
Above the Invasive and Ornamental Attributes of the Traveler’s Palm: An In Vitro and In Silico Insight into the Anti-Oxidant, Anti-Enzymatic, Cytotoxic and Phytochemical Characterization of Ravenala madagascariensis

 ,
,  , , , ,
, , , ,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction of Phytochemicals
2.2. Phytochemical Composition
2.3. HPLC-ESI-Q-TOF-MS-MS Analysis
2.4. Biological Activities Evaluation
2.4.1. Antioxidant Assays
2.4.2. Anticholinesterase Activities
2.4.3. Anti-Tyrosinase Activities
2.4.4. Alpha-Amylase and Alpha-Glucosidase Assays
2.5. Cell Viability Assay
2.6. In Silico Docking Studies
2.7. Statistical Analysis
3. Results
3.1. Total Bioactive Components
3.2. Characterization of Phytochemicals by HPLC-ESI-Q-TOF-MS-MS
3.3. Antioxidant Properties
3.4. Enzyme Inhibitory Properties
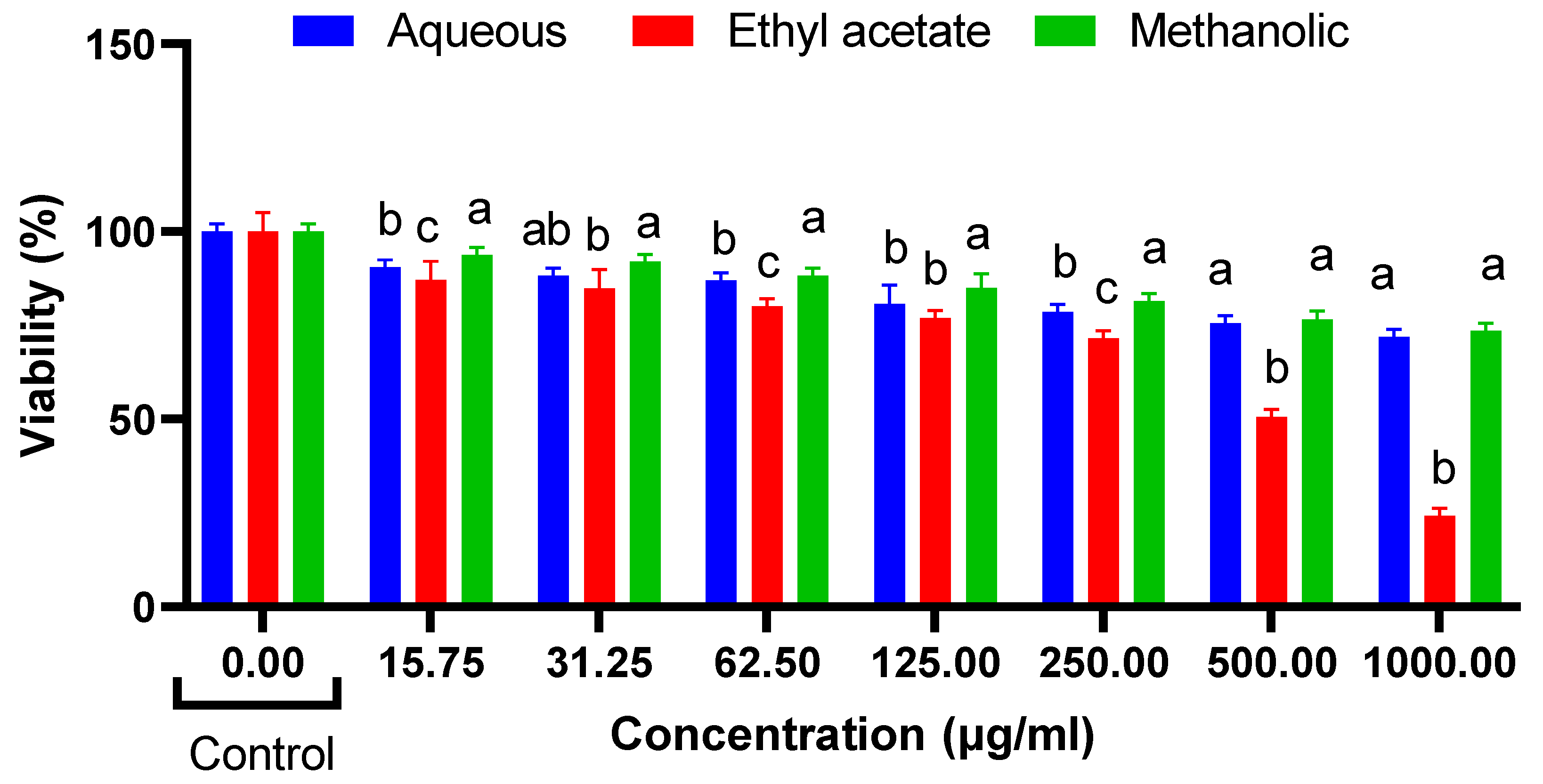
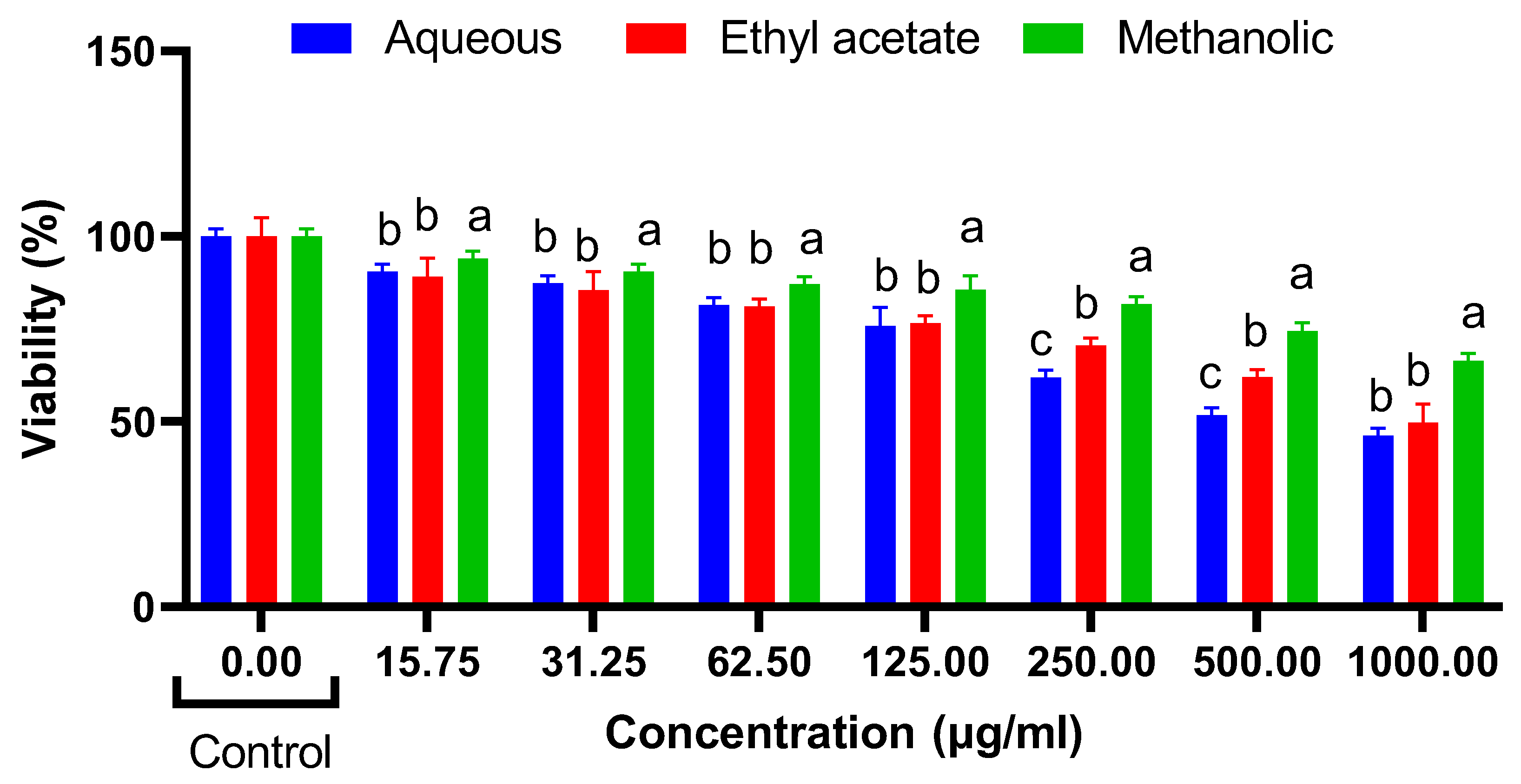
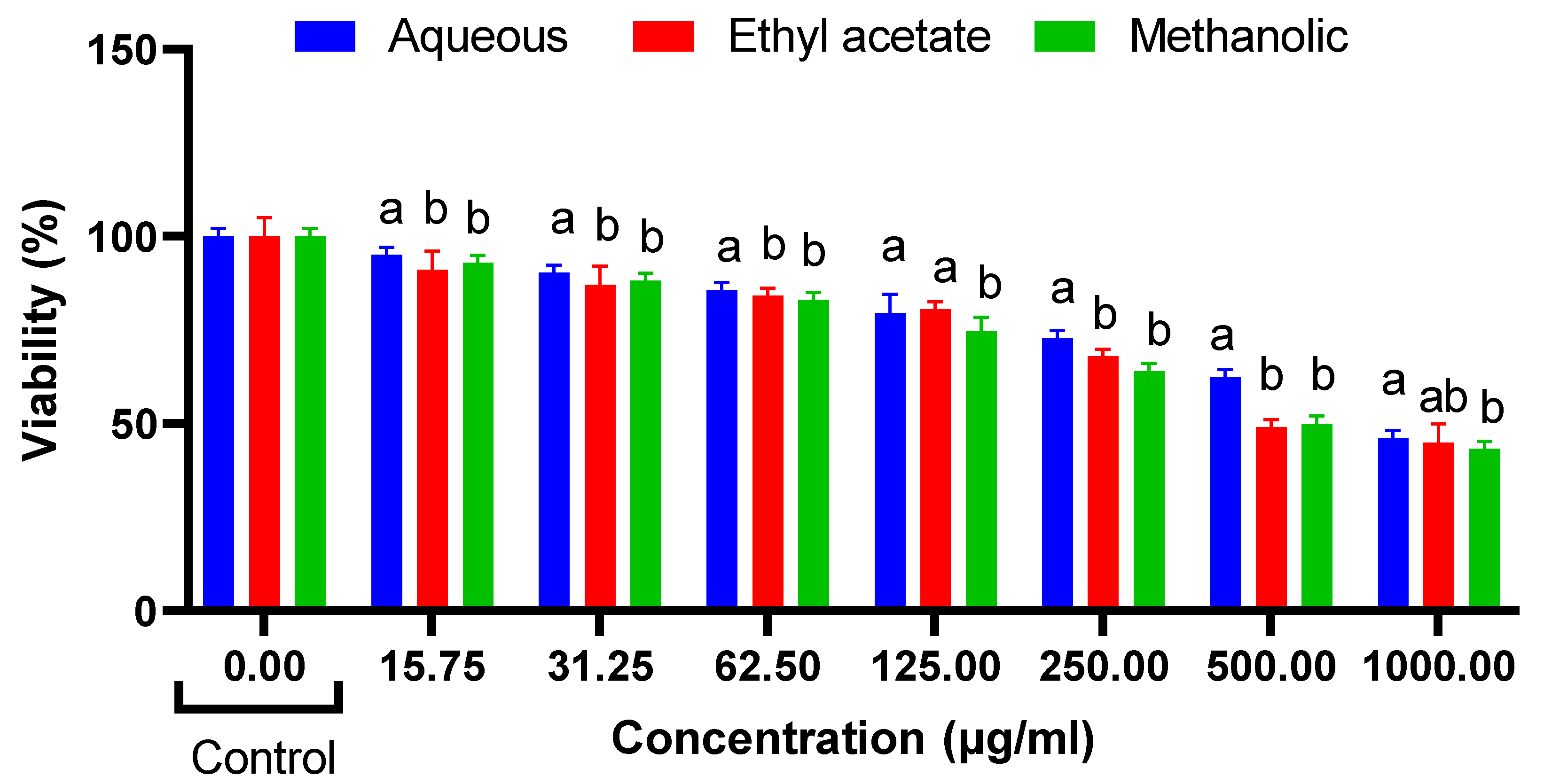
Cytotoxic Effects
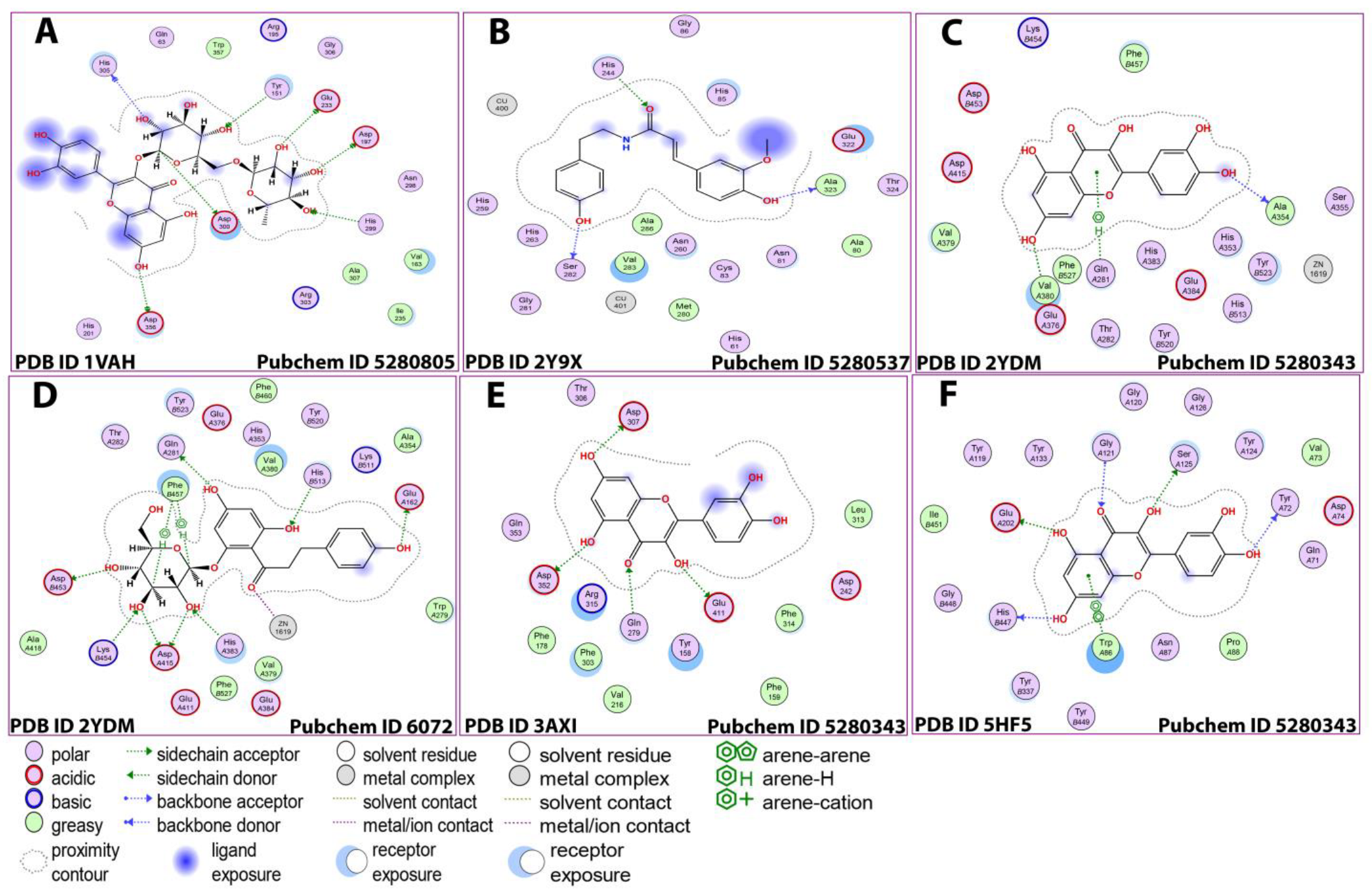
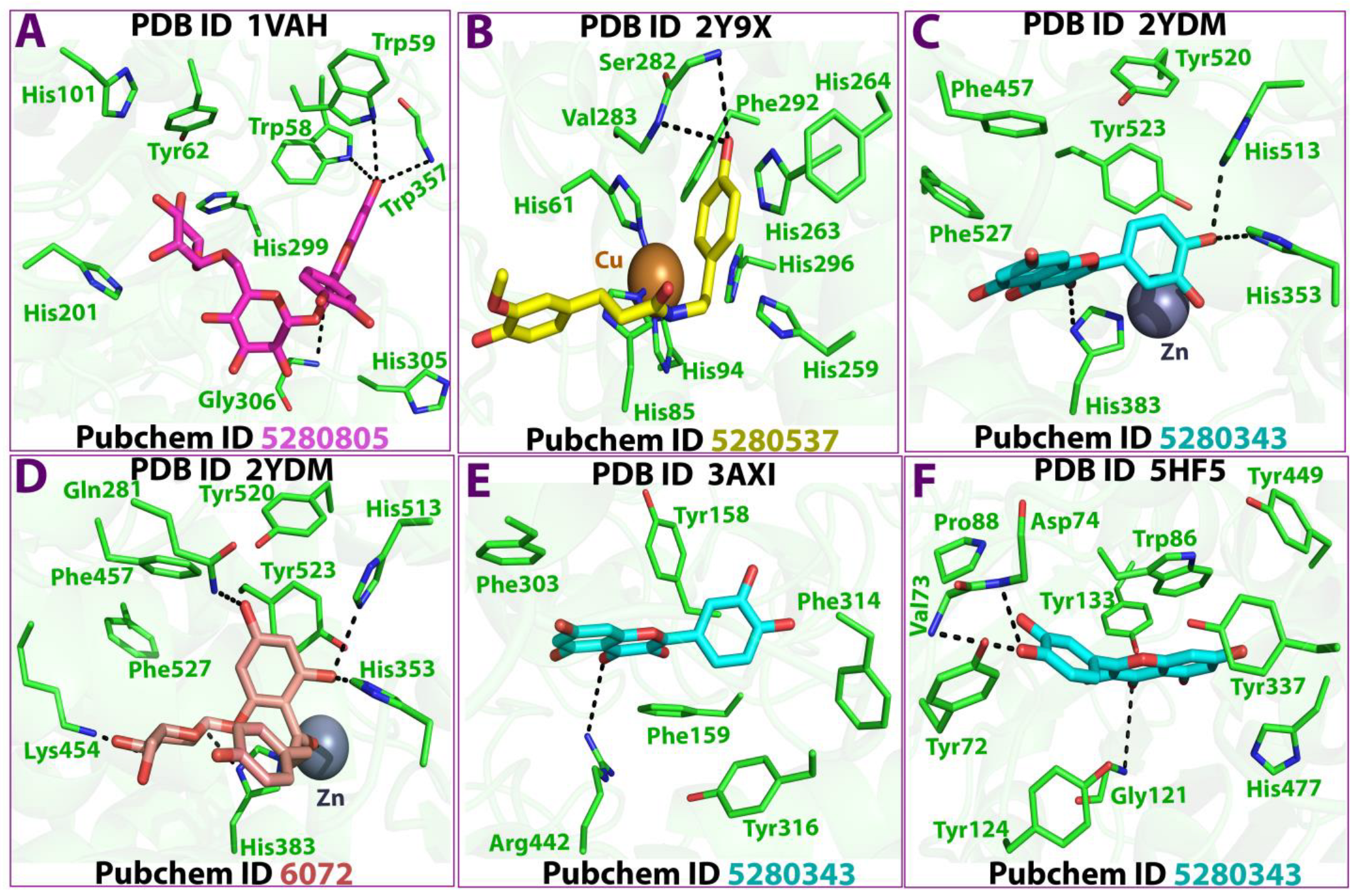
3.5. In Silico Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hladik, M.; Blanc, P.; Hladik, A. L’arbre du voyageur: Des usages et de la diffusion horticole du ravenala. Hommes Plantes 2002, 40, 39–47. [Google Scholar]
- Rakotoarivelo, N.; Razanatsima, A.; Rakotoarivony, F.; Rasoaviety, L.; Ramarosandratana, A.V.; Jeannoda, V.; Kuhlman, A.R.; Randrianasolo, A.; Bussmann, R.W. Ethnobotanical and economic value of Ravenala madagascariensis Sonn. in Eastern Madagascar. J. Ethnobiol. Ethnomed. 2014, 10, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakotoarivelo, N.H.; Rakotoarivony, F.; Ramarosandratana, A.V.; Jeannoda, V.H.; Kuhlman, A.R.; Randrianasolo, A.; Bussmann, R.W. Medicinal plants used to treat the most frequent diseases encountered in Ambalabe rural community, Eastern Madagascar. J. Ethnobiol. Ethnomed. 2015, 11, 68. [Google Scholar] [CrossRef] [Green Version]
- Ramiarantsoa, H.; Yao-Kouassi, P.A.; Kanko, C.; Assi, K.M.; Djakoure, A.L.; Tonzibo, F.Z. Chemical constituents of the antidiabetic plant Ravenala madagascariensis. Int. J. Pharm. Sci. Res. 2014, 5, 5503–5510. [Google Scholar]
- Duvale, S.A.H.; Judicael, R.L.; Ranjana, R.H.; Doll, R.D.A.; Louis, J.V. Antimicrobial and antioxidant activities of the fruits of bemavo, a variety of Ravenala madagascariensis Sonn. (Strelitziaceae). World J. Biol. Pharm. Health Sci. 2020, 2, 030–039. [Google Scholar]
- Priyadarsini, S.S.; Vadivu, R.; Vijayalakshmi, A.; Kumar, P.R. Antioxidant activity of Ravenala madagascariensis Sonn. leaves on alloxan induced diabetic rats. Int. J. PharmTech Res. 2013, 5, 1823–1827. [Google Scholar]
- Sharmin, T.; Choudhury, S.; Chowdhury, M.; Mian, M.; Hoque, M.; Sumsujjaman, M.; Nahar, F. Evaluation of antimicrobial activities of some Bangladeshi medicinal plants. World J. Pharm. Sci. 2014, 2, 137–209. [Google Scholar]
- Rabearivony, A.D.; Kuhlman, A.R.; Razafiariso, Z.L.; Raharimalala, F.; Rakotoarivony, F.; Randrianarivony, T.; Rakotoarivelo, N.; Randrianasolo, A.; Bussmann, R.W. Ethnobotanical Study of the Medicinal Plants Known by Men in Ambalabe, Madagascar. Ethnobot. Res. Appl. 2015, 14, 123–138. [Google Scholar] [CrossRef] [Green Version]
- Razafindraibe, M.; Kuhlman, A.R.; Rabarison, H.; Rakotoarimanana, V.; Rajeriarison, C.; Rakotoarivelo, N.; Randrianarivony, T.; Rakotoarivony, F.; Ludovic, R.; Randrianasolo, A.; et al. Medicinal plants used by women from Agnalazaha littoral forest (Southeastern Madagascar). J. Ethnobiol. Ethnomed. 2013, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, S.; Sharmin, T.; Hoque, M.; Sumsujjaman, M.; Das, M.; Nahar, F. Evaluation of thrombolytic and membrane stabilizing activities of four medicinal plants of Bangladesh. Int. J. Pharm. Sci. Res. 2013, 4, 4223–4227. [Google Scholar]
- Onifade, A.K.; Bello, M.O.; Fadipe, D. Bioassay directed fractionation of antibacterial compounds from traveller’s tree (ravenala Madagascariensis sonnerat) and its phytochemical constituents. Int. J. Bioassays 2015, 4, 4299–4304. [Google Scholar]
- Ganzera, M.; Sturm, S. Recent advances on HPLC/MS in medicinal plant analysis—An update covering 2011–2016. J. Pharm. Biomed. Anal. 2018, 147, 211–233. [Google Scholar] [CrossRef]
- Forli, S.; Huey, R.; Pique, M.E.; Sanner, M.F.; Goodsell, D.S.; Olson, A.J. Computational protein–ligand docking and virtual drug screening with the AutoDock suite. Nat. Protoc. 2016, 11, 905–919. [Google Scholar] [CrossRef] [Green Version]
- Uysal, S.; Aktumsek, A. A phytochemical study on Potentilla anatolica: An endemic Turkish plant. Ind. Crops Prod. 2015, 76, 1001–1007. [Google Scholar] [CrossRef]
- Zengin, G.; Guler, G.O.; Aktumsek, A.; Ceylan, R.; Picot, C.M.N.; Mahomoodally, M.F. Enzyme inhibitory properties, antioxidant activities, and phytochemical profile of three medicinal plants from Turkey. Adv. Pharmacol. Sci. 2015, 2015, 410675. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Poyatos, M.D.P.; Ruiz-Medina, A.; Zengin, G.; Llorent-Martínez, E.J. Phenolic Characterization, Antioxidant Activity, and Enzyme Inhibitory Properties of Berberis thunbergii DC. Leaves: A Valuable Source of Phenolic Acids. Molecules 2019, 24, 4171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mocan, A.; Zengin, G.; Simirgiotis, M.; Schafberg, M.; Mollica, A.; Vodnar, D.C.; Crişan, G.; Rohn, S. Functional constituents of wild and cultivated Goji (L. barbarum L.) leaves: Phytochemical characterization, biological profile, and computational studies. J. Enzym. Inhib. Med. Chem. 2017, 32, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Llorent-Martínez, E.J.; Ruiz-Medina, A.; Zengin, G.; Ak, G.; Jugreet, S.; Mahomoodally, M.F.; Emre, G.; Orlando, G.; Libero, M.L.; Acquaviva, A.; et al. New Biological and Chemical Evidences of Two LamiaceaeSpecies (Thymbra capitata and Thymus sipyleus subsp. rosulans): In Vitro, In Silico and Ex Vivo Approaches. Molecules 2022, 27, 9029. [Google Scholar] [CrossRef]
- Llorent-Martínez, E.J.; Spínola, V.; Gouveia, S.; Castilho, P.C. HPLC-ESI-MSn characterization of phenolic compounds, terpenoid saponins, and other minor compounds in Bituminaria bituminosa. Ind. Crops Prod. 2015, 69, 80–90. [Google Scholar] [CrossRef]
- de Souza, L.M.; Cipriani, T.R.; Iacomini, M.; Gorin, P.A.; Sassaki, G.L. HPLC/ESI-MS and NMR analysis of flavonoids and tannins in bioactive extract from leaves of Maytenus ilicifolia. J. Pharm. Biomed. Anal. 2008, 47, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Hamed, A.I.; Al-Ayed, A.S.; Moldoch, J.; Piacente, S.; Oleszek, W.; Stochmal, A. Profiles analysis of proanthocyanidins in the argun nut (Medemia argun--an ancient Egyptian palm) by LC-ESI-MS/MS. J. Mass Spectrom. 2014, 49, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Mateos-Martín, M.L.; Fuguet, E.; Jiménez-Ardón, A.; Herrero-Uribe, L.; Tamayo-Castillo, G.; Torres, J.L. Identification of polyphenols from antiviral Chamaecrista nictitans extract using high-resolution LC-ESI-MS/MS. Anal. Bioanal. Chem. 2014, 406, 5501–5506. [Google Scholar] [CrossRef] [PubMed]
- Reyad-ul-Ferdous, M.; Nasir, U.; Shahjahan, D.; Mosharaf, H.; Arman, M.; Ariful, I. Preliminary in-vitro potential phytochemicals investigation of barks of Ravenala madagascariensis Sonnerat. World J. Pharm. Pharm. Sci. 2014, 3, 1418–1423. [Google Scholar]
- Kamzolova, S.V.; Samoilenko, V.A.; Lunina, J.N.; Morgunov, I.G. Isocitric Acid Production from Ethanol Industry Waste by Yarrowia lipolytica. Fermentation 2021, 7, 146. [Google Scholar] [CrossRef]
- Akaranta, O.; Akaho, A.A. Synergic Effect of Citric Acid and Peanut Skin Extract on the Oxidative Stability of Vegetable Oil. J. Appl. Sci. Environ. Manag. 2012, 16, 345–351. [Google Scholar]
- Ribeiro, B.; Lopes, R.; Andrade, P.B.; Seabra, R.M.; Gonçalves, R.F.; Baptista, P.; Quelhas, I.; Valentão, P.C. Comparative study of phytochemicals and antioxidant potential of wild edible mushroom caps and stipes. Food Chem. 2008, 110, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Ricciutelli, M.; Bartolucci, G.; Campana, R.; Salucci, S.; Benedetti, S.; Caprioli, G.; Maggi, F.; Sagratini, G.; Vittori, S.; Lucarini, S. Quantification of 2- and 3-isopropylmalic acids in forty Italian wines by UHPLC-MS/MS triple quadrupole and evaluation of their antimicrobial, antioxidant activities and biocompatibility. Food Chem. 2020, 321, 126726. [Google Scholar] [CrossRef] [PubMed]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Zhang, Y.; Tang, C.; Hou, Y.; Ai, X.; Chen, X.; Zhang, Y.; Wang, X.; Meng, X. Gallic acid: Pharmacological activities and molecular mechanisms involved in inflammation-related diseases. Biomed. Pharmacother. 2021, 133, 110985. [Google Scholar] [CrossRef]
- Randjelovic, P.; Veljković, S.; Stojiljković, N.; Sokolovic, D.; Ilić, I.; Laketić, D.; Randjelović, D.; Randjelović, N. The Beneficial Biological Properties of Salicylic Acid. Acta Fac. Med. Naissensis 2015, 32, 259–265. [Google Scholar] [CrossRef]
- Qu, Z.; Liu, A.; Li, P.; Liu, C.; Xiao, W.; Huang, J.; Liu, Z.; Zhang, S. Advances in physiological functions and mechanisms of (-)-epicatechin. Crit. Rev. Food Sci. Nutr. 2021, 61, 211–233. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G.E.; Beshbishy, A.M.; Ikram, M.; Mulla, Z.S.; El-Hack, M.E.A.; Taha, A.E.; Algammal, A.M.; Elewa, Y.H.A. The Pharmacological Activity, Biochemical Properties, and Pharmacokinetics of the Major Natural Polyphenolic Flavonoid: Quercetin. Foods 2020, 9, 374. [Google Scholar] [CrossRef] [Green Version]
- Calderón-Montaño, J.M.; Burgos-Morón, E.; Pérez-Guerrero, C.; López-Lázaro, M. A review on the dietary flavonoid kaempferol. Mini Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef]
- Santos, T.C.d.; Gomes, T.M.; Pinto, B.A.S.; Camara, A.L.; Paes, A.M.d.A. Naturally Occurring Acetylcholinesterase Inhibitors and Their Potential Use for Alzheimer’s Disease Therapy. Front. Pharmacol. 2018, 9, 1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mervić, M.; Bival Štefan, M.; Kindl, M.; Blažeković, B.; Marijan, M.; Vladimir-Knežević, S. Comparative Antioxidant, Anti-Acetylcholinesterase and Anti-α-Glucosidase Activities of Mediterranean Salvia Species. Plants 2022, 11, 625. [Google Scholar] [CrossRef]
- Taqui, R.; Debnath, M.; Ahmed, S.; Ghosh, A. Advances on plant extracts and phytocompounds with acetylcholinesterase inhibition activity for possible treatment of Alzheimer’s disease. Phytomed. Plus 2022, 2, 100184. [Google Scholar] [CrossRef]
- Younis, M.M.; Ayoub, I.M.; Mostafa, N.M.; El Hassab, M.A.; Eldehna, W.M.; Al-Rashood, S.T.; Eldahshan, O.A. GC/MS Profiling, Anti-Collagenase, Anti-Elastase, Anti-Tyrosinase and Anti-Hyaluronidase Activities of a Stenocarpus sinuatus Leaves Extract. Plants 2022, 11, 918. [Google Scholar] [CrossRef]
- Ekins, S.; Mestres, J.; Testa, B. In silico pharmacology for drug discovery: Methods for virtual ligand screening and profiling. Br. J. Pharmacol. 2007, 152, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Yarley, O.P.N.; Kojo, A.B.; Zhou, C.; Yu, X.; Gideon, A.; Kwadwo, H.H.; Richard, O. Reviews on mechanisms of in vitro antioxidant, antibacterial and anticancer activities of water-soluble plant polysaccharides. Int. J. Biol. Macromol. 2021, 183, 2262–2271. [Google Scholar] [CrossRef]
- Wong, M.C.; Jiang, J.Y.; Goggins, W.B.; Liang, M.; Fang, Y.; Fung, F.D.; Leung, C.; Wang, H.H.; Wong, G.L.; Wong, V.W.; et al. International incidence and mortality trends of liver cancer: A global profile. Sci. Rep. 2017, 7, 45846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Mullins, C.S.; Schafmayer, C.; Zeißig, S.; Linnebacher, M. A global assessment of recent trends in gastrointestinal cancer and lifestyle-associated risk factors. Cancer Commun. 2021, 41, 1137–1151. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Phenolic Content (mg GAE/g) | Total Flavonoid Content (mg RE/g) | Total Phenolic Acid Content (mg CAE/g) | Total Flavonol Content (mg CE/g) |
|---|---|---|---|---|
| RM-EA | 41.13 ± 0.2 c | 25.04 ± 0.29 b | 7.65 ± 0.52 c | 32.22 ± 0.46 a |
| RM-MEOH | 61.06 ± 0.75 b | 32.26 ± 0.48 a | 16.44 ± 0.47 a | 33.73 ± 0.53 a |
| RM-Aq | 64.47 ± 0.19 a | 25.30 ± 0.16 b | 14.05 ± 0.49 b | 2.87 ± 0.08 b |
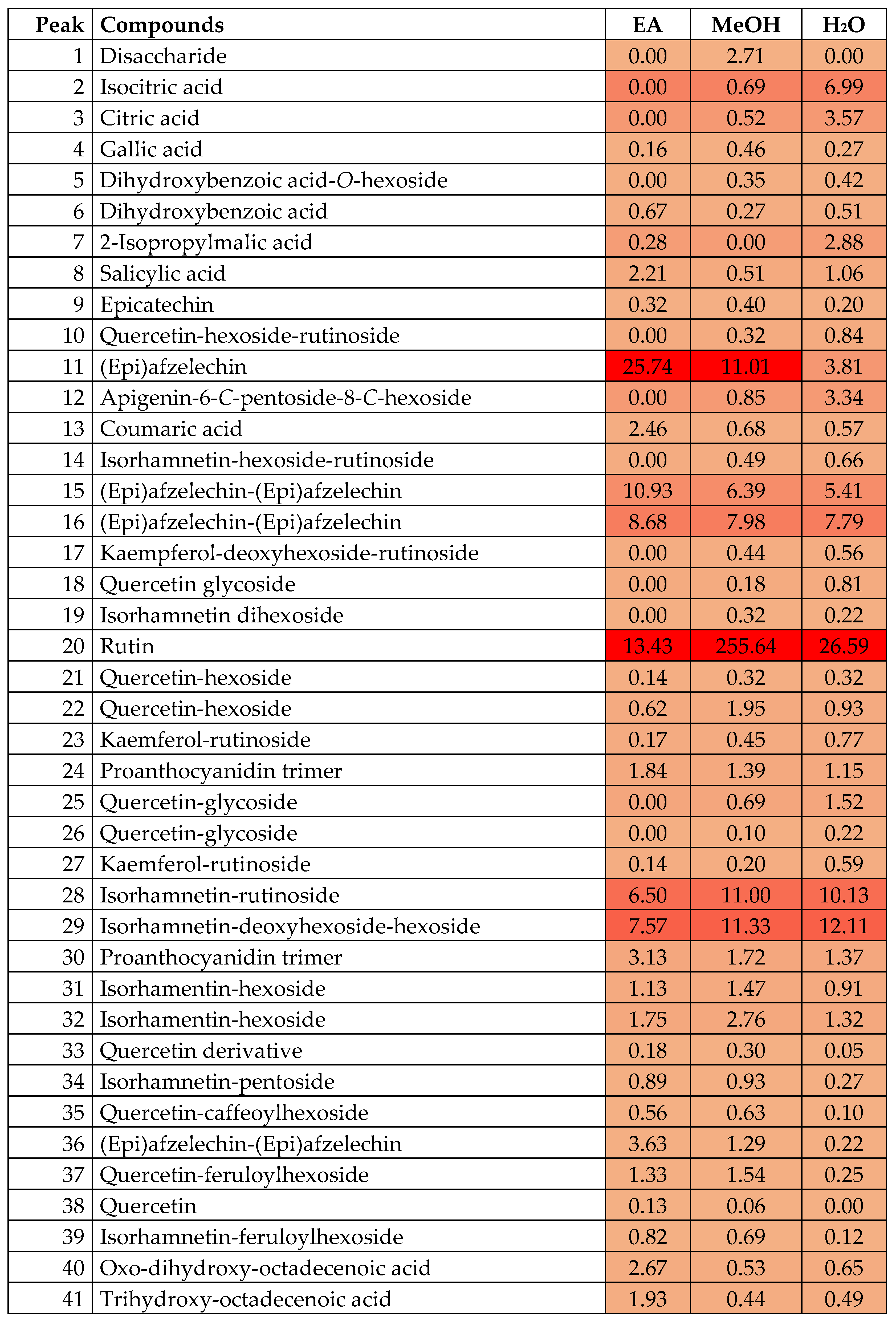
| No. | tR (min) | Observed [M-H]- | Molecular Formula | Error (ppm) | Fragment Ions | Assigned Identification | EA | MeOH | H2O |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 1.8 | 341.1094 | C12H22O11 | −1.36 | 179.0562, 161.0453, 131.0341, 119.0347, 89.0246 | Disaccharide (two hexosides) | ✓ | ||
| 2 | 2.0 | 191.0207 | C6H8O7 | −4.82 | 173.0092, 129.0193, 111.0086 | Isocitric acid | ✓ | ✓ | |
| 3 | 2.7 | 191.0202 | C6H8O7 | −2.36 | 173.0088, 129.0175, 111.0081 | Citric acid | ✓ | ✓ | |
| 4 | 3.2 | 169.0145 | C7H6O5 | −1.38 | 125.0242 | Gallic acid | ✓ | ✓ | ✓ |
| 5 | 3.6 | 315.0727 | C13H16O9 | −2.54 | 153.0187, 109.0294 | Dihydroxybenzoic acid-O-hexoside | ✓ | ✓ | |
| 6 | 5.4 | 153.0193 | C7H6O4 | −1.63 | 109.0293 | Dihydroxybenzoic acid | ✓ | ✓ | ✓ |
| 7 | 6.8 | 175.0615 | C7H12O5 | −1.92 | 157.0505, 131.0713, 115.0399, 85.0661 | 2-Isopropylmalic acid | ✓ | ✓ | |
| 8 | 8.8 | 137.0242 | C7H6O3 | 1.13 | 93.0347 | Salicylic acid | ✓ | ✓ | ✓ |
| 9 | 12.1 | 289.0725 | C15H14O6 | −3.06 | 245.0797, 205.0482, 203.0709, 109.0288 | Epicatechin | ✓ | ✓ | ✓ |
| 10 | 14.4 | 771.1996 | C33H40O21 | −1.21 | 301.0339, 300.0275, 178.9983, 151.0013 | Quercetin-hexoside-rutinoside | ✓ | ✓ | |
| 11 | 15.4 | 273.0777 | C15H14O5 | −3.16 | 255.0653, 229.0882, 205.0868, 187.0749, 137.0237, 107.0494, 97.0291 | (Epi)afzelechin | ✓ | ✓ | |
| 12 | 15.8 | 563.1419 | C26H28O14 | −2.11 | 545.1310, 503.1201, 473.1100, 443.0993, 383.0775, 353.0672 | Apigenin-6-C-pentoside-8-C-hexoside | ✓ | ✓ | |
| 13 | 16.5 | 163.0401 | C9H8O3 | −0.57 | 119.0499 | Coumaric acid | ✓ | ✓ | ✓ |
| 14 | 17.2 | 785.2153 | C34H42O21 | −1.06 | 315.0496, 300.0267, 151.0020 | Isorhamnetin-hexoside-rutinoside | ✓ | ✓ | |
| 15 | 17.6 | 545.1467 | C30H26O10 | −2.43 | 419.1139, 409.0929, 273.0768, 164.0108, 125.0243, 97.0297 | (Epi)afzelechin-(Epi)afzelechin | ✓ | ✓ | ✓ |
| 16 | 18.2 | 545.1470 | C30H26O10 | −2.89 | 419.1140, 409.0932, 273.0770, 164.0113, 125.0241, 97.0292 | (Epi)afzelechin-(Epi)afzelechin | ✓ | ✓ | ✓ |
| 17 | 18.4 | 739.2099 | C33H40O19 | −1.01 | 285.0386, 284.0327, 257.0454, 255.0291, 227.0342, 178.9996, 151.0027 | Kaempferol-deoxyhexoside-rutinoside | ✓ | ✓ | |
| 18 | 18.9 | 933.2302 | C42H46O24 | 0.57 | 771.1996, 631.1852, 301.0345, 300.0286, 178.9988, 151.0019 | Quercetin glycoside | ✓ | ✓ | |
| 19 | 19.1 | 639.1580 | C28H32O17 | −1.81 | 315.0486, 300.0268 | Isorhamnetin dihexoside | ✓ | ✓ | |
| 20 | 19.9 | 609.1478 | C27H30O16 | −2.75 | 301.0342, 300.0281, 178.9985, 151.0031 | Rutin | ✓ | ✓ | ✓ |
| 21 | 20.7 | 463.0891 | C21H20O12 | −2.03 | 301.0351, 178.9992, 151.0035 | Quercetin-hexoside | ✓ | ✓ | ✓ |
| 22 | 21.5 | 463.0886 | C21H20O12 | −2.39 | 301.0343, 178.9979, 151.0029 | Quercetin-hexoside | ✓ | ✓ | ✓ |
| 23 | 22.0 | 593.1523 | C27H30O15 | −1.79 | 285.0396, 284.324, 255.0305, 227.0343, 151.0043 | Kaemferol-rutinoside | ✓ | ✓ | ✓ |
| 24 | 22.6 | 817.2146 | C45H38O15 | −0.83 | 545.1451, 543.1301, 419.1144, 273.0762, 271.0616, 164.0117, 125.0232 | Proanthocyanidin trimer | ✓ | ✓ | ✓ |
| 25 | 22.6 | 947.2466 | C43H48O24 | −0.25 | 771.1975, 301.0338, 178.9961 | Quercetin-glycoside | ✓ | ✓ | |
| 26 | 23.1 | 917.2357 | C42H46O23 | 0.71 | 771.1977, 301.0328, 300.0290, 178.9973, 151.0397 | Quercetin-glycoside | ✓ | ✓ | |
| 27 | 23.3 | 593.1523 | C27H30O15 | −1.70 | 285.0403, 255.0309 | Kaemferol-rutinoside | ✓ | ✓ | |
| 28 | 23.4 | 623.1629 | C28H32O16 | −1.89 | 315.0517, 300.0270, 151.0034 | Isorhamnetin-rutinoside | ✓ | ✓ | ✓ |
| 29 | 24.0 | 623.1639 | C28H32O16 | −3.23 | 477.1082, 315.0517, 300.0275, 151.0038 | Isorhamnetin-deoxyhexoside-hexoside | ✓ | ✓ | ✓ |
| 30 | 24.2 | 817.2148 | C45H38O15 | −0.98 | 545.1428, 543.1283, 409.0903, 273.0766, 271.0617, 164.0106, 125.0239 | Proanthocyanidin trimer | ✓ | ✓ | ✓ |
| 31 | 24.6 | 477.1049 | C22H22O12 | −2.36 | 315.0496, 314.0437, 300.0270, 151.0038 | Isorhamentin-hexoside | ✓ | ✓ | ✓ |
| 32 | 25.3 | 477.1052 | C22H22O12 | −3.11 | 315.0484, 314.0434, 300.0254, 151.0032 | Isorhamentin-hexoside | ✓ | ✓ | ✓ |
| 33 | 27.1 | 613.1208 | C29H26O15 | −1.50 | 301.0376, 178.9995, 151.0033 | Quercetin derivative | ✓ | ✓ | ✓ |
| 34 | 27.4 | 447.0941 | C21H20O11 | −1.76 | 315.0469, 314.0437, 301.0337, 300.0260, 151.0013 | Isorhamnetin-pentoside | ✓ | ✓ | ✓ |
| 35 | 28.2 | 625.1211 | C30H26O15 | −2.04 | 463.0881, 301.0361, 178.9991, 151.0033 | Quercetin-caffeoylhexoside | ✓ | ✓ | ✓ |
| 36 | 28.6 | 545.1465 | C30H26O10 | −2.25 | 419.1143, 409.0924, 273.0765, 164.0113, 125.0238, 97.0289 | (Epi)afzelechin-(Epi)afzelechin | ✓ | ✓ | ✓ |
| 37 | 33.0 | 639.1355 | C31H28O15 | −0.91 | 463.0712, 301.0370, 178.9976, 151.0045 | Quercetin-feruloylhexoside | ✓ | ✓ | ✓ |
| 38 | 35.6 | 301.0353 | C15H10O7 | −1.72 | 178.9964, 151.0021 | Quercetin | ✓ | ✓ | |
| 39 | 37.7 | 653.1525 | C32H30O15 | −1.83 | 477.1042, 315.0510, 300.0272, 299.0195 | Isorhamnetin-feruloylhexoside | ✓ | ✓ | ✓ |
| 40 | 38.9 | 327.2184 | C18H32O5 | −2.29 | 291.1993, 229.1430, 211.1325, 171.1020 | Oxo-dihydroxy-octadecenoic acid | ✓ | ✓ | ✓ |
| 41 | 40.4 | 329.2340 | C18H34O5 | −3.00 | 311.2177, 229.1441, 211.1333, 171.1021 | Trihydroxy-octadecenoic acid | ✓ | ✓ | ✓ |
| Samples | DPPH (mgTE/g) | ABTS (mgTE/g) | FRAP (mgTE/g) | CUPRAC (mgTE/g) | Metal Chelating (mgEDTAE/g) | Phosphomolybdenum (mmolTE/g) |
|---|---|---|---|---|---|---|
| RM-EA | 76.43 ± 1.52 b | 438.46 ± 1.69 c | 128.10 ± 1.49 c | 219.81 ± 3.82 | 5.67 ± 0.26 b | 1.78 ± 0.07 c |
| RM-MEOH | 152.28 ± 2.40 a | 482.57 ± 0.89 a | 205.92 ± 7.24 b | 380.14 ± 1.38 | 6.73 ± 0.14 b | 1.98 ± 0.02 b |
| RM-Aq | 154.08 ± 2.43 a | 477.02 ± 1.09 b | 249.40 ± 3.01 a | 384.57 ± 1.99 | 29.68 ± 0.74 a | 2.38 ± 0.07 a |
| Samples | AChE Inhibition (mgGALAE/g) | BchE Inhibition (mgGALAE/g) | Tyrosinase Inhibition (mgKAE/g) | Alpha-Amylase Inhibition (mmolACAE/g) | Alpha-Glucosidase Inhibition (mmolACAE/g) |
|---|---|---|---|---|---|
| RM-EA | 4.94 ± 0.07 a | 6.48 ± 0.62 a | 139.84 ± 0.67 a | 0.85 ± 0.04 a | 1.76 ± 0.02 b |
| RM-MEOH | 5.02 ± 0.06 a | 5.27 ± 0.44 b | 139.08 ± 0.51 a | 0.68 ± 0.03 b | 1.79 ± 0.01 a |
| RM-Aq | na | na | 19.36 ± 2.36 b | 0.08 ± 0.01 c | 1.75 ± 0.01 b |
| Metabolite/Pubchem ID | AChE (PDB:2YDM) | Alpha-Amylase (PDB:1VAH) | Alpha-Glucosidase (PDB:3AXI) | Tyrosinase (PDB:2Y9X) | BChE (PDB:5HF5) |
|---|---|---|---|---|---|
| Binding Energies | |||||
| 2-Isopropylmalic acid/77 | −5.1 | −5.3 | −5.4 | −4.7 | −5.7 |
| Citric acid/311 | −5.5 | −5.3 | −5.5 | −5 | −5.7 |
| Salicylic acid/338 | −5.6 | −4.9 | −5.3 | −6.1 | −6.7 |
| Gallic acid/370 | −5.9 | −5.3 | −5.8 | −5.9 | −7.1 |
| Isocitric acid/1198 | −5.7 | −5.4 | −5.5 | −5.5 | −6.1 |
| Epicatechin/72276 | −8.3 | −6.8 | −8.1 | −6 | −10 |
| (Epi)afzelechin/282014 | −7.8 | −6.2 | −7.4 | −6.5 | −9.7 |
| Quercetin/5280343 | −8.7 | −7 | −8.6 | −6.1 | −10.7 |
| Rutin/5280805 | −8.4 | −8.2 | −3 | −7.2 | −8.8 |
| Isorhamnetin Rutinoside/133562525 | −8.4 | −7.6 | −0.6 | −6.8 | −4.7 |
| Co-crystal to 2ydm (control) | −6.3 | ||||
| Co-crystal to 1Vah (control) | −4.9 | ||||
| Co-crystal to 3axi (control) | −5.6 | ||||
| Co-crystal to 2y9x (control) | −6.1 | ||||
| Co-crystal to 5hf5 (control) | −4.5 | ||||
| Ligand | Receptor | Residue | Interaction | Distance | E (Kcal/mol) |
|---|---|---|---|---|---|
| Alpha-Amylase-Rutin Docking | |||||
| C9 10 | OD1 | ASP 300 (A) | H-donor | 3.53 | −0.6 |
| O14 41 | OD1 | ASP 356 (A) | H-donor | 2.53 | −1.0 |
| O7 43 | O | HIS 305 (A) | H-donor | 3.07 | −1.7 |
| O9 66 | OD1 | ASP 197 (A) | H-donor | 2.95 | −0.9 |
| O9 66 | OD2 | ASP 197 (A) | H-donor | 2.75 | −3.9 |
| O8 68 | OE2 | GLU 233 (A) | H-donor | 2.75 | −4.4 |
| O10 64 | NE2 | HIS 299 (A) | H-acceptor | 2.86 | −3.5 |
| O4 72 | OH | TYR 151 (A) | H-acceptor | 2.77 | −2.1 |
| Alpha-Glucosidase-Quercetin Docking | |||||
| O2 27 | OE2 | GLU 411 (A) | H-donor | 2.53 | −4.8 |
| O3 29 | OD2 | ASP 352 (A) | H-donor | 2.58 | −0.9 |
| O5 31 | OD2 | ASP 307 (A) | H-donor | 2.59 | −4.9 |
| O4 2 | NE2 | GLN 279 (A) | H-acceptor | 2.91 | −1.1 |
| Butyrylcholinesterase-Quercetin Docking | |||||
| O7 25 | O | TYR 72 (A) | H-donor | 2.75 | −2.2 |
| O2 27 | OG | SER 125 (A) | H-donor | 2.78 | −2.6 |
| O3 29 | OE2 | GLU 202 (A) | H-donor | 2.50 | 0.5 |
| O5 31 | O | HIS 447 (A) | H-donor | 2.61 | −2.6 |
| O4 2 | N | GLY 121 (A) | H-acceptor | 2.90 | −2.1 |
| Acetylcholinesterase-Quercetin Docking | |||||
| O7 25 | O | ALA 354 (A) | H-donor | 2.99 | −2.3 |
| O5 31 | OE2 | GLU 376 (A) | H-donor | 2.53 | 0.2 |
| ZN 1619 | OE 1 | GLU 384 (A) | metal | 1.76 | −5.2 |
| ZN 1619 | OE 2 | GLU 384 (A) | metal | 1.75 | −5.2 |
| ZN 1619 | OE1 | GLU 411 (A) | metal | 1.77 | −5.3 |
| ZN 1619 | OE2 | GLU 411 (A) | metal | 1.79 | −5.2 |
| ZN 1619 | OE1 | GLU 384 (A) | ionic | 1.76 | −21.5 |
| ZN 1619 | OE2 | GLU 384 (A) | ionic | 1.75 | −21.6 |
| ZN 1619 | OE1 | GLU 411 (A) | ionic | 1.77 | −21.2 |
| ZN 1619 | OE2 | GLU 411 (A) | ionic | 1.79 | −20.6 |
| 6-ring | NE2 | GLN 281 (A) | pi-H | 4.50 | −1.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suroowan, S.; Llorent-Martínez, E.J.; Zengin, G.; Dall’Acqua, S.; Sut, S.; Buskaran, K.; Fakurazi, S.; Le Van, B.; Abdalla, M.; Abdalla, A.N.; et al. Above the Invasive and Ornamental Attributes of the Traveler’s Palm: An In Vitro and In Silico Insight into the Anti-Oxidant, Anti-Enzymatic, Cytotoxic and Phytochemical Characterization of Ravenala madagascariensis. Antioxidants 2023, 12, 184. https://doi.org/10.3390/antiox12010184
Suroowan S, Llorent-Martínez EJ, Zengin G, Dall’Acqua S, Sut S, Buskaran K, Fakurazi S, Le Van B, Abdalla M, Abdalla AN, et al. Above the Invasive and Ornamental Attributes of the Traveler’s Palm: An In Vitro and In Silico Insight into the Anti-Oxidant, Anti-Enzymatic, Cytotoxic and Phytochemical Characterization of Ravenala madagascariensis. Antioxidants. 2023; 12(1):184. https://doi.org/10.3390/antiox12010184
Chicago/Turabian StyleSuroowan, Shanoo, Eulogio Jose Llorent-Martínez, Gokhan Zengin, Stefano Dall’Acqua, Stefania Sut, Kalaivani Buskaran, Sharida Fakurazi, Bao Le Van, Mohnad Abdalla, Ashraf N. Abdalla, and et al. 2023. "Above the Invasive and Ornamental Attributes of the Traveler’s Palm: An In Vitro and In Silico Insight into the Anti-Oxidant, Anti-Enzymatic, Cytotoxic and Phytochemical Characterization of Ravenala madagascariensis" Antioxidants 12, no. 1: 184. https://doi.org/10.3390/antiox12010184