
Intermediate-Term Storage of Spotted Halibut (Verasper variegatus) Sperm: Effects of Storage Methods, Extenders Supplemented with Antibiotics and Antioxidants on Sperm Quality

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Broodstock and Collection of Gamates
2.2. Sperm Motility, Duration Motility, and Cell Viability
2.3. Experimental Design
2.3.1. Effect of Diluent
2.3.2. Effect of Temperature
2.3.3. Effect of Dilution Ratio
2.3.4. Effect of Antibiotics
2.3.5. Effect of Antioxidants
2.3.6. Statistical Analysis
3. Results
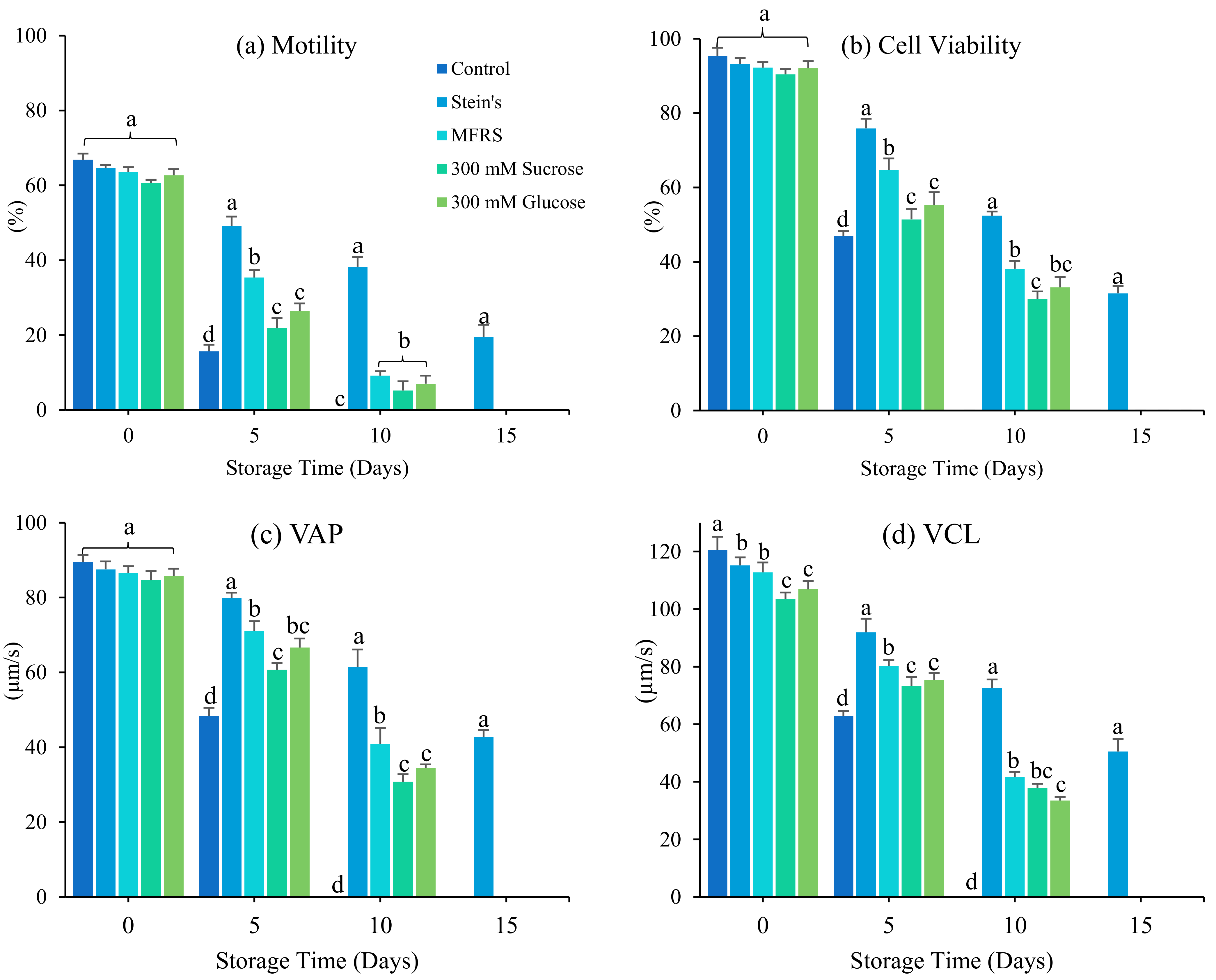
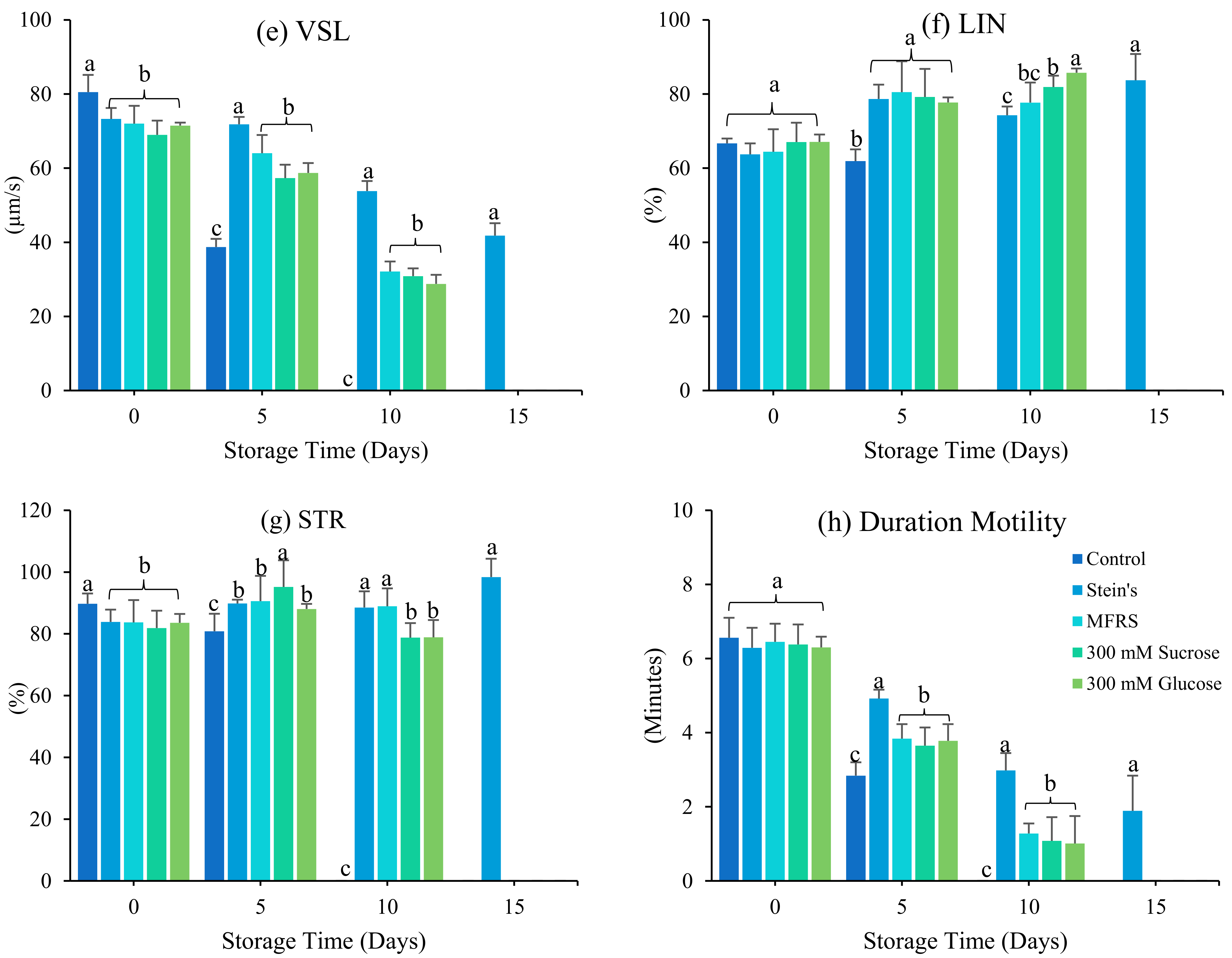
3.1. Effect of Diluents
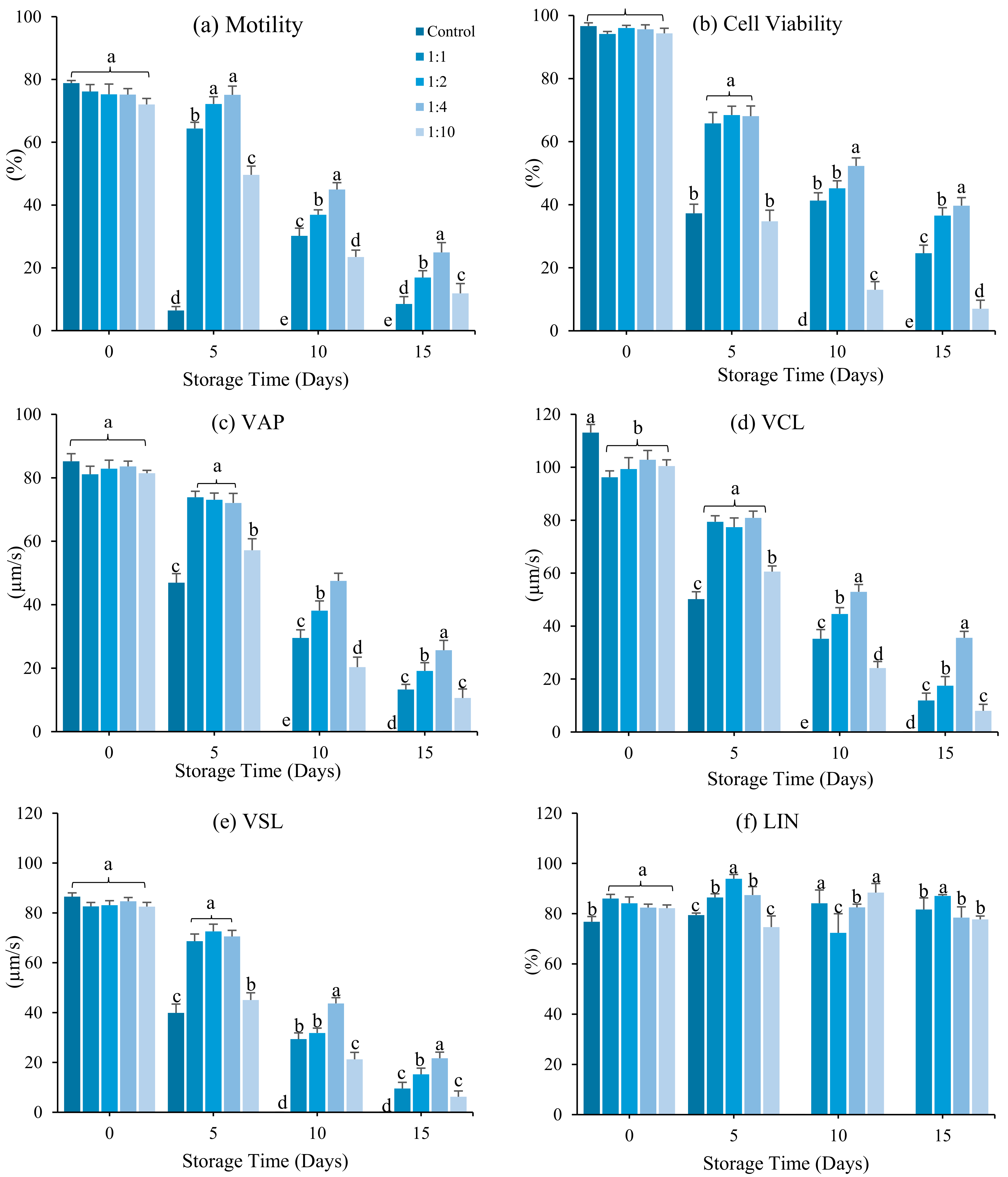
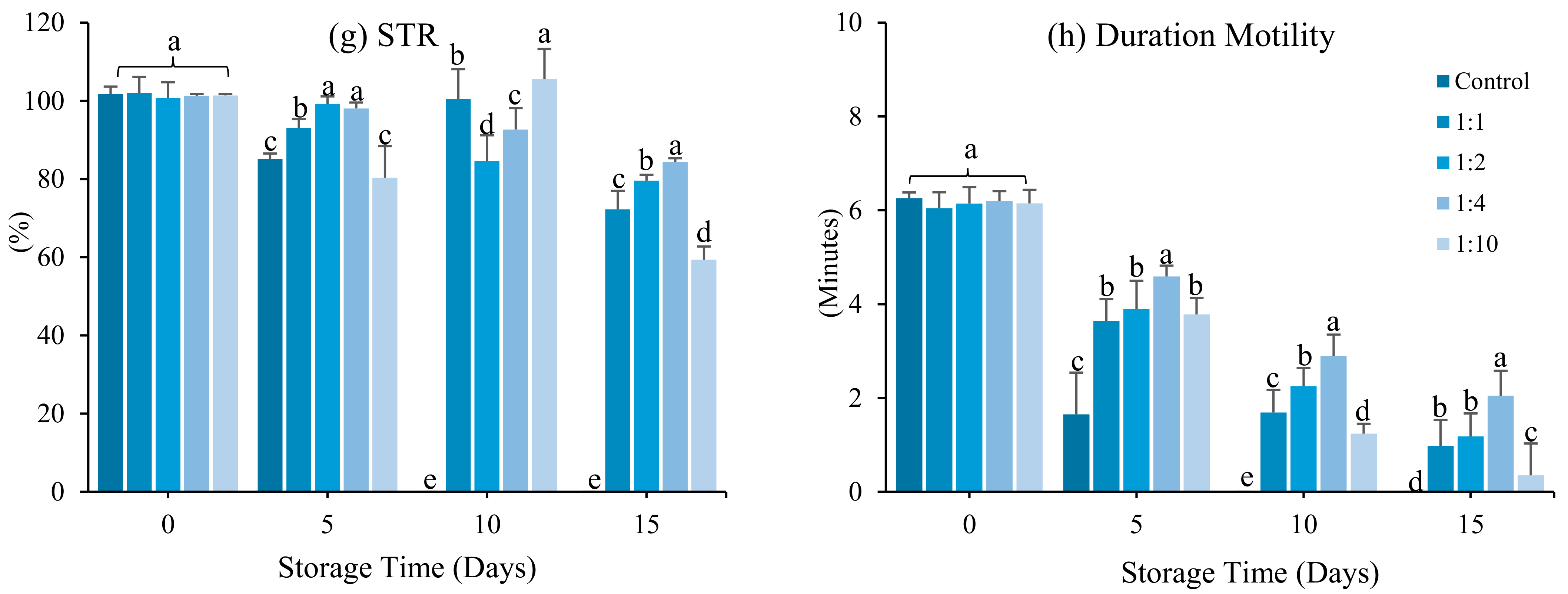
3.2. Effect of Dilution Ratio
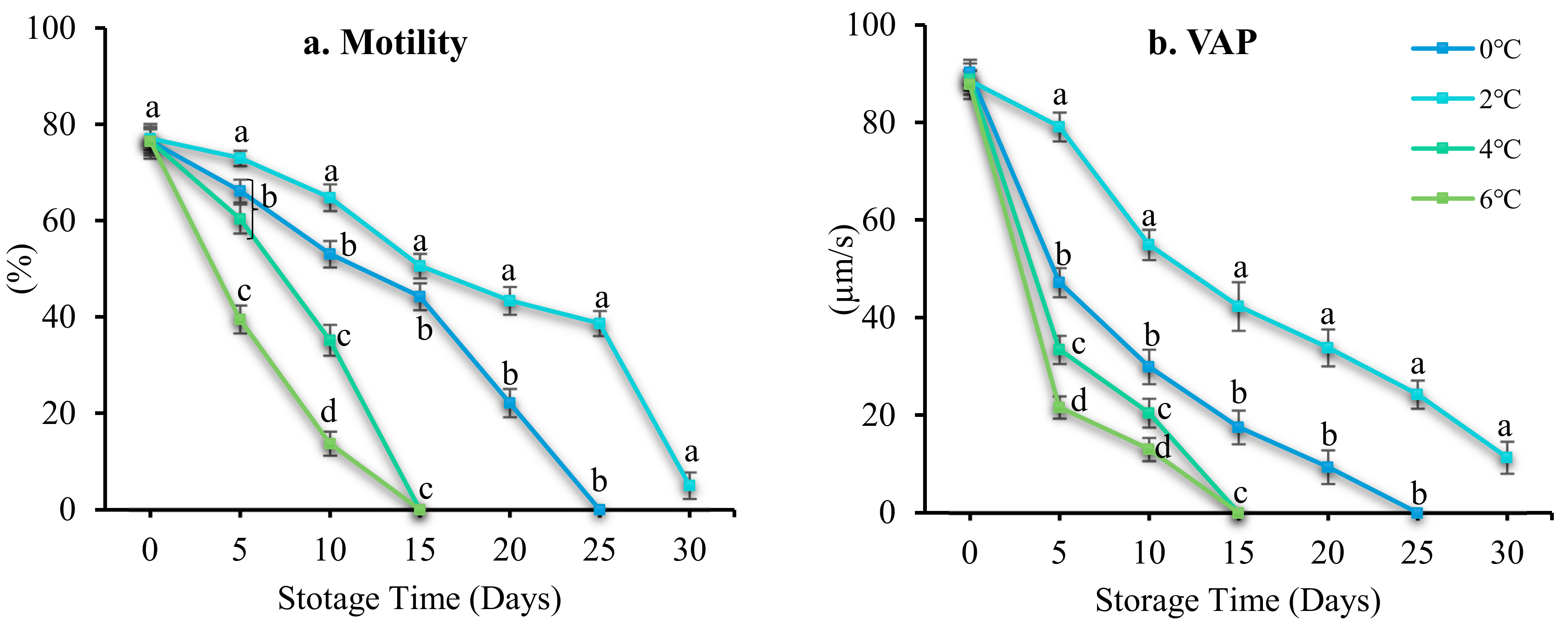
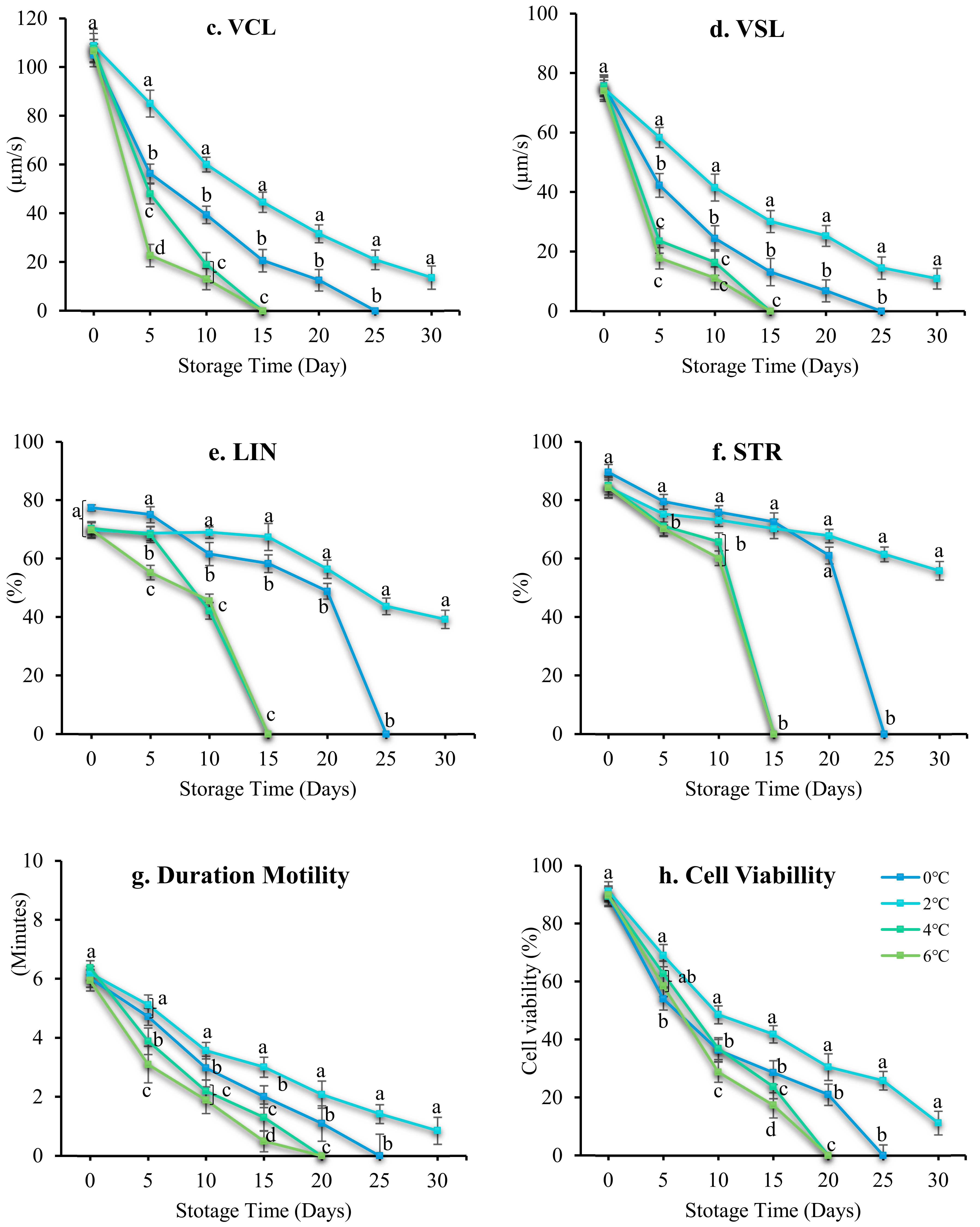
3.3. Effect of Storage Temperature
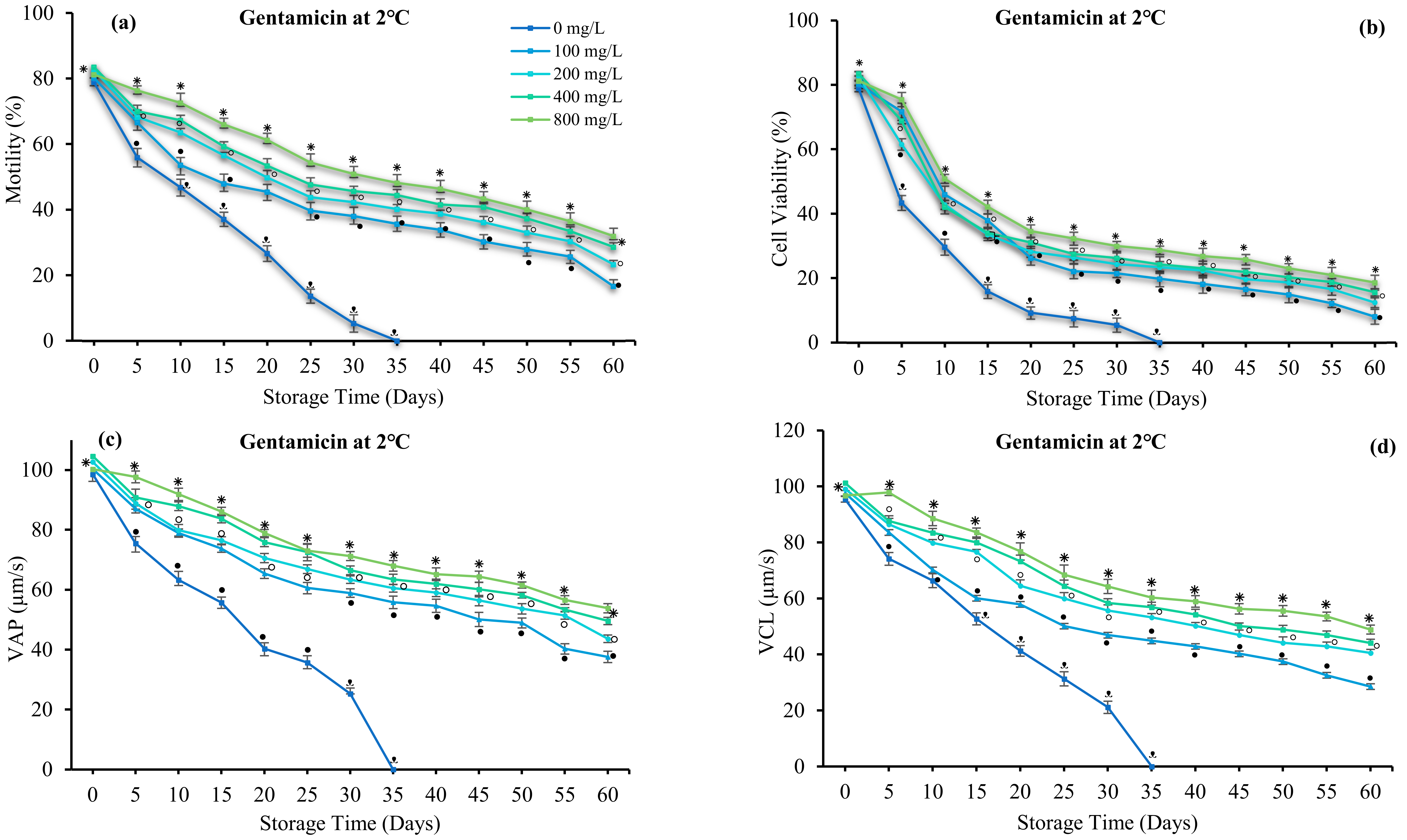
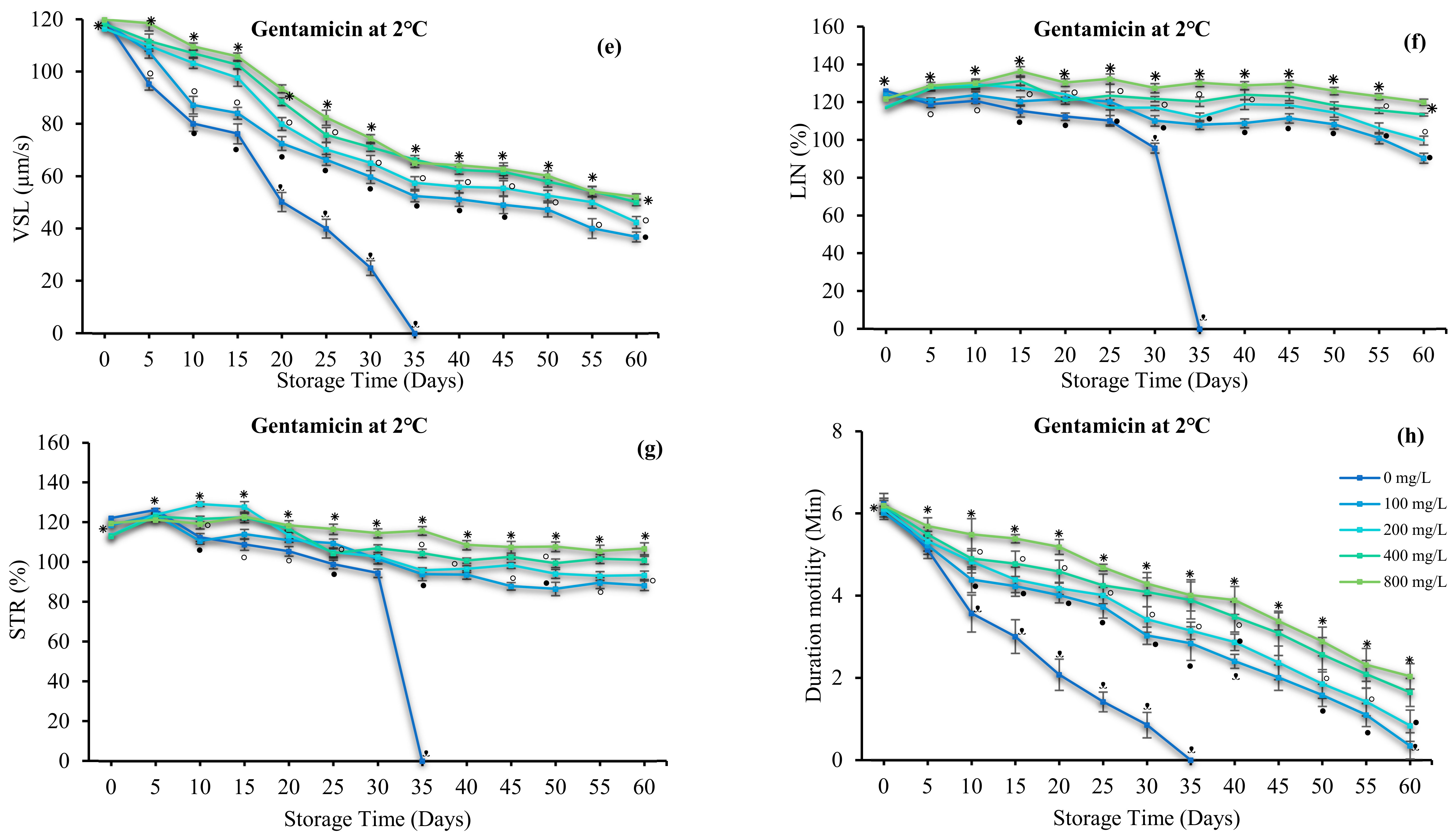
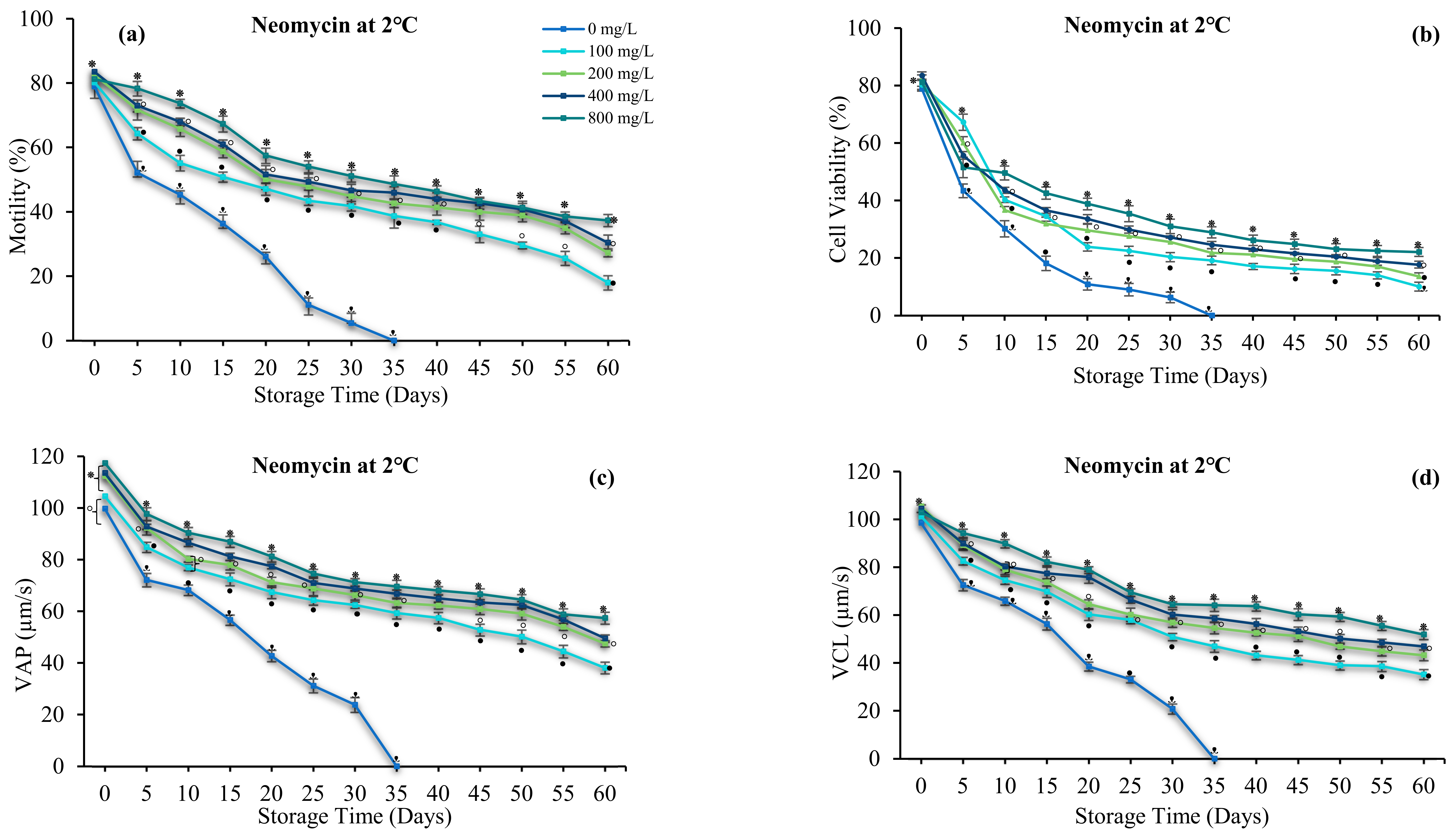
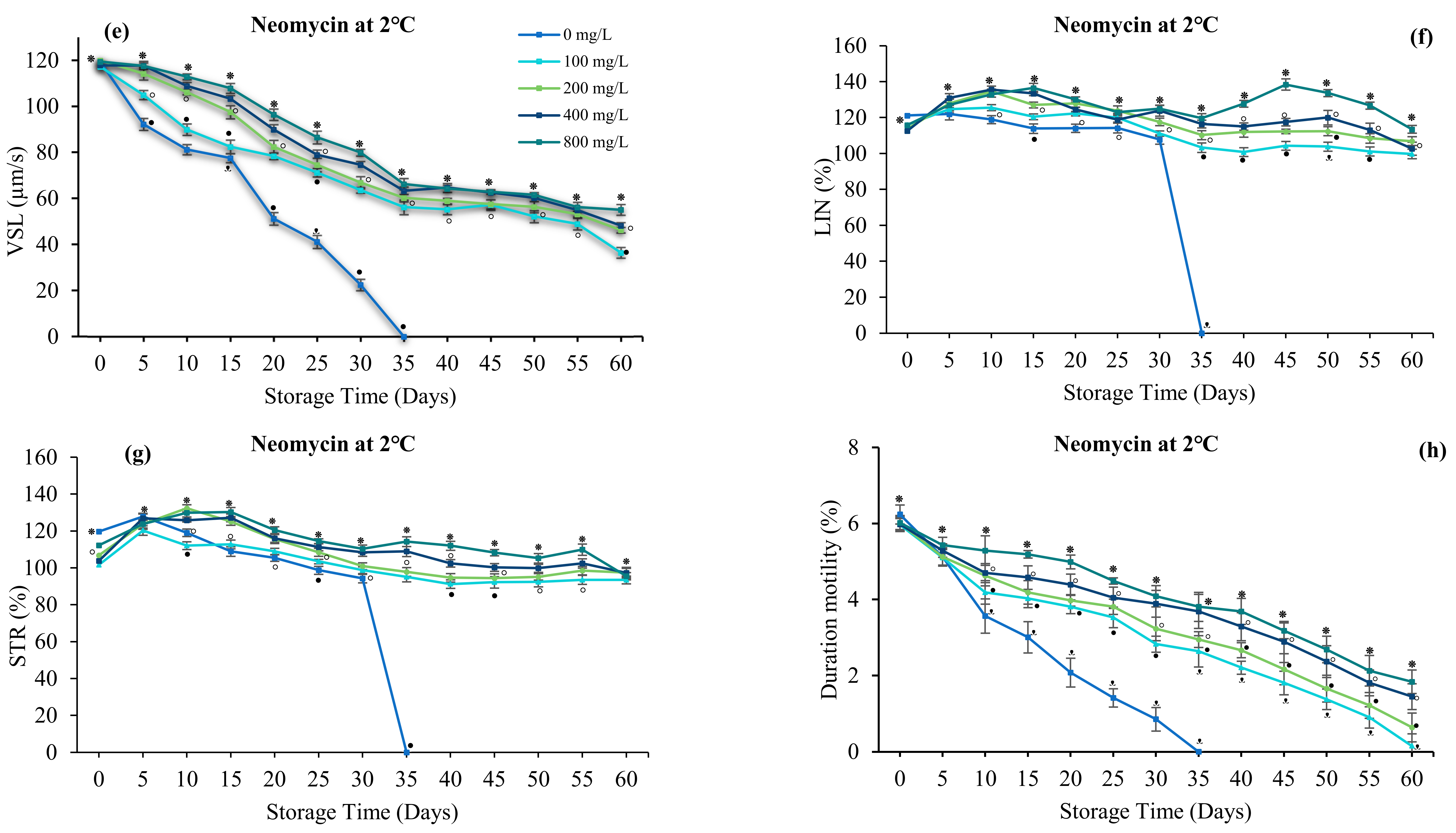
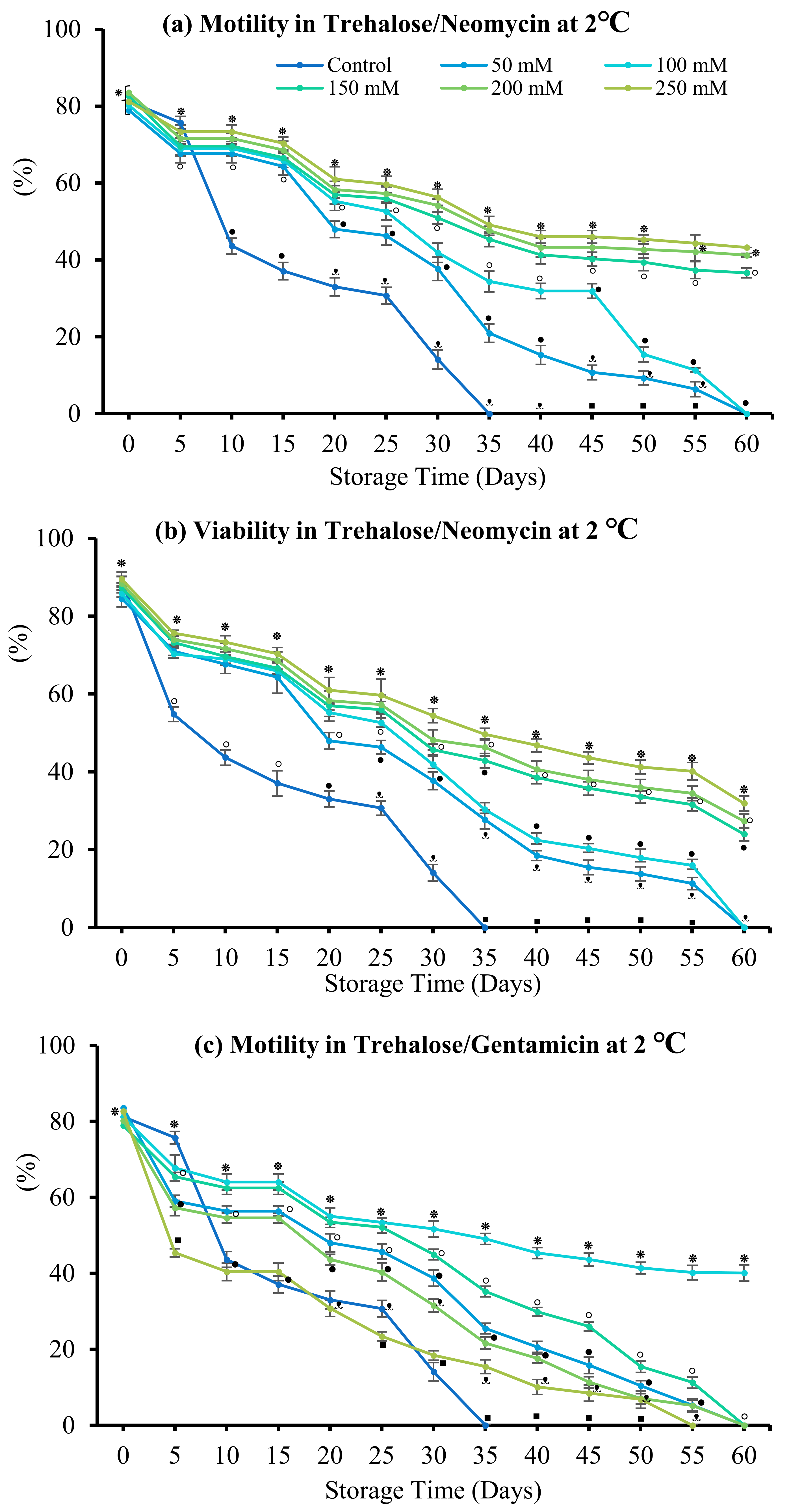
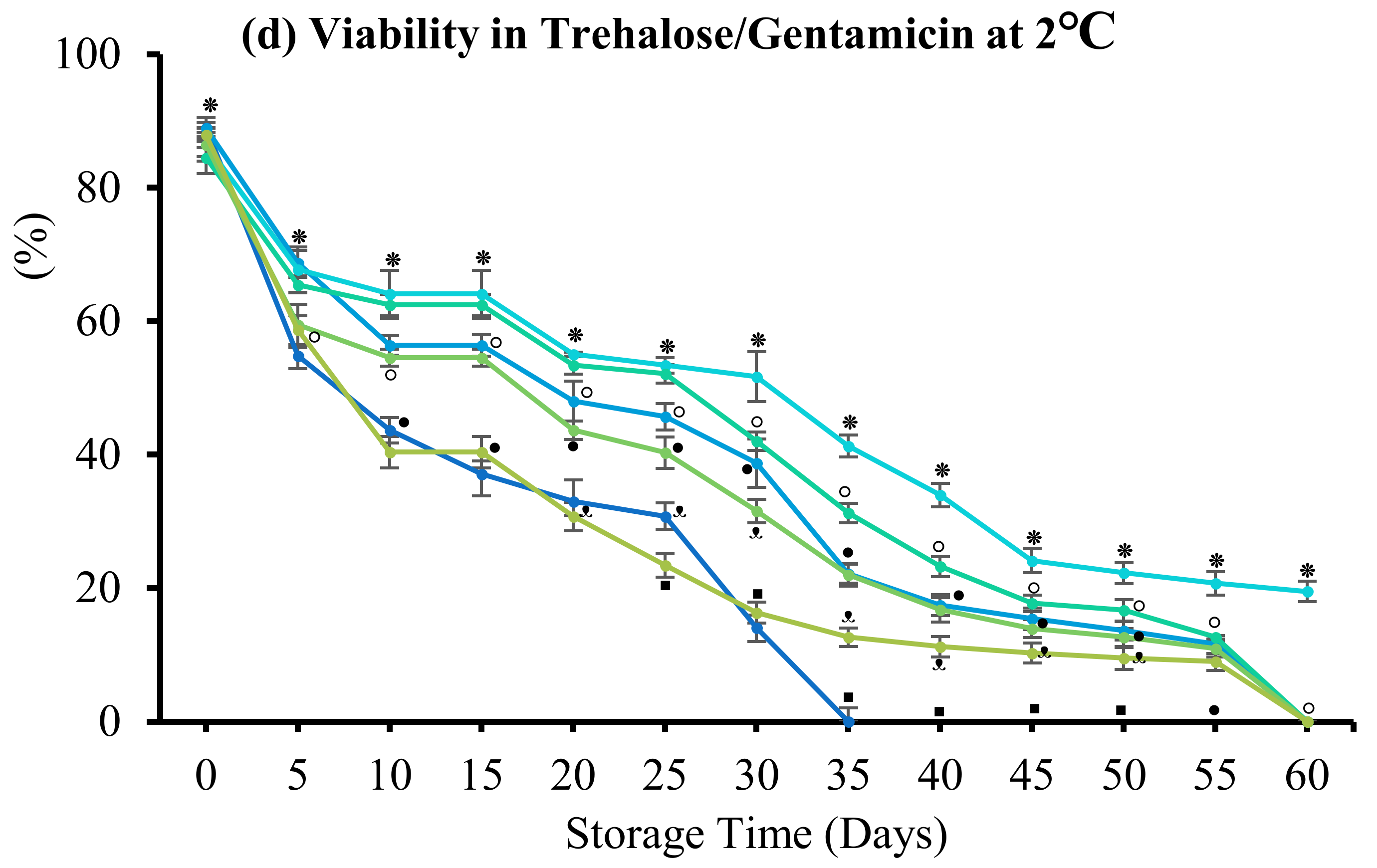
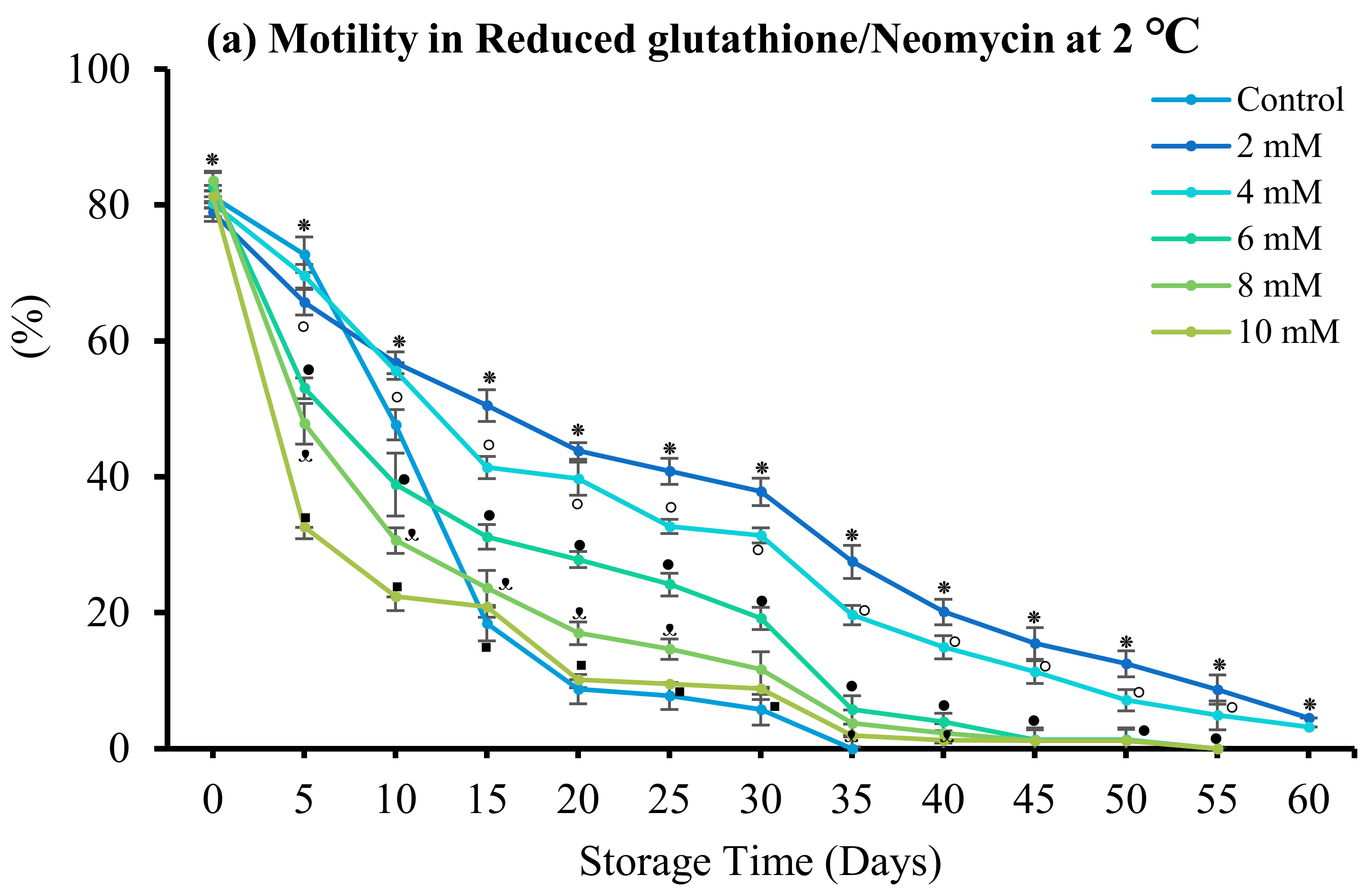
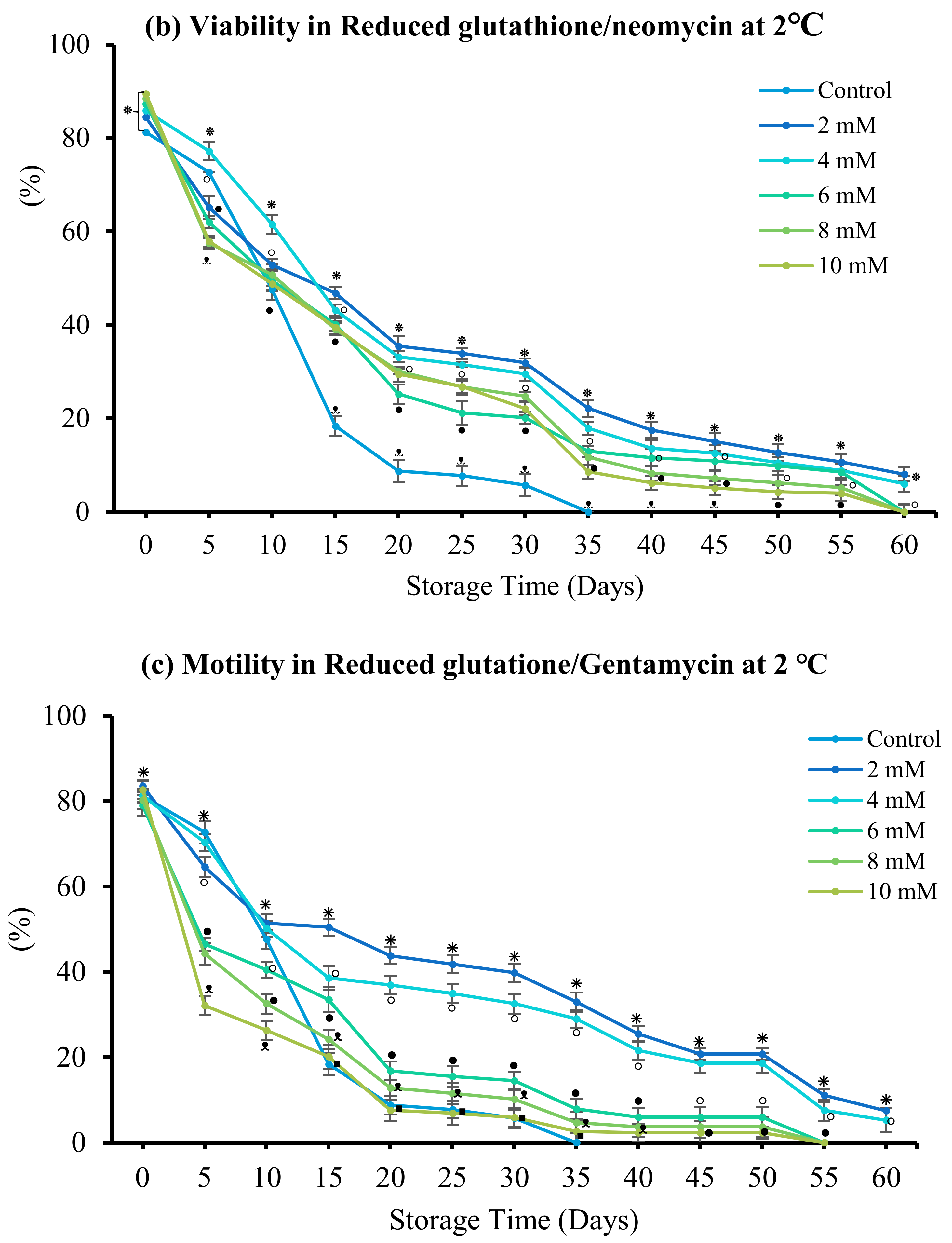
3.4. Effect of Antibiotics
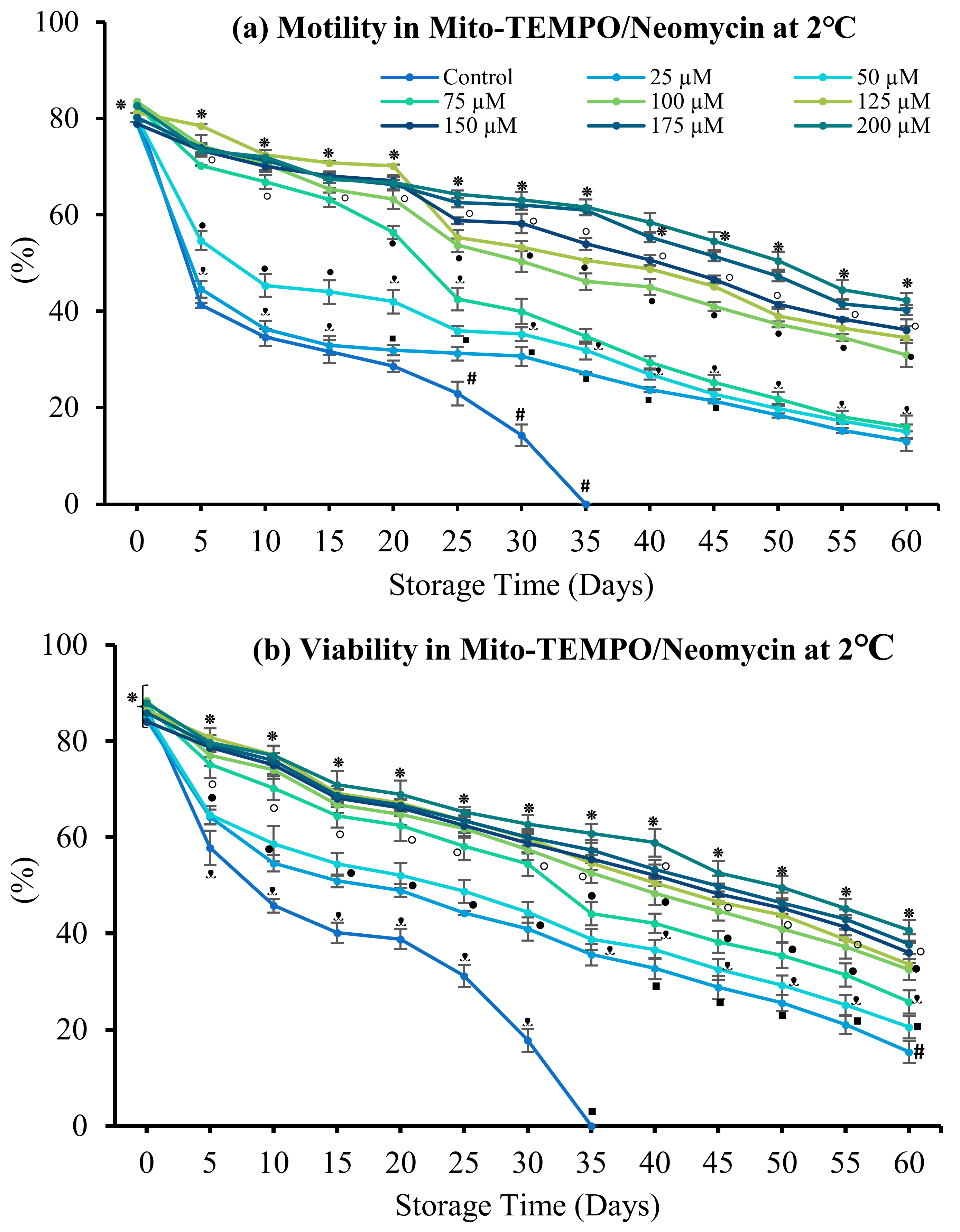
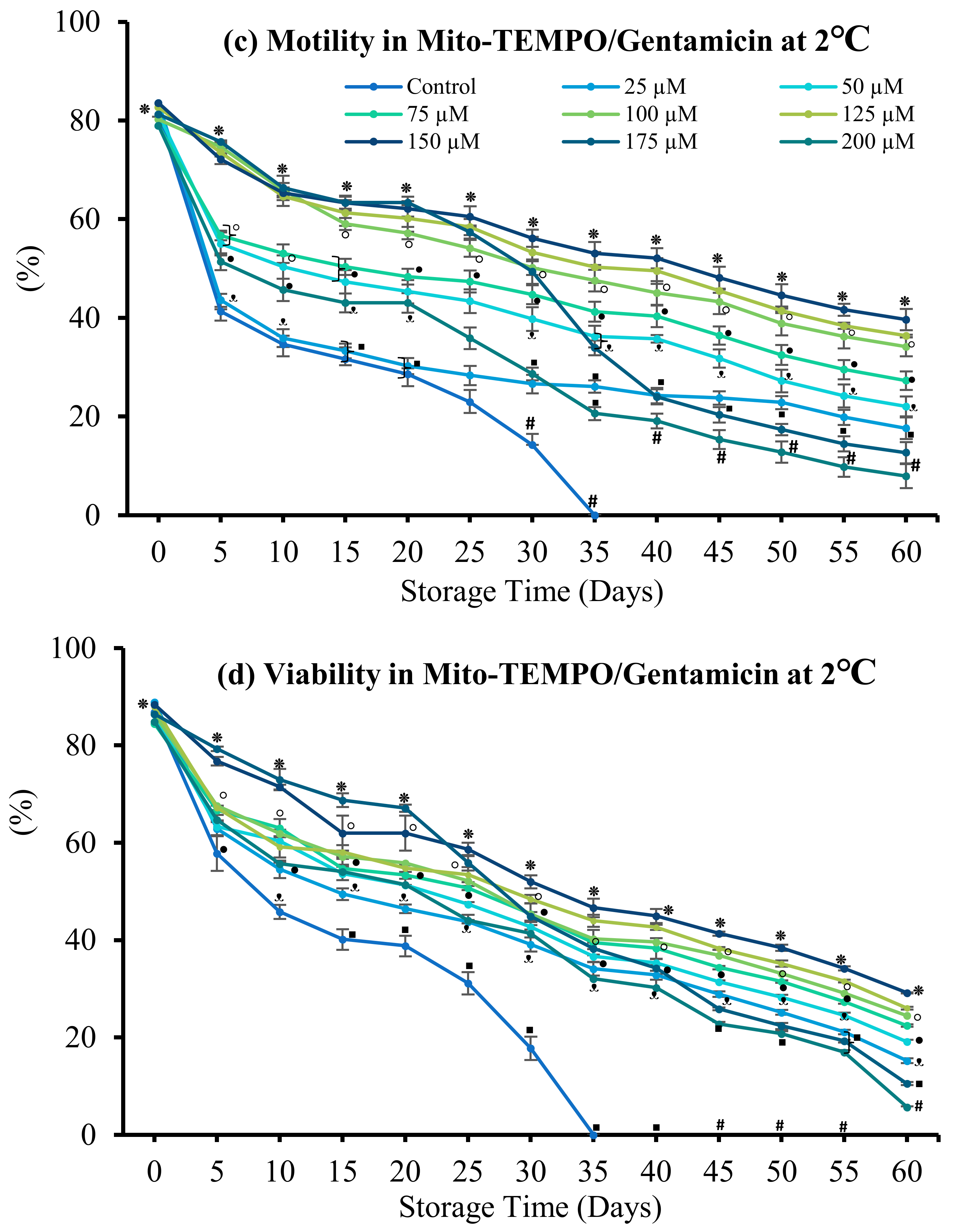
3.4.1. Mito-TEMPO
3.4.2. Trehalose
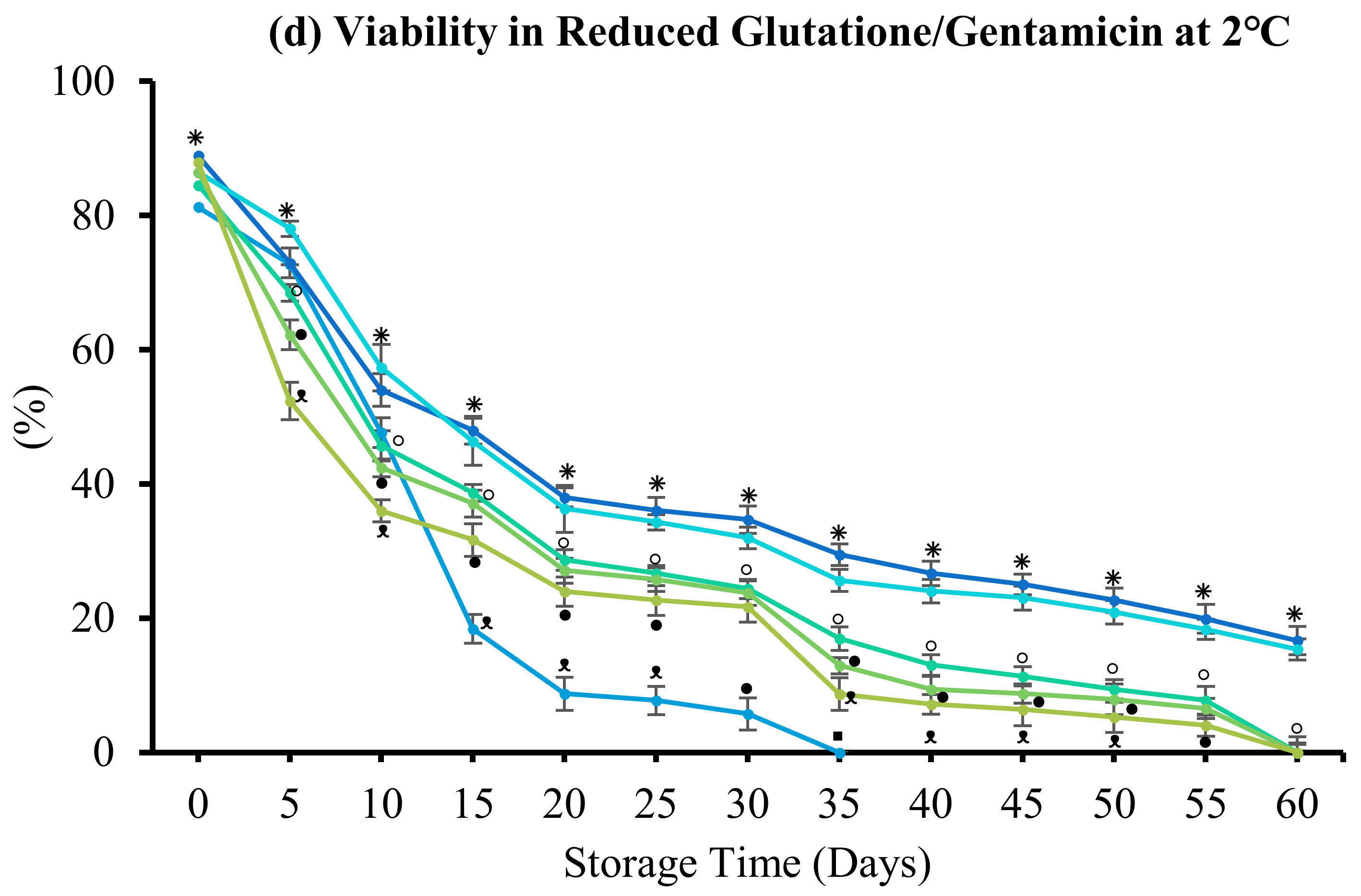
3.4.3. Reduced Glutathione
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zidni, I.; Lee, Y.H.; Park, J.Y.; Lee, H.B.; Hur, J.W.; Lim, H.K. Effects of Cryoprotective Medium Composition, Dilution Ratio, and Freezing Rates on Spotted Halibut (Verasper Variegatus) Sperm Cryopreservation. Animals 2020, 10, 2153. [Google Scholar] [CrossRef] [PubMed]
- Magnotti, C.; Cerqueira, V.; Lee-Estevez, M.; Farias, J.G.; Valdebenito, I.; Figueroa, E. Cryopreservation and Vitrification of Fish Semen: A Review with Special Emphasis on Marine Species. Rev. Aquac. 2018, 10, 15–25. [Google Scholar] [CrossRef]
- Shaliutina, A.; Hulak, M.; Gazo, I.; Linhartova, P.; Linhart, O. Effect of Short-Term Storage on Quality Parameters, DNA Integrity, and Oxidative Stress in Russian (Acipenser gueldenstaedtii) and Siberian (Acipenser baerii) Sturgeon Sperm. Anim. Reprod. Sci. 2013, 139, 127–135. [Google Scholar] [CrossRef]
- Riley, P. Refrigerated Storage and Cryopreservation of Sperm for the Production of Red Snapper and Snapper Hybrids. Master’s Thesis, Louisiana State University, Baton Rouge, LA, USA, 2002. [Google Scholar]
- Lim, H.K.; An, C.M.; Son, M.H.; Park, M.-W.; Park, Y.J. Effect of Diluents for Cold Storage of Olive Flounder (Paralichthys Olivaceus) Sperm. Korean J. Fish. Aquat. Sci. 2005, 38, 239–244. [Google Scholar]
- Chang, Y.J.; Chang, Y.J.; Lim, H.K.; Lee, J.K.; Park, Y.J. Cold Storage of Milt from Four Species of Flatfish. Fish. Aquat. Sci. 2002, 5, 64–74. [Google Scholar] [CrossRef]
- Lim, H.-K.; An, C.M.; Son, M.H.; Park, M.-W.; Kim, E.O.; Byun, S.-G. Effect of Diluents and Temperature on Sperm Storage in Starry Flounder (Platichthys stellatus). Korean Aquaculture Society. J. Aquac. 2006, 19, 47–51. [Google Scholar]
- Le, M.H.; Lim, H.K.; Min, B.H.; Park, M.S. Storage of Yellow Croaker Larimichthys Polyactis Semen. Busan 608-. Isr. J. Aquac. Bamidgeh 2011, 63, 7. [Google Scholar]
- Yang, Y.; Wang, T.; Yang, S.; Wu, X.; Huang, W.; Fan, B.; Xia, J.; Meng, Z.; Liu, X. The Effects of Storage in Vitro on Functions, Transcriptome, Proteome, and Oxidation Resistance of Giant Grouper Sperm. Front. Mar. Sci. 2021, 8, 716047. [Google Scholar] [CrossRef]
- Merino, O.; Figueroa, E.; Cheuquemán, C.; Valdebenito, I.; Isachenko, V.; Isachenko, E.; Sánchez, R.; Farías, J.; Risopatrón, J. Short-Term Storage of Salmonids Semen in a Sodium Alginate-Based Extender. Andrologia 2017, 49, e12661. [Google Scholar] [CrossRef]
- Boonthai, T.; Khaopong, W.; Sangsong, J.; Vuthiphandchai, V.; Nimrat, S. In Vitro Inoculation of Aeromonas hydrophila and Pseudomonas fluorescens in Cryopreserved Silver Barb (Barbodes gonionotus) Milt: Effect on Fertilization Capacity and Transmission Potential to Embryos. Theriogenology 2018, 108, 1–6. [Google Scholar] [CrossRef]
- Segovia, M.; Jenkins, J.A.; Paniagua-Chavez, C.; Tiersch, T.R. Flow cytometric evaluation of antibiotic effects on viability and mitochondrial function of refrigerated spermatozoa of nile tilapia. Theriogenology 1999, 53, 1489–1499. [Google Scholar] [CrossRef]
- Figueroa, E.; Risopatrón, J.; Sánchez, R.; Isachenko, E.; Merino, O.; Isachenko, V.; Valdebenito, I. Spermatozoa Vitrification of Sex-Reversed Rainbow Trout (Oncorhynchus Mykiss): Effect of Seminal Plasma on Physiological Parameters. Aquaculture 2013, 372–375, 119–126. [Google Scholar] [CrossRef]
- Zidni, I.; Bin, H.; Hye, J.; Yeol, J.; Dae, Y.; Seok, H.; Su, Y.; Young, I.; Kyu, H. Effect of Antioxidants in Cryopreservation Media on Spotted Halibut (Verasfer variegatus) Sperm Quality during Cryopreservation. Aquaculture 2022, 557, 738351. [Google Scholar] [CrossRef]
- Félix, F.; Oliveira, C.C.V.; Cabrita, E. Antioxidants in Fish Sperm and the Potential Role of Melatonin. Antioxidants 2021, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Abualreesh, M.; Myers, J.N.; Gurbatow, J.; Johnson, A.; Xing, D.; Wang, J.; Li, S.; Coogan, M.; Vo, K.; El Husseini, N.; et al. Effects of Antioxidants and Antifreeze Proteins on Cryopreservation of Blue Catfish (Ictalurus furcatus) Spermatogonia. Aquaculture 2021, 531, 735966. [Google Scholar] [CrossRef]
- Herranz-Jusdado, J.G.; Gallego, V.; Rozenfeld, C.; Morini, M.; Pérez, L.; Asturiano, J.F. European Eel Sperm Storage: Optimization of Short-Term Protocols and Cryopreservation of Large Volumes. Aquaculture 2019, 506, 42–50. [Google Scholar] [CrossRef]
- Le, M.H.; Nguyen, T.T.; Pham, P.L. The Open Access Israeli Journal of Aquaculture—Bamidgeh Saprolegnia Pathogen from Pengze Crucian Carp. Isr. J. Aquac. Bamidgeh 2010, 57, 90–96. [Google Scholar]
- Contreras, P.; Dumorné, K.; Ulloa-Rodríguez, P.; Merino, O.; Figueroa, E.; Farías, J.G.; Valdebenito, I.; Risopatrón, J. Effects of Short-Term Storage on Sperm Function in Fish Semen: A Review. Rev. Aquac. 2020, 12, 1373–1389. [Google Scholar] [CrossRef]
- Chang, Y.-J.; Chang, Y.-J.; Lim, H.-K. Cryopreservation of Tiger Puffer (Takifugu Rubripes) Sperm. Dev. Reprod. 1997, 1, 29–36. [Google Scholar]
- Yasui, G.S.; Senhorini, J.A.; Shimoda, E.; Pereira-Santos, M.; Nakaghi, L.S.O.; Fujimoto, T.; Arias-Rodriguez, L.; Silva, L.A. Improvement of Gamete Quality and Its Short-Term Storage: An Approach for Biotechnology in Laboratory Fish. Animal 2015, 9, 464–470. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Zhao, E.; Lu, Y.; Yan, M.; Huang, C.; Dong, Q. Evaluation of Activation and Storage Conditions for Sperm of Yellow Drum Nibea Albiflora. Aquaculture 2012, 324–325, 319–322. [Google Scholar] [CrossRef]
- DeGraaf, J.D.; Berlinsky, D.L. Cryogenic and Refrigerated Storage of Rainbow Smelt Osmerus Mordax Spermatozoa. J. World Aquac. Soc. 2004, 35, 209–216. [Google Scholar] [CrossRef]
- Santos, M.; Soares, F.; Moreira, M.; Beirão, J. Evaluation of Different Extenders for the Cold Storage of Meagre (Argyrosomus regius) Semen. Aquac. Res. 2018, 49, 2723–2731. [Google Scholar] [CrossRef]
- Bobe, J.; Labbe, C. Chilled Storage of Sperm and Eggs. In Methods in Reproductive Aquaculture: Marine and Freshwater Species; CRC Press: Boca Raton, FL, USA, 2009; pp. 219–235. ISBN 9780849380549. [Google Scholar]
- Agarwal, A.; Gupta, S.; Sharma, R. Andrological Evaluation of Male Infertility; Springer: Cham, Switzerland, 2016; pp. 73–77. [Google Scholar] [CrossRef]
- Garcia, R.R.F.; Vasconcelos, A.C.N.; Povh, J.A.; Oberst, E.R.; Eloy, L.R.; Streit, D.P. Different Extenders Solutions for Tambaqui Semen Cooling. Pesqui. Agropecu. Bras. 2016, 51, 780–784. [Google Scholar] [CrossRef]
- Vuthiphandchai, V.; Thadsri, I.; Nimrat, S. Chilled Storage of Walking Catfish (Clarias macrocephalus) Semen. Aquaculture 2009, 296, 58–64. [Google Scholar] [CrossRef]
- Ulloa-Rodríguez, P.; Contreras, P.; Dumorné, K.; Lee-Estevez, M.; Díaz, R.; Figueroa, E.; Valdebenito, I.; Risopatrón, J.; Farías, J.G. Patagonian Blenny (Eleginops maclovinus) Spermatozoa Quality after Storage at 4 °C in Cortland Medium. Anim. Reprod. Sci. 2018, 197, 117–125. [Google Scholar] [CrossRef]
- Brown, G.G.; Mims, S.D. Storage, Transportation, and Fertility of Undiluted and Diluted Paddlefish Milt. Progress. Fish-Cult. 1995, 57, 64–69. [Google Scholar] [CrossRef]
- Harvey, B.; Kelley, R.N. Chilled Storage of Sarotherodon Mossambicus Milt. Aquaculture 1984, 36, 85–95. [Google Scholar] [CrossRef]
- Jenkins-Keeran, K.; Schreuders, P.; Edwards, K.; Woods, L.C. The Effects of Oxygen on the Short-Term Storage of Striped Bass Semen. N. Am. J. Aquac. 2001, 63, 238–241. [Google Scholar] [CrossRef]
- Parodi, J.; Guerra, G.; Cuevas, M.; Ramírez-Reveco, A.; Romero, F. Effects of Storage Time on the Motility, Mortality and Calcium Levels of Atlantic Salmon Salmo Salar Spermatozoa. J. Fish Biol. 2017, 90, 1506–1516. [Google Scholar] [CrossRef]
- Ubilla, A.; Fornari, D.; Figueroa, E.; Effer, B.; Valdebenito, I. Short-Term Cold Storage of the Semen of Rainbow Trout Oncorhynchus Mykiss (Walbaum, 1792) Incorporating DMSO in the Sperm Diluent. Effects on Motility and Fertilizing Capacity. Aquac. Res. 2015, 46, 37–44. [Google Scholar] [CrossRef]
- Trigo, P.; Merino, O.; Figueroa, E.; Valdebenito, I.; Sánchez, R.; Risopatrón, J. Effect of Short-Term Semen Storage in Salmon (Oncorhynchus mykiss) on Sperm Functional Parameters Evaluated by Flow Cytometry. Andrologia 2015, 47, 407–411. [Google Scholar] [CrossRef]
- Risopatrón, J.; Merino, O.; Cheuquemán, C.; Figueroa, E.; Sánchez, R.; Farías, J.G.; Valdebenito, I. Effect of the Age of Broodstock Males on Sperm Function during Cold Storage in the Trout (Oncorhynchus Mykiss). Andrologia 2018, 50, e12857. [Google Scholar] [CrossRef]
- Hassan, M.M.; Nahiduzzaman, M.; Al Mamun, S.N.; Taher, M.A.; Hossain, M.A.R. Fertilization by Refrigerator Stored Sperm of the Indian Major Carp, Labeo calbasu (Hamilton, 1822). Aquac. Res. 2013, 45, 150–158. [Google Scholar] [CrossRef]
- Sarosiek, B.; Dryl, K.; Kucharczyk, D.; Żarski, D.; Kowalski, R.K. Motility Parameters of Perch Spermatozoa (Perca fluviatilis L.) during Short-Term Storage with Antioxidants Addition. Aquac. Int. 2014, 22, 159–165. [Google Scholar] [CrossRef]
- Sarosiek, B.; Judycka, S.; Kowalski, R.K. Influence of Antioxidants on Spermatozoa in the Short-Term Storage of Salmonidae Milt. Polish J. Nat. Sci. 2013, 28, 379–384. [Google Scholar]
- Christen, R.; Gatti, J.-L.; Billard, R. Trout Sperm Motility: The Transient Movement of Trout Sperm Is Related to Changes in the Concentration of ATP Following the Activation of the Flagellar Movement. Eur. J. Biochem. 1987, 166, 667–671. [Google Scholar] [CrossRef]
- Gardiner, D.M. Utilization of Extracellular Glucose by Spermatozoa of Two Viviparous Fishes. Comp. Biochem. Physiol. Part A Physiol. 1978, 59, 165–168. [Google Scholar] [CrossRef]
- Babiak, I.; Ottesen, O.; Rudolfsen, G.; Johnsen, S. Chilled Storage of Semen from Atlantic Halibut, Hippoglossus Hippoglossus, L. I: Optimizing the Protocol. Theriogenology 2006, 66, 2025–2035. [Google Scholar] [CrossRef]
- Mansour, N.; Lahnsteiner, F.; Berger, B. Characterization of the Testicular Semen of the African Catfish, Clarias Gariepinus (Burchell, 1822), and Its Short-Term Storage. Aquac. Res. 2004, 35, 232–244. [Google Scholar] [CrossRef]
- Cabrita, E.; Sarasquete, C.; Martínez-Páramo, S.; Robles, V.; Beirão, J.; Pérez-Cerezales, S.; Herráez, M.P. Cryopreservation of Fish Sperm: Applications and Perspectives. J. Appl. Ichthyol. 2010, 26, 623–635. [Google Scholar] [CrossRef]
- Tiersch, T.R.; Goudie, C.A.; Carmichael, G.J. Cryopreservation of Channel Catfish Sperm: Storage in Cryoprotectants, Fertilization Trials, and Growth of Channel Catfish Produced with Cryopreserved Sperm. Trans. Am. Fish. Soc. 1994, 123, 580–586. [Google Scholar] [CrossRef]
- Chao, N.H.; Tsai, H.P.; Liao, I. Short and Long-Term Cryopreservation of Sperm and Sperm Suspension of the Grouper, Epinephelus Malabaricus (Bloch and Schneider). Asian Fish. Sci. 1992, 5, 103–116. [Google Scholar] [CrossRef]
- Chang, Y.J.; Chang, Y.J. Milt Properties of Four Flatfish Species and Fine Structure of Their Cryopreserved Spermatozoa. J. Fish. Sci. Technol. 2002, 5, 87–96. [Google Scholar] [CrossRef]
- Rahimi, R.; Hajirezaee, S.; Shaluei, F.; Kaboutari Katadj, J. Antibiotics, Penicillin and Sreptomycin Improve Semen Quality Indices of Endangered Caspian Brown Trout, Salmo Trutta Caspius (Kessler, 1870) during in Vitro Short-Term Storage. Aquac. Res. 2016, 47, 3662–3666. [Google Scholar] [CrossRef]
- Wild, T.; Kaurova, S.A.; Browne, R.K.; Uteshev, V.K. Antibiotics for the Refrigerated Storage at 4 °C of Hormonally Induced European Common Frog (Rana temporaria) Spermatozoa. Theriogenology Wild 2022, 1, 100009. [Google Scholar] [CrossRef]
- Stoss, J.; Refstie, T. Short-Term Storage and Cryopreservation of Milt from Atlantic Salmon and Sea Trout. Aquaculture 1983, 30, 229–236. [Google Scholar] [CrossRef]
- Jenkins, J.A.; Tiersch, T.R. A Preliminary Bacteriological Study of Refrigerated Channel Catfish Sperm. J. World Aquac. Soc. 1997, 28, 282–288. [Google Scholar] [CrossRef]
- Judycka, S.; Cejko, B.I.; Dryl, K.; Dobosz, S.; Grudniewska, J.; Kowalski, R.K. The Effect of Supplementation of a Trehalose-Based Extender with KCl on Rainbow Trout (Oncorhynchus mykiss) Sperm Freezability and Post-Thaw Motility. Aquaculture 2016, 465, 303–310. [Google Scholar] [CrossRef]
- Sarosiek, B.; Dryl, K.; Judycka, S.; Dobosz, S.; Grudniewska, J.; Kowalski, R.K. Cryopreservation Method for Whitefish (Coregonus lavaretus) Semen Possible for Use in Large-Scale Fertilization. Aquac. Res. 2016, 47, 4038–4042. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, X.; Wang, W.; Zhang, X.; Xu, S.; Ma, D.; Xiao, Z.; Xiao, Y.; Li, J. Effect of the Addition of Six Antioxidants on Sperm Motility, Membrane Integrity and Mitochondrial Function in Red Seabream (Pagrus major) Sperm Cryopreservation. Fish Physiol. Biochem. 2015, 41, 413–422. [Google Scholar] [CrossRef]
- Gheller, S.M.M.; Corcini, C.D.; de Brito, C.R.C.; Acosta, I.B.; Tavares, G.C.; Soares, S.L.; Silva, A.C.; Pires, D.M.; Varela Junior, A.S. Use of Trehalose in the Semen Cryopreservation of Amazonian Catfish Leiarius Marmoratus. Cryobiology 2019, 87, 74–77. [Google Scholar] [CrossRef]
- Aisen, E.; Quintana, M.; Medina, V.; Morello, H.; Venturino, A. Ultramicroscopic and Biochemical Changes in Ram Spermatozoa Cryopreserved with Trehalose-Based Hypertonic Extenders. Cryobiology 2005, 50, 239–249. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Ding, W.; Wang, Y. Mito-TEMPO Alleviates Renal Fibrosis by Reducing Inflammation, Mitochondrial Dysfunction, and Endoplasmic Reticulum Stress. Oxid. Med. Cell. Longev. 2018, 2018, 5828120. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Pesini, E.; Díez-Sánchez, C.; López-Pérez, M.J.; Enríquez, J.A. The Role of the Mitochondrion in Sperm Function: Is There a Place for Oxidative Phosphorylation or Is This a Purely Glycolytic Process? Curr. Top. Dev. Biol. 2007, 77, 3–19. [Google Scholar] [CrossRef]
- Masoudi, R.; Asadzadeh, N.; Sharafi, M. The Mitochondria-Targeted Antioxidant Mito-TEMPO Conserves Rooster’s Cooled Semen Quality and Fertility Potential. Theriogenology 2020, 156, 236–241. [Google Scholar] [CrossRef]
- Dickinson, D.A.; Forman, H.J. Cellular Glutathione and Thiols Metabolism. Biochem. Pharmacol. 2002, 64, 1019–1026. [Google Scholar] [CrossRef]
- Pereira, F.A.; Corcini, C.D.; Da Silva, A.C.; Pires, D.M.; Pereira, J.R.; Gheller, S.M.M.; Streit, D.P.; Maria, A.N.; Varela, A.S. Reduced Glutathione and ATP in the Seminal Cryopreservation of Tambaqui. Cryo-Letters 2018, 39, 371–379. [Google Scholar]
- Masoudi, R.; Sharafi, M.; Shahneh, A.Z.; Khodaei-Motlagh, M. Effects of Reduced Glutathione on the Quality of Rooster Sperm during Cryopreservation. Theriogenology 2019, 128, 149–155. [Google Scholar] [CrossRef]
- Estrada, E.; Rivera del Álamo, M.M.; Rodríguez-Gil, J.E.; Yeste, M. The Addition of Reduced Glutathione to Cryopreservation Media Induces Changes in the Structure of Motile Subpopulations of Frozen-Thawed Boar Sperm. Cryobiology 2017, 78, 56–64. [Google Scholar] [CrossRef]
- Silva, S.V.; Soares, A.T.; Batista, A.M.; Almeida, F.C.; Nunes, J.F.; Peixoto, C.A.; Guerra, M.M.P. In Vitro and In Vivo Evaluation of Ram Sperm Frozen in Tris Egg-Yolk and Supplemented with Superoxide Dismutase and Reduced Glutathione. Reprod. Domest. Anim. 2011, 46, 874–881. [Google Scholar] [CrossRef]
- Angrimani, D.S.R.; Nichi, M.; Brito, M.M.; Kawai, G.K.V.; Rui, B.R.; Losano, J.D.A.; Vieira, N.M.G.; Francischini, M.C.P.; Cruz, D.S.G.; Queiroz-Hazarbassanov, N.; et al. The Use of Reduced Glutathione (GSH) as Antioxidant for Cryopreserved Sperm in Dogs. Arq. Bras. Med. Veterinária Zootec. 2018, 70, 419–428. [Google Scholar] [CrossRef]

















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zidni, I.; Lee, H.-B.; Yoon, J.-H.; Park, J.-Y.; Jang, H.-S.; Co, Y.-S.; Pratiwi, D.Y.; Lim, H.-K. Intermediate-Term Storage of Spotted Halibut (Verasper variegatus) Sperm: Effects of Storage Methods, Extenders Supplemented with Antibiotics and Antioxidants on Sperm Quality. Antioxidants 2023, 12, 122. https://doi.org/10.3390/antiox12010122
Zidni I, Lee H-B, Yoon J-H, Park J-Y, Jang H-S, Co Y-S, Pratiwi DY, Lim H-K. Intermediate-Term Storage of Spotted Halibut (Verasper variegatus) Sperm: Effects of Storage Methods, Extenders Supplemented with Antibiotics and Antioxidants on Sperm Quality. Antioxidants. 2023; 12(1):122. https://doi.org/10.3390/antiox12010122
Chicago/Turabian StyleZidni, Irfan, Hyo-Bin Lee, Ji-Hye Yoon, Jung-Yeol Park, Hyun-Seok Jang, Youn-Su Co, Dian Yuni Pratiwi, and Han-Kyu Lim. 2023. "Intermediate-Term Storage of Spotted Halibut (Verasper variegatus) Sperm: Effects of Storage Methods, Extenders Supplemented with Antibiotics and Antioxidants on Sperm Quality" Antioxidants 12, no. 1: 122. https://doi.org/10.3390/antiox12010122