
Dietary Taurine Improves Growth Performance and Intestine Health via the GSH/GSSG Antioxidant System and Nrf2/ARE Signaling Pathway in Weaned Piglets
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Material
2.2. Experimental Design and Diets
2.3. Sample Collection
2.4. Biochemical Parameters of Oxidative Stress and Indicators of Intestinal Barrier Integrity
2.5. Real-Time RT-PCR
2.6. H and E Staining
2.7. Cecal Microbial 16S rDNA Sequencing Analysis
2.8. Data Treatments and Statistic Analysis
3. Results
3.1. Growth Performance
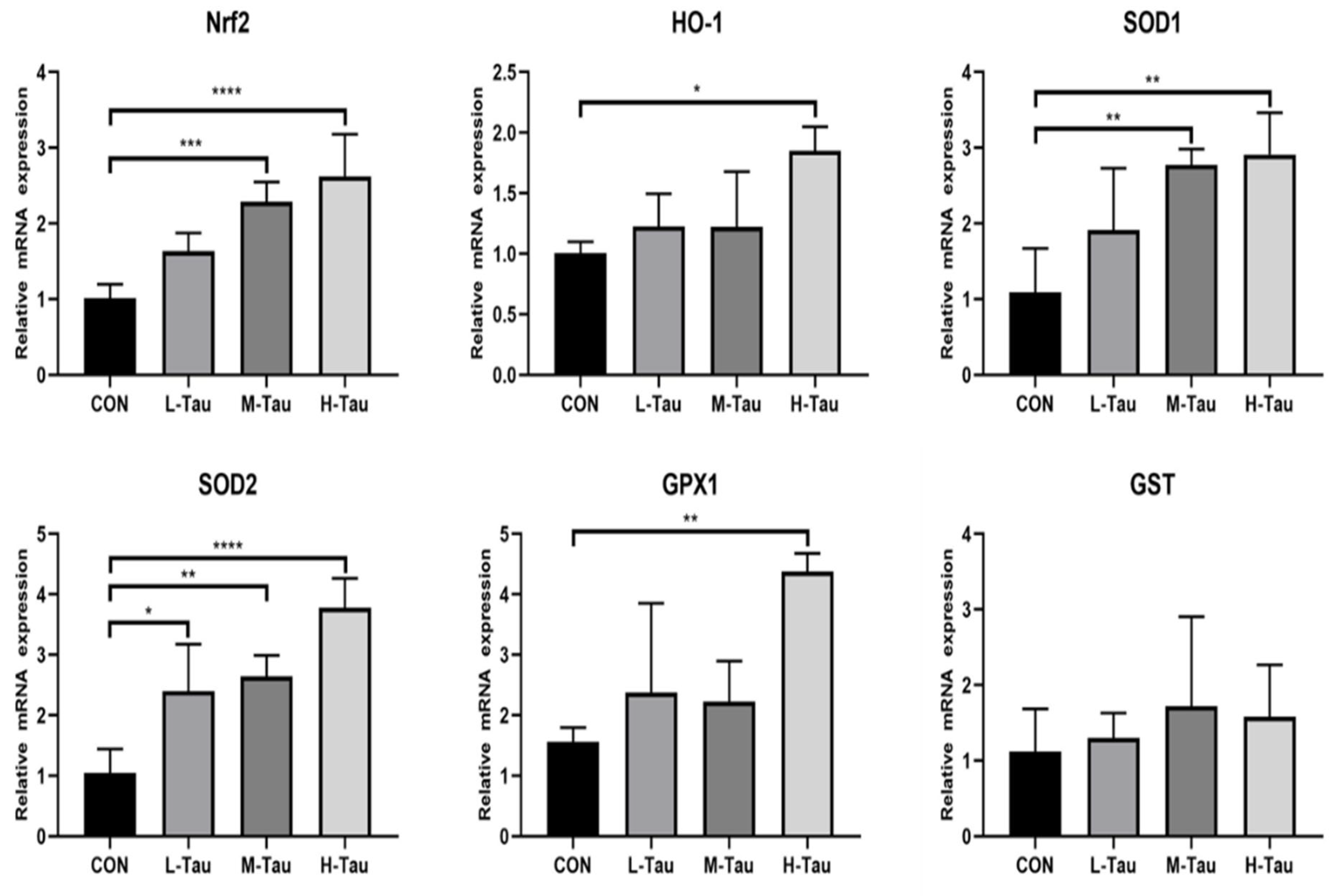
3.2. Antioxygenic Capacity
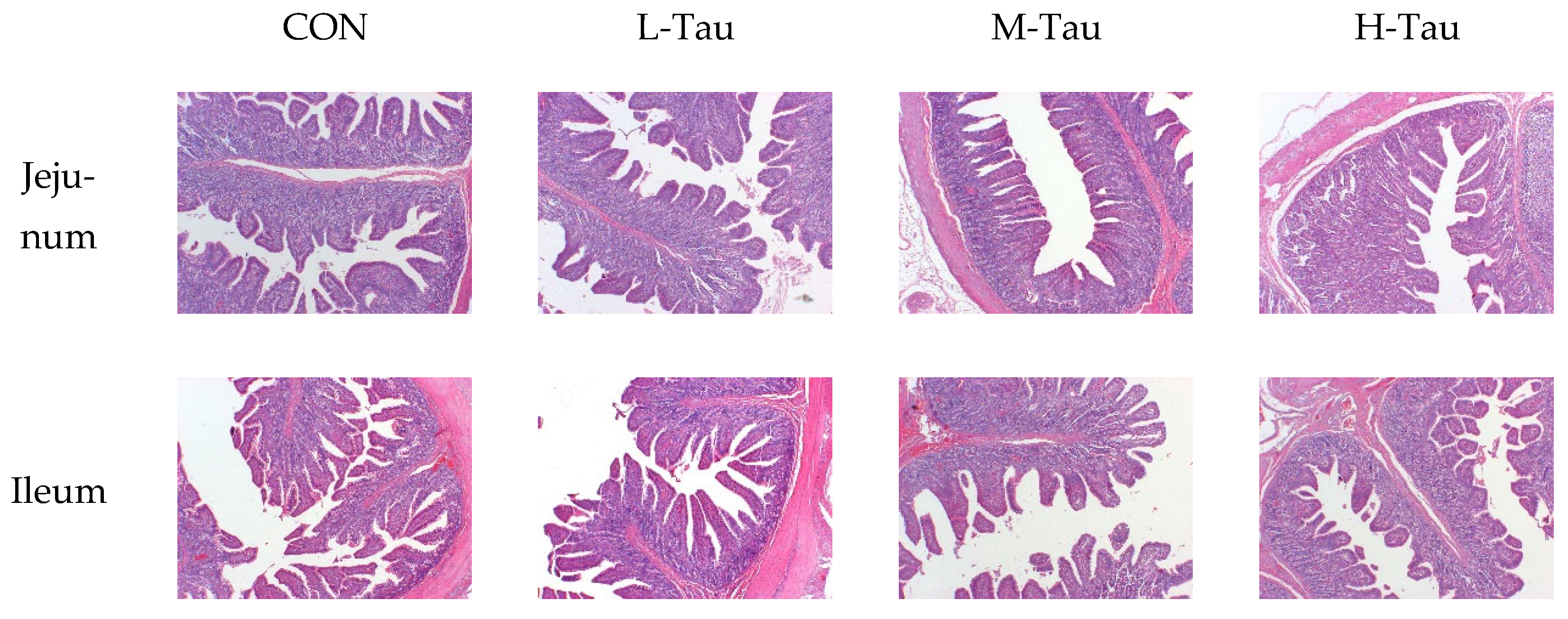
3.3. Intestinal Morphology
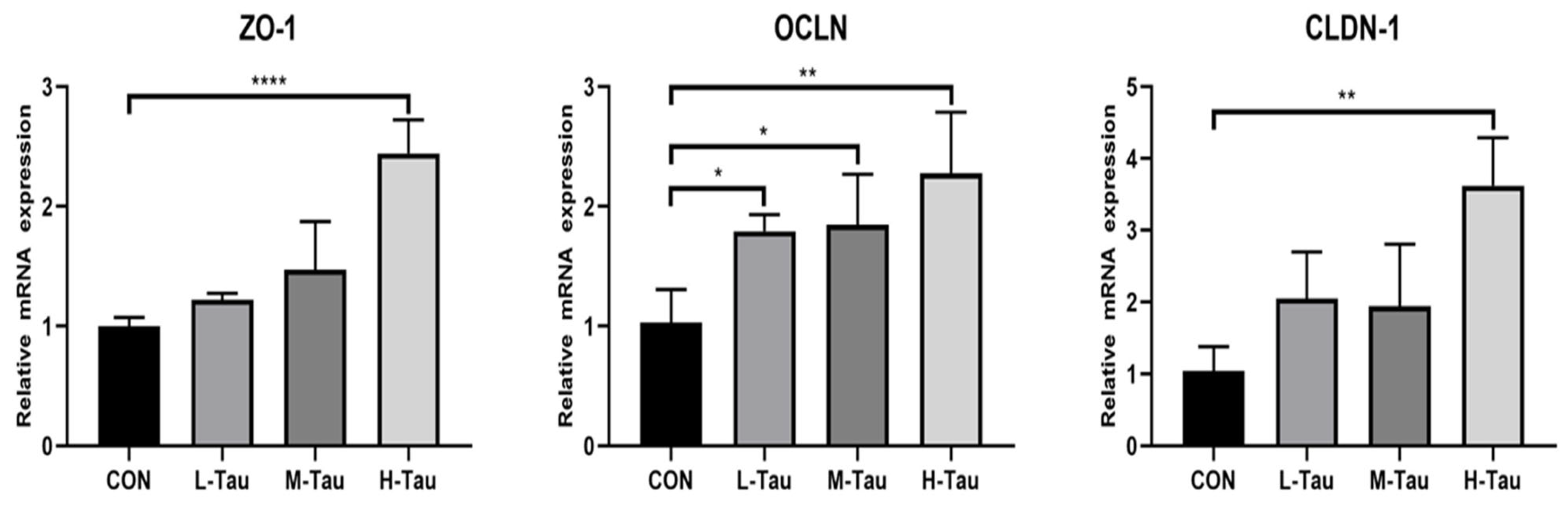
3.4. Intestinal Mucosal Barrier
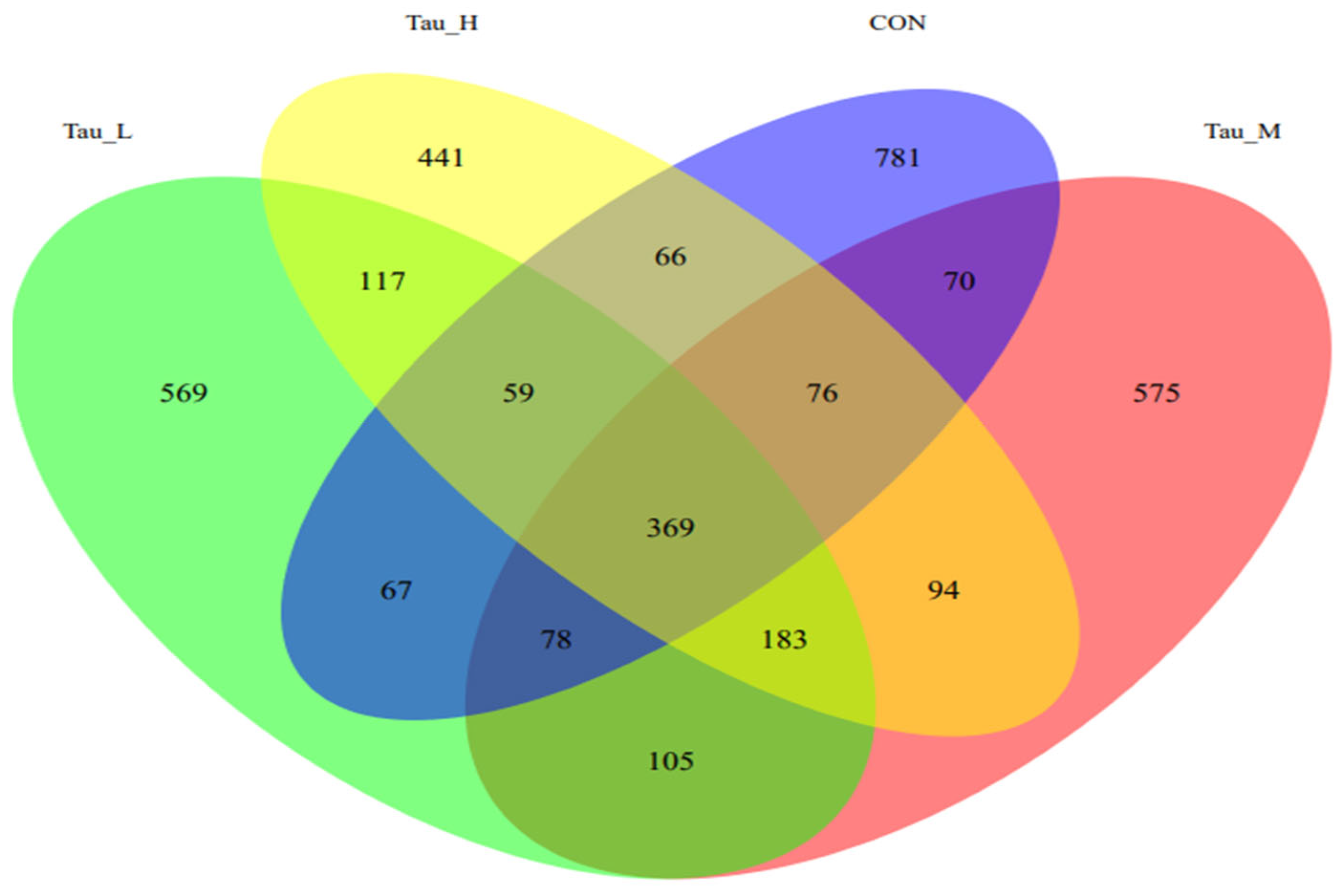
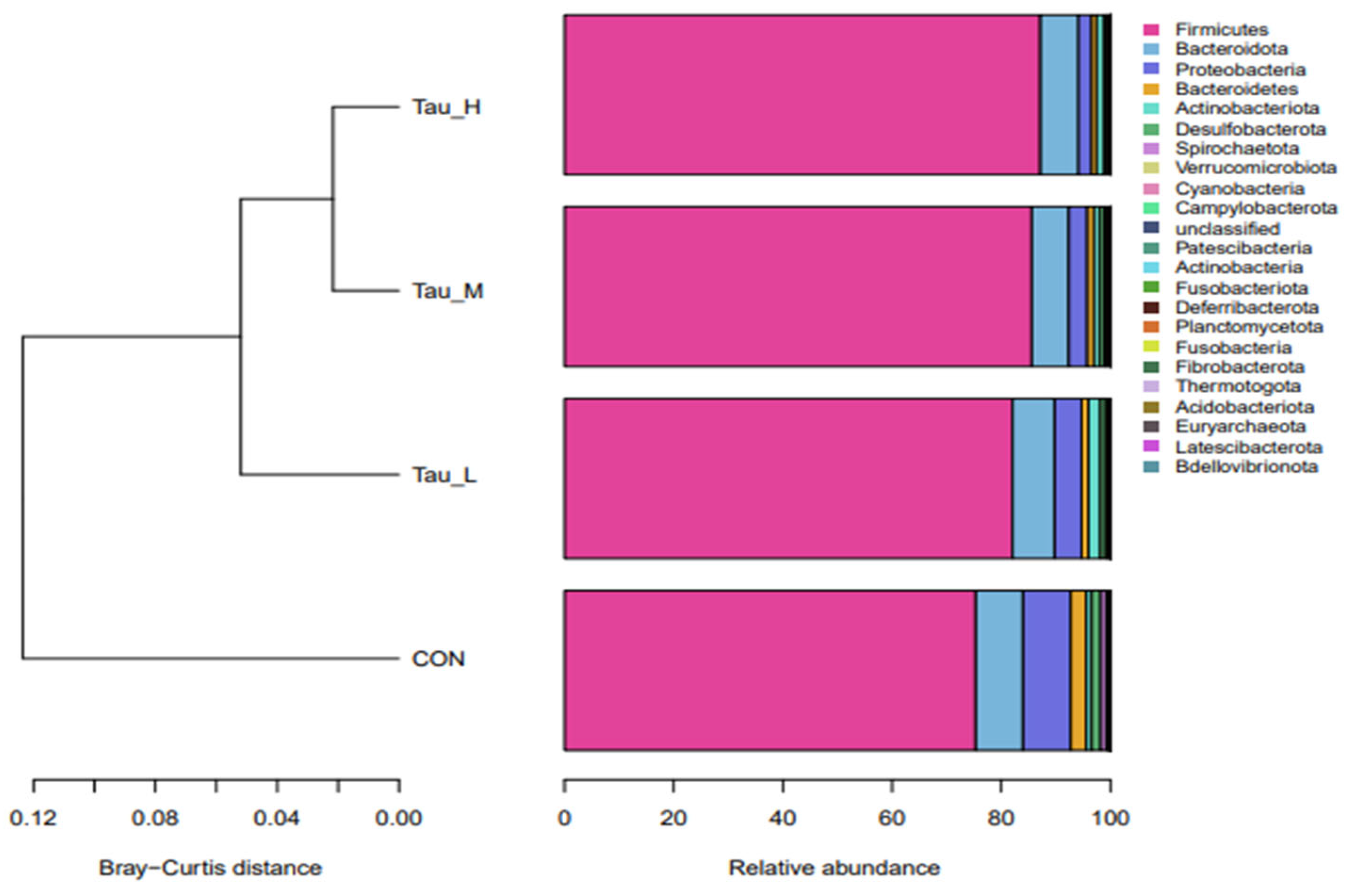
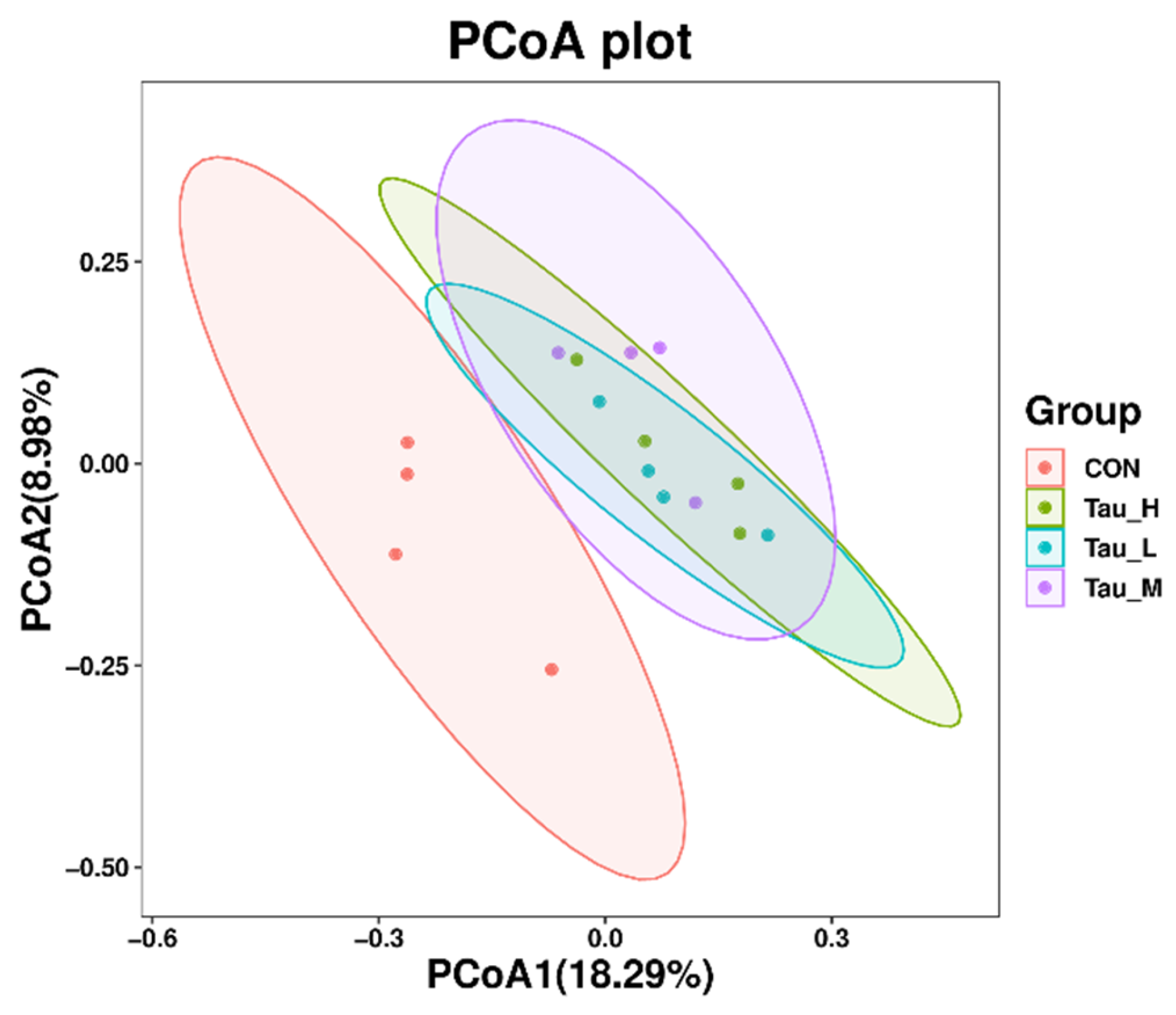
3.5. Cecum Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ipema, A.F.; Bokkers, E.A.M.; Gerrits, W.J.J.; Kemp, B.; Bolhuis, J.E. Providing live black soldier fly larvae (Hermetia illucens) improves welfare while maintaining performance of piglets post-weaning. Sci. Rep. 2021, 11, 7371. [Google Scholar] [CrossRef]
- Pluske, J.R.; Hampson, D.J.; Williams, I.H. Factors influencing the structure and function of the small intestine in the weaned pig: A review. Livest. Prod. Sci. 1997, 51, 215–236. [Google Scholar] [CrossRef]
- Weary, D.M.; Jasper, J.; Hötzel, M.J. Understanding weaning distress. Appl. Anim. Behav. Sci. 2008, 110, 24–41. [Google Scholar] [CrossRef]
- Novais, A.K.; Deschêne, K.; Martel-Kennes, Y.; Roy, C.; Laforest, J.P.; Lessard, M.; Matte, J.J.; Lapointe, J. Weaning differentially affects mitochondrial function, oxidative stress, inflammation and apoptosis in normal and low birth weight piglets. PLoS ONE 2021, 16, e0247188. [Google Scholar] [CrossRef]
- Zhu, L.; Zhao, K.; Chen, X.; Xu, J. Impact of weaning and an antioxidant blend on intestinal barrier function and antioxidant status in pigs. J. Anim. Sci. 2013, 91, 1522, Erratum in J. Anim. Sci. 2013, 91, 1522. [Google Scholar] [CrossRef]
- In, J.; Ren, W.K.; Wu, X.S. Oxidative stress-mediated signaling pathways: A review. J. Agr. Food Chem. 2013, 11, 132–139. [Google Scholar]
- Huxtable, R.J. Physiological actions of taurine. Physiol. Rev. 1992, 72, 101–163. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, J.T.; Brosnan, M.E. The sulfur-containing amino acids: An overview. J. Nutr. 2006, 136, 1636–1640. [Google Scholar] [CrossRef]
- Kulczyński, B.; Sidor, A.; Gramza-Michałowska, A. Characteristics of Selected Antioxidative and Bioactive Compounds in Meat and Animal Origin Products. Antioxidants 2019, 8, 335. [Google Scholar] [CrossRef]
- Surai, P.F.; Earle-Payne, K.; Kidd, M.T. Taurine as a Natural Antioxidant: From Direct Antioxidant Effects to Protective Action in Various Toxicological Models. Antioxidants 2021, 10, 1876. [Google Scholar] [CrossRef]
- Jong, C.J.; Azuma, J.; Schaffer, S. Mechanism underlying the antioxidant activity of taurine: Prevention of mitochondrial oxidant production. Amino Acids 2012, 42, 2223–2232. [Google Scholar] [CrossRef] [PubMed]
- Jong, C.J.; Azuma, J.; Schaffer, S.W. Role of mitochondrial permeability transition in taurine deficiency-induced apoptosis. Exp. Clin. Cardiol. 2011, 16, 125–128. [Google Scholar] [PubMed]
- Ricci, C.; Pastukh, V.; Leonard, J.; Turrens, J.; Wilson, G.; Schaffer, D.; Schaffer, S.W. Mitochondrial DNA damage triggers mitochondrial-superoxide generation and apoptosis. Am. J. Physiol. Cell Physiol. 2008, 294, C413–C422. [Google Scholar] [CrossRef] [PubMed]
- Shetewy, A.; Shimada-Takaura, K.; Warner, D.; Jong, C.J.; Mehdi, A.B.; Alexeyev, M.; Takahashi, K.; Schaffer, S.W. Mitochondrial defects associated with β-alanine toxicity: Relevance to hyper-beta-alaninemia. Mol. Cell. Biochem. 2016, 416, 11–22. [Google Scholar] [CrossRef]
- Cheong, S.H.; Lee, D.S. Taurine Chloramine Prevents Neuronal HT22 Cell Damage Through Nrf2-Related Heme Oxygenase-1. Adv. Exp. Med. Biol. 2017, 975 Pt 1, 145–157. [Google Scholar] [CrossRef]
- Hansen, S.H.; Grunnet, N. Taurine, glutathione and bioenergetics. Adv. Exp. Med. Biol. 2013, 776, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, Y.; Yan, G.; Bu, Q.; Lv, L.; Yang, Y.; Zhao, J.; Shao, X.; Deng, Y.; Zhu, R.; et al. Protective role of taurine against morphine-induced neurotoxicity in C6 cells via inhibition of oxidative stress. Neurotox. Res. 2011, 20, 334–342. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Swine, 11th ed.; National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Hart, G.K.; Dobb, G.J. Effect of a fecal bulking agent on diarrhea during enteral feeding in the critically ill. JPEN J. Parenter. Enter. Nutr. 1988, 12, 465–468. [Google Scholar] [CrossRef]
- Yin, J.; Wu, M.M.; Xiao, H.; Ren, W.K.; Duan, J.L.; Yang, G.; Li, T.J.; Yin, Y.L. Development of an antioxidant system after early weaning in piglets. J. Anim. Sci. 2014, 92, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Cai, X.; Xu, C.; Luo, Z.; Sheng, Y.; Bao, J.; Chen, X.; Xu, J. Effects of dietary supplementation with N-acetyl cysteine on antioxidant capacities and the expression of inflammatory cytokines in weaned piglets. J. Anim. Sci. 2016, 15, 634–641. [Google Scholar] [CrossRef]
- Luo, Z.; Zhu, W.; Guo, Q.; Luo, W.; Zhang, J.; Xu, W.; Xu, J. Weaning Induced Hepatic Oxidative Stress, Apoptosis, and Aminotransferases through MAPK Signaling Pathways in Piglets. Oxidative Med. Cell. Longev. 2016, 2016, 4768541. [Google Scholar] [CrossRef]
- Zhao, Y.; Niu, Y.; He, J.; Zhang, L.; Wang, C.; Wang, T. Dietary Dihydroartemisinin Supplementation Attenuates Hepatic Oxidative Damage of Weaned Piglets with Intrauterine Growth Retardation through the Nrf2/ARE Signaling Pathway. Animals 2019, 9, 1144. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Yamamoto, M. Stress-sensing mechanisms and the physiological roles of the Keap1-Nrf2 system during cellular stress. J. Biol. Chem. 2017, 292, 16817–16824. [Google Scholar] [CrossRef]
- Wu, S.; Lu, H.; Bai, Y. Nrf2 in cancers: A double-edged sword. Cancer Med. 2019, 8, 2252–2267. [Google Scholar] [CrossRef]
- Tomoda, E.; Nagao, A.; Shirai, Y.; Asano, K.; Suzuki, T.; Battersby, B.J.; Suzuki, T. Restoration of mitochondrial function through activation of hypomodified tRNAs with pathogenic mutations associated with mitochondrial diseases. Nucleic Acids Res. 2023, 51, 7563–7579. [Google Scholar] [CrossRef]
- Kirino, Y.; Goto, Y.; Campos, Y.; Arenas, J.; Suzuki, T. Specific correlation between the wobble modification deficiency in mutant tRNAs and the clinical features of a human mitochondrial disease. Proc. Natl. Acad. Sci. USA 2005, 102, 7127–7132. [Google Scholar] [CrossRef]
- Kirino, Y.; Yasukawa, T.; Ohta, S.; Akira, S.; Ishihara, K.; Watanabe, K.; Suzuki, T. Codon-specific translational defect caused by a wobble modification deficiency in mutant tRNA from a human mitochondrial disease. Proc. Natl. Acad. Sci. USA 2004, 101, 15070–15075. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Lv, Q.; Zhao, D.; Wang, J.; Fu, Y.; Li, C.; Wu, G.; Liu, M.; Hu, J.; Lin, S.; et al. Intervention Effect of Taurine on LPS-Induced Intestinal Mechanical Barrier Injury in Piglets. Adv. Exp. Med. Biol. 2022, 1370, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Biehs, B.; Warming, S.; Leong, K.G.; Rangell, L.; Klein, O.D.; De Sauvage, F.J. A reserve stem cell population in small intestine renders Lgr5-positive cells dispensable. Nature 2011, 478, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Chia, L.A.; Kuo, C.J. The intestinal stem cell. Prog. Mol. Biol. Transl. Sci. 2010, 96, 157–173. [Google Scholar] [CrossRef]
- Umar, S. Intestinal stem cells. Curr. Gastroenterol. Rep. 2010, 12, 340–348. [Google Scholar] [CrossRef]
- Lou, W.; Liu, D. Hyperoxia reduces the expression of Bmi1 and Lgr5 in intestinal stem cells of neonatal rats. Basic Clin. Med. 2019, 39, 22–26. [Google Scholar]
- Li, Y.; Ma, S.; Zhang, Y.; Yao, M.; Zhu, X.; Guan, F. (-)-Epicatechin mitigates radiation-induced intestinal injury and promotes intestinal regeneration via suppressing oxidative stress. Free Radic. Res. 2019, 53, 851–864. [Google Scholar] [CrossRef]
- Wakabayashi, N.; Chartoumpekis, D.V.; Kensler, T.W. Crosstalk between Nrf2 and Notch signaling. Free Radic. Biol. Med. 2015, 88 Pt B, 158–167. [Google Scholar] [CrossRef]
- Yagishita, Y.; McCallum, M.L.; Kensler, T.W.; Wakabayashi, N. Constitutive Activation of Nrf2 in Mice Expands Enterogenesis in Small Intestine Through Negative Regulation of Math1. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 503–524. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Wu, H.; Wang, C.; Zhang, Q.; Jiao, L.; Lin, F.; Hu, C. Diquat-induced oxidative stress increases intestinal permeability, impairs mitochondrial function, and triggers mitophagy in piglets. J. Anim. Sci. 2018, 96, 1795–1805. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Li, W.; Su, Y.; Liu, S.; Kan, X.; Ran, X.; Cao, Y.; Fu, S.; Liu, J. GPR109A alleviate mastitis and enhances the blood milk barrier by activating AMPK/Nrf2 and autophagy. Int. J. Biol. Sci. 2021, 17, 4271–4284. [Google Scholar] [CrossRef]
- Singh, R.; Chandrashekharappa, S.; Bodduluri, S.R.; Baby, B.V.; Hegde, B.; Kotla, N.G.; Hiwale, A.A.; Saiyed, T.; Patel, P.; Vijay-Kumar, M.; et al. Enhancement of the gut barrier integrity by a microbial metabolite through the Nrf2 pathway. Nat. Commun. 2019, 10, 89. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Liu, W.; Li, X.; Chen, W.; Liu, Z.; Wen, J.; Liu, Z. A Protective Role of the NRF2-Keap1 Pathway in Maintaining Intestinal Barrier Function. Oxid. Med. Cell. Longev. 2019, 2019, 1759149. [Google Scholar] [CrossRef]
- Chen, H.; Hu, Y.; Fang, Y.; Djukic, Z.; Yamamoto, M.; Shaheen, N.J.; Orlando, R.C.; Chen, X. Nrf2 deficiency impairs the barrier function of mouse oesophageal epithelium. Gut 2014, 63, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Liu, J.; Chen, Z.; Huang, K.; Li, Y.; Li, C. Protective effects of sugar cane extract against oxidative damage induced by diquat and mechanism mediated by intestinal microbiota in weaned piglets. J. Nanjing Agric. Univ. 2020, 3, 514–522. [Google Scholar]
- Baarlen, P.V.; Wells, J.M.; Kleerebezem, M. Regulation of intestinal homeostasis and immunity with probiotic lactobacilli. Trends Immunol. 2013, 34, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Miquel, S.; Martín, R.; Rossi, O.; Bermúdez-Humarán, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium prausnitzii and human intestinal health. Curr. Opin. Microbiol. 2013, 16, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.A.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.K.; Lynch, S.V.; Knight, R. Current understanding of the human microbiome. Nat. Med. 2018, 24, 392–400. [Google Scholar] [CrossRef]
- Dai, J. Effects of Taurine on LPS Mediated Caecumepithelial Tight Junction Protein, Flora and Metabolites in Piglets; Shenyang Agricultural University: Shenyang, China, 2022. [Google Scholar] [CrossRef]
- Thomas, F.; Hehemann, J.H.; Rebuffet, E.; Czjzek, M.; Michel, G. Environmental and gut bacteroidetes: The food connection. Front. Microbiol. 2011, 2, 93. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Bervoets, L.; Van Hoorenbeeck, K.; Kortleven, I.; Van Noten, C.; Hens, N.; Vael, C.; Goossens, H.; Desager, K.N.; Vankerckhoven, V. Differences in gut microbiota composition between obese and lean children: A cross-sectional study. Gut Pathog. 2013, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Yu, Y.; Wan, J.; Mao, Y.; Zhang, H.; Zhang, J.; Tian, X.; Zhao, Q. Progress in research on the relationship between Proteobacteria and the imbalance of mammalian colonic intestinal flora. Chin. J. Microecol. 2022, 34, 479–484. [Google Scholar] [CrossRef]
- Råsbäck, T.; Jansson, D.S.; Johansson, K.E.; Fellström, C. A novel enteropathogenic, strongly haemolytic spirochaete isolated from pig and mallard, provisionally designated ‘Brachyspira suanatina’ sp. Environ. Microbiol. 2007, 9, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Liu, X. Effects of Different Fermented Feeds on Growth Performance and Meat Quality and Intestinal Microorganism; Henan Agricultural University: Zhengzhou, China, 2019. [Google Scholar] [CrossRef]
- Bauchart-Thevret, C.; Stoll, B.; Chacko, S.; Burrin, D.G. Sulfur amino acid deficiency upregulates intestinal methionine cycle activity and suppresses epithelial growth in neonatal pigs. Am. J. Physiol. Endocrinol. Metab. 2009, 296, 1239–1250. [Google Scholar] [CrossRef]
- Li, L. Diversity and Metabolic Characteristics of Lactic Acid Bacteria and Lactate-Degrading Bacteria in Fermented Grains of Luzhou-Flavor Liquor; Jiangnan University: Wuxi, China, 2016. [Google Scholar]
- Song, Z.; Jiang, N.; Zhang, A. Research progress on biological functions of glutathione. Feed Res. 2008, 9, 25–27. [Google Scholar]
- Liu, X. Antioxidative Defence Mechanism of Glutathione on Litopenaeus vannamei; Huazhong Agricultural University: Wuhan, China, 2010. [Google Scholar]
- Ziegler, D.M. Role of reversible oxidation-reduction of enzyme thiols-disulfides in metabolic regulation. Annu. Rev. Biochem. 1985, 54, 305–329. [Google Scholar] [CrossRef] [PubMed]
- Long, M. A Study of Effects of N-acetylcysteine on Growth and Viability of Intestinal Porcine Epithelial Cell and the Related Mechanisms; Wuhan Polytechnic University: Wuhan, China, 2014. [Google Scholar]
- Huang, R.; Peng, Z. Effect of diet type and dietary turine supplementation on growth performance of weaning pigs. Cereal Feed Ind. 2008, 9, 44–45. [Google Scholar]
- Spitze, A.R.; Wong, D.L.; Rogers, Q.R.; Fascetti, A.J. Taurine concentrations in animal feed ingredients; cooking influences taurine content. J. Anim. Physiol. Anim. Nutr. 2003, 87, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.S.; Ki, K.N.; Chung, H.Y. Proximate composition, amino Acid, mineral, and heavy metal content of dried laver. Prev. Nutr. Food Sci. 2013, 18, 139–144. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Contents | Nutritional Level 2 | Contents |
|---|---|---|---|
| Corn (%) | 61.49 | DE(MJ/kg) | 14.34 |
| Soybean meal (%) | 12.00 | CP (%) | 18.42 |
| Puffed soybean meal (%) | 6.08 | Ca (%) | 0.78 |
| Fish meal (%) | 5.00 | Total P (%) | 0.67 |
| Whey powder (%) | 10.00 | Available P (%) | 0.40 |
| Fatty powder (%) | 1.59 | Lysine (%) | 1.42 |
| Calcium hydrophosphate (%) | 0.93 | Methionine (%) | 0.51 |
| Mountain flour (%) | 0.64 | Lysine + Cystine (%) | 0.78 |
| NaCl (%) | 0.30 | Threonine (%) | 0.90 |
| L-lysine hydrochloride (%) | 0.57 | Tryptophan (%) | 0.21 |
| Methionine (%) | 0.21 | Taurine | 0.037 |
| Threonine (%) | 0.20 | ||
| Premix 1 (%) | 1.00 | ||
| Total (%) | 100 |
| Gene Name | ID Number | Primer Sequence | Product Length (bp) |
|---|---|---|---|
| GAPDH | XM_021091114 | F: CGAGATCCCGCCAACATCAA R: CCCCACCCTTCAAGTGAGC | 109 |
| Nrf2 | MH101365.1 | F: TGCAGCTTTTGGCAGAGACA R: AGGAGCAATGAAGACTGGGC | 119 |
| ZO-1 | AJ318101.1 | F: ATTCCTGAGGGAATTGGGCAG R: GCAGGGTTTCACCTTTCTCCT | 94 |
| OCLN | NM_001163647.2 | F: ATATGGTGGAGAGACGCCCA R: AAGGGTAGCCCATACCACCT | 230 |
| SOD2 | NM_214127.2 | F: TTGCAGATTGCCGCTTGTTC R: TTACTTTTTGCAAGCGGCGT | 195 |
| GPX1 | NM_214201.1 | F: GAAGTGTGAGGTGAATGGCG R: TGCGATGTCATTGCGACACA | 145 |
| GST | NM_214300.2 | F: CTCAAGCTTTGGCAAGGGAG R: TGGGCTCTTCGTACACGTTC | 78 |
| HO-1 | NM_001004027.1 | F: TACCGCTCCCGAATGAACAC R: GTCACGGGAGTGGAGTCTTG | 209 |
| SOD1 | NM_001190422.1 | F: AAGGCCGTGTGTGTGCTGAA R: AGTGGCCACACCATCTTTGC | 279 |
| CLDN-1 | XM_005670262.3 | F: CTAGTGATGAGGCAGATGAA R: AGATAGGTCCGAAGCAGAT | 250 |
| Items | Treatments | SEM | p Value | |||||
|---|---|---|---|---|---|---|---|---|
| CON | L-Tau | M-Tau | H-Tau | Linear | Quadratic | Cubic | ||
| Initial weight (kg) | 6.73 | 6.74 | 6.73 | 6.73 | 0.22 | 1.00 | 1.00 | 1.00 |
| Final weight (kg) | 12.03 b | 13.09 a | 13.22 a | 13.54 a | 0.255 | 0.001 | 0.026 | 0.078 |
| Weight gain (kg) | 5.81 b | 6.26 b | 6.49 a | 7.05 a | 0.156 | 0.001 | 0.028 | 0.058 |
| ADG (kg/d) | 166 b | 179 b | 186 a | 194 a | 4.46 | 0.047 | 0.028 | 0.058 |
| ADFI (kg/d) | 442 | 458 | 459 | 462 | 17.87 | 0.837 | 0.773 | 0.637 |
| Feed conversion rate | 0.38 b | 0.39 b | 0.40 a | 0.42 a | 0.015 | 0.03 | 0.036 | 0.091 |
| Diarrhea index | 1.28 a | 1.05 b | 0.87 bc | 0.72 c | 0.22 | <0.001 | 0.573 | 0.973 |
| Items | Treatments | SEM | p Value | |||||
|---|---|---|---|---|---|---|---|---|
| CON | L-Tau | M-Tau | H-Tau | Linear | Quadratic | Cubic | ||
| Serum | ||||||||
| MDA (nmol/L) | 11.2 a | 12.5 a | 9.66 ab | 6.97 b | 1.86 | 0.003 | 0.053 | 0.320 |
| NO (μmoL/mL) | 0.34 a | 0.34 a | 0.25 b | 0.24 b | 0.04 | 0.001 | 0.935 | 0.100 |
| SOD (U/mL) | 209 b | 197 b | 249 b | 387 a | 46.51 | <0.001 | 0.007 | 0.809 |
| GPX (U/mL) | 671 b | 701 b | 841 a | 842 a | 64.46 | 0.003 | 0.686 | 0.118 |
| CAT (U/mL) | 17.40 b | 15.72 b | 24.16 a | 24.14 a | 2.57 | 0.001 | 0.546 | 0.009 |
| POD (U/mL) | 6.25 b | 6.52 b | 8.75 ab | 12.99 a | 2.82 | 0.007 | 0.204 | 0.996 |
| T-AOC (μmol/mL) | 0.12 c | 0.13 bc | 0.15 b | 0.19 a | 0.02 | <0.001 | 0.162 | 0.833 |
| O2•− clearance rate (%) | 17.35 b | 16.96 b | 17.81 ab | 18.64 a | 0.73 | 0.013 | 0.119 | 0.457 |
| •OH clearance rate (%) | 3.50 b | 3.54 b | 3.65 b | 3.97 a | 0.20 | <0.001 | 0.187 | 0.787 |
| GSH (μmol/mL) | 0.09 ab | 0.08 b | 0.13 ab | 0.19 a | 0.06 | 0.033 | 0.310 | 0.717 |
| GSSG (nmol/mL) | 3.67 ab | 3.81 a | 3.61 ab | 2.71 b | 0.62 | 0.050 | 0.138 | 0.810 |
| GSH/GSSG | 25.29 b | 22.71 b | 36.75 ab | 80.79 a | 30.52 | 0.019 | 0.145 | 0.845 |
| Liver | ||||||||
| O2•− clearance rate (%) | 17.37 b | 16.99 b | 18.21 a | 18.24 a | 0.42 | 0.004 | 0.396 | 0.021 |
| •OH clearance rate (%) | 3.38 b | 3.64 ab | 3.79 ab | 4.00 a | 0.23 | 0.005 | 0.867 | 0.763 |
| POD (U/mg prot) | 4.71 ab | 3.93 b | 5.62 ab | 6.62 a | 1.41 | 0.040 | 0.252 | 0.372 |
| GSHS (U/g) | 0.42 c | 0.39 c | 0.49 b | 0.58 a | 0.04 | <0.001 | 0.013 | 0.099 |
| Jejunal mucosa | ||||||||
| POD (U/mg prot) | 2.91 b | 2.94 b | 4.79 ab | 6.33 a | 1.76 | 0.011 | 0.429 | 0.621 |
| O2•− clearance rate (%) | 16.37 b | 15.88 b | 16.63 b | 18.03 a | 0.45 | <0.001 | 0.002 | 0.572 |
| •OH clearance rate (%) | 3.36 b | 3.48 b | 3.91 a | 4.20 a | 0.20 | <0.001 | 0.413 | 0.366 |
| GSHS (U/g) | 0.38 b | 0.34 b | 0.41 b | 0.69 a | 0.05 | <0.001 | <0.001 | 0.423 |
| Items | Treatments | SEM | p Value | |||||
|---|---|---|---|---|---|---|---|---|
| CON | L-Tau | M-Tau | H-Tau | Linear | Quadratic | Cubic | ||
| Jejunum | ||||||||
| Weight (g) | 277.01 | 309.64 | 279.62 | 335.56 | 53.33 | 0.245 | 0.670 | 0.237 |
| Weight index (g/kg) | 26.57 | 28.17 | 25.17 | 26.62 | 6.11 | 0.838 | 0.981 | 0.521 |
| Length (cm) | 744.50 | 785.00 | 787.50 | 857.50 | 132.48 | 0.271 | 0.828 | 0.728 |
| Length index (cm/kg) | 71.22 | 70.57 | 71.05 | 68.09 | 14.27 | 0.785 | 0.874 | 0.888 |
| VH (μm) | 235.95 c | 286.75 bc | 320.24 ab | 368.00 a | 47.55 | 0.003 | 0.952 | 0.774 |
| CD (μm) | 82.11 | 74.37 | 73.00 | 71.80 | 18.38 | 0.447 | 0.728 | 0.883 |
| VH/CD | 3.46 b | 3.98 b | 4.65 ab | 6.17 a | 1.19 | 0.011 | 0.432 | 0.800 |
| Ileum | ||||||||
| Weight (g) | 117.5 | 155.11 | 149.28 | 127.01 | 45.45 | 0.839 | 0.232 | 0.797 |
| Weight index (g/kg) | 11.03 | 13.98 | 12.87 | 10.03 | 3.67 | 0.649 | 0.159 | 0.783 |
| Length (cm) | 238.25 | 366.25 | 373.00 | 287.50 | 104.61 | 0.521 | 0.064 | 0.903 |
| Length index (cm/kg) | 22.79 | 32.77 | 32.49 | 22.68 | 8.34 | 0.974 | 0.035 | 0.970 |
| VH (μm) | 286.69 b | 301.40 b | 335.97 ab | 384.53 a | 46.13 | 0.019 | 0.512 | 0.957 |
| CD (μm) | 106.96 | 91.19 | 80.22 | 73.04 | 25.13 | 0.068 | 0.739 | 0.986 |
| VH/CD | 2.92 b | 3.56 b | 4.61 ab | 5.81 a | 1.13 | 0.004 | 0.650 | 0.915 |
| Items | Treatments | SEM | p Value | |||||
|---|---|---|---|---|---|---|---|---|
| CON | L-Tau | M-Tau | H-Tau | Linear | Quadratic | Cubic | ||
| Serum | ||||||||
| D-LA (μmol/L) | 122.51 a | 107.10 a | 94.74 ab | 76.32 b | 17.98 | 0.003 | 0.870 | 0.825 |
| Jejunal mucosa | ||||||||
| CLDN-1 (ng/g) | 586.88 c | 632.24 c | 1017.64 b | 1469.29 a | 126.29 | <.001 | 0.007 | 0.351 |
| ZO-1 (ng/g) | 756.72 b | 714.90 b | 879.10 b | 1291.94 a | 133.31 | <0.001 | 0.007 | 0.890 |
| OCLN (ng/g) | 39.78 c | 32.09 c | 66.02 b | 136.85 a | 9.54 | <0.001 | <0.001 | 0.841 |
| Items | Treatments | SEM | p Value | |||||
|---|---|---|---|---|---|---|---|---|
| CON | L-Tau | M-Tau | H-Tau | Linear | Quadratic | Cubic | ||
| Alpha diversity in | ||||||||
| Observed_otus | 594.00 | 632.25 | 610.00 | 554.50 | 106.52 | 0.566 | 0.396 | 0.911 |
| Shannon | 7.19 | 6.73 | 6.30 | 6.35 | 0.77 | 0.111 | 0.520 | 0.788 |
| Simpson | 0.97 | 0.95 | 0.92 | 0.93 | 0.04 | 0.202 | 0.507 | 0.528 |
| Chao1 | 595.23 | 641.05 | 618.02 | 555.85 | 110.07 | 0.577 | 0.346 | 0.906 |
| Pielou_e | 0.78 | 0.72 | 0.68 | 0.70 | 0.07 | 0.084 | 0.319 | 0.775 |
| Goods_coverage | 1.00 | 1.00 | 1.00 | 1.00 | 0.00 | 0.809 | 0.032 | 0.872 |
| Phylum level in | ||||||||
| Firmicutes | 75.33 b | 82.06 a | 85.60 a | 87.17 a | 4.17 | 0.001 | 0.240 | 0.898 |
| Proteobacteria | 10.48 a | 5.88 ab | 4.25 b | 2.84 b | 3.23 | 0.005 | 0.342 | 0.710 |
| Bacteroidetes | 11.49 | 8.94 | 8.03 | 7.94 | 4.75 | 0.297 | 0.614 | 0.939 |
| Actinobacteriota | 0.94 | 2.23 | 1.06 | 1.26 | 0.82 | 0.914 | 0.216 | 0.060 |
| Spirochaetota | 0.95 a | 0.18 b | 0.22 b | 0.11 b | 0.33 | 0.006 | 0.07 | 0.233 |
| Verrucomicrobiota | 0.23 | 0.41 | 0.21 | 0.34 | 0.38 | 0.899 | 0.891 | 0.418 |
| Cyanobacteria | 0.09 | 0.11 | 0.52 | 0.16 | 0.28 | 0.340 | 0.191 | 0.085 |
| Generic level | ||||||||
| Clostridium_sensu _stricto_1 | 5.25 b | 24.10 a | 37.08 a | 34.71 a | 11.55 | 0.002 | 0.091 | 0.721 |
| Streptococcus | 13.49 | 5.92 | 2.72 | 5.42 | 6.89 | 0.101 | 0.162 | 0.922 |
| Lachnospiraceae | 6.08 | 5.96 | 4.56 | 2.65 | 4.48 | 0.266 | 0.696 | 0.938 |
| Terrisporobacter | 0.56 b | 4.19 a | 4.74 a | 3.12 ab | 1.93 | 0.081 | 0.019 | 0.836 |
| UCG-005 | 5.40 a | 1.47 b | 2.39 b | 2.29 b | 1.32 | 0.015 | 0.014 | 0.072 |
| T34_unclassified | 4.93 | 2.38 | 2.25 | 1.08 | 2.42 | 0.053 | 0.579 | 0.533 |
| Agathobacter | 3.33 | 2.53 | 2.44 | 1.62 | 1.69 | 0.190 | 0.990 | 0.707 |
| Turicibacter | 0.42 b | 1.93 ab | 3.59 a | 3.39 a | 1.45 | 0.007 | 0.263 | 0.543 |
| Ruminococcus | 3.13 | 1.37 | 1.75 | 1.87 | 1.22 | 0.234 | 0.146 | 0.395 |
| Clostridium_sensu_stricto_2 | 2.15 | 1.57 | 1.81 | 2.51 | 1.61 | 0.716 | 0.440 | 0.918 |
| Streptococcus | 0.53 | 2.52 | 1.74 | 1.78 | 1.45 | 0.380 | 0.203 | 0.292 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Jiang, L.; Chu, Y.; Feng, F.; Tang, W.; Chen, C.; Qiu, Y.; Hu, Z.; Diao, H.; Tang, Z. Dietary Taurine Improves Growth Performance and Intestine Health via the GSH/GSSG Antioxidant System and Nrf2/ARE Signaling Pathway in Weaned Piglets. Antioxidants 2023, 12, 1852. https://doi.org/10.3390/antiox12101852
Wang L, Jiang L, Chu Y, Feng F, Tang W, Chen C, Qiu Y, Hu Z, Diao H, Tang Z. Dietary Taurine Improves Growth Performance and Intestine Health via the GSH/GSSG Antioxidant System and Nrf2/ARE Signaling Pathway in Weaned Piglets. Antioxidants. 2023; 12(10):1852. https://doi.org/10.3390/antiox12101852
Chicago/Turabian StyleWang, Lingang, Liwen Jiang, Yunyun Chu, Fu Feng, Wenjie Tang, Chen Chen, Yibin Qiu, Zhijin Hu, Hui Diao, and Zhiru Tang. 2023. "Dietary Taurine Improves Growth Performance and Intestine Health via the GSH/GSSG Antioxidant System and Nrf2/ARE Signaling Pathway in Weaned Piglets" Antioxidants 12, no. 10: 1852. https://doi.org/10.3390/antiox12101852