
Potential Role of Oxidative Stress in the Production of Volatile Organic Compounds in Obesity
 ,
,
Abstract
:1. Introduction
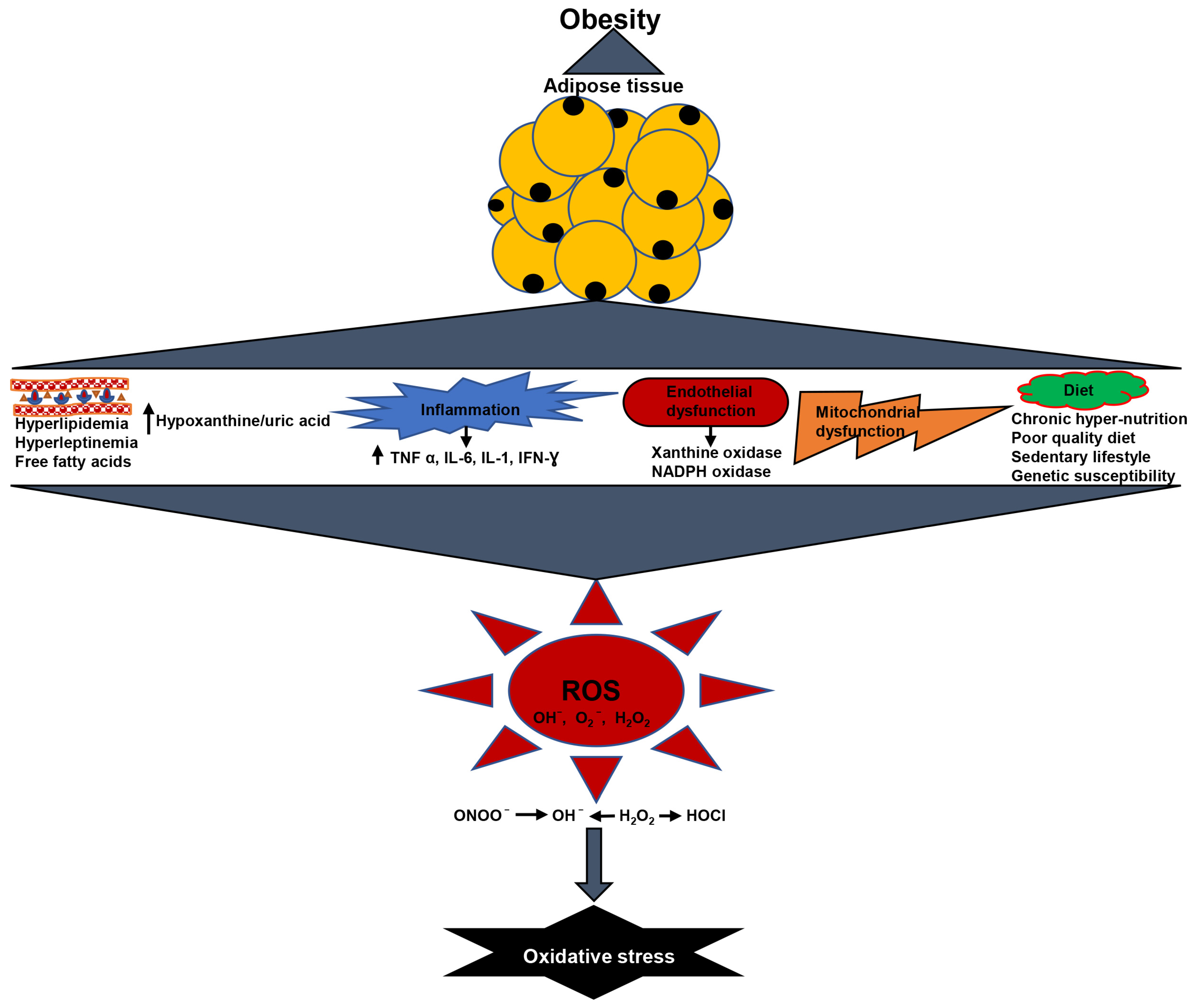
2. Oxidative Stress in Obesity
2.1. Obesity Is Associated with Elevated Lipid Levels and Oxidative Stress
2.2. Chronic Low-Grade Inflammation Produced Oxidative Stress in Obesity
2.3. Obesity Causes Increased Hypoxanthine/Uric Acid and Oxidative Stress
2.4. Obesity Causes Endothelial Dysfunction and Oxidative Stress
2.5. Obesity Causes Mitochondrial Dysfunction and Oxidative Stress
2.6. Diet and Oxidative Stress in Obesity
3. Volatile Organic Compounds in Obesity
3.1. Volatile Organic Compounds in Biological Samples
3.2. The Biochemical Pathway of VOCs
3.3. Analytical Techniques for VOCs Detection
4. Interconnection of Volatile Organic Compounds and Oxidative Stress in Obesity
Potential Role of Oxidative Stress-Generated VOC in Obesity-Related Problem
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, Z.Y.; Wang, M.W. Obesity, a health burden of a global nature. Acta Pharmacol. Sin. 2012, 33, 145–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pi-Sunyer, F.X. Health implications of obesity. Am. J. Clin. Nutr. 1991, 53, 1595S–1603S. [Google Scholar] [CrossRef] [PubMed]
- Despres, J.P.; Moorjani, S.; Lupien, P.J.; Tremblay, A.; Nadeau, A.; Bouchard, C. Regional distribution of body fat, plasma lipoproteins, and cardiovascular disease. Arteriosclerosis 1990, 10, 497–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, Á.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, Oxidative Stress, and Obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef] [Green Version]
- Calenic, B.; Miricescu, D.; Greabu, M.; Kuznetsov, A.V.; Troppmair, J.; Ruzsanyi, V.; Amann, A. Oxidative stress and volatile organic compounds: Interplay in pulmonary, cardio-vascular, digestive tract systems and cancer. Open Chem. 2015, 13, 1020–1030. [Google Scholar] [CrossRef]
- de Lacy Costello, B.; Amann, A.; Al-Kateb, H.; Flynn, C.; Filipiak, W.; Khalid, T.; Osborne, D.; Ratcliffe, N.M. A review of the volatiles from the healthy human body. J. Breath Res. 2014, 8, 014001. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.; Preti, G. Challenges in the Investigation of Volatile Disease Biomarkers in Urine. Volatile Biomark. 2013, 394–404. [Google Scholar] [CrossRef]
- Buljubasic, F.; Buchbauer, G. scent of human diseases: A review on specific volatile organic compounds as diagnostic biomarkers. Flavour Fragr. J. 2015, 30, 5–25. [Google Scholar] [CrossRef]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef] [Green Version]
- Satapati, S.; Sunny, N.E.; Kucejova, B.; Fu, X.; He, T.T.; Mendez-Lucas, A.; Shelton, J.M.; Perales, J.C.; Browning, J.D.; Burgess, S.C. Elevated TCA cycle function in the pathology of diet-induced hepatic insulin resistance and fatty liver. J. Lipid Res. 2012, 53, 1080–1092. [Google Scholar] [CrossRef]
- Pennathur, S.; Heinecke, J.W. Mechanisms of oxidative stress in diabetes: Implications for the pathogenesis of vascular disease and antioxidant therapy. Front. Biosci. 2004, 9, 565–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [Green Version]
- Egan, B.M.; Greene, E.L.; Goodfriend, T.L. Insulin resistance and cardiovascular disease. Am. J. Hypertens. 2001, 14, 116S–125S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Meco, M.T.; Moscat, J. The atypical PKCs in inflammation: NF-kappaB and beyond. Immunol. Rev. 2012, 246, 154–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piperi, C.; Adamopoulos, C.; Dalagiorgou, G.; Diamanti-Kandarakis, E.; Papavassiliou, A.G. Crosstalk between advanced glycation and endoplasmic reticulum stress: Emerging therapeutic targeting for metabolic diseases. J. Clin. Endocrinol. Metab. 2012, 97, 2231–2242. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Schmeisser, A.; Garlichs, C.D.; Plotze, K.; Damme, U.; Mugge, A.; Daniel, W.G. Angiotensin II-induced superoxide anion generation in human vascular endothelial cells: Role of membrane-bound NADH-/NADPH-oxidases. Cardiovasc. Res. 1999, 44, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Aronson, D.; Rayfield, E.J. How hyperglycemia promotes atherosclerosis: Molecular mechanisms. Cardiovasc. Diabetol. 2002, 1, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baynes, J.W. Role of oxidative stress in development of complications in diabetes. Diabetes 1991, 40, 405–412. [Google Scholar] [CrossRef]
- Bondia-Pons, I.; Ryan, L.; Martinez, J.A. Oxidative stress and inflammation interactions in human obesity. J. Physiol. Biochem. 2012, 68, 701–711. [Google Scholar] [CrossRef]
- Bakker, S.J.; RG, I.J.; Teerlink, T.; Westerhoff, H.V.; Gans, R.O.; Heine, R.J. Cytosolic triglycerides and oxidative stress in central obesity: The missing link between excessive atherosclerosis, endothelial dysfunction, and beta-cell failure? Atherosclerosis 2000, 148, 17–21. [Google Scholar] [CrossRef]
- Inoguchi, T.; Li, P.; Umeda, F.; Yu, H.Y.; Kakimoto, M.; Imamura, M.; Aoki, T.; Etoh, T.; Hashimoto, T.; Naruse, M.; et al. High glucose level and free fatty acid stimulate reactive oxygen species production through protein kinase C--dependent activation of NAD(P)H oxidase in cultured vascular cells. Diabetes 2000, 49, 1939–1945. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.P.; Gastaldi, G.; Bobbioni-Harsch, E.; Arboit, P.; Gobelet, C.; Deriaz, O.; Golay, A.; Witztum, J.L.; Giacobino, J.P. Lipid peroxidation in skeletal muscle of obese as compared to endurance-trained humans: A case of good vs. bad lipids? FEBS Lett. 2003, 551, 104–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, H.K.; Powers, S.K.; Dirks, A.J.; Scarpace, P.J. Mechanism for obesity-induced increase in myocardial lipid peroxidation. Int. J. Obes. Relat. Metab. Disord. 2001, 25, 378–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, S.; Riserus, U.; Turpeinen, A.; Vessby, B. Conjugated linoleic acid induces lipid peroxidation in men with abdominal obesity. Clin. Sci. 2000, 99, 511–516. [Google Scholar] [CrossRef]
- Fonseca-Alaniz, M.H.; Takada, J.; Alonso-Vale, M.I.; Lima, F.B. Adipose tissue as an endocrine organ: From theory to practice. J. Pediatr. (Rio J.) 2007, 83, S192–S203. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.J.; McAllister, M.J.; Slusher, A.L.; Webb, H.E.; Mock, J.T.; Acevedo, E.O. Obesity-Related Oxidative Stress: The Impact of Physical Activity and Diet Manipulation. Sports Med. Open 2015, 1, 32. [Google Scholar] [CrossRef] [Green Version]
- Beltowski, J.; Wojcicka, G.; Jamroz, A. Leptin decreases plasma paraoxonase 1 (PON1) activity and induces oxidative stress: The possible novel mechanism for proatherogenic effect of chronic hyperleptinemia. Atherosclerosis 2003, 170, 21–29. [Google Scholar] [CrossRef]
- Parola, M.; Marra, F. Adipokines and redox signaling: Impact on fatty liver disease. Antioxid. Redox Signal. 2011, 15, 461–483. [Google Scholar] [CrossRef] [Green Version]
- German, A.J.; Ryan, V.H.; German, A.C.; Wood, I.S.; Trayhurn, P. Obesity, its associated disorders and the role of inflammatory adipokines in companion animals. Vet. J. 2010, 185, 4–9. [Google Scholar] [CrossRef]
- Maingrette, F.; Renier, G. Leptin increases lipoprotein lipase secretion by macrophages: Involvement of oxidative stress and protein kinase C. Diabetes 2003, 52, 2121–2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saiki, S.; Sato, T.; Kohzuki, M.; Kamimoto, M.; Yosida, T. Changes in serum hypoxanthine levels by exercise in obese subjects. Metabolism 2001, 50, 627–630. [Google Scholar] [CrossRef] [PubMed]
- Sorlie, D.; Myhre, K.; Saugstad, O.D.; Giercksky, K.E. Release of hypoxanthine and phosphate from exercising human legs with and without arterial insufficiency. Acta Med. Scand. 1982, 211, 281–286. [Google Scholar] [CrossRef]
- Sahlin, K.; Ekberg, K.; Cizinsky, S. Changes in plasma hypoxanthine and free radical markers during exercise in man. Acta Physiol. Scand. 1991, 142, 275–281. [Google Scholar] [CrossRef]
- Harkness, R.A.; Simmonds, R.J.; Coade, S.B. Purine transport and metabolism in man: The effect of exercise on concentrations of purine bases, nucleosides and nucleotides in plasma, urine, leucocytes and erythrocytes. Clin. Sci. 1983, 64, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellsten-Westing, Y.; Sollevi, A.; Sjodin, B. Plasma accumulation of hypoxanthine, uric acid and creatine kinase following exhausting runs of differing durations in man. Eur. J. Appl. Physiol. Occup. Physiol. 1991, 62, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.L. Exercise, oxidative stress, and antioxidants. Am. J. Sports Med. 1996, 24, S20–S24. [Google Scholar] [CrossRef] [PubMed]
- Vincent, H.K.; Taylor, A.G. Biomarkers and potential mechanisms of obesity-induced oxidant stress in humans. Int. J. Obes. 2006, 30, 400–418. [Google Scholar] [CrossRef] [Green Version]
- Wolin, M.S.; Ahmad, M.; Gupte, S.A. The sources of oxidative stress in the vessel wall. Kidney Int. 2005, 67, 1659–1661. [Google Scholar] [CrossRef] [Green Version]
- Konior, A.; Schramm, A.; Czesnikiewicz-Guzik, M.; Guzik, T.J. NADPH oxidases in vascular pathology. Antioxid. Redox Signal. 2014, 20, 2794–2814. [Google Scholar] [CrossRef]
- Kaminski, K.A.; Bonda, T.A.; Korecki, J.; Musial, W.J. Oxidative stress and neutrophil activation--the two keystones of ischemia/reperfusion injury. Int. J. Cardiol. 2002, 86, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Wheatcroft, S.B.; Williams, I.L.; Shah, A.M.; Kearney, M.T. Pathophysiological implications of insulin resistance on vascular endothelial function. Diabet. Med. 2003, 20, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Harrison, D.G. Endothelial dysfunction in cardiovascular diseases: The role of oxidant stress. Circ. Res. 2000, 87, 840–844. [Google Scholar] [CrossRef] [Green Version]
- Schiffrin, E.L.; Canadian Institutes of Health Research Multidisciplinary Research Group on Hypertension. Beyond blood pressure: The endothelium and atherosclerosis progression. Am. J. Hypertens. 2002, 15, 115S–122S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dandona, P.; Kumar, V.; Aljada, A.; Ghanim, H.; Syed, T.; Hofmayer, D.; Mohanty, P.; Tripathy, D.; Garg, R. Angiotensin II receptor blocker valsartan suppresses reactive oxygen species generation in leukocytes, nuclear factor-kappa B, in mononuclear cells of normal subjects: Evidence of an antiinflammatory action. J. Clin. Endocrinol. Metab. 2003, 88, 4496–4501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frisbee, J.C.; Maier, K.G.; Stepp, D.W. Oxidant stress-induced increase in myogenic activation of skeletal muscle resistance arteries in obese Zucker rats. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H2160–H2168. [Google Scholar] [CrossRef] [Green Version]
- de Mello, A.H.; Costa, A.B.; Engel, J.D.G.; Rezin, G.T. Mitochondrial dysfunction in obesity. Life Sci. 2018, 192, 26–32. [Google Scholar] [CrossRef]
- Bogacka, I.; Xie, H.; Bray, G.A.; Smith, S.R. Pioglitazone induces mitochondrial biogenesis in human subcutaneous adipose tissue in vivo. Diabetes 2005, 54, 1392–1399. [Google Scholar] [CrossRef] [Green Version]
- Martinez, J.A. Mitochondrial oxidative stress and inflammation: An slalom to obesity and insulin resistance. J. Physiol. Biochem. 2006, 62, 303–306. [Google Scholar] [CrossRef]
- Patel, C.; Ghanim, H.; Ravishankar, S.; Sia, C.L.; Viswanathan, P.; Mohanty, P.; Dandona, P. Prolonged reactive oxygen species generation and nuclear factor-kappaB activation after a high-fat, high-carbohydrate meal in the obese. J. Clin. Endocrinol. Metab. 2007, 92, 4476–4479. [Google Scholar] [CrossRef]
- Vincent, H.K.; Bourguignon, C.M.; Taylor, A.G. Relationship of the dietary phytochemical index to weight gain, oxidative stress and inflammation in overweight young adults. J. Hum. Nutr. Diet. 2010, 23, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Kneepkens, C.M.; Lepage, G.; Roy, C.C. The potential of the hydrocarbon breath test as a measure of lipid peroxidation. Free Radic. Biol. Med. 1994, 17, 127–160. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.; Cataneo, R.N.; Greenberg, J.; Gunawardena, R.; Naidu, A.; Rahbari-Oskoui, F. Effect of age on the breath methylated alkane contour, a display of apparent new markers of oxidative stress. J. Lab. Clin. Med. 2000, 136, 243–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cozzolino, R.; De Giulio, B.; Marena, P.; Martignetti, A.; Gunther, K.; Lauria, F.; Russo, P.; Stocchero, M.; Siani, A. Urinary volatile organic compounds in overweight compared to normal-weight children: Results from the Italian I.Family cohort. Sci. Rep. 2017, 7, 15636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kistler, M.; Muntean, A.; Szymczak, W.; Rink, N.; Fuchs, H.; Gailus-Durner, V.; Wurst, W.; Hoeschen, C.; Klingenspor, M.; Hrabe de Angelis, M.; et al. Diet-induced and mono-genetic obesity alter volatile organic compound signature in mice. J. Breath Res. 2016, 10, 016009. [Google Scholar] [CrossRef] [Green Version]
- Kistler, M.; Szymczak, W.; Fedrigo, M.; Fiamoncini, J.; Hollriegl, V.; Hoeschen, C.; Klingenspor, M.; Hrabe de Angelis, M.; Rozman, J. Effects of diet-matrix on volatile organic compounds in breath in diet-induced obese mice. J. Breath Res. 2014, 8, 016004. [Google Scholar] [CrossRef] [PubMed]
- Alkhouri, N.; Eng, K.; Cikach, F.; Patel, N.; Yan, C.; Brindle, A.; Rome, E.; Hanouneh, I.; Grove, D.; Lopez, R.; et al. Breathprints of childhood obesity: Changes in volatile organic compounds in obese children compared with lean controls. Pediatr. Obes. 2015, 10, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Alkhouri, N.; Cikach, F.; Eng, K.; Moses, J.; Patel, N.; Yan, C.; Hanouneh, I.; Grove, D.; Lopez, R.; Dweik, R. Analysis of breath volatile organic compounds as a noninvasive tool to diagnose nonalcoholic fatty liver disease in children. Eur. J. Gastroenterol. Hepatol. 2014, 26, 82–87. [Google Scholar] [CrossRef]
- Uchikawa, M.; Kato, M.; Nagata, A.; Sanada, S.; Yoshikawa, Y.; Tsunematsu, Y.; Sato, M.; Suzuki, T.; Hashidume, T.; Watanabe, K.; et al. Elevated levels of proinflammatory volatile metabolites in feces of high fat diet fed KK-A(y) mice. Sci. Rep. 2020, 10, 5681. [Google Scholar] [CrossRef] [Green Version]
- Klemenz, A.C.; Meyer, J.; Ekat, K.; Bartels, J.; Traxler, S.; Schubert, J.K.; Kamp, G.; Miekisch, W.; Peters, K. Differences in the Emission of Volatile Organic Compounds (VOCs) between Non-Differentiating and Adipogenically Differentiating Mesenchymal Stromal/Stem Cells from Human Adipose Tissue. Cells 2019, 8, 697. [Google Scholar] [CrossRef]
- Dragonieri, S.; Porcelli, F.; Longobardi, F.; Carratu, P.; Aliani, M.; Ventura, V.A.; Tutino, M.; Quaranta, V.N.; Resta, O.; de Gennaro, G. An electronic nose in the discrimination of obese patients with and without obstructive sleep apnoea. J. Breath Res. 2015, 9, 026005. [Google Scholar] [CrossRef]
- Rowan, D.D. Volatile metabolites. Metabolites 2011, 1, 41–63. [Google Scholar] [CrossRef] [Green Version]
- Longo, V.; Forleo, A.; Provenzano, S.P.; Coppola, L.; Zara, V.; Ferramosca, A.; Siciliano, P.; Capone, S. HS-SPME-GC-MS metabolomics approach for sperm quality evaluation by semen volatile organic compounds (VOCs) analysis. Biomed. Phys. Eng. Express 2018, 5, 015006. [Google Scholar] [CrossRef]
- Baranska, A.; Smolinska, A.; Boots, A.W.; Dallinga, J.W.; van Schooten, F.J. Dynamic collection and analysis of volatile organic compounds from the headspace of cell cultures. J. Breath Res. 2015, 9, 047102. [Google Scholar] [CrossRef] [PubMed]
- Sohrabi. Mohsen; Zhang. Li; Zhang. Kai; Ahmetagic. Adnan; Wei. Ming, Q. Volatile Organic Compounds as Novel Markers for the Detection of Bacterial Clin. Microbial. 2014, 3, 151. [CrossRef] [Green Version]
- Forleo, A.; Capone, S.; Longo, V.; Casino, F.; Radogna, A.V.; Siciliano, P.; Massaro, M.; Scoditti, E.; Calabriso, N.; Carluccio, M.A. Evaluation of the Volatile Organic Compounds Released from Peripheral Blood Mononuclear Cells and THP1 Cells Under Normal and Proinflammatory Conditions. In Proceedings of the AISEM Annual Conference on Sensors and Microsystems 2017, Lecce, Italy, 21–23 February; pp. 269–277.
- Khalid, T.; Aggio, R.; White, P.; De Lacy Costello, B.; Persad, R.; Al-Kateb, H.; Jones, P.; Probert, C.S.; Ratcliffe, N. Urinary Volatile Organic Compounds for the Detection of Prostate Cancer. PLoS ONE 2015, 10, e0143283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thriumani, R.; Zakaria, A.; Hashim, Y.Z.H.; Jeffree, A.I.; Helmy, K.M.; Kamarudin, L.M.; Omar, M.I.; Shakaff, A.Y.M.; Adom, A.H.; Persaud, K.C. A study on volatile organic compounds emitted by in-vitro lung cancer cultured cells using gas sensor array and SPME-GCMS. BMC Cancer 2018, 18, 362. [Google Scholar] [CrossRef]
- Ambrosone, C.B. Oxidants and antioxidants in breast cancer. Antioxid. Redox Signal. 2000, 2, 903–917. [Google Scholar] [CrossRef]
- Hakim, M.; Broza, Y.Y.; Barash, O.; Peled, N.; Phillips, M.; Amann, A.; Haick, H. Volatile organic compounds of lung cancer and possible biochemical pathways. Chem. Rev. 2012, 112, 5949–5966. [Google Scholar] [CrossRef]
- Watanabe, M. Polymorphic CYP genes and disease predisposition--what have the studies shown so far? Toxicol. Lett. 1998, 102-103, 167–171. [Google Scholar] [CrossRef]
- Murray, G.I. The role of cytochrome P450 in tumour development and progression and its potential in therapy. J. Pathol. 2000, 192, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Chen, S. Aromatase and breast cancer. Front. Biosci. 1998, 3, d922–d933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terelius, Y.; Ingelman-Sundberg, M. Metabolism of n-pentane by ethanol-inducible cytochrome P-450 in liver microsomes and reconstituted membranes. Eur. J. Biochem. 1986, 161, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Kohlmuller, D.; Kochen, W. Is n-pentane really an index of lipid peroxidation in humans and animals? A methodological reevaluation. Anal. Biochem. 1993, 210, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Risby, T.H.; Sehnert, S.S. Clinical application of breath biomarkers of oxidative stress status. Free Radic. Biol. Med. 1999, 27, 1182–1192. [Google Scholar] [CrossRef] [PubMed]
- Marchitti, S.A.; Brocker, C.; Stagos, D.; Vasiliou, V. Non-P450 aldehyde oxidizing enzymes: The aldehyde dehydrogenase superfamily. Expert Opin. Drug Metab. Toxicol. 2008, 4, 697–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, I.; van Schadewijk, A.A.; Crowther, A.J.; Hiemstra, P.S.; Stolk, J.; MacNee, W.; De Boer, W.I. 4-Hydroxy-2-nonenal, a specific lipid peroxidation product, is elevated in lungs of patients with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2002, 166, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Vaz, A.D.; Coon, M.J. Hydrocarbon formation in the reductive cleavage of hydroperoxides by cytochrome P-450. Proc. Natl. Acad. Sci. USA 1987, 84, 1172–1176. [Google Scholar] [CrossRef] [Green Version]
- Buszewski, B.; Kesy, M.; Ligor, T.; Amann, A. Human exhaled air analytics: Biomarkers of diseases. Biomed. Chromatogr. 2007, 21, 553–566. [Google Scholar] [CrossRef]
- Gordon, S.M.; Szidon, J.P.; Krotoszynski, B.K.; Gibbons, R.D.; O’Neill, H.J. Volatile organic compounds in exhaled air from patients with lung cancer. Clin. Chem. 1985, 31, 1278–1282. [Google Scholar] [CrossRef]
- Hakim, M.; Billan, S.; Tisch, U.; Peng, G.; Dvrokind, I.; Marom, O.; Abdah-Bortnyak, R.; Kuten, A.; Haick, H. Diagnosis of head-and-neck cancer from exhaled breath. Br. J. Cancer 2011, 104, 1649–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, I.; Lazar, Z.; Gyulai, N.; Kollai, M.; Losonczy, G. Exhaled biomarkers in lung cancer. Eur. Respir. J. 2009, 34, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Branton, P.J.; McAdam, K.G.; Winter, D.B.; Liu, C.; Duke, M.G.; Proctor, C.J. Reduction of aldehydes and hydrogen cyanide yields in mainstream cigarette smoke using an amine functionalised ion exchange resin. Chem. Cent. J. 2011, 5, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahotupa, M.; Bussacchini-Griot, V.; Bereziat, J.C.; Camus, A.M.; Bartsch, H. Rapid oxidative stress induced by N-nitrosamines. Biochem. Biophys. Res. Commun. 1987, 146, 1047–1054. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.O.; Slater, G.; Aufses, A.H., Jr.; Cohen, G. Production of ethane by rats treated with the colon carcinogen, 1,2-dimethylhydrazine. Biochem. Pharmacol. 1988, 37, 2967–2971. [Google Scholar] [CrossRef]
- Burdock, G.; Fenaroli, G. Fenaroli’s Handbook of Flavor Ingredients.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2010. [Google Scholar]
- Yannai, S. Dictionary of Food Compounds with CD-ROM: Additives, Flavors, and Ingredients; Chapman & Hall/CRC: Boca Raton, FL, USA, 2004. [Google Scholar]
- Murray, R.; Granner, D.; Mayes, P.; Rodwell, V. Harper’s Illustrated Biochemistry; McGrawHill Medical: New York, NY, USA, 2006. [Google Scholar]
- Smith, D.; Wang, T.; Spanel, P. On-line, simultaneous quantification of ethanol, some metabolites and water vapour in breath following the ingestion of alcohol. Physiol. Meas. 2002, 23, 477–489. [Google Scholar] [CrossRef]
- Lagg, A.; Taucher, J.; Lindinger, W. Application of proton transfer reactions to gas analysis. Int. J. Mass Spectrom. Ion Process. 1994, 1, 55–66. [Google Scholar] [CrossRef]
- Xu, Z.Q.; Broza, Y.Y.; Ionsecu, R.; Tisch, U.; Ding, L.; Liu, H.; Song, Q.; Pan, Y.Y.; Xiong, F.X.; Gu, K.S.; et al. A nanomaterial-based breath test for distinguishing gastric cancer from benign gastric conditions. Br. J. Cancer 2013, 108, 941–950. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Mohan, A.; Guleria, R. Biomarkers in cancer screening, research and detection: Present and future: A review. Biomarkers 2006, 11, 385–405. [Google Scholar] [CrossRef]
- Riemenschneider, W.; Bolt, H. Esters, Organic; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2005. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.; Cross, C.E. Free radicals, antioxidants, and human disease: Where are we now? J. Lab. Clin. Med. 1992, 119, 598–620. [Google Scholar]
- Guengerich, F.P.; Shimada, T. Oxidation of toxic and carcinogenic chemicals by human cytochrome P-450 enzymes. Chem. Res. Toxicol. 1991, 4, 391–407. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, M. Toxicological Review of Acetonitrile; U.S. Environmental Protection Agency: Washington, DC, USA, 1999. [Google Scholar]
- Shibamoto, T. Analytical methods for trace levels of reactive carbonyl compounds formed in lipid peroxidation systems. J. Pharm. Biomed. Anal. 2006, 41, 12–25. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997, 82, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Peng, G.; Hakim, M.; Broza, Y.Y.; Billan, S.; Abdah-Bortnyak, R.; Kuten, A.; Tisch, U.; Haick, H. Detection of lung, breast, colorectal, and prostate cancers from exhaled breath using a single array of nanosensors. Br. J. Cancer 2010, 103, 542–551. [Google Scholar] [CrossRef]
- Kasapovic, J.; Pejic, S.; Todorovic, A.; Stojiljkovic, V.; Pajovic, S.B. Antioxidant status and lipid peroxidation in the blood of breast cancer patients of different ages. Cell Biochem. Funct. 2008, 26, 723–730. [Google Scholar] [CrossRef]
- Mense, S.M.; Remotti, F.; Bhan, A.; Singh, B.; El-Tamer, M.; Hei, T.K.; Bhat, H.K. Estrogen-induced breast cancer: Alterations in breast morphology and oxidative stress as a function of estrogen exposure. Toxicol. Appl. Pharmacol. 2008, 232, 78–85. [Google Scholar] [CrossRef] [Green Version]
- Phillips, M.; Cataneo, R.N.; Ditkoff, B.A.; Fisher, P.; Greenberg, J.; Gunawardena, R.; Kwon, C.S.; Tietje, O.; Wong, C. Prediction of breast cancer using volatile biomarkers in the breath. Breast Cancer Res. Treat. 2006, 99, 19–21. [Google Scholar] [CrossRef]
- Ebeler, S.E.; Clifford, A.J.; Shibamoto, T. Quantitative analysis by gas chromatography of volatile carbonyl compounds in expired air from mice and human. J. Chromatogr. B Biomed. Sci. Appl. 1997, 702, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Peng, Y.; Liu, Y.; Li, W.; Jin, Y.; Tang, Z.; Duan, Y. Investigation of potential breath biomarkers for the early diagnosis of breast cancer using gas chromatography-mass spectrometry. Clin. Chim. Acta 2014, 436, 59–67. [Google Scholar] [CrossRef]
- Haick, H.; Broza, Y.Y.; Mochalski, P.; Ruzsanyi, V.; Amann, A. Assessment, origin, and implementation of breath volatile cancer markers. Chem. Soc. Rev. 2014, 43, 1423–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kneepkens, C.M.; Ferreira, C.; Lepage, G.; Roy, C.C. The hydrocarbon breath test in the study of lipid peroxidation: Principles and practice. Clin. Investig. Med. 1992, 15, 163–186. [Google Scholar]
- Liu, Y.; Li, W.; Duan, Y. Effect of H2O2 induced oxidative stress (OS) on volatile organic compounds (VOCs) and intracellular metabolism in MCF-7 breast cancer cells. J. Breath Res. 2019, 13, 036005. [Google Scholar] [CrossRef]
- Cikach, F.S., Jr.; Dweik, R.A. Cardiovascular biomarkers in exhaled breath. Prog. Cardiovasc. Dis. 2012, 55, 34–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, J.F.; Maier, C.S. Acrolein: Sources, metabolism, and biomolecular interactions relevant to human health and disease. Mol. Nutr. Food Res. 2008, 52, 7–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghe, A.; Ghare, S.; Lamoreau, B.; Mohammad, M.; Barve, S.; McClain, C.; Joshi-Barve, S. Molecular mechanisms of acrolein toxicity: Relevance to human disease. Toxicol. Sci. 2015, 143, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Clemens, M.R.; Remmer, H.; Waller, H.D. Phenylhydrazine-induced lipid peroxidation of red blood cells in vitro and in vivo: Monitoring by the production of volatile hydrocarbons. Biochem. Pharmacol. 1984, 33, 1715–1718. [Google Scholar] [CrossRef] [PubMed]
- Kessler, W.; Remmer, H. Generation of volatile hydrocarbons from amino acids and proteins by an iron/ascorbate/GSH system. Biochem. Pharmacol. 1990, 39, 1347–1351. [Google Scholar] [CrossRef]
- Edson, K.Z.; Rettie, A.E. CYP4 enzymes as potential drug targets: Focus on enzyme multiplicity, inducers and inhibitors, and therapeutic modulation of 20-hydroxyeicosatetraenoic acid (20-HETE) synthase and fatty acid omega-hydroxylase activities. Curr. Top. Med. Chem. 2013, 13, 1429–1440. [Google Scholar] [CrossRef]
- Lieber, C.S. Cytochrome P-4502E1: Its physiological and pathological role. Physiol. Rev. 1997, 77, 517–544. [Google Scholar] [CrossRef] [PubMed]
- Raucy, J.L.; Lasker, J.M.; Kraner, J.C.; Salazar, D.E.; Lieber, C.S.; Corcoran, G.B. Induction of cytochrome P450IIE1 in the obese overfed rat. Mol. Pharmacol. 1991, 39, 275–280. [Google Scholar]
- Weltman, M.D.; Farrell, G.C.; Hall, P.; Ingelman-Sundberg, M.; Liddle, C. Hepatic cytochrome P450 2E1 is increased in patients with nonalcoholic steatohepatitis. Hepatology 1998, 27, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.; Leclercq, I.; Farrell, G.C. Nonalcoholic steatosis and steatohepatitis. II. Cytochrome P-450 enzymes and oxidative stress. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G1135–G1139. [Google Scholar] [CrossRef] [PubMed]
- Aghdassi, E.; Allard, J.P. Breath alkanes as a marker of oxidative stress in different clinical conditions. Free Radic. Biol. Med. 2000, 28, 880–886. [Google Scholar] [CrossRef]
- Bernhard, D.; Wang, X.L. Smoking, oxidative stress and cardiovascular diseases-do anti-oxidative therapies fail? Curr. Med. Chem. 2007, 14, 1703–1712. [Google Scholar] [CrossRef] [PubMed]
- Allard, J.P.; Royall, D.; Kurian, R.; Muggli, R.; Jeejeebhoy, K.N. Effects of beta-carotene supplementation on lipid peroxidation in humans. Am. J. Clin. Nutr. 1994, 59, 884–890. [Google Scholar] [CrossRef]
- Gelmont, D.; Stein, R.A.; Mead, J.F. The bacterial origin of rat breath pentane. Biochem. Biophys. Res. Commun. 1981, 102, 932–936. [Google Scholar] [CrossRef]
- Smith, U.; Hammersten, J.; Bjorntorp, P.; Kral, J.G. Regional differences and effect of weight reduction on human fat cell metabolism. Eur. J. Clin. Investig. 1979, 9, 327–332. [Google Scholar] [CrossRef]
- Mittelman, S.D.; Van Citters, G.W.; Kirkman, E.L.; Bergman, R.N. Extreme insulin resistance of the central adipose depot in vivo. Diabetes 2002, 51, 755–761. [Google Scholar] [CrossRef] [Green Version]
- Meyer, J.; Salamon, A.; Mispagel, S.; Kamp, G.; Peters, K. Energy metabolic capacities of human adipose-derived mesenchymal stromal cells in vitro and their adaptations in osteogenic and adipogenic differentiation. Exp. Cell Res. 2018, 370, 632–642. [Google Scholar] [CrossRef]
- Drolet, R.; Richard, C.; Sniderman, A.D.; Mailloux, J.; Fortier, M.; Huot, C.; Rheaume, C.; Tchernof, A. Hypertrophy and hyperplasia of abdominal adipose tissues in women. Int. J. Obes. 2008, 32, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Jo, J.; Gavrilova, O.; Pack, S.; Jou, W.; Mullen, S.; Sumner, A.E.; Cushman, S.W.; Periwal, V. Hypertrophy and/or Hyperplasia: Dynamics of Adipose Tissue Growth. PLoS Comput. Biol. 2009, 5, e1000324. [Google Scholar] [CrossRef]
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Naslund, E.; Britton, T.; et al. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Zeve, D.; Tang, W.; Graff, J. Fighting fat with fat: The expanding field of adipose stem cells. Cell Stem Cell 2009, 5, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Tsushima, Y.; Nishizawa, H.; Tochino, Y.; Nakatsuji, H.; Sekimoto, R.; Nagao, H.; Shirakura, T.; Kato, K.; Imaizumi, K.; Takahashi, H.; et al. Uric acid secretion from adipose tissue and its increase in obesity. J. Biol. Chem. 2013, 288, 27138–27149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kushch, I.; Arendacka, B.; Stolc, S.; Mochalski, P.; Filipiak, W.; Schwarz, K.; Schwentner, L.; Schmid, A.; Dzien, A.; Lechleitner, M.; et al. Breath isoprene--aspects of normal physiology related to age, gender and cholesterol profile as determined in a proton transfer reaction mass spectrometry study. Clin. Chem. Lab. Med. 2008, 46, 1011–1018. [Google Scholar] [CrossRef]
- Simonen, P.; Kotronen, A.; Hallikainen, M.; Sevastianova, K.; Makkonen, J.; Hakkarainen, A.; Lundbom, N.; Miettinen, T.A.; Gylling, H.; Yki-Jarvinen, H. Cholesterol synthesis is increased and absorption decreased in non-alcoholic fatty liver disease independent of obesity. J. Hepatol. 2011, 54, 153–159. [Google Scholar] [CrossRef]
- Salerno-Kennedy, R.; Cashman, K.D. Potential applications of breath isoprene as a biomarker in modern medicine: A concise overview. Wien. Klin. Wochenschr. 2005, 117, 180–186. [Google Scholar] [CrossRef]
- Miekisch, W.; Schubert, J.K.; Noeldge-Schomburg, G.F. Diagnostic potential of breath analysis--focus on volatile organic compounds. Clin. Chim. Acta 2004, 347, 25–39. [Google Scholar] [CrossRef]
- Franzese, A.; Vajro, P.; Argenziano, A.; Puzziello, A.; Iannucci, M.P.; Saviano, M.C.; Brunetti, F.; Rubino, A. Liver involvement in obese children. Ultrasonography and liver enzyme levels at diagnosis and during follow-up in an Italian population. Dig. Dis. Sci. 1997, 42, 1428–1432. [Google Scholar] [CrossRef]
- Schwimmer, J.B.; Deutsch, R.; Kahen, T.; Lavine, J.E.; Stanley, C.; Behling, C. Prevalence of fatty liver in children and adolescents. Pediatrics 2006, 118, 1388–1393. [Google Scholar] [CrossRef]
- Hecht, S.S. Tobacco carcinogens, their biomarkers and tobacco-induced cancer. Nat. Rev. Cancer 2003, 3, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [Green Version]
- Chien, P.J.; Suzuki, T.; Tsujii, M.; Ye, M.; Minami, I.; Toda, K.; Otsuka, H.; Toma, K.; Arakawa, T.; Araki, K.; et al. Biochemical Gas Sensors (Biosniffers) Using Forward and Reverse Reactions of Secondary Alcohol Dehydrogenase for Breath Isopropanol and Acetone as Potential Volatile Biomarkers of Diabetes Mellitus. Anal. Chem. 2017, 89, 12261–12268. [Google Scholar] [CrossRef]
- Capurso, C.; Capurso, A. From excess adiposity to insulin resistance: The role of free fatty acids. Vascul. Pharmacol. 2012, 57, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Grabowska-Polanowska, B.; Skowron, M.; Miarka, P.; Pietrzycka, A.; Sliwka, I. The application of chromatographic breath analysis in the search of volatile biomarkers of chronic kidney disease and coexisting type 2 diabetes mellitus. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017, 1060, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Leiherer, A.; Geiger, K.; Muendlein, A.; Drexel, H. Hypoxia induces a HIF-1alpha dependent signaling cascade to make a complex metabolic switch in SGBS-adipocytes. Mol. Cell. Endocrinol. 2014, 383, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Solga, S.F.; Alkhuraishe, A.; Cope, K.; Tabesh, A.; Clark, J.M.; Torbenson, M.; Schwartz, P.; Magnuson, T.; Diehl, A.M.; Risby, T.H. Breath biomarkers and non-alcoholic fatty liver disease: Preliminary observations. Biomarkers 2006, 11, 174–183. [Google Scholar] [CrossRef]
- Raman, M.; Ahmed, I.; Gillevet, P.M.; Probert, C.S.; Ratcliffe, N.M.; Smith, S.; Greenwood, R.; Sikaroodi, M.; Lam, V.; Crotty, P.; et al. Fecal microbiome and volatile organic compound metabolome in obese humans with nonalcoholic fatty liver disease. Clin. Gastroenterol. Hepatol. 2013, 11, 868–875.e3. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| No. | Volatile Metabolite | Study | Method | Fluid |
|---|---|---|---|---|
| 1 | 5-methyl-3-hexanone, 1-heptanol, 4-methyl-2-heptanone, 2-hexanol, dimethyl sulfone, formamide N, N-dibutyl, 1-hexanol, 2-pentanone, 2,4,6-trimethyl-pyridine, 3-hexanone, 3-octanone, 2,4,4-trimethyl-1-pentanol | Cozzolino et al. [54] | SPME and GC-MS | Human urine |
| 2 | Acetic acid, methanol, carbon dioxide, (methylthio)methanethiol, acrolein, methylacetate, ammonia, fragments of aldehyde (butanal, hexanal, octanal or nonanal), acetone, propanol | Kistler et al. [55] | Proton-transfer reaction time-of-flight mass spectrometry (PTF-MS) | Mouse breath |
| 3 | Methanol, methylacetate, propionate, dimethyl disulfone | Kistler et al. [56] | PTF-MS | Mouse breath |
| 4 | Acetaldehyde, acetone, isoprene, 1-decene, 1-octene, ammonia, hydrogen sulfide | Alkhouri et al. [57] | SIFT-MS | Human breath |
| 5 | 2-propanol, acetaldehyde, acetone, acrylonitrite, benzene, carbon disulfide, dimethylsulfide, ethanol, isoprene, pentane, 1-decene, 1-heptane, 1-nonene,1-octene, 3-methylhexane, 2-nonene, ammonia, ethane, hydrogen sulfide, triethylamine, trimethylamine | Alkhouri et al. [58] | SIFT-MS | Human breath |
| 6 | Acetaldehyde, acetone, 2-methyl-butanal, 3-methyl butanal, 5-octadecene, 3-methyl butanol, 1-pentanol, methylpyrazine, 2,6 dimethyl pyrazine, dimethylsulfide, nonanal, methional, 3-octadecene, phenol | Uchikawa et al. [59] | Headspace sampler GC-MS | Mouse feces |
| 7 | Acetaldehyde, pentane, 1,3-bis-(1,1- dimethylethyl) benzene, ethylbenzene, benzaldehyde, heptanal and octanal | Klemenz et al. [60] | Needle trap micro extraction and GC-MS | Adipogenically differentiated mesenchymal stromal/stem cells from human adipose tissue |
| 8 | Tetrachloroethane, 2,3,5 trimethyl-hexane, beta-pinene, 1,3,5 trimethyl benzene, 9-methyl acridine, tetradecane, 6,10 dimethyl-5,9 undecadien-2-one, beta-ionone | Dragonieri et al. [61] | Electronic nose TD-GC-MS | Human breath |
| No. | Class of VOC | Metabolic Pathway | Examples of the VOC |
|---|---|---|---|
| 1 | Hydrocarbon | Lipid peroxidation | Ethane, pentane, decane, hexane, dodecane, branched chain: 3-ethyl hexane, tetradecane, tridecane. 2,4 dimethyl eicosane |
| 2 | Alcohol | Production of hydrocarbon metabolism, alcohol metabolism by alcohol dehydrogenase and cytochrome p450 (CYP2E1) | Cyclohexanol, propanol, 1-decanol, 3-octanol |
| 3 | Aldehydes | Alcohol metabolism, secondary product of lipid peroxidation, detoxification process by cytochrome p450, smoking and diet | Nonanal, decanal, butanal, pentanal |
| 4 | Ketones | Fatty acid oxidation and protein metabolism | Heptanone, acetone, 2-pentanone, 4-octanone |
| 5 | Esters | Lipid hydrolysis | Isopropylmyristate |
| 6 | Nitriles and aromatic compounds | Exogenous origin that are highly reactive resulting in peroxidative damage to PUFA, DNA, and protein. | Acetonitrile |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oyerinde, A.S.; Selvaraju, V.; Babu, J.R.; Geetha, T. Potential Role of Oxidative Stress in the Production of Volatile Organic Compounds in Obesity. Antioxidants 2023, 12, 129. https://doi.org/10.3390/antiox12010129
Oyerinde AS, Selvaraju V, Babu JR, Geetha T. Potential Role of Oxidative Stress in the Production of Volatile Organic Compounds in Obesity. Antioxidants. 2023; 12(1):129. https://doi.org/10.3390/antiox12010129
Chicago/Turabian StyleOyerinde, Adebowale Samuel, Vaithinathan Selvaraju, Jeganathan Ramesh Babu, and Thangiah Geetha. 2023. "Potential Role of Oxidative Stress in the Production of Volatile Organic Compounds in Obesity" Antioxidants 12, no. 1: 129. https://doi.org/10.3390/antiox12010129