-Y.; Wang, Y.; et al. TMCO1 is essential for ovarian follicle development by regulating ER Ca 2+ store of granulosa cells. Cell Death Differ. 2018, 25, 1686–1701. [Google Scholar] [CrossRef] [PubMed]
- Harder, J.D.; Jackson, L.M. Male pheromone stimulates ovarian follicular development and body growth in juvenile female opossums (Monodelphis domestica). Reprod. Biol. Endocrinol. 2003, 1, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Sabharwal, S.S.; Schumacker, P.T. Mitochondrial ROS in cancer: Initiators, amplifiers or an Achilles’ heel? Nat. Rev. Cancer 2014, 14, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Belousov, V.V.; Chandel, N.S.; Davies, M.J.; Winterbourn, C. Defining roles of specific reactive oxygen species (ROS) in cell biology and physiology. Nat. Rev. Mol. Cell Biol. 2022, 23, 499–515. [Google Scholar] [CrossRef]
- Matzuk, M.M.; Dionne, L.; Guo, Q.; Kumar, T.R.; Lebovitz, R.M. Ovarian function in superoxide dismutase 1 and 2 knockout mice. Endocrinology 1998, 139, 4008–4011. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Camarillo, C.; González-González, A.; Vergara-Onofre, M.; González-Padilla, E.; Avalos-Rodriguez, A.; Gutierrez-Rodriguez, M.E.; Arriaga-Pizano, L.; Cruz, M.; Baiza-Gutman, L.A.; Diaz-Flores, M. Changes in the glucose-6-phosphate dehydrogenase activity in granulosa cells during follicular atresia in ewes. Reproduction 2009, 137, 979–986. [Google Scholar] [CrossRef]
- Tilly, J.L.; Tilly, K.I.; Kenton, M.L.; Johnson, A.L. Expression of members of the bcl-2 gene family in the immature rat ovary: Equine chorionic gonadotropin-mediated inhibition of granulosa cell apoptosis is associated with decreased bax and constitutive bcl-2 and bcl-xlong messenger ribonucleic acid levels. Endocrinology 1995, 136, 232–241. [Google Scholar] [CrossRef]
- Gupta, R.K.; Miller, K.P.; Babus, J.K.; Flaws, J.A. Methoxychlor inhibits growth and induces atresia of antral follicles through an oxidative stress pathway. Toxicol. Sci. Off. J. Soc. Toxicol. 2006, 93, 382–389. [Google Scholar] [CrossRef]
- Venegas, C.; García, J.A.; Escames, G.; Ortiz, F.; López, A.; Doerrier, C.; García-Corzo, L.; López, L.C.; Reiter, R.J.; Acuña-Castroviejo, D. Extrapineal melatonin: Analysis of its subcellular distribution and daily fluctuations. J. Pineal Res. 2012, 52, 217–227. [Google Scholar] [CrossRef]
- Díaz-Casado, M.E.; López, L.C.; Acu?a-Castroviejo, D.; Venegas, C.; Tan, D.X.; Lima-Cabello, E.; Escames, G.; Rosales-Corral, S.; Reiter, R.J. Extrapineal melatonin: Sources, regulation, and potential functions. Cell. Mol. Life Sci. CMLS 2014, 71, 2997–3025. [Google Scholar]
- Sakaguchi, K.; Itoh, M.T.; Takahashi, N.; Tarumi, W.; Ishizuka, B. The rat oocyte synthesises melatonin. Reprod. Fertil. Dev. 2013, 25, 674. [Google Scholar] [CrossRef]
- El-Raey, M.; Geshi, M.; Somfai, T.; Kaneda, M.; Hirako, M.; Abdel-Ghaffar, A.E.; Sosa, G.A.; El-Roos, M.E.A.A.; Nagai, T. Evidence of melatonin synthesis in the cumulus oocyte complexes and its role in enhancing oocyte maturation in vitro in cattle. Mol. Reprod. Dev. 2011, 78, 250–262. [Google Scholar] [CrossRef]
- Amireault, P. Serotonin and Its Antidepressant-Sensitive Transport in Mouse Cumulus-Oocyte Complexes and Early Embryos. Biol. Reprod. 2005, 73, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Cheng, K.; Zhang, B.; Xu, H.; Cao, Y.; Guo, F.; Feng, X.; Xia, Q. Novel mechanisms for superoxide-scavenging activity of human manganese superoxide dismutase determined by the K68 key acetylation site. Free Radic. Biol. Med. Off. J. Oxyg. Soc. 2015, 85, 114–126. [Google Scholar] [CrossRef]
- Pi, H.; Xu, S.; Reiter, R.J.; Guo, P.; Zhang, L.; Li, Y.; Li, M.; Cao, Z.; Tian, L.; Xie, J. SIRT3-SOD2-mROS-dependent autophagy in cadmium-induced hepatotoxicity and salvage by melatonin. Autophagy 2015, 11, 1037–1051. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.Y.; Sun, X.F.; Li, L.; Ma, J.M.; Zhang, R.Q.; Li, N.; Liu, X.-L.; Dyce, P.W.; Shen, W. Ochratoxin A Exposure Impairs Porcine Granulosa Cell Growth via the PI3K/AKT Signaling Pathway. J Agric Food Chem. 2019, 67, 2679–2690. [Google Scholar] [CrossRef]
- Liu, Z.; Li, C.; Wu, G.; Li, W.; Zhang, X.; Zhou, J.; Zhang, L.; Tao, J.; Shen, M.; Liu, H. Involvement of JNK/FOXO1 pathway in apoptosis induced by severe hypoxia in porcine granulosa cells. Theriogenology 2020, 154, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, S.; Zhou, J.; Pang, X.; Wang, L. RNAi-mediated knockdown of MTNR1B without disrupting the effects of melatonin on apoptosis and cell cycle in bovine granulose cells. PeerJ 2018, 6, e4463. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, W.; Wen, A.; Yang, B.; Pang, X. Luzindole and 4P-PDOT block the effect of melatonin on bovine granulosa cell apoptosis and cell cycle depending on its concentration. PeerJ 2021, 9, e10627. [Google Scholar] [CrossRef]
- Lee, S.; Jin, J.X.; Taweechaipaisankul, A.; Kim, G.A.; Lee, B.C. Stimulatory Effects of Melatonin on Porcine In Vitro Maturation Are Mediated by MT2 Receptor. Int. J. Mol. Sci. 2018, 19, 1581. [Google Scholar] [CrossRef]
- Aglan, H.S.; Gebremedhn, S.; Salilew-Wondim, D.; Neuhof, C.; Tholen, E.; Holker, M.; Schellander, K.; Tesfaye, D. Regulation of Nrf2 and NF-kappaB during lead toxicity in bovine granulosa cells. Cell Tissue Res. 2020, 380, 643–655. [Google Scholar] [CrossRef]
- Qin, X.; Dang, W.; Yang, X.; Wang, K.; Kebreab, E.; Lyu, L. Neddylation inactivation affects cell cycle and apoptosis in sheep follicular granulosa cells. J. Cell. Physiol. 2022, 237, 3278–3291. [Google Scholar] [CrossRef]
- Tao, J.L.; Zhang, X.; Zhou, J.Q.; Li, C.Y.; Liu, H.L. Melatonin Alleviates Hypoxia-Induced Apoptosis of Granulosa Cells by Reducing ROS and Activating MTNR1B–PKA–Caspase8/9 Pathway. Antioxidants 2021, 10, 184. [Google Scholar] [CrossRef]
- Du, X.; Li, Q.; Cao, Q.; Wang, S.; Li, Q. Integrated Analysis of miRNA-mRNA Interaction Network in Porcine Granulosa Cells Undergoing Oxidative Stress. Oxidative Med. Cell. Longev. 2019, 1041583. [Google Scholar] [CrossRef]
- Wang, F.; Yin, Y.; Nie, X.; Zou, Y.; Tong, X.; Tong, Y.; Zhou, J.; Tan, Y. Protocatechuic acid alleviates polycystic ovary syndrome symptoms in mice by PI3K signaling in granulosa cells to relieve ROS pressure and apoptosis. Gynecol. Endocrinol. 2023, 39, 2228917. [Google Scholar] [CrossRef]
- Shen, M.; Cao, Y.; Jiang, Y.; Wei, Y.; Liu, H. melatonin protects mouse granulosa cells against oxidative damage by inhibiting foxo1-mediated autophagy: Implication of an antioxidation-independent mechanism. Redox Biol. 2018, 18, 138–157. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Wan, S.; Liu, S.; Wang, W.; Tang, M.; Bai, L.; Zhu, Y. LARS2 Regulates Apoptosis via ROS-Mediated Mitochondrial Dysfunction and Endoplasmic Reticulum Stress in Ovarian Granulosa Cells. Oxidative Med. Cell. Longev. 2022, 2022, 5501346. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.R.; Hay, M.P. Targeting hypoxia in cancer therapy. Nat. Rev. Cancer 2011, 11, 393–410. [Google Scholar] [CrossRef] [PubMed]
- Keith, B.; Simon, M.C.; Nakazawa, M.S. Oxygen availability and metabolic adaptations. Nat. Rev. Cancer 2016, 16, 663–673. [Google Scholar]
- Huang, M.; Li, X.; Jia, S.; Liu, S.; Yang, M. Bisphenol AF induces apoptosis via estrogen receptor beta (ERβ) and ROS-ASK1-JNK MAPK pathway in human granulosa cell line KGN. Environ. Pollut. 2020, 270, 116051. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, Q.; Hu, Y.; Xu, L.; Jiang, Y.; Zhang, C.; Ding, L.; Jiang, R.; Sun, J.; Sun, H.; et al. miR-181a increases FoxO1 acetylation and promotes granulosa cell apoptosis via SIRT1 downregulation. Cell Death Dis. 2017, 8, e3088. [Google Scholar] [CrossRef]
- Yadav, A.K.; Yadav, P.K.; Chaudhary, G.R.; Tiwari, M.; Gupta, A.; Sharma, A.; Pandey, A.N.; Pandey, A.K.; Chaube, S.K. Autophagy in hypoxic ovary. Cell. Mol. Life Sci. 2019, 76, 3311–3322. [Google Scholar] [CrossRef]
- Yang, H.; Xie, Y.; Yang, D.; Ren, D. Oxidative stress-induced apoptosis in granulosa cells involves JNK, p53 and Puma. Oncotarget 2017, 8, 25310–25322. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.C.; Duan, C.C.; Jin, S.; Sheng, C.B.; Wang, Y.S.; Yue, Z.P.; Guo, B. HB-EGF induces mitochondrial dysfunction via estrogen hypersecretion in granulosa cells dependent on cAMP-PKA-JNK/ERK-Ca(2+)-FOXO1 pathway. Int. J. Biol. Sci. 2022, 18, 2047–2059. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.P.; Jian, Z.; Ma, R.Y.; Cao, Z.Z.; Zhu, Y.; Zhu, Y.; Tang, F.Q.; Xiao, Y.B. Melatonin alleviates hypoxia-induced cardiac apoptosis through PI3K/Akt pathway. Int. J. Clin. Exp. Pathol. 2018, 11, 5840–5849. [Google Scholar]
- Zhao, S.; Wang, Y.; Zhang, X.; Zheng, L.; Zhu, B.; Yao, S.; Yang, L.; Du, J. Melatonin Protects Against Hypoxia/Reoxygenation-Induced Dysfunction of Human Umbilical Vein Endothelial Cells Through Inhibiting Reactive Oxygen Species Generation. Acta Cardiol. Sin. 2018, 34, 424–431. [Google Scholar]
- Zhao, X.M.; Wang, N.; Hao, H.S.; Li, C.Y.; Zhao, Y.H.; Yan, C.L.; Wang, H.Y.; Du, W.H.; Wang, D.; Liu, Y.; et al. Melatonin improves the fertilization capacity and developmental ability of bovine oocytes by regulating cytoplasmic maturation events. J. Pineal Res. 2018, 64, e12445. [Google Scholar] [CrossRef]
- He, Y.; Deng, H.; Jiang, Z.; Li, Q.; Shi, M.; Chen, H.; Han, Z. Effects of melatonin on follicular atresia and granulosa cell apoptosis in the porcine. Mol. Reprod. Dev. 2016, 83, 692–700. [Google Scholar] [CrossRef]
- Muller, F.L.; Lustgarten, M.S.; Jang, Y.; Richardson, A.; Remmen, H.V. Trends in oxidative aging theories. Free Radic. Biol. Med. 2007, 43, 477–503. [Google Scholar] [CrossRef] [PubMed]
- Elmorsy, E.; Al-Ghafari, A.; Aggour, A.M.; Khan, R.; Amer, S. The role of oxidative stress in antipsychotics induced ovarian toxicity. Toxicol. Vitr. 2017, 44, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, E.R. Importance of individuality in oxidative stress and aging—ScienceDirect. Free Radic. Biol. Med. 2002, 33, 597–604. [Google Scholar] [CrossRef]
- Liu, Z.F.; Liu, K.; Liu, Z.Q.; Cong, L.; Lei, M.Y.; Li, J.; Ma, Z.; Deng, Y.; Liu, W.; Xu, B. Melatonin attenuates manganese-induced mitochondrial fragmentation by suppressing the Mst1/JNK signaling pathway in primary mouse neurons. Sci Total Environ. 2022, 844, 157134. [Google Scholar] [CrossRef] [PubMed]
- Nikmard, F.; Hosseini, E.; Bakhtiyari, M.; Ashrafi, M.; Amidi, F.; Aflatoonian, R. Effects of melatonin on oocyte maturation in PCOS mouse model. Anim. Sci. J. = Nihon Chikusan Gakkaiho 2017, 88, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Tagliaferri, V.; Romualdi, D.; Scarinci, E.; Cicco, S.D.; Florio, C.D.; Immediata, V.; Tropea, A.; Santarsiero, C.M.; Lanzone, A.; Apa, R. Melatonin Treatment May Be Able to Restore Menstrual Cyclicity in Women With PCOS: A Pilot Study. Reprod. Sci. 2018, 25, 269–275. [Google Scholar] [CrossRef]
- Xie, Q.E.; Wang, M.Y.; Cao, Z.P.; Du, X.; Ji, D.M.; Liang, D.; Cao, Y.X.; Liu, Y.J. Melatonin protects against excessive autophagy-induced mitochondrial and ovarian reserve function deficiency though ERK signaling pathway in Chinese hamster ovary (CHO) cells. Mitochondrion 2021, 61, 44–53. [Google Scholar] [CrossRef]
- Abo El Gheit, R.E.; Soliman, N.A.; Nagla, S.A.; El-Sayed, R.M.; Badawi, G.A.; Emam, M.N.; Abdel Ghafar, M.T.; Ibrahim, M.A.; Elswaidy, N.R.; Radwan, D.A.; et al. Melatonin epigenetic potential on testicular functions and fertility profile in varicocele rat model is mediated by silent information regulator 1. Br. J. Pharmacol. 2022, 179, 3363–3381. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (5′–3′) | GenBank Accession NO. |
|---|---|---|
| HMOX-1 | F: CCGAGAAGGCTTTAAGCTGGT R: GGAAGTAGAGGGGCGTGTAG | NM_001004027.1 |
| TRX1 | F: CTTTACCTTATTGCCCGGGT | NM_214313.2 |
| R: GTTCACCGATTTTGTTGGCC | ||
| SOD2 | F: AGGCGCTGAAAAAGGGTGAT R: AAGTCGCGTTTGATGGCTTC | NM_214127.2 |
| CAT | F: CACACATACCCATTCGTCACT R: CAGCCCTAACCTTCACTTACC | NM_214301.2 |
| GPX4 | F: ATTCTCAGCCAAGGACATCG R: CCTCATTGAGAGGCCACATT | NM_214407.1 |
| TUBA1A | F: AAGAGTCGCGCTGTAAGAAG R: AATGACTGTGGGTTCCAGGTC | NM_001315710.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zhang, D.; Li, H.; Liu, Z.; Yang, Y.; Li, J.; Tang, L.; Tao, J.; Liu, H.; Shen, M. Melatonin-Mediated Suppression of mtROS-JNK-FOXO1 Pathway Alleviates Hypoxia-Induced Apoptosis in Porcine Granulosa Cells. Antioxidants 2023, 12, 1881. https://doi.org/10.3390/antiox12101881
Zhang X, Zhang D, Li H, Liu Z, Yang Y, Li J, Tang L, Tao J, Liu H, Shen M. Melatonin-Mediated Suppression of mtROS-JNK-FOXO1 Pathway Alleviates Hypoxia-Induced Apoptosis in Porcine Granulosa Cells. Antioxidants. 2023; 12(10):1881. https://doi.org/10.3390/antiox12101881
Chicago/Turabian StyleZhang, Xuan, Dingding Zhang, Hongmin Li, Zhaojun Liu, Yatong Yang, Jiameng Li, Lishiyuan Tang, Jingli Tao, Honglin Liu, and Ming Shen. 2023. "Melatonin-Mediated Suppression of mtROS-JNK-FOXO1 Pathway Alleviates Hypoxia-Induced Apoptosis in Porcine Granulosa Cells" Antioxidants 12, no. 10: 1881. https://doi.org/10.3390/antiox12101881