
Potential Antioxidative Activity of Homocysteine in Erythrocytes under Oxidative Stress

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Blood Sampling
2.3. Determination of Plasma Total-Hcy after Whole Blood Kept at Room Temperature for Different Durations
2.4. The Production of Homocysteine from Blood Cells
2.5. The Production of Total-Hcy from Erythrocytes after Incubation with Methionine
2.6. The Effects of r-Hcy and Methionine on Reactive Oxygen Species (ROS) Production in Erythrocytes
2.7. Measurement of Erythrocyte Osmotic Fragility
2.8. The Effects of r-Hcy or Methionine on Ferryl Hb Decay
2.9. Measurement of Methemoglobin (MetHb) in Erythrocytes
2.10. The Contents of Total-Hcy and Glutathione (GSH) in Rosup-Treated Erythrocytes
2.11. HPLC-MS Determination of Total-Hcy and r-Hcy Concentrations
2.12. HPLC Determination of GSH
2.13. Statistical Analysis
3. Results
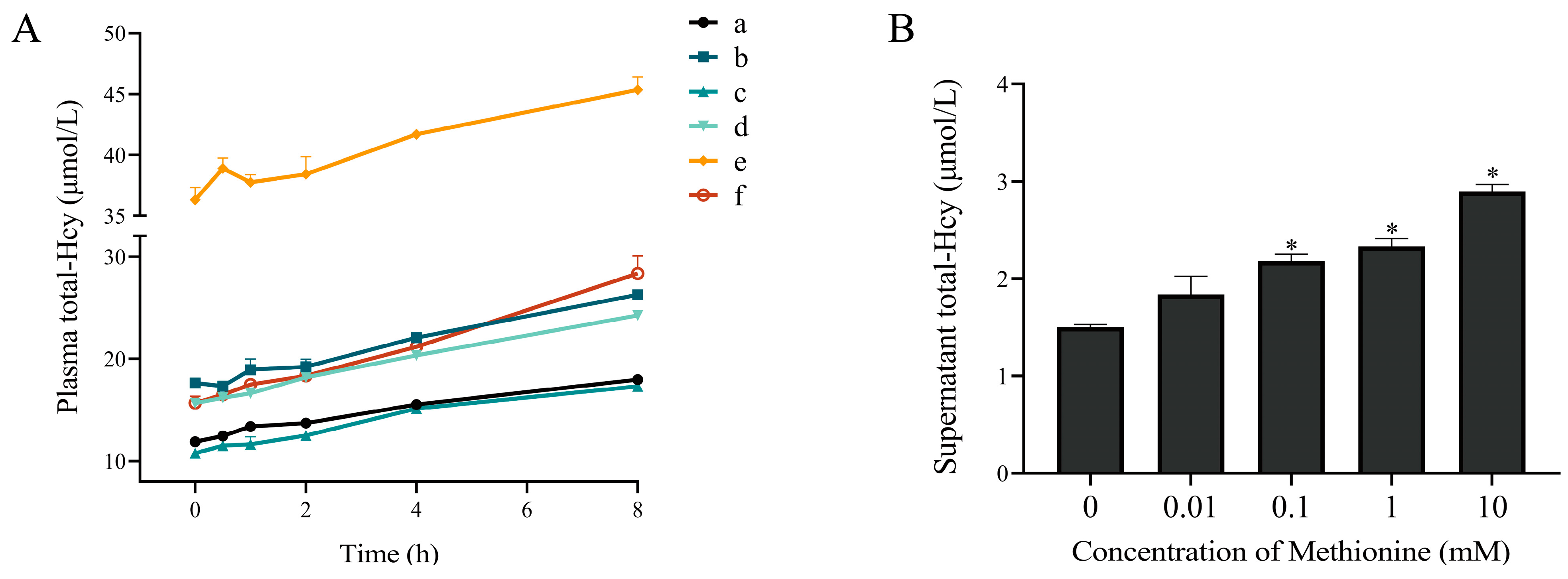
3.1. The Plasma Total-Hcy Increased after Whole Blood Samples Kept at Room Temperature for Different Durations
3.2. The Production of Homocysteine from Erythrocytes and Leukocytes
3.3. Incubation of Erythrocytes with Methionine Increased Total-Hcy Production
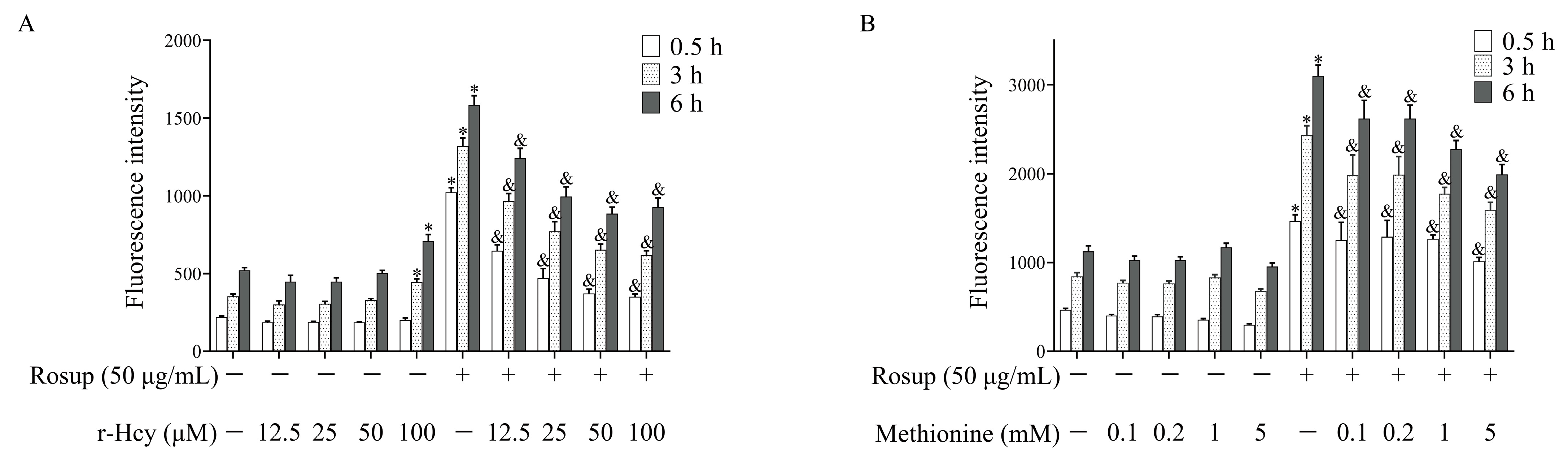
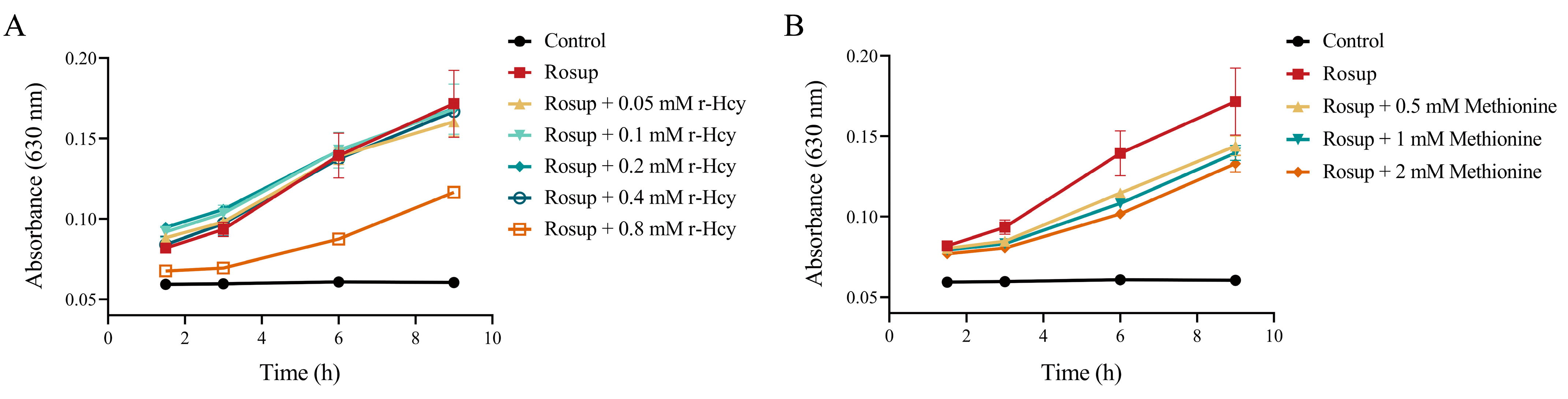
3.4. r-Hcy and Methionine Inhibited the Production of Erythrocytes ROS Induced by Rosup
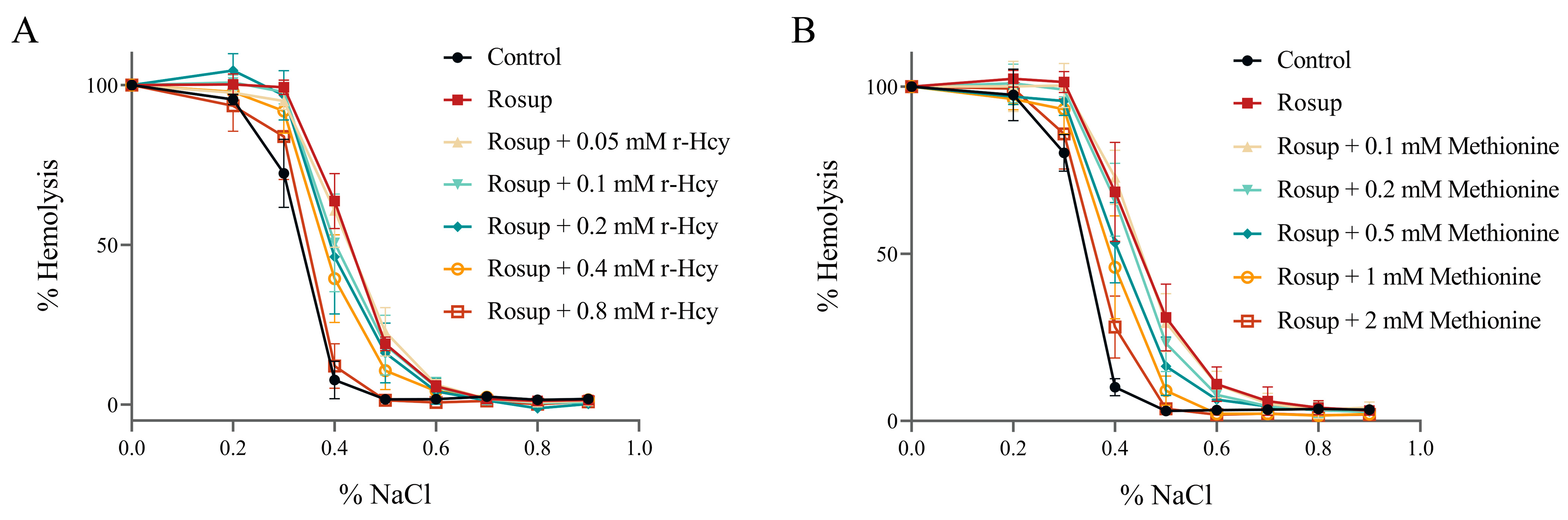
3.5. The Effects of r-Hcy and Methionine on Erythrocytes Osmotic Fragility under Oxidative Stress
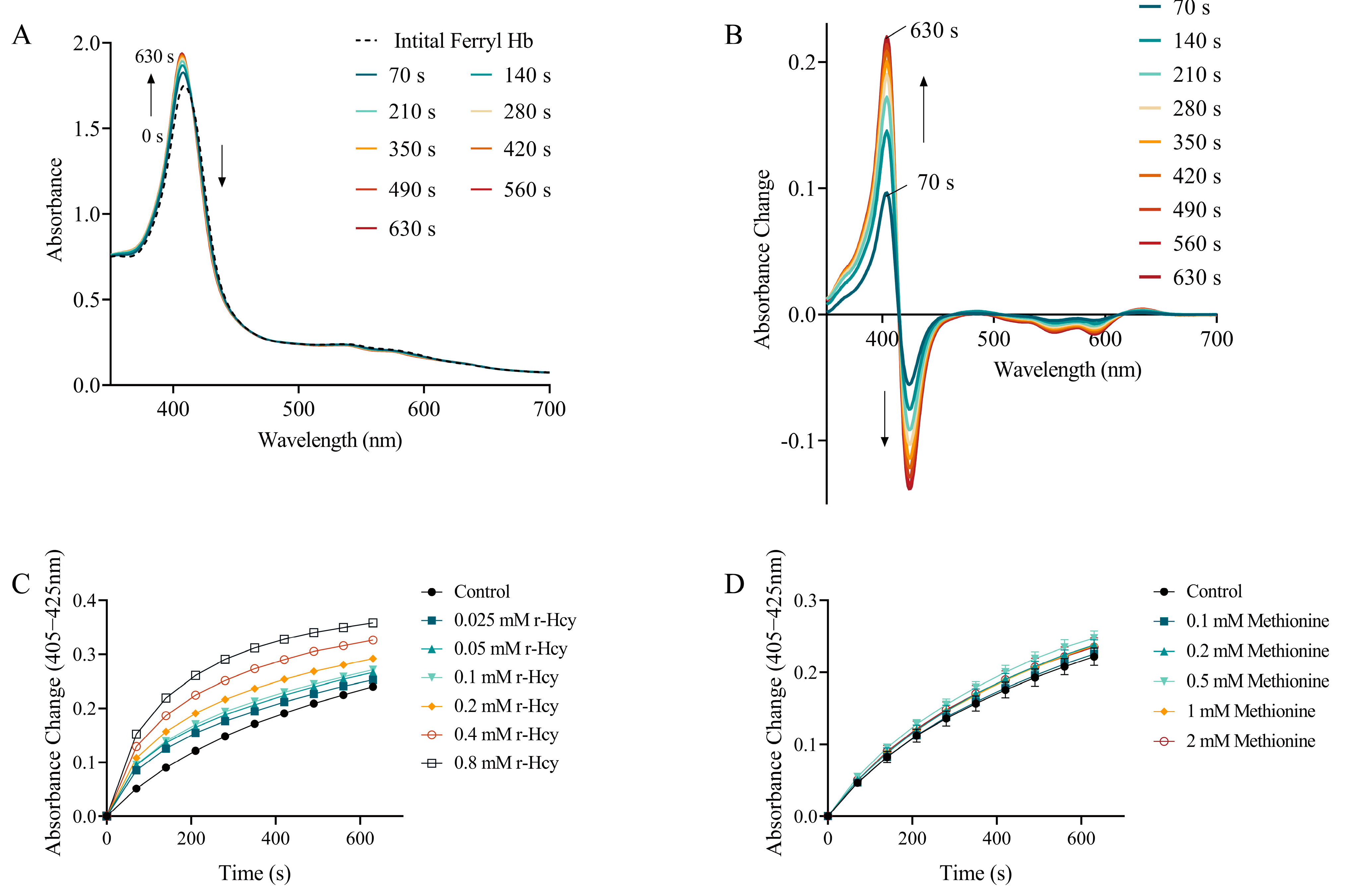
3.6. The Effects of r-Hcy and Methionine on Ferryl Hb Decay
3.7. The Effects of r-Hcy and Methionine on MetHb Formation in Rosup-Treated Erythrocytes
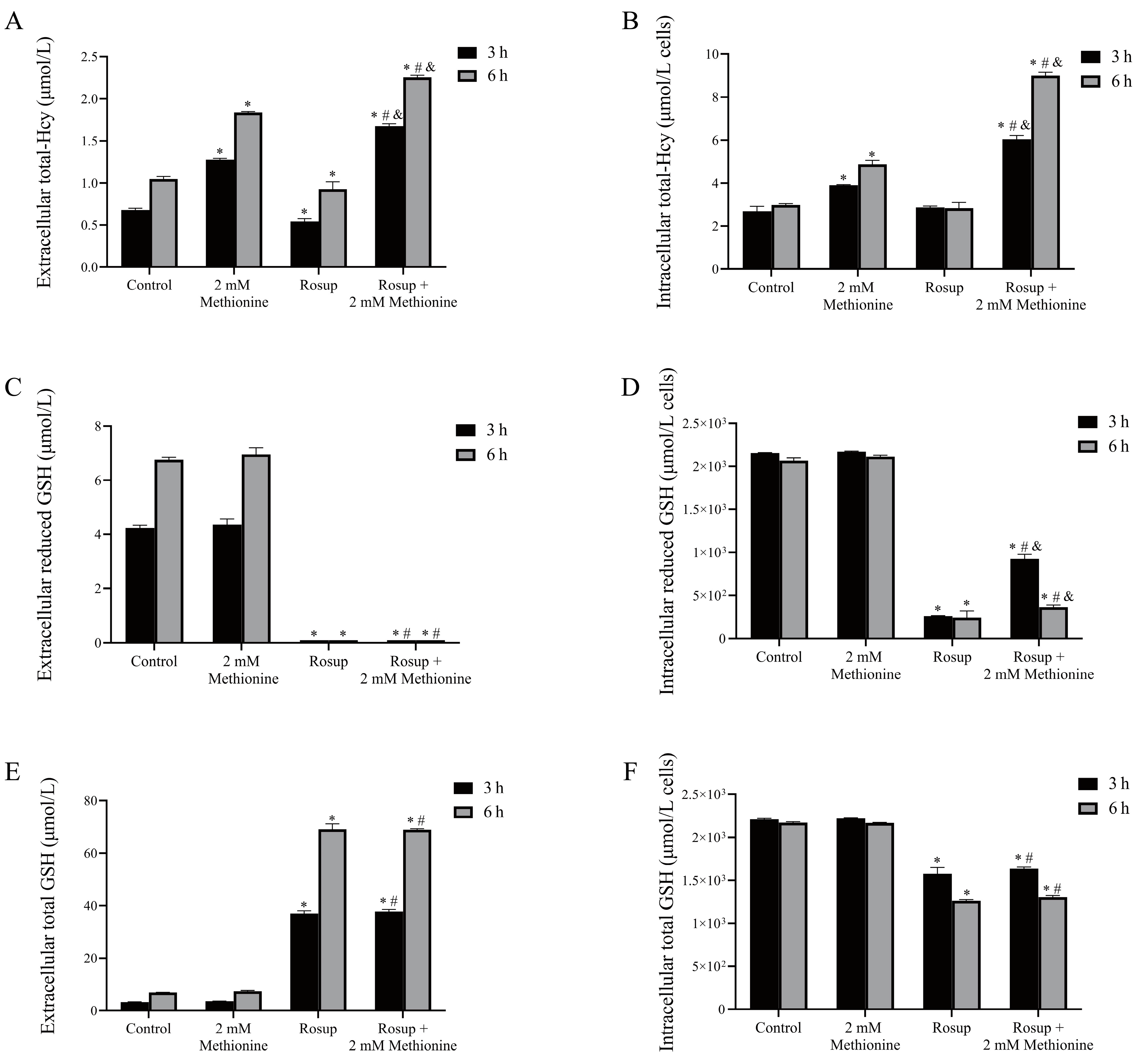
3.8. The Alterations of Total-Hcy and GSH Levels in Erythrocytes after Treatment with Rosup
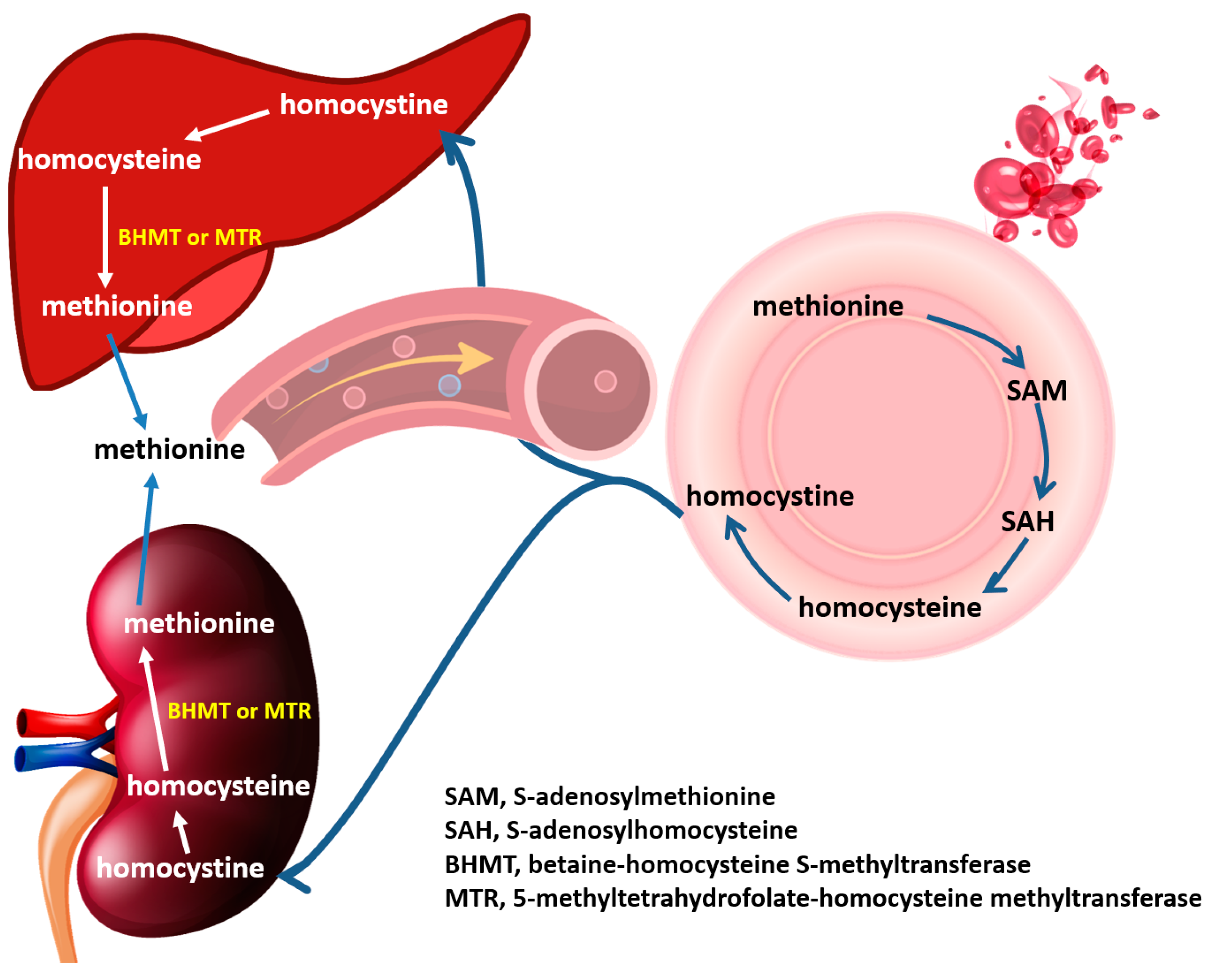
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jakubowski, H. Homocysteine Modification in Protein Structure/Function and Human Disease. Physiol. Rev. 2019, 99, 555–604. [Google Scholar] [CrossRef] [PubMed]
- Ueland, P.M.; Refsum, H.; Stabler, S.P.; Malinow, M.R.; Andersson, A.; Allen, R.H. Total homocysteine in plasma or serum: Methods and clinical applications. Clin. Chem. 1993, 39, 1764–1779. [Google Scholar] [CrossRef]
- Sawuła, W.; Banecka-Majkutewicz, Z.; Kadziński, L.; Jakóbkiewicz-Banecka, J.; Wegrzyn, G.; Nyka, W.; Banecki, B. Improved HPLC method for total plasma homocysteine detection and quantification. Acta. Biochim. Pol. 2008, 55, 119–125. [Google Scholar] [CrossRef]
- Škovierová, H.; Vidomanová, E.; Mahmood, S.; Sopková, J.; Drgová, A.; Červeňová, T.; Halašová, E.; Lehotský, J. The Molecular and Cellular Effect of Homocysteine Metabolism Imbalance on Human Health. Int. J. Mol. Sci. 2016, 17, 1733. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Palfrey, H.; Pathak, R.; Kadowitz, P.; Gettys, T.; Murthy, S. The metabolism and significance of homocysteine in nutrition and health. Nutr. Metab. 2017, 14, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kim, H.; Roh, H.; Kwon, Y. Causes of hyperhomocysteinemia and its pathological significance. Arch. Pharm. Res. 2018, 41, 372–383. [Google Scholar] [CrossRef]
- Paganelli, F.; Mottola, G.; Fromonot, J.; Marlinge, M.; Deharo, P.; Guieu, R.; Ruf, J. Hyperhomocysteinemia and Cardiovascular Disease: Is the Adenosinergic System the Missing Link? Int. J. Mol. Sci. 2021, 22, 1690. [Google Scholar] [CrossRef] [PubMed]
- Chwatko, G.; Bald, E. Determination of different species of homocysteine in human plasma by high-performance liquid chromatography with ultraviolet detection. J. Chromatogr. A 2002, 949, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Głowacki, R.; Bald, E. Fully automated method for simultaneous determination of total cysteine, cysteinylglycine, glutathione and homocysteine in plasma by HPLC with UV absorbance detection. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2009, 877, 3400–3404. [Google Scholar] [CrossRef]
- Ji, Y.; Luo, H.; Li, H.; Lin, Z.; Luo, W. Determination of plasma homocysteine with a UHPLC-MS/MS method: Application to analyze the correlation between plasma homocysteine and whole blood 5-methyltetrahydrofolate in healthy volunteers. Biomed. Chromatogr. 2020, 34, e4845. [Google Scholar] [CrossRef]
- Fiskerstrand, T.; Refsum, H.; Kvalheim, G.; Ueland, P.M. Homocysteine and other thiols in plasma and urine: Automated determination and sample stability. Clin. Chem. 1993, 39, 263–271. [Google Scholar] [CrossRef]
- Bianconi, E.; Piovesan, A.; Facchin, F.; Beraudi, A.; Casadei, R.; Frabetti, F.; Vitale, L.; Pelleri, M.C.; Tassani, S.; Piva, F. An estimation of the number of cells in the human body. Ann. Hum. Biol. 2013, 40, 463–471. [Google Scholar] [CrossRef]
- Bettiol, A.; Galora, S.; Argento, F.R.; Fini, E.; Emmi, G.; Mattioli, I.; Bagni, G.; Fiorillo, C.; Becatti, M. Erythrocyte oxidative stress and thrombosis. Expert Rev. Mol. Med. 2022, 24, e31. [Google Scholar] [CrossRef]
- Lai, W.K.; Kan, M.Y. Homocysteine-Induced Endothelial Dysfunction. Ann. Nutr. Metab. 2015, 67, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.-H.; Guo, Z.; Feng, M.; Wu, Z.-Z.; He, Z.-M.; Xiong, Y. Protection of DDAH2 overexpression against homocysteine-induced impairments of DDAH/ADMA/NOS/NO pathway in endothelial cells. Cell Physiol. Biochem. 2012, 30, 1413–1422. [Google Scholar] [CrossRef]
- Esse, R.; Barroso, M.; de Almeida, I.T.; Castro, R. The Contribution of Homocysteine Metabolism Disruption to Endothelial Dysfunction: State-of-the-Art. Int. J. Mol. Sci. 2019, 20, 867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faverzani, J.L.; Hammerschmidt, T.G.; Sitta, A.; Deon, M.; Wajner, M.; Vargas, C.R. Oxidative Stress in Homocystinuria Due to Cystathionine ß-Synthase Deficiency: Findings in Patients and in Animal Models. Cell. Mol. Neurobiol. 2017, 37, 1477–1485. [Google Scholar] [CrossRef]
- Jacobsen, D.W. Hyperhomocysteinemia and oxidative stress: Time for a reality check? Arterioscler. Thromb. Vasc Biol. 2000, 20, 1182–1184. [Google Scholar] [CrossRef] [Green Version]
- Martí-Carvajal, A.; Solà, I.; Lathyris, D.; Dayer, M. Homocysteine-lowering interventions for preventing cardiovascular events. Cochrane Database Syst. Rev. 2017, 8, Cd006612. [Google Scholar] [PubMed]
- Bald, E.; Glowacki, R. 2-Chloro-1-Methylquinolinium Tetrafluoroborate As An Effective And Thiol Specific Uv-Tagging Reagent For Liquid Chromatography. J. Liq. Chromatogr. Relat. Technol. 2001, 24, 1323–1339. [Google Scholar] [CrossRef]
- Tan, W.S.D.; Lee, J.J.; Satish, R.; Ang, E.-T. Detectability of secretagogin in human erythrocytes. Neurosci. Lett. 2012, 526, 59–62. [Google Scholar] [CrossRef]
- Xu, M.; Du, J.; Cui, J.; Zhang, S.; Zhang, S.; Deng, M.; Zhang, W.; Li, H.; Yu, Z. Cell-Free Fat Extract Prevents Tail Suspension-Induced Bone Loss by Inhibiting Osteocyte Apoptosis. Front. Bioeng. Biotechnol. 2022, 10, 818572. [Google Scholar] [CrossRef]
- Jin, Y.; Tan, Y.; Zhao, P.; Guo, Y.; Chen, S.; Wu, J.; Ren, Z. Glutathione S-transferase Mu 2 inhibits hepatic steatosis via ASK1 suppression. Commun. Biol. 2022, 5, 326. [Google Scholar] [CrossRef] [PubMed]
- Chihuailaf, R.H.; Contreras, P.; Wittwer, F. Pathogenesis of oxidative stress: Consequences and evaluation in animal health. Veterinaria México 2002, 33, 265. [Google Scholar]
- Sharma, B.; Rai, D.; Rai, P.K.; Rizvi, S.I.; Watal, G. Determination of erythrocyte fragility as a marker of pesticide-induced membrane oxidative damage. Methods Mol. Biol. 2010, 594, 123–128. [Google Scholar] [PubMed]
- Veena, C.K.; Josephine, A.; Preetha, S.P.; Varalakshmi, P. Effect of sulphated polysaccharides on erythrocyte changes due to oxidative and nitrosative stress in experimental hyperoxaluria. Hum. Exp. Toxicol. 2007, 26, 923–932. [Google Scholar] [CrossRef]
- Lu, N.; He, Y.; Chen, C.; Tian, R.; Xiao, Q.; Peng, Y.-Y. Tyrosine can protect against oxidative stress through ferryl hemoglobin reduction. Toxicol. Vitr. 2014, 28, 847–855. [Google Scholar] [CrossRef]
- Reeder, B.; Cutruzzola, F.; Bigotti, M.G.; Hider, R.; Wilson, M. Tyrosine as a redox-active center in electron transfer to ferryl heme in globins. Free Radic. Biol. Med. 2008, 44, 274–283. [Google Scholar] [CrossRef]
- Das, D.; Sen, S.; Sen, K. Disparity of selenourea and selenocystine on methaemoglobinemia in non-diabetics and diabetics. J. Biochem. 2021, 169, 371–382. [Google Scholar] [CrossRef]
- Bald, E.; Chwatko, G.; Głowacki, R.; Kuśmierek, K. Analysis of plasma thiols by high-performance liquid chromatography with ultraviolet detection. J. Chromatogr. A 2004, 1032, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Rocha, S.; Gomes, D.; Lima, M.; Bronze-da-Rocha, E.; Santos-Silva, A. Peroxiredoxin 2, glutathione peroxidase, and catalase in the cytosol and membrane of erythrocytes under H2O2-induced oxidative stress. Free Radic. Res. 2015, 49, 990–1003. [Google Scholar] [CrossRef]
- Harley, J.D. Role of reduced glutathione in human erythrocytes. Nature 1965, 206, 1054–1055. [Google Scholar] [CrossRef]
- DeLeve, L.D.; Kaplowitz, N. Glutathione metabolism and its role in hepatotoxicity. Pharmacol. Ther. 1991, 52, 287–305. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M. The definition and measurement of antioxidants in biological systems. Free Radic. Biol. Med. 1995, 18, 125–126. [Google Scholar] [CrossRef]
- Ji, L.L.; Yeo, D. Oxidative stress: An evolving definition. Fac. Rev. 2021, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.S.; Bhattacharya, R.; Singh, L.R. Functional inhibition of redox regulated heme proteins: A novel mechanism towards oxidative stress induced by homocysteine. Redox Biol. 2021, 46, 102080. [Google Scholar] [CrossRef] [PubMed]
- Fowler, B. Homocysteine: Overview of biochemistry, molecular biology, and role in disease processes. Semin. Vasc. Med. 2005, 5, 77–86. [Google Scholar] [CrossRef]
- Baggott, J.E.; Tamura, T. Iron-dependent formation of homocysteine from methionine and other thioethers. Eur. J. Clin. Nutr. 2007, 61, 1359–1363. [Google Scholar] [CrossRef] [Green Version]
- Lutz, H.U.; Bogdanova, A. Mechanisms tagging senescent red blood cells for clearance in healthy humans. Front. Physiol. 2013, 4, 387. [Google Scholar] [CrossRef] [Green Version]
- Remigante, A.; Morabito, R.; Marino, A. Band 3 protein function and oxidative stress in erythrocytes. J. Cell. Physiol. 2021, 236, 6225–6234. [Google Scholar] [CrossRef]
- Remigante, A.; Spinelli, S.; Basile, N.; Caruso, D.; Falliti, G.; Dossena, S.; Marino, A.; Morabito, R. Oxidation Stress as a Mechanism of Aging in Human Erythrocytes: Protective Effect of Quercetin. Int. J. Mol. Sci. 2022, 23, 7781. [Google Scholar] [CrossRef]
- Reeder, B.J. The redox activity of hemoglobins: From physiologic functions to pathologic mechanisms. Antioxid. Redox Signal. 2010, 13, 1087–1123. [Google Scholar] [CrossRef]
- George, P.; Irvine, D.H. The reaction between metmyoglobin and hydrogen peroxide. Biochem. J. 1952, 52, 511–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutaud, O.; Moore, K.; Reeder, B.; Harry, D.; Howie, A.; Wang, S.; Carney, C.; Masterson, T.; Amin, T.; Wright, D. Acetaminophen inhibits hemoprotein-catalyzed lipid peroxidation and attenuates rhabdomyolysis-induced renal failure. Proc. Natl. Acad. Sci. USA 2010, 107, 2699–2704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeder, B.J.; Wilson, M.T. Mechanism of reaction of myoglobin with the lipid hydroperoxide hydroperoxyoctadecadienoic acid. Biochem. J. 1998, 330 Pt 3, 1317–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, P.; Stratmann, C.J. The oxidation of myoglobin to metmyoglobin by oxygen. I. Biochem. J. 1952, 51, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Buehler, P.W.; Boykins, R.A.; Venable, R.M.; Alayash, A.I. Structural basis of peroxide-mediated changes in human hemoglobin: A novel oxidative pathway. J. Biol. Chem. 2007, 282, 4894–4907. [Google Scholar] [CrossRef] [Green Version]
- Portillo, F.; Vázquez, J.; Pajares, M.A. Protein-protein interactions involving enzymes of the mammalian methionine and homocysteine metabolism. Biochimie 2020, 173, 33–47. [Google Scholar] [CrossRef]
- Pajares, M.A.; Pérez-Sala, D. Betaine homocysteine S-methyltransferase: Just a regulator of homocysteine metabolism? Cell. Mol. Life Sci. 2006, 63, 2792–2803. [Google Scholar] [CrossRef] [Green Version]
- Finkelstein, J.D.; Martin, J.J. Methionine metabolism in mammals. Distribution of homocysteine between competing pathways. J. Biol. Chem. 1984, 259, 9508–9513. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, M.; Li, H.; Luo, H.; Zhou, Y.; Luo, W.; Lin, Z. Potential Antioxidative Activity of Homocysteine in Erythrocytes under Oxidative Stress. Antioxidants 2023, 12, 202. https://doi.org/10.3390/antiox12010202
Ye M, Li H, Luo H, Zhou Y, Luo W, Lin Z. Potential Antioxidative Activity of Homocysteine in Erythrocytes under Oxidative Stress. Antioxidants. 2023; 12(1):202. https://doi.org/10.3390/antiox12010202
Chicago/Turabian StyleYe, Mingxin, Hui Li, Hongjun Luo, Yongyin Zhou, Wenhong Luo, and Zhexuan Lin. 2023. "Potential Antioxidative Activity of Homocysteine in Erythrocytes under Oxidative Stress" Antioxidants 12, no. 1: 202. https://doi.org/10.3390/antiox12010202