

Lipid Adaptations against Oxidative Challenge in the Healthy Adult Human Brain
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction

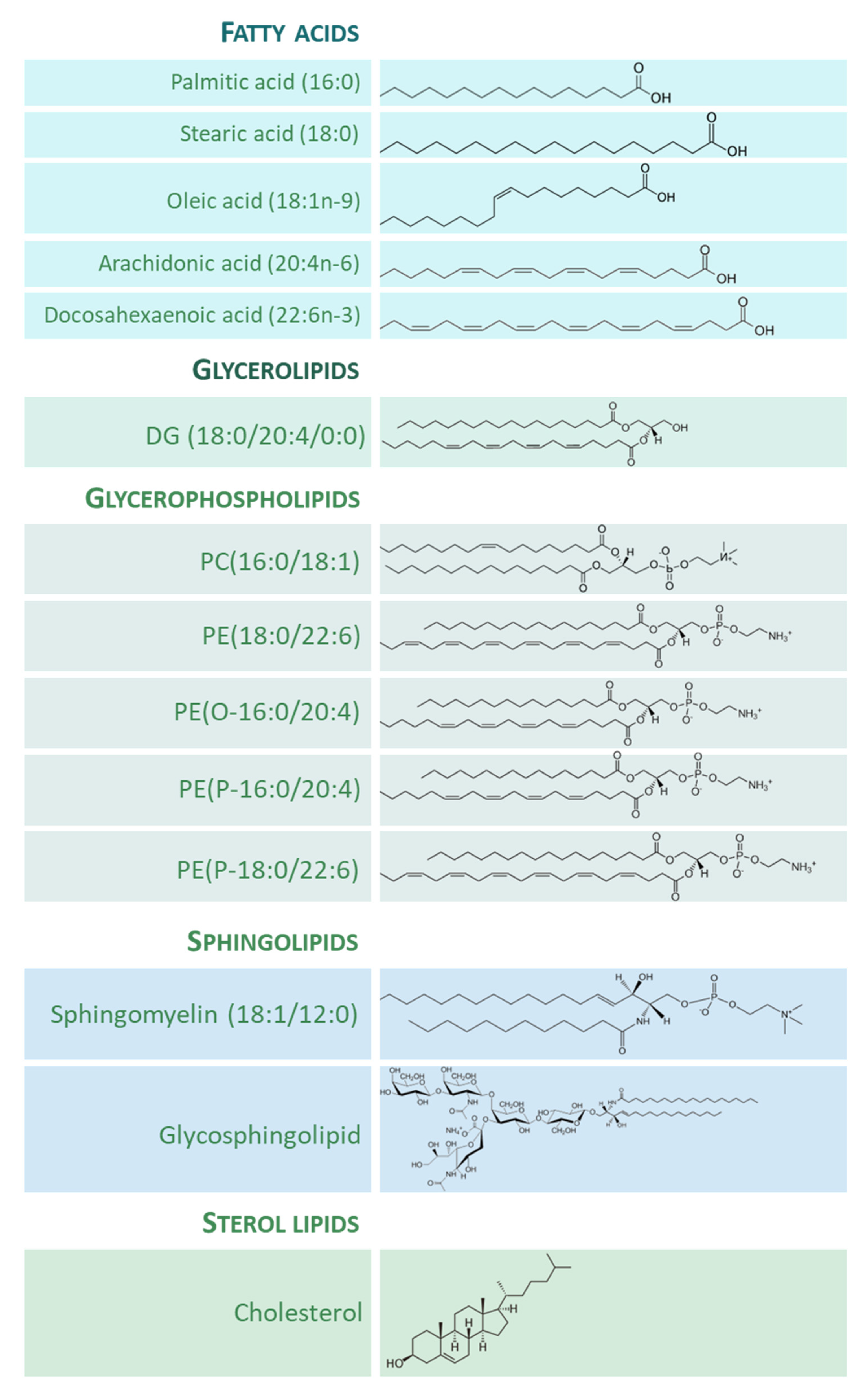
2. Lipids in the Adult whole Human Brain
3. Functional Properties of Lipid Species in the Human Brain
3.1. Integrity of Neural Cell Membranes
3.2. Lipid Signaling in the Human Brain
3.3. Chemical Reactivity of the Acyl Chains in the Human Brain
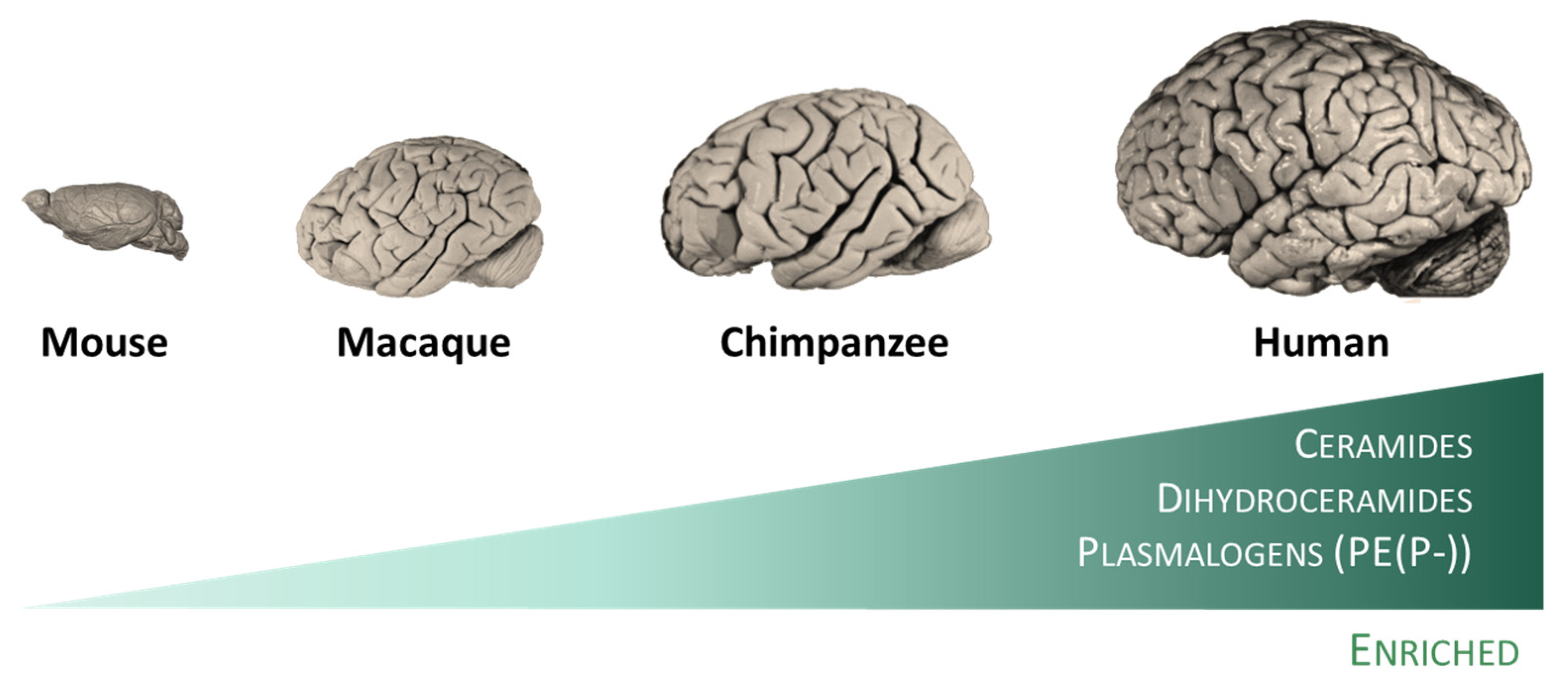
4. Evolution of the Human Brain Lipid Composition
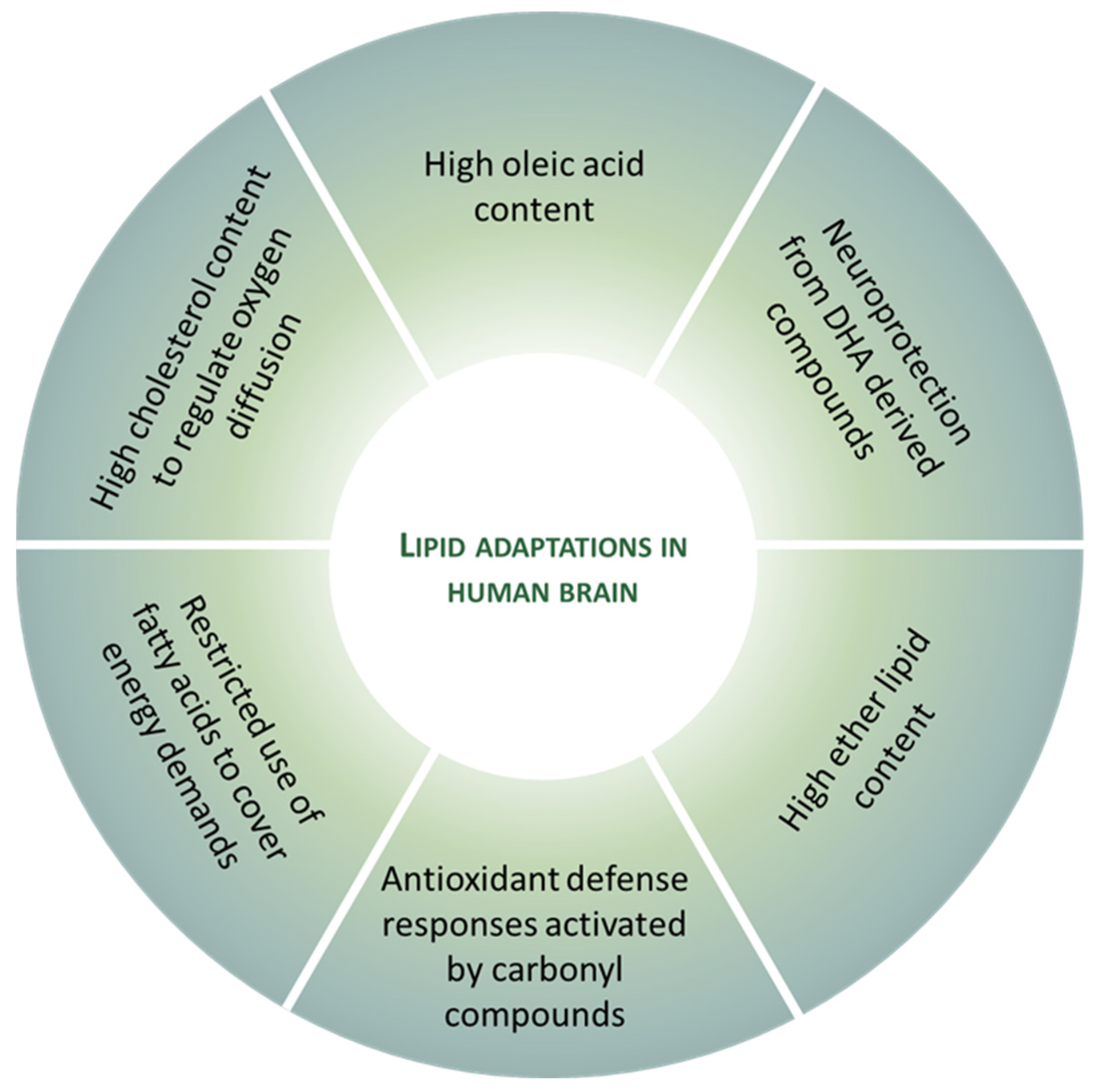
5. Lipid Adaptations against Oxidative Challenge in the Healthy Human Brain
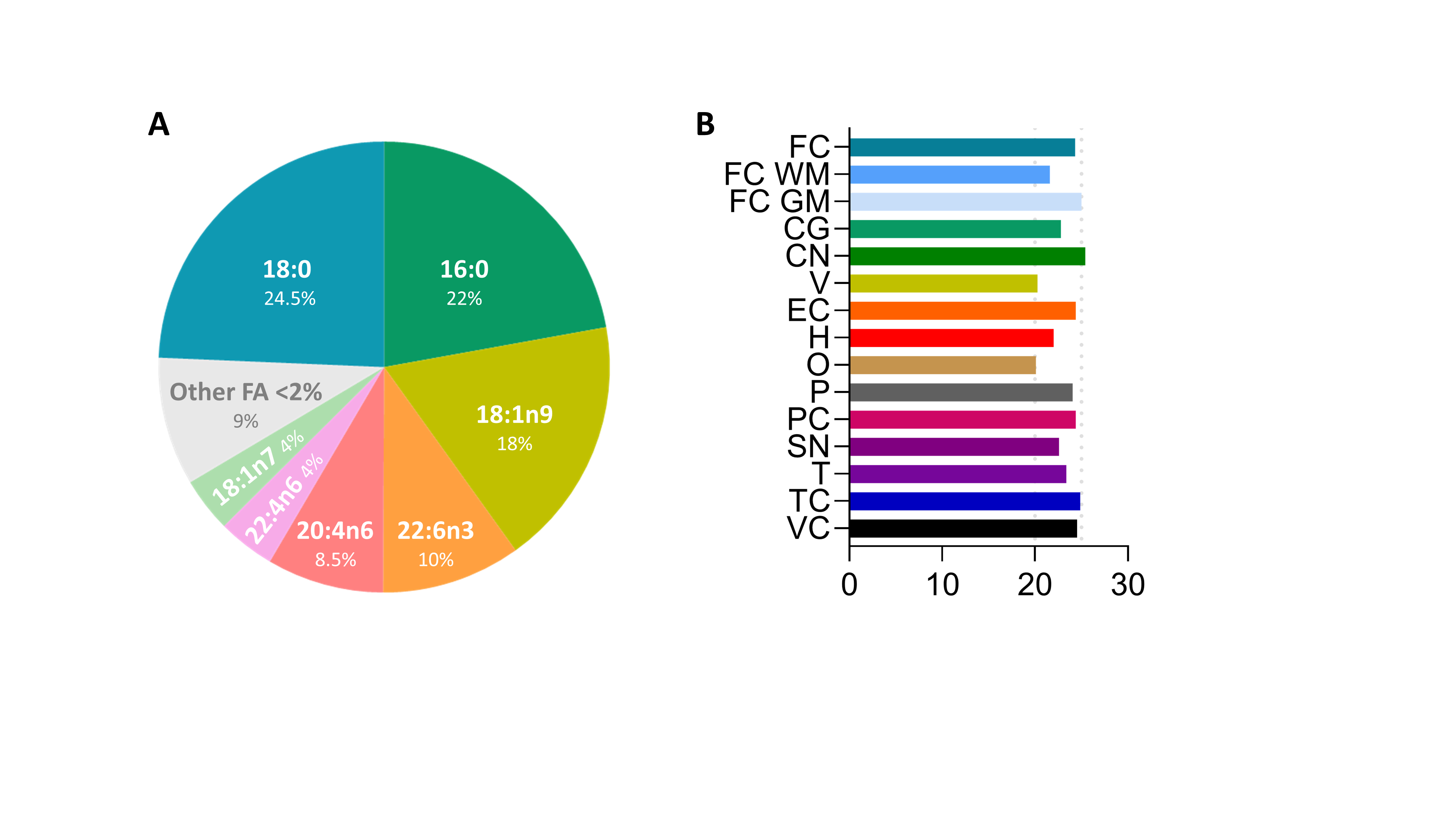
5.1. Oleic Acid Is the Main Fatty Acid in Neural Membranes
5.2. DHA-Derived Compounds Possess a Protective Role against Neural Oxidative Damage
5.3. Ether Lipids as Adaptive Antioxidant System in the Brain
5.4. Carbonyl Compounds Derived from Lipid Peroxidation Are Signaling Compounds which Induce Antioxidant Responses
5.5. The Non-Use of Fatty Acids by Neurons to Cover Energy Demands as Adaptive Non-Oxidant Mechanism
5.6. High Cholesterol Content to Regulate Oxygen Diffusion into Neural Cells
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fahy, E.; Cotter, D.; Sud, M.; Subramaniam, S. Lipid classification, structures and tools. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2011, 1811, 637–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yetukuri, L.; Ekroos, K.; Vidal-Puig, A.; Orešič, M. Informatics and computational strategies for the study of lipids. Mol. Biosyst. 2008, 4, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Han, X. Neurolipidomics: Challenges and developments. Front. Biosci. 2007, 12, 2601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Gross, R.W. The foundations and development of lipidomics. J. Lipid Res. 2022, 63, 100164. [Google Scholar] [CrossRef]
- Fahy, E.; Subramaniam, S.; Murphy, R.C.; Nishijima, M.; Raetz, C.R.H.; Shimizu, T.; Spener, F.; van Meer, G.; Wakelam, M.J.O.; Dennis, E.A. Update of the LIPID MAPS comprehensive classification system for lipids. J. Lipid Res. 2009, 50, S9–S14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramaniam, S.; Fahy, E.; Gupta, S.; Sud, M.; Byrnes, R.W.; Cotter, D.; Dinasarapu, A.R.; Maurya, M.R. Bioinformatics and systems biology of the lipidome. Chem. Rev. 2011, 111, 6452–6490. [Google Scholar] [CrossRef] [Green Version]
- Fahy, E.; Subramaniam, S.; Alex Brown, H.; Glass, C.K.; Merrill, A.H.; Murphy, R.C.; Raetz, C.R.H.; Russell, D.W.; Seyama, Y.; Shaw, W.; et al. A comprehensive classification system for lipids. J. Lipid Res. 2005, 46, 839–861. [Google Scholar] [CrossRef] [Green Version]
- Sastry, P.S. Lipids of nervous tissue: Composition and metabolism. Prog. Lipid Res. 1985, 24, 69–176. [Google Scholar] [CrossRef]
- Thudichum, J.L. A Treatise on the Chemical Constitution of the Brain; Archon Books: Hamden, CT, USA, 1962. [Google Scholar]
- Thudichum, J.L.W. A treatise on the chemical constitution of the brain: Based throughout upon original researches. Glasgow Med. J. 1884, 22, 363–364. [Google Scholar]
- Halliwell, B. Reactive oxygen species and the central nervous system. J. Neurochem. 1992, 59, 1609–1623. [Google Scholar] [CrossRef]
- Cobley, J.N.; Fiorello, M.L.; Miles Bailey, D. 13 reasons why the brain is susceptible to oxidative stress. Redox Biol. 2018, 15, 490–503. [Google Scholar] [CrossRef] [PubMed]
- Naudí, A.; Cabré, R.; Ayala, V.; Jové, M.; Mota-Martorell, N.; Portero-Otín, M.; Pamplona, R. Region-specific vulnerability to lipid peroxidation and evidence of neuronal mechanisms for polyunsaturated fatty acid biosynthesis in the healthy adult human central nervous system. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2017, 1862, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Mota-Martorell, N.; Andrés-Benito, P.; Martín-Gari, M.; Galo-Licona, J.D.; Sol, J.; Fernández-Bernal, A.; Portero-Otín, M.; Ferrer, I.; Jove, M.; Pamplona, R. Selective brain regional changes in lipid profile with human aging. GeroScience 2022, 44, 763–783. [Google Scholar] [CrossRef] [PubMed]
- Merrill, A.H.; Sullards, M.C.; Allegood, J.C.; Kelly, S.; Wang, E. Sphingolipidomics: High-throughput, structure-specific, and quantitative analysis of sphingolipids by liquid chromatography tandem mass spectrometry. Methods 2005, 36, 207–224. [Google Scholar] [CrossRef]
- Dietschy, J.M.; Turley, S.D. Cholesterol metabolism in the brain. Curr. Opin. Lipidol. 2001, 12, 105–112. [Google Scholar] [CrossRef]
- Rouser, G.; Galli, C.; Kritchevsky, G. Lipid class composition of normal human brain and variations in metachromatic leucodystrophy, tay-sachs, niemann-pick, chronic gaucher’s and alzheimer’s diseases. J. Am. Oil Chem. Soc. 1965, 42, 404–410. [Google Scholar] [CrossRef]
- Rouser, G.; Feldman, G.; Galli, C. Fatty acid compositions of human brain lecithin and sphingomyelin in normal individuals, senile cerebral cortical atrophy, alzheimer’s disease, metachromatic leucodystrophy, tay-sachs and niemann-pick diseases. J. Am. Oil Chem. Soc. 1965, 42, 411–412. [Google Scholar] [CrossRef]
- Rouser, G.; Yamamoto, A. Curvilinear regression course of human brain lipid composition changes with age. Lipids 1968, 3, 284–287. [Google Scholar] [CrossRef]
- O’brien, J.S.; Sampson, E.L. Lipid composition of the normal human brain: Gray matter, white matter, and myelin. J. Lipid Res. 1965, 6, 537. [Google Scholar] [CrossRef]
- Panganamala, R.V.; Horrocks, L.A.; Geer, J.C.; Cornwell, D.G. Positions of double bonds in the monounsaturated Alk-1-Enyl groups from the plasmalogens of human heart and brain. Chem. Phys. Lipids 1971, 6, 97–102. [Google Scholar] [CrossRef]
- Kahma, K.; Brotherus, J.; Haltia, M.; Renkonen, O. Low and moderate concentrations of lysobisphosphatidic acid in brain and liver of patients affected by some storage diseases. Lipids 1976, 11, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Maceyka, M.; Spiegel, S. Sphingolipid metabolites in inflammatory disease. Nature 2014, 510, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Sonnino, S.; Chigorno, V. Ganglioside molecular species containing C18- and C20-sphingosine in mammalian nervous tissues and neuronal cell cultures. Biochim. Biophys. Acta-Rev. Biomembr. 2000, 1469, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Piomelli, D.; Astarita, G.; Rapaka, R. A Neuroscientist’s guide to lipidomics. Nat. Rev. Neurosci. 2007, 8, 743–754. [Google Scholar] [CrossRef] [Green Version]
- Piomelli, D. The challenge of brain lipidomics. Prostaglandins Other Lipid Mediat. 2005, 77, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Farooqui, A.A. Lipid mediators in the neural cell nucleus: Their metabolism, signaling, and association with neurological disorders. Neuroscientist 2009, 15, 392–407. [Google Scholar] [CrossRef]
- Farooqui, A.A. Lipid mediators and their metabolism in the nucleus: Implications for alzheimer’s disease. J. Alzheimer’s Dis. 2012, 30, S163–S178. [Google Scholar] [CrossRef]
- Pamplona, R. Membrane phospholipids, lipoxidative damage and molecular integrity: A causal role in aging and longevity. Biochim. Biophys. Acta 2008, 1777, 1249–1262. [Google Scholar] [CrossRef] [Green Version]
- Holman, R.T. Autoxidation of fats and related substances. Prog. Chem. Fats Other Lipids 1954, 2, 51–98. [Google Scholar] [CrossRef]
- Bielski, B.H.J.; Arudi, R.L.; Sutherland, M.W. A study of the reactivity of HO2/O2− with unsaturated fatty acids. J. Biol. Chem. 1983, 258, 4759–4761. [Google Scholar] [CrossRef]
- Yin, H.; Xu, L.; Porter, N.A. Free radical lipid peroxidation: Mechanisms and analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef] [PubMed]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef] [PubMed]
- Catalá, A. Lipid peroxidation of membrane phospholipids generates hydroxy-alkenals and oxidized phospholipids active in physiological and/or pathological conditions. Chem. Phys. Lipids 2009, 157, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fritz, K.S.; Petersen, D.R. An overview of the chemistry and biology of reactive aldehydes. Free Radic. Biol. Med. 2013, 59, 85–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorpe, S.R.; Baynes, J.W. Maillard reaction products in tissue proteins: New products and new perspectives. Amino Acids 2003, 25, 275–281. [Google Scholar] [CrossRef] [PubMed]
- West, J.D.; Marnett, L.J. Endogenous reactive intermediates as modulators of cell signaling and cell death. Chem. Res. Toxicol. 2006, 19, 173–194. [Google Scholar] [CrossRef]
- Naudí, A.; Jové, M.; Ayala, V.; Cabré, R.; Portero-Otín, M.; Pamplona, R. Non-enzymatic modification of aminophospholipids by carbonyl-amine reactions. Int. J. Mol. Sci. 2013, 14, 3285–3313. [Google Scholar] [CrossRef] [Green Version]
- Pamplona, R. Advanced lipoxidation end-products. Chem. Biol. Interact. 2011, 192, 14–20. [Google Scholar] [CrossRef]
- Caro, P.; Gomez, J.; Sanchez, I.; Naudi, A.; Ayala, V.; López-Torres, M.; Pamplona, R.; Barja, G. Forty percent methionine restriction decreases mitochondrial oxygen radical production and leak at complex i during forward electron flow and lowers oxidative damage to proteins and mitochondrial DNA in rat kidney and brain mitochondria. Rejuven. Res. 2009, 12, 421–434. [Google Scholar] [CrossRef]
- Pamplona, R.; Portero-Otín, M.; Sanz, A.; Ayala, V.; Vasileva, E.; Barja, G. Protein and lipid oxidative damage and complex I content are lower in the brain of budgerigar and canaries than in mice. Relation to aging rate. Age 2005, 27, 267–280. [Google Scholar] [CrossRef] [Green Version]
- Naudí, A.; Caro, P.; Jové, M.; Gómez, J.; Boada, J.; Ayala, V.; Portero-Otín, M.; Barja, G.; Pamplona, R. Methionine restriction decreases endogenous oxidative molecular damage and increases mitochondrial biogenesis and uncoupling protein 4 in rat brain. Rejuvena. Res. 2007, 10, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Aviram, R.; Manella, G.; Kopelman, N.; Neufeld-Cohen, A.; Zwighaft, Z.; Elimelech, M.; Adamovich, Y.; Golik, M.; Wang, C.; Han, X.; et al. Lipidomics analyses reveal temporal and spatial lipid organization and uncover daily oscillations in intracellular organelles. Mol. Cell 2016, 62, 636–648. [Google Scholar] [CrossRef]
- Khrameeva, E.; Kurochkin, I.; Bozek, K.; Giavalisco, P.; Khaitovich, P. Lipidome evolution in mammalian tissues. Mol. Biol. Evol. 2018, 35, 1947–1957. [Google Scholar] [CrossRef] [Green Version]
- Pradas, I.; Huynh, K.; Cabré, R.; Ayala, V.; Meikle, P.J.; Jové, M.; Pamplona, R. Lipidomics reveals a tissue-specific fingerprint. Front. Physiol. 2018, 9, 1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jové, M.; Mota-Martorell, N.; Pradas, I.; Galo-Licona, J.D.; Martín-Gari, M.; Obis, È.; Sol, J.; Pamplona, R. The Lipidome Fingerprint of Longevity. Molecules 2020, 25, 4343. [Google Scholar] [CrossRef] [PubMed]
- Bozek, K.; Wei, Y.; Yan, Z.; Liu, X.; Xiong, J.; Sugimoto, M.; Tomita, M.; Pääbo, S.; Sherwood, C.C.; Hof, P.R.; et al. Organization and evolution of brain lipidome revealed by large-scale analysis of human, chimpanzee, macaque, and mouse tissues. Neuron 2015, 85, 695–702. [Google Scholar] [CrossRef] [Green Version]
- Söderberg, M.; Edlund, C.; Kristensson, K.; Dallner, G. Lipid compositions of different regions of the human brain during aging. J. Neurochem. 1990, 54, 415–423. [Google Scholar] [CrossRef]
- Silvius, J. Thermotropic phase transitions of pure lipids in model membranes and their modifications by membrane proteins. Lipid-Protein Interact. 1982, 2, 239–281. [Google Scholar]
- Obis, E.; Sol, J.; Andres-Benito, P.; Martín-Gari, M.; Mota-Martorell, N.; Daniel Galo-Licona, J.; Piñol-Ripoll, G.; Portero-Otin, M.; Ferrer, I.; Jové, M.; et al. Lipidomic alterations in the cerebral cortex and white matter in sporadic alzheimer’s disease. bioRxiv 2022. [Google Scholar] [CrossRef]
- Brenner, R.R. Effect of unsaturated acids on membrane structure and enzyme kinetics. Prog. Lipid Res. 1984, 23, 69–96. [Google Scholar] [CrossRef]
- Hulbert, A.J. The under-appreciated fats of life: The two types of polyunsaturated fats. J. Exp. Biol. 2021, 224, jeb232538. [Google Scholar] [CrossRef] [PubMed]
- Svennerholm, L. Distribution and fatty acid composition of phosphoglycerides in normal human brain. J. Lipid Res. 1968, 9, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Spector, A.A. Plasma free fatty acid and lipoproteins as sources of polyunsaturated fatty acid for the brain. J. Mol. Neurosci. 2001, 16, 159–165. [Google Scholar] [CrossRef]
- Kim, H.Y. Novel metabolism of docosahexaenoic acid in neural cells. J. Biol. Chem. 2007, 282, 18661–18665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.T.; Green, J.T.; Orr, S.K.; Bazinet, R.P. Regulation of brain polyunsaturated fatty acid uptake and turnover. Prostaglandins Leukot. Essent. Fat. Acids 2008, 79, 85–91. [Google Scholar] [CrossRef]
- Bazan, N.G.; Molina, M.F.; Gordon, W.C. Docosahexaenoic acid signalolipidomics in nutrition: Significance in aging, neuroinflammation, macular degeneration, alzheimer’s, and other neurodegenerative diseases. Annu. Rev. Nutr. 2011, 31, 321–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapoport, S.I.; Rao, J.; Igarashi, M. Brain metabolism of nutritionally essential polyunsaturated fatty acids depends on both the diet and the liver. Prostaglandins Leukot. Essent. Fat. Acids 2007, 77, 251–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashima, T. ‘PUFA–GPR40–CREB signaling’ hypothesis for the adult primate neurogenesis. Prog. Lipid Res. 2012, 51, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Brenna, J.T.; Salem, N.; Sinclair, A.J.; Cunnane, S.C. α-Linolenic acid supplementation and conversion to n-3 long-chain polyunsaturated fatty acids in humans. Prostaglandins Leukot. Essent. Fat. Acids 2009, 80, 85–91. [Google Scholar] [CrossRef]
- Crawford, M.A.; Leigh Broadhurst, C.; Guest, M.; Nagar, A.; Wang, Y.; Ghebremeskel, K.; Schmidt, W.F. A quantum theory for the irreplaceable role of docosahexaenoic acid in neural cell signalling throughout evolution. Prostaglandins Leukot. Essent. Fat. Acids 2013, 88, 5–13. [Google Scholar] [CrossRef]
- Casañas-Sánchez, V.; Pérez, J.A.; Fabelo, N.; Quinto-Alemany, D.; Díaz, M.L. Docosahexaenoic (DHA) modulates phospholipid-hydroperoxide glutathione peroxidase (Gpx4) gene expression to ensure self-protection from oxidative damage in hippocampal cells. Front. Physiol. 2015, 6, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz, M.; Mesa-Herrera, F.; Marín, R. DHA and its elaborated modulation of antioxidant defenses of the brain: Implications in aging and AD neurodegeneration. Antioxidants 2021, 10, 907. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, M.; De Oliveira, E.; Odena, M.A.; Portero, M.; Pamplona, R.; Ferrer, I. Redox proteomic profiling of neuroketal-adducted proteins in human brain: Regional vulnerability at middle age increases in the elderly. Free Radic. Biol. Med. 2016, 95, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Dean, J.M.; Lodhi, I.J. Structural and functional roles of ether lipids. Protein Cell 2018, 9, 196–206. [Google Scholar] [CrossRef]
- Yavin, E.; Gatt, S. Oxygen-dependent cleavage of the vinyl-ether linkage of plasmalogens. Eur. J. Biochem. 1972, 25, 437–446. [Google Scholar] [CrossRef]
- Yavin, E.; Gatt, S. Oxygen-dependent cleavage of the vinyl-ether linkage of plasmalogen. Eur. J. Biochem. 1972, 25, 431–436. [Google Scholar] [CrossRef]
- Goldfine, H. The appearance, disappearance and reappearance of plasmalogens in evolution. Prog. Lipid Res. 2010, 49, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Morand, O.H.; Zoeller, R.A.; Raetz, C.R.H. Disappearance of plasmalogens from membranes of animal cells subjected to photosensitized oxidation. J. Biol. Chem. 1988, 263, 11597–11606. [Google Scholar] [CrossRef]
- Khaselev, N.; Murphy, R.C. Susceptibility of plasmenyl glycerophosphoethanolamine lipids containing arachidonate to oxidative degradation. Free Radic. Biol. Med. 1999, 26, 275–284. [Google Scholar] [CrossRef]
- Maeba, R.; Sawada, Y.; Shimasaki, H.; Takahashi, I.; Ueta, N. Ethanolamine plasmalogens protect cholesterol-rich liposomal membranes from oxidation caused by free radicals. Chem. Phys. Lipids 2002, 120, 145–151. [Google Scholar] [CrossRef]
- Skaff, O.; Pattison, D.I.; Davies, M.J. The vinyl ether linkages of plasmalogens are favored targets for myeloperoxidase-derived oxidants: A kinetic study. Biochemistry 2008, 47, 8237–8245. [Google Scholar] [CrossRef] [PubMed]
- Broniec, A.; Klosinski, R.; Pawlak, A.; Wrona-Krol, M.; Thompson, D.; Sarna, T. Interactions of plasmalogens and their diacyl analogs with singlet oxygen in selected model systems. Free Radic. Biol. Med. 2011, 50, 892–898. [Google Scholar] [CrossRef] [PubMed]
- Zoeller, R.A.; Morand, O.H.; Raetz, C.R.H. A possible role for plasmalogens in protecting animal cells against photosensitized killing. J. Biol. Chem. 1988, 263, 11590–11596. [Google Scholar] [CrossRef]
- Reiss, D.; Beyer, K.; Engelmann, B. Delayed oxidative degradation of polyunsaturated diacyl phospholipids in the presence of plasmalogen phospholipids in vitro. Biochem. J. 1997, 323, 807–814. [Google Scholar] [CrossRef] [Green Version]
- Luoma, A.M.; Kuo, F.; Cakici, O.; Crowther, M.N.; Denninger, A.R.; Avila, R.L.; Brites, P.; Kirschner, D.A. Plasmalogen phospholipids protect internodal myelin from oxidative damage. Free Radic. Biol. Med. 2015, 84, 296–310. [Google Scholar] [CrossRef]
- Higdon, A.; Diers, A.R.; Oh, J.Y.; Landar, A.; Darley-Usmar, V.M. Cell signalling by reactive lipid species: New concepts and molecular mechanisms. Biochem. J. 2012, 442, 453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echtay, K.S.; Esteves, T.C.; Pakay, J.L.; Jekabsons, M.B.; Lambert, A.J.; Portero-Otín, M.; Pamplona, R.; Vidal-Puig, A.J.; Wang, S.; Roebuck, S.J.; et al. A signalling role for 4-hydroxy-2-nonenal in regulation of mitochondrial uncoupling. EMBO J. 2003, 22, 4103–4110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand, M.D.; Affourtit, C.; Esteves, T.C.; Green, K.; Lambert, A.J.; Miwa, S.; Pakay, J.L.; Parker, N. Mitochondrial superoxide: Production, biological effects, and activation of uncoupling proteins. Free Radic. Biol. Med. 2004, 37, 755–767. [Google Scholar] [CrossRef]
- Kim-Han, J.S.; Dugan, L.L. Mitochondrial uncoupling proteins in the central nervous system. Antioxid. Redox Signal. 2005, 7, 1173–1181. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Chan, S.L.; de Souza-Pinto, N.C.; Slevin, J.R.; Wersto, R.P.; Zhan, M.; Mustafa, K.; de Cabo, R.; Mattson, M.P. Mitochondrial UCP4 mediates an adaptive shift in energy metabolism and increases the resistance of neurons to metabolic and oxidative stress. Neuromol. Med. 2006, 8, 389–414. [Google Scholar] [CrossRef] [Green Version]
- Ramsden, D.B.; Ho, P.W.L.; Ho, J.W.M.; Liu, H.F.; So, D.H.F.; Tse, H.M.; Chan, K.H.; Ho, S.L. Human neuronal uncoupling proteins 4 and 5 (UCP4 and UCP5): Structural properties, regulation, and physiological role in protection against oxidative stress and mitochondrial dysfunction. Brain Behav. 2012, 2, 468–478. [Google Scholar] [CrossRef]
- Wakabayashi, N.; Dinkova-Kostova, A.T.; Holtzclaw, W.D.; Kang, M.I.; Kobayashi, A.; Yamamoto, M.; Kensler, T.W.; Talalay, P. Protection against electrophile and oxidant stress by induction of the phase 2 response: Fate of cysteines of the keap1 sensor modified by inducers. Proc. Natl. Acad. Sci. USA 2004, 101, 2040. [Google Scholar] [CrossRef]
- Giles, G.I. Redox-controlled transcription factors and gene expression. In Redox Signaling and Regulation in Biology and Medicine; Wiley: Hoboken, NJ, USA, 2009; pp. 245–270. [Google Scholar] [CrossRef]
- Maher, J.; Yamamoto, M. The rise of antioxidant signaling--the evolution and hormetic actions of Nrf2. Toxicol. Appl. Pharmacol. 2010, 244, 4–15. [Google Scholar] [CrossRef]
- Imai, H.; Nakagawa, Y. Biological significance of phospholipid hydroperoxide glutathione peroxidase (PHGPx, GPx4) in mammalian cells. Free Radic. Biol. Med. 2003, 34, 145–169. [Google Scholar] [CrossRef]
- Brigelius-Flohé, R. Glutathione peroxidases and redox-regulated transcription factors. Biol. Chem. 2006, 387, 1329–1335. [Google Scholar] [CrossRef]
- Conrad, M.; Schneider, M.; Seiler, A.; Bornkamm, G.W. Physiological role of phospholipid hydroperoxide glutathione peroxidase in mammals. Biol. Chem. 2007, 388, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, J.; Buckingham, J.A.; Roebuck, S.J.; Brand, M.D. Topology of superoxide production from different sites in the mitochondrial electron transport chain. J. Biol. Chem. 2002, 277, 44784–44790. [Google Scholar] [CrossRef] [Green Version]
- Lambertucci, R.H.; Hirabara, S.M.; Silveira, L.D.R.; Levada-Pires, A.C.; Curi, R.; Pithon-Curi, T.C. Palmitate increases superoxide production through mitochondrial electron transport chain and NADPH oxidase activity in skeletal muscle cells. J. Cell. Physiol. 2008, 216, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Schönfeld, P.; Reiser, G. Why does brain metabolism not favor burning of fatty acids to provide energy?-Reflections on disadvantages of the use of free fatty acids as fuel for brain. J. Cereb. Blood Flow Metab. 2013, 33, 1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonvento, G.; Bolaños, J.P. Astrocyte-neuron metabolic cooperation shapes brain activity. Cell Metab. 2021, 33, 1546–1564. [Google Scholar] [CrossRef] [PubMed]
- Schönfeld, P.; Reiser, G. How the brain fights fatty acids’ toxicity. Neurochem. Int. 2021, 148, 105050. [Google Scholar] [CrossRef] [PubMed]
- Galea, A.M.; Brown, A.J. Special relationship between sterols and oxygen: Were sterols an adaptation to aerobic life? Free Radic. Biol. Med. 2009, 47, 880–889. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Galea, A.M. Cholesterol as an evolutionary response to living with oxygen. Evolution 2010, 64, 2179–2183. [Google Scholar] [CrossRef] [PubMed]
- Zuniga-Hertz, J.P.; Patel, H.H. The evolution of cholesterol-rich membrane in oxygen adaption: The respiratory system as a model. Front. Physiol. 2019, 10, 1340. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological System | Animal Species | Concentration | Reference |
|---|---|---|---|
| Brain mitochondria | Rat | 571 ± 30 | [40] |
| Whole brain | Mouse | 374 ± 23 | [41] |
| Whole brain | Parakeet | 305 ± 27 | [41] |
| Whole brain | Canary | 259 ± 22 | [41] |
| Whole brain | Rat | 337 ± 18 | [42] |
| Amygdala | Human | 431 ± 32 | [42] |
| Cerebellum | Human | 203 ± 20 | [42] |
| Entorhinal cortex | Human | 283 ± 28 | [42] |
| Frontal cortex | Human | 185 ± 12 | [42] |
| Hippocampus | Human | 221 ± 25 | [42] |
| Medulla oblongata | Human | 340 ± 19 | [42] |
| Occipital cortex | Human | 219 ± 16 | [42] |
| Spinal cord | Human | 352 ± 11 | [42] |
| Striatum | Human | 450 ± 52 | [42] |
| Substantia nigra | Human | 590 ± 29 | [42] |
| Temporal cortex | Human | 164 ± 9 | [42] |
| Thalamus | Human | 481 ± 42 | [42] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jové, M.; Mota-Martorell, N.; Obis, È.; Sol, J.; Martín-Garí, M.; Ferrer, I.; Portero-Otín, M.; Pamplona, R. Lipid Adaptations against Oxidative Challenge in the Healthy Adult Human Brain. Antioxidants 2023, 12, 177. https://doi.org/10.3390/antiox12010177
Jové M, Mota-Martorell N, Obis È, Sol J, Martín-Garí M, Ferrer I, Portero-Otín M, Pamplona R. Lipid Adaptations against Oxidative Challenge in the Healthy Adult Human Brain. Antioxidants. 2023; 12(1):177. https://doi.org/10.3390/antiox12010177
Chicago/Turabian StyleJové, Mariona, Natàlia Mota-Martorell, Èlia Obis, Joaquim Sol, Meritxell Martín-Garí, Isidre Ferrer, Manuel Portero-Otín, and Reinald Pamplona. 2023. "Lipid Adaptations against Oxidative Challenge in the Healthy Adult Human Brain" Antioxidants 12, no. 1: 177. https://doi.org/10.3390/antiox12010177