
Co-Treatments of Gardeniae Fructus and Silymarin Ameliorates Excessive Oxidative Stress-Driven Liver Fibrosis by Regulation of Hepatic Sirtuin1 Activities Using Thioacetamide-Induced Mice Model
 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Preparation of the Plant Material and Finger Printing Analysis of GF
2.2. Mice Treatment
2.3. Histological Examination
2.4. Analysis of Serum Biochemistry
2.5. Cell Death Signaling
2.6. Analysis of Immunohistochemistry (IHC)
2.7. Measurement of ROS, TBA-Reactive Substance (TBARS) Levels and Pro-Inflammatory Cytokine
2.8. Cell Cultures
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
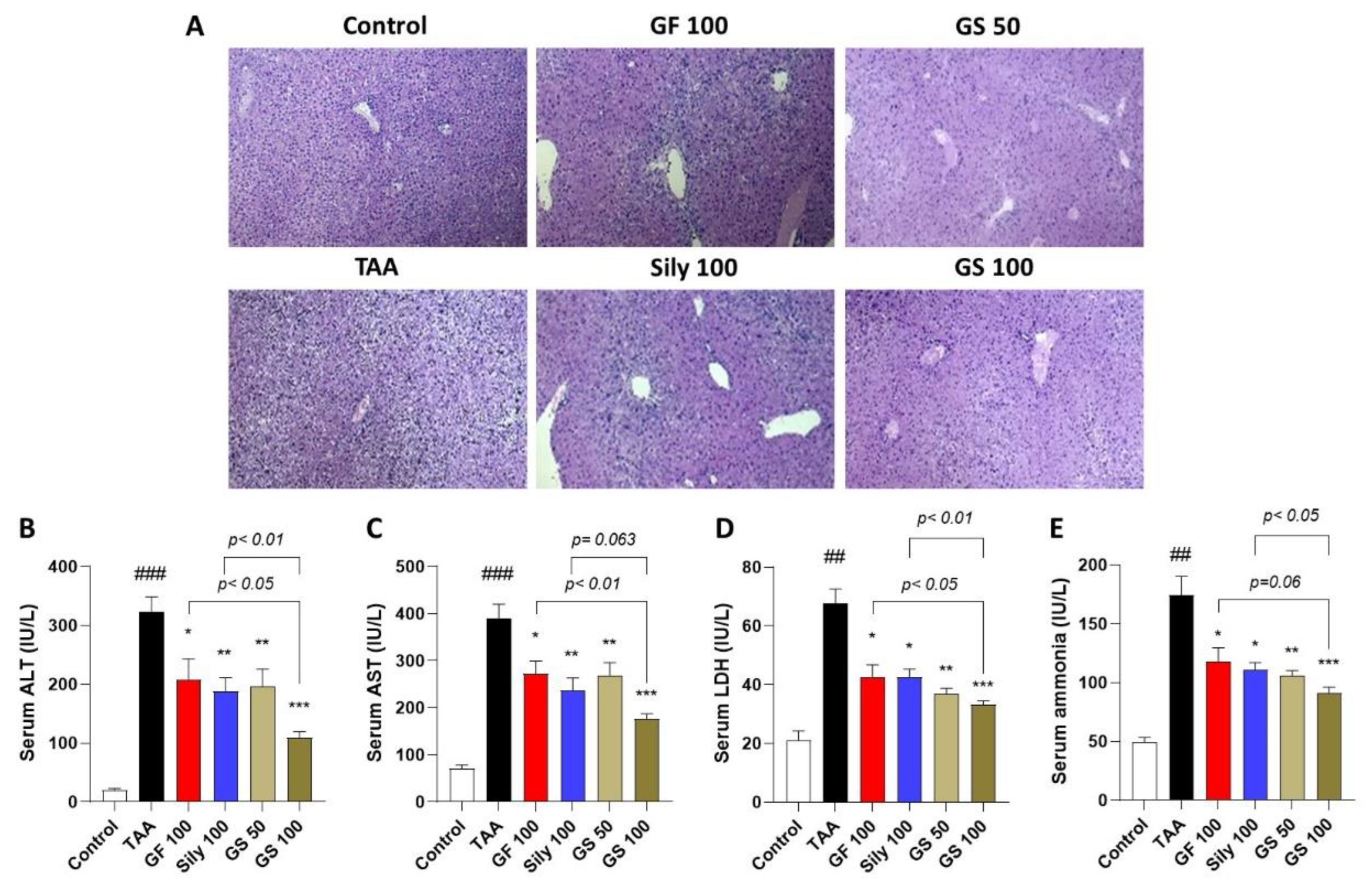
3.1. Co-Treatments of GF and Silymarin Ameliorate TAA-Injected Hepatic Injury
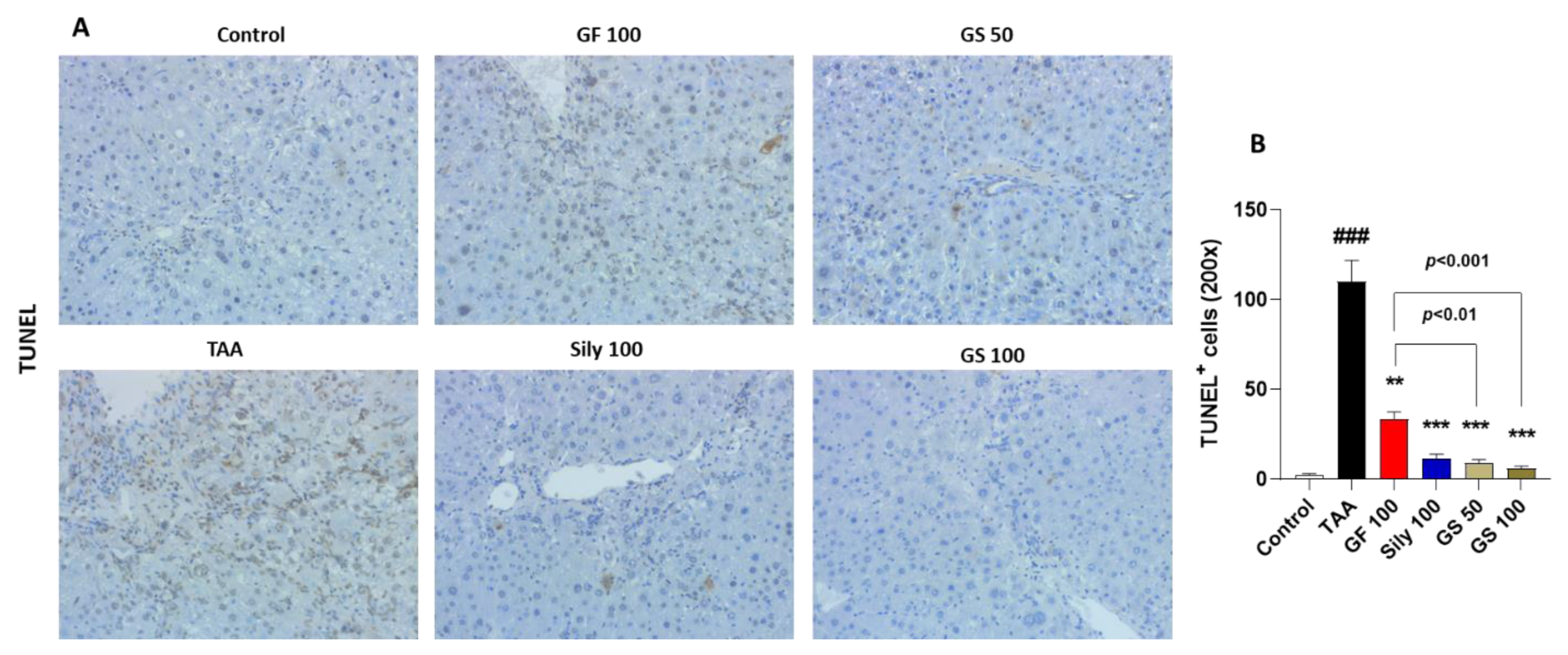
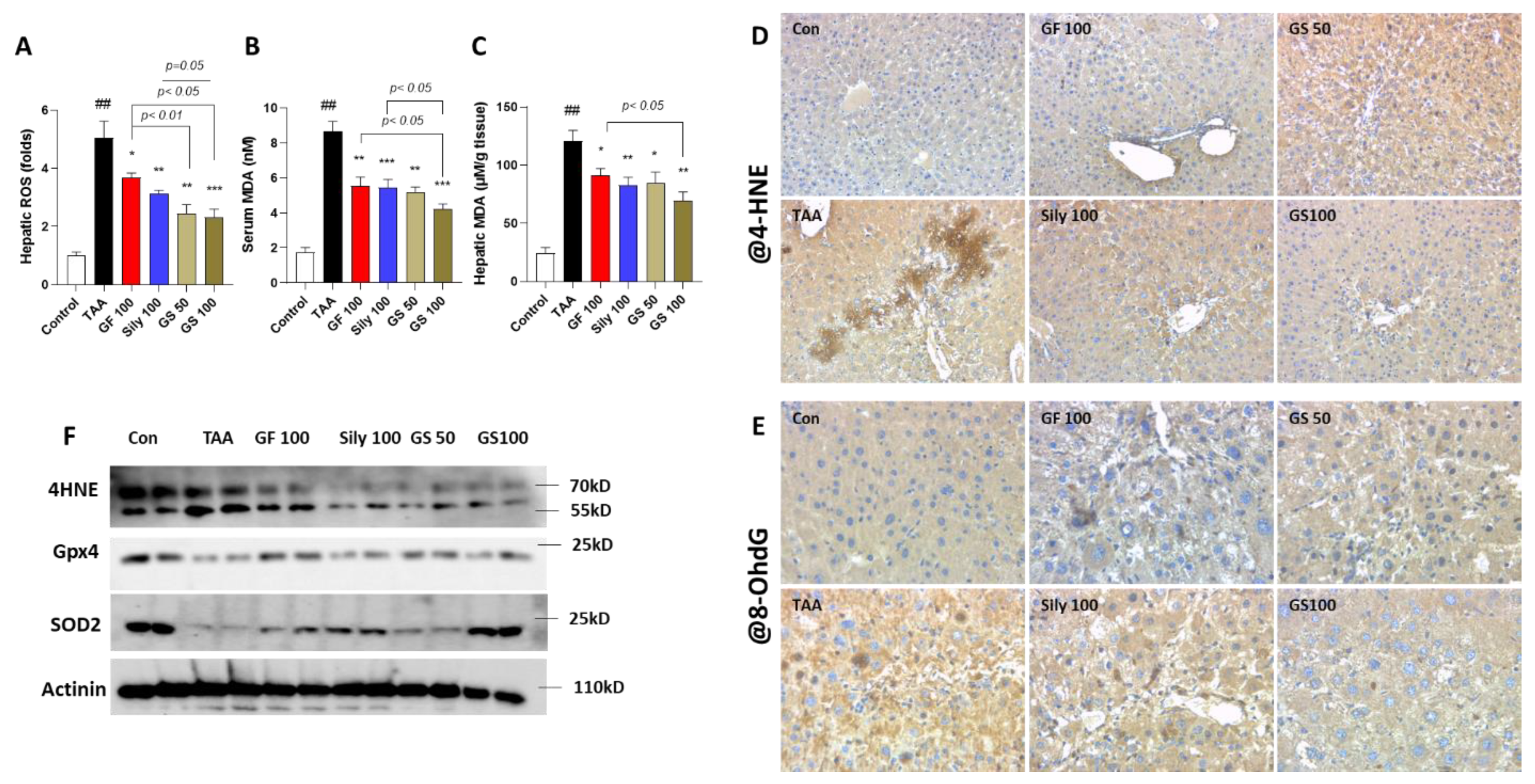
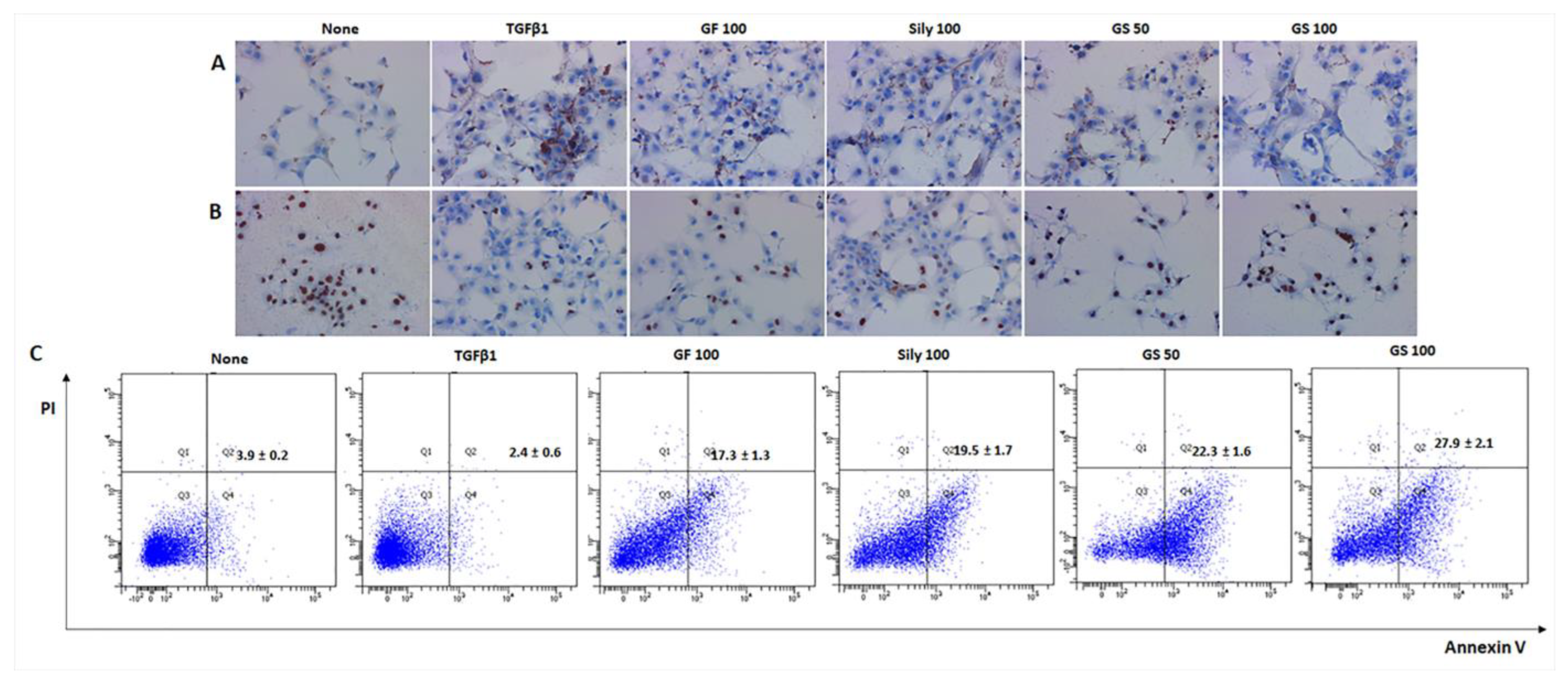
3.2. Synergistic Effects of GS on the Liver Cell Deaths by Attenuations of Oxidative Stress
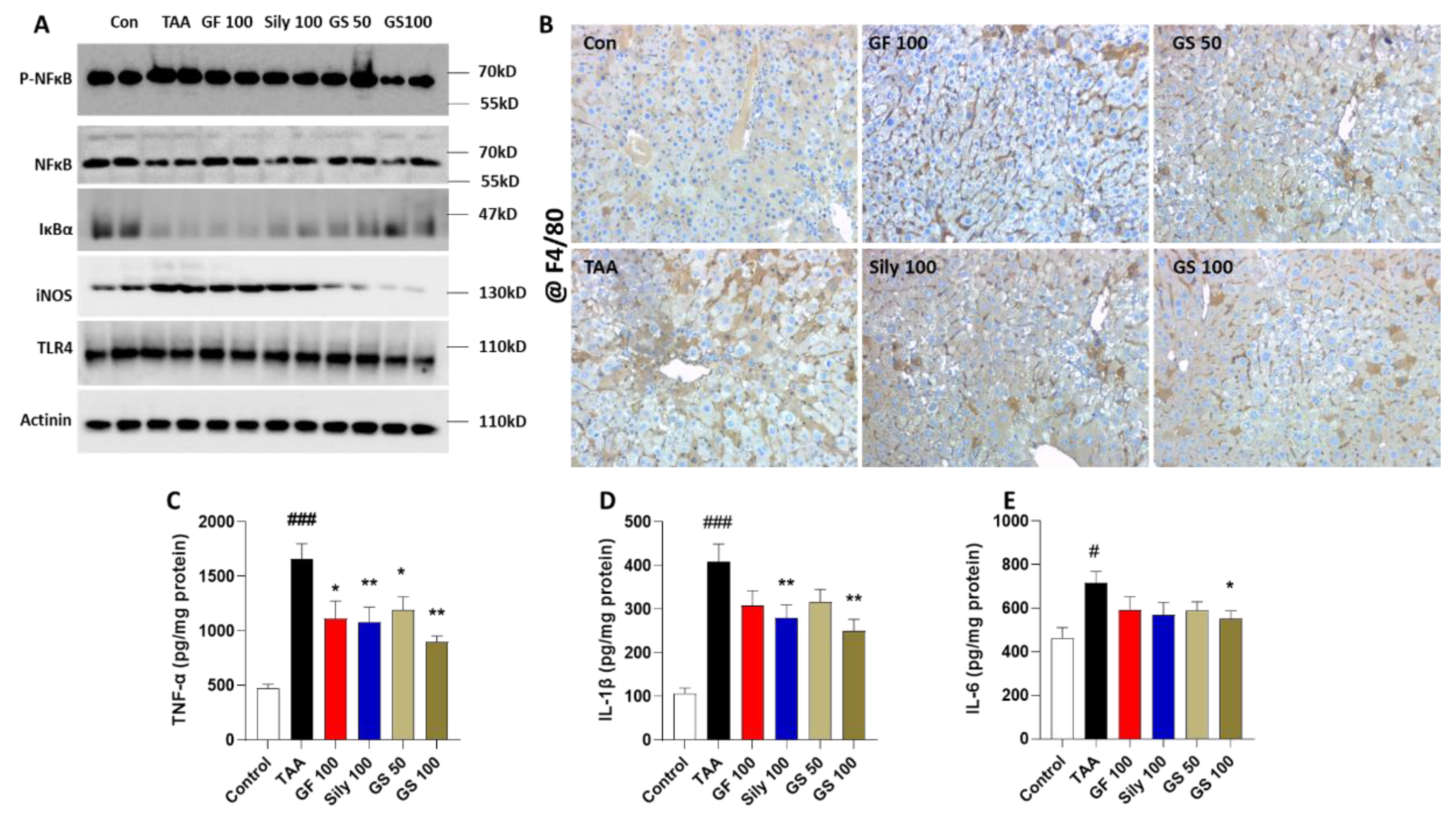
3.3. Anti-Inflammatory Effects of GS in the Liver Tissue
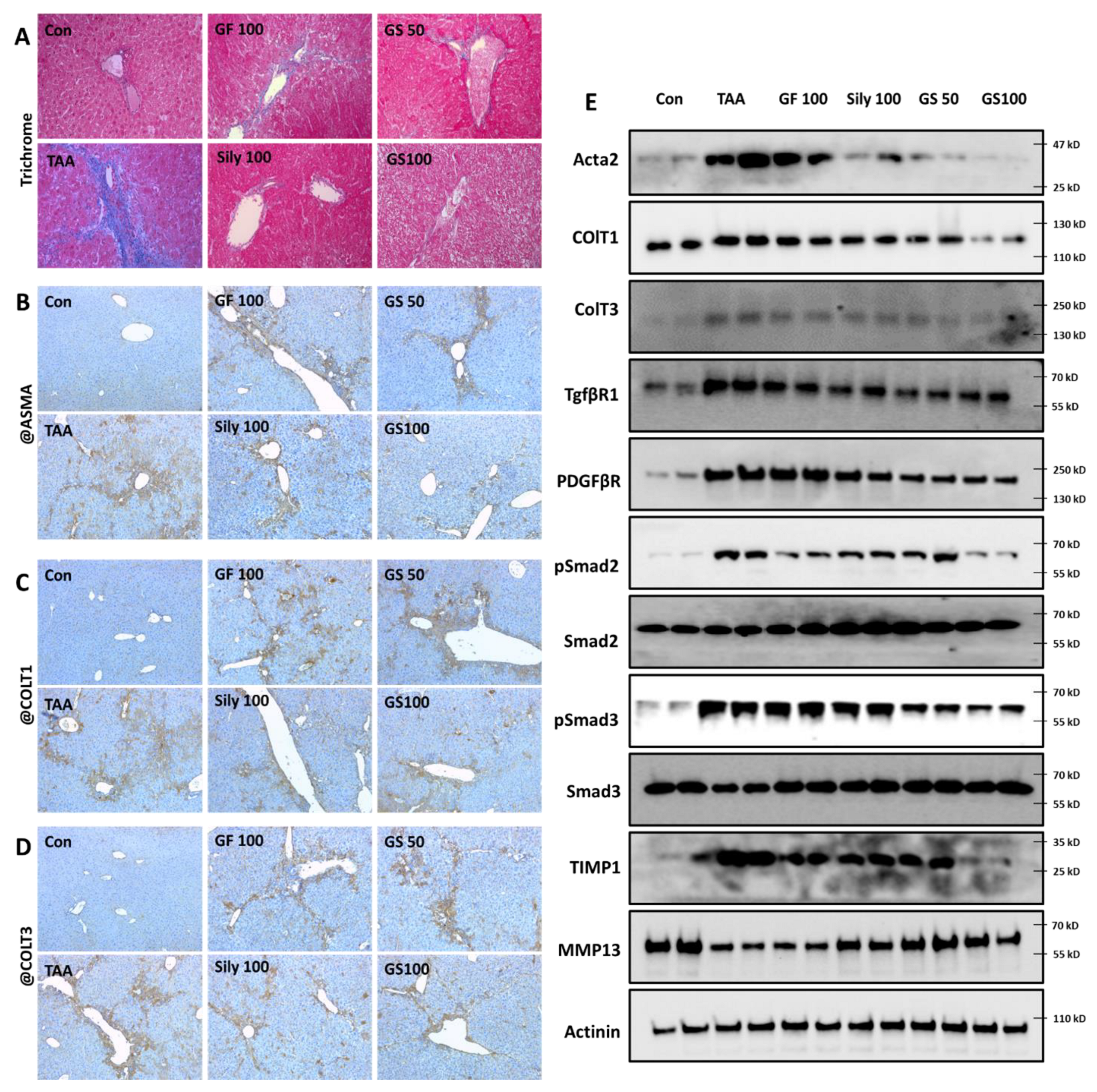
3.4. Anti-Liver Fibrosis Effects of GS on the Liver Tissue with Synergistic Manner
3.5. Pharmacological Actions of GS through Nrf2/HO-1 Signaling Pathway via Regulations of Hepatic Sirt1 Activation
3.6. Selective Effects of Sirt6 by Induction of HSCs during Fibrosis Development
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wong, R.J.; Kachru, N.; Martinez, D.J.; Moynihan, M.; Ozbay, A.B.; Gordon, S.C. Real-world Comorbidity Burden, Health Care Utilization, and Costs of Nonalcoholic Steatohepatitis Patients With Advanced Liver Diseases. J. Clin. Gastroenterol. 2021, 55, 891–902. [Google Scholar] [CrossRef]
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of liver diseases in the world. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Zhai, M.; Long, J.; Gong, Y.; Ren, C.; Zhang, D.; Lin, X.; Liu, S. The burden of liver cirrhosis in mortality: Results from the global burden of disease study. Front. Public Health 2022, 10, 909455. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Kisseleva, T. Reversibility of liver fibrosis. Clin. Res. Hepatol. Gastroenterol. 2015, 39 (Suppl S1), S60–S63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismail, M.H.; Pinzani, M. Reversal of liver fibrosis. Saudi J. Gastroenterol. 2009, 15, 72–79. [Google Scholar] [CrossRef]
- Garrido, A.; Djouder, N. Cirrhosis: A Questioned Risk Factor for Hepatocellular Carcinoma. Trends Cancer 2021, 7, 29–36. [Google Scholar] [CrossRef]
- Caligiuri, A.; Gentilini, A.; Pastore, M.; Gitto, S.; Marra, F. Cellular and Molecular Mechanisms Underlying Liver Fibrosis Regression. Cells 2021, 10, 2759. [Google Scholar] [CrossRef]
- Kisseleva, T.; Brenner, D. Molecular and cellular mechanisms of liver fibrosis and its regression. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 151–166. [Google Scholar] [CrossRef]
- Wu, J.; Zern, M.A. Hepatic stellate cells: A target for the treatment of liver fibrosis. J. Gastroenterol. 2000, 35, 665–672. [Google Scholar] [CrossRef]
- Li, Y.; Pu, S.; Liu, Q.; Li, R.; Zhang, J.; Wu, T.; Chen, L.; Li, H.; Yang, X.; Zou, M.; et al. An integrin-based nanoparticle that targets activated hepatic stellate cells and alleviates liver fibrosis. J. Control. Release 2019, 303, 77–90. [Google Scholar] [CrossRef]
- Latief, U.; Ahmad, R. Herbal remedies for liver fibrosis: A review on the mode of action of fifty herbs. J. Tradit. Complement. Med. 2018, 8, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.R.; Chen, X.P.; Lu, J.J.; Wang, Y.; Wang, Y.T. Potent natural products and herbal medicines for treating liver fibrosis. Chin. Med. 2015, 10, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joung, J.Y.; Kim, H.G.; Lee, J.S.; Cho, J.H.; Ahn, Y.C.; Lee, D.S.; Son, C.G. Anti-hepatofibrotic effects of CGX, a standardized herbal formula: A multicenter randomized clinical trial. Biomed. Pharmacother. 2020, 126, 110105. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.S.; Chae, Y.S.; Kim, D.Y.; Seo, S.W.; Park, H.J.; Bae, G.S.; Kim, T.H.; Oh, H.J.; Yun, K.J.; Park, R.K.; et al. Gardenia jasminoides protects against cerulein-induced acute pancreatitis. World J. Gastroenterol. 2008, 14, 6188–6194. [Google Scholar] [CrossRef]
- Li, C.; Gao, X.; Gao, X.; Lv, J.; Bian, X.; Lv, J.; Sun, J.; Luo, G.; Zhang, H. Effects of medicine food Fructus Gardeniae on liver and kidney functions after oral administration to rats for 12 weeks. J. Food Biochem. 2021, 45, e13752. [Google Scholar] [CrossRef]
- Tang, Z.; Li, L.; Xia, Z. Exploring Anti-Nonalcoholic Fatty Liver Disease Mechanism of Gardeniae Fructus by Network Pharmacology, Molecular Docking, and Experiment Validation. ACS Omega 2022, 7, 25521–25531. [Google Scholar] [CrossRef]
- Ma, Z.G.; Kong, C.Y.; Song, P.; Zhang, X.; Yuan, Y.P.; Tang, Q.Z. Geniposide Protects against Obesity-Related Cardiac Injury through AMPKalpha- and Sirt1-Dependent Mechanisms. Oxid. Med. Cell. Longev. 2018, 2018, 6053727. [Google Scholar] [CrossRef] [Green Version]
- He, T.; Shen, H.; Zhu, J.; Zhu, Y.; He, Y.; Li, Z.; Lu, H. Geniposide attenuates cadmium-induced oxidative stress injury via Nrf2 signaling in osteoblasts. Mol. Med. Rep. 2019, 20, 1499–1508. [Google Scholar] [CrossRef] [Green Version]
- Gillessen, A.; Schmidt, H.H. Silymarin as Supportive Treatment in Liver Diseases: A Narrative Review. Adv. Ther. 2020, 37, 1279–1301. [Google Scholar] [CrossRef] [Green Version]
- Golestaneh, E.; Aslani, A.; Aghaei, M.; Hashemnia, M.; Aarabi, M.H. Preparation and characterisation of a new form of silymarin as a potential antidiabetic agent in the adult male rat. Arch. Physiol. Biochem. 2021, 1–11. [Google Scholar] [CrossRef]
- Di Costanzo, A.; Angelico, R. Formulation Strategies for Enhancing the Bioavailability of Silymarin: The State of the Art. Molecules 2019, 24, 2155. [Google Scholar] [CrossRef] [Green Version]
- Yousaf, A.M.; Malik, U.R.; Shahzad, Y.; Mahmood, T.; Hussain, T. Silymarin-laden PVP-PEG polymeric composite for enhanced aqueous solubility and dissolution rate: Preparation and in vitro characterization. J. Pharm. Anal. 2019, 9, 34–39. [Google Scholar] [CrossRef]
- Vaziri, H.; Dessain, S.K.; Ng Eaton, E.; Imai, S.I.; Frye, R.A.; Pandita, T.K.; Guarente, L.; Weinberg, R.A. hSIR2(SIRT1) functions as an NAD-dependent p53 deacetylase. Cell 2001, 107, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Shin, M.R.; Lee, J.A.; Kim, M.; Lee, S.; Oh, M.; Moon, J.; Nam, J.W.; Choi, H.; Mun, Y.J.; Roh, S.S. Gardeniae Fructus Attenuates Thioacetamide-Induced Liver Fibrosis in Mice via Both AMPK/SIRT1/NF-kappaB Pathway and Nrf2 Signaling. Antioxidants 2021, 10, 1837. [Google Scholar] [CrossRef]
- Sun, W.; He, S.; Martínez-Romero, C.; Kouznetsova, J.; Tawa, G.; Xu, M.; Shinn, P.; Fisher, E.; Long, Y.; Motabar, O.; et al. Synergistic drug combination effectively blocks Ebola virus infection. Antivir. Res. 2017, 137, 165–172. [Google Scholar] [CrossRef]
- Yue, W.; Yang, C.S.; DiPaola, R.S.; Tan, X.L. Repurposing of metformin and aspirin by targeting AMPK-mTOR and inflammation for pancreatic cancer prevention and treatment. Cancer Prev. Res. 2014, 7, 388–397. [Google Scholar] [CrossRef] [Green Version]
- Ledda-Columbano, G.M.; Coni, P.; Curto, M.; Giacomini, L.; Faa, G.; Oliverio, S.; Piacentini, M.; Columbano, A. Induction of two different modes of cell death, apoptosis and necrosis, in rat liver after a single dose of thioacetamide. Am. J. Pathol. 1991, 139, 1099–1109. [Google Scholar]
- Lim, S.; Lee, S.J.; Nam, K.W.; Kim, K.H.; Mar, W. Hepatoprotective effects of reynosin against thioacetamide-induced apoptosis in primary hepatocytes and mouse liver. Arch. Pharm. Res. 2013, 36, 485–494. [Google Scholar] [CrossRef]
- Đurašević, S.; Pejić, S.; Grigorov, I.; Nikolić, G.; Mitić-Ćulafić, D.; Dragićević, M.; Đorđević, J.; Vukotić, N.T.; Đorđević, N.; Todorović, A.; et al. Effects of C60 Fullerene on Thioacetamide-Induced Rat Liver Toxicity and Gut Microbiome Changes. Antioxidants 2021, 10, 911. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.M.; Subeq, Y.M.; Lee, R.P.; Chiou, T.W.; Hsu, B.G. Single dose intravenous thioacetamide administration as a model of acute liver damage in rats. Int. J. Exp. Pathol. 2008, 89, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Chilakapati, J.; Shankar, K.; Korrapati, M.C.; Hill, R.A.; Mehendale, H.M. Saturation toxicokinetics of thioacetamide: Role in initiation of liver injury. Drug Metab. Dispos. 2005, 33, 1877–1885. [Google Scholar] [CrossRef] [PubMed]
- Bruck, R.; Aeed, H.; Schey, R.; Matas, Z.; Reifen, R.; Zaiger, G.; Hochman, A.; Avni, Y. Pyrrolidine dithiocarbamate protects against thioacetamide-induced fulminant hepatic failure in rats. J. Hepatol. 2002, 36, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Diez-Fernandez, C.; Sanz, N.; Bosca, L.; Hortelano, S.; Cascales, M. Involvement of nitric oxide synthesis in hepatic perturbations induced in rats by a necrogenic dose of thioacetamide. Br. J. Pharmacol. 1997, 121, 820–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenner, D.A. Molecular pathogenesis of liver fibrosis. Trans. Am. Clin. Climatol. Assoc. 2009, 120, 361–368. [Google Scholar] [PubMed]
- Lee, U.E.; Friedman, S.L. Mechanisms of hepatic fibrogenesis. Best Pract. Res. Clin. Gastroenterol. 2011, 25, 195–206. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, Y.; Sun, B. The Molecular Mechanisms of Liver Fibrosis and Its Potential Therapy in Application. Int. J. Mol. Sci. 2022, 23, 2572. [Google Scholar] [CrossRef]
- Bao, Y.L.; Wangm, L.; Pan, H.T.; Zhang, T.R.; Chen, Y.H.; Xu, S.J.; Mao, X.L.; Li, S.W. Animal and Organoid Models of Liver Fibrosis. Front. Physiol. 2021, 12, 666138. [Google Scholar] [CrossRef]
- Tan, Z.; Sun, H.; Xue, T.; Gan, C.; Liu, H.; Xie, Y.; Yao, Y.; Ye, T. Liver Fibrosis: Therapeutic Targets and Advances in Drug Therapy. Front. Cell Dev. Biol. 2021, 9, 730176. [Google Scholar] [CrossRef]
- Ong, A.L.C.; Ramasamy, T.S. Role of Sirtuin1-p53 regulatory axis in aging, cancer and cellular reprogramming. Ageing Res. Rev. 2018, 43, 64–80. [Google Scholar] [CrossRef]
- Alves-Fernandes, D.K.; Jasiulionis, M.G. The Role of SIRT1 on DNA Damage Response and Epigenetic Alterations in Cancer. Int. J. Mol. Sci. 2019, 20, 3153. [Google Scholar] [CrossRef] [Green Version]
- Ren, R.; Wang, Z.; Wu, M.; Wang, H. Emerging Roles of SIRT1 in Alcoholic Liver Disease. Int. J. Biol. Sci. 2020, 16, 3174–3183. [Google Scholar] [CrossRef] [PubMed]
- de Gregorio, E.; Colell, A.; Morales, A.; Mari, M. Relevance of SIRT1-NF-kappaB Axis as Therapeutic Target to Ameliorate Inflammation in Liver Disease. Int. J. Mol. Sci. 2020, 21, 3858. [Google Scholar] [CrossRef] [PubMed]
- Farghali, H.; Kemelo, M.K.; Canova, N.K. SIRT1 Modulators in Experimentally Induced Liver Injury. Oxid. Med. Cell. Longev. 2019, 2019, 8765954. [Google Scholar] [CrossRef]
- Blokker, B.A.; Maijo, M.; Echeandia, M.; Galduroz, M.; Patterson, A.M.; Ten, A.; Philo, M.; Schungel, R.; Gutierrez-de Juan, V.; Halilbasic, E.; et al. Fine-Tuning of Sirtuin 1 Expression Is Essential to Protect the Liver From Cholestatic Liver Disease. Hepatology 2019, 69, 699–716. [Google Scholar] [CrossRef] [Green Version]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Crosstalk between Oxidative Stress and SIRT1: Impact on the Aging Process. Int. J. Mol. Sci. 2013, 14, 3834–3859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, T.; Huang, J.; Nisar, M.F.; Wan, C.; Huang, W. The Beneficial Roles of SIRT1 in Drug-Induced Liver Injury. Oxid. Med. Cell. Longev. 2019, 2019, 8506195. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, Y.; Wang, Y.; Chao, Y.; Zhang, J.; Jia, Y.; Tie, J.; Hu, D. Regulation of SIRT1 and Its Roles in Inflammation. Front. Immunol. 2022, 13, 831168. [Google Scholar] [CrossRef]
- Qiang, L.; Sample, A.; Liu, H.; Wu, X.; He, Y.Y. Epidermal SIRT1 regulates inflammation, cell migration, and wound healing. Sci. Rep. 2017, 7, 14110. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Hong, W.; Hao, C.; Li, L.; Wu, D.; Shen, A.; Lu, J.; Zheng, Y.; Li, P.; Xu, Y. SIRT1 antagonizes liver fibrosis by blocking hepatic stellate cell activation in mice. FASEB J. 2018, 32, 500–511. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Hong, W.; Hao, C.; Li, L.; Xu, H.; Li, P.; Xu, Y. Hepatic stellate cell-specific deletion of SIRT1 exacerbates liver fibrosis in mice. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 3202–3211. [Google Scholar] [CrossRef]
- Yi, J.; Wu, S.; Tan, S.; Qin, Y.; Wang, X.; Jiang, J.; Liu, H.; Wu, B. Berberine alleviates liver fibrosis through inducing ferrous redox to activate ROS-mediated hepatic stellate cells ferroptosis. Cell Death Discov. 2021, 7, 374. [Google Scholar] [CrossRef] [PubMed]
- Dognin, J.; Crichton, R.R. Mobilisation of iron from ferritin fractions of defined iron content by biological reductants. FEBS Lett. 1975, 54, 234–236. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.A.; Shin, M.-R.; Choi, J.; Kim, M.; Park, H.-J.; Roh, S.-S. Co-Treatments of Gardeniae Fructus and Silymarin Ameliorates Excessive Oxidative Stress-Driven Liver Fibrosis by Regulation of Hepatic Sirtuin1 Activities Using Thioacetamide-Induced Mice Model. Antioxidants 2023, 12, 97. https://doi.org/10.3390/antiox12010097
Lee JA, Shin M-R, Choi J, Kim M, Park H-J, Roh S-S. Co-Treatments of Gardeniae Fructus and Silymarin Ameliorates Excessive Oxidative Stress-Driven Liver Fibrosis by Regulation of Hepatic Sirtuin1 Activities Using Thioacetamide-Induced Mice Model. Antioxidants. 2023; 12(1):97. https://doi.org/10.3390/antiox12010097
Chicago/Turabian StyleLee, Jin A, Mi-Rae Shin, JeongWon Choi, MinJu Kim, Hae-Jin Park, and Seong-Soo Roh. 2023. "Co-Treatments of Gardeniae Fructus and Silymarin Ameliorates Excessive Oxidative Stress-Driven Liver Fibrosis by Regulation of Hepatic Sirtuin1 Activities Using Thioacetamide-Induced Mice Model" Antioxidants 12, no. 1: 97. https://doi.org/10.3390/antiox12010097