
Glibenclamide Increases Nitric Oxide Levels and Decreases Oxidative Stress in an In Vitro Model of Preeclampsia

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Plasma Collection and Processing
2.3. Culture of HUVEC
2.4. Cell Viability Analysis of HUVECs against Incubation with Plasma and Glibenclamide
2.5. Evaluation of Cellular Production of Nitric Oxide (NO) and Nitrite/Nitrate (Total NOx) in the Cell Culture Supernatant
2.6. Reactive Oxygen Species (ROS)
2.7. Assessment of Lipid Peroxidation in Supernatants
2.8. Antioxidant Capacity
2.9. Statistical Analysis
3. Results
3.1. Clinical Parameters of Patients Used to Constitute Pooled Plasma
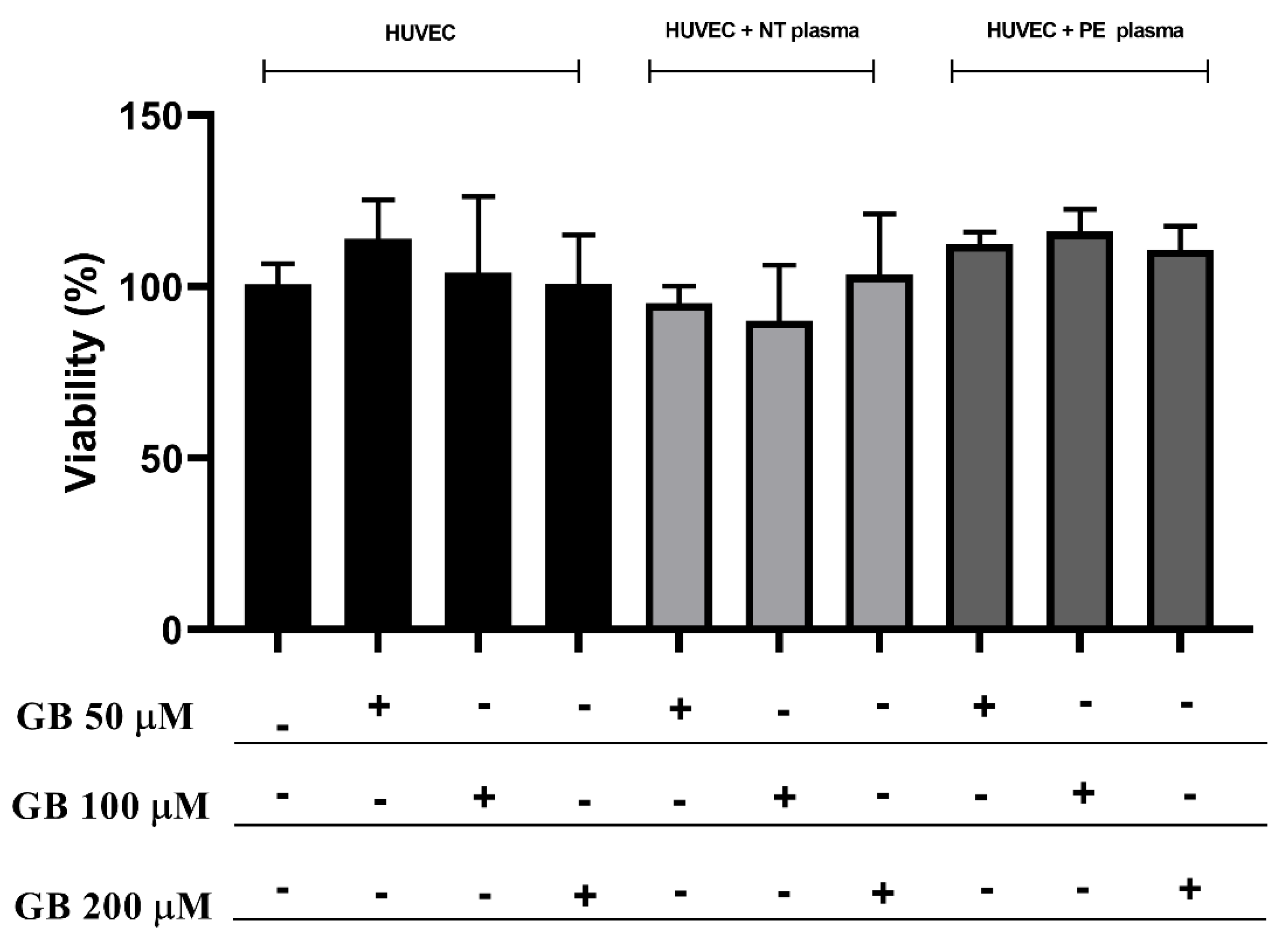
3.2. HUVEC Incubated with Plasma and Glibenclamide did Not Show Differences in Viability
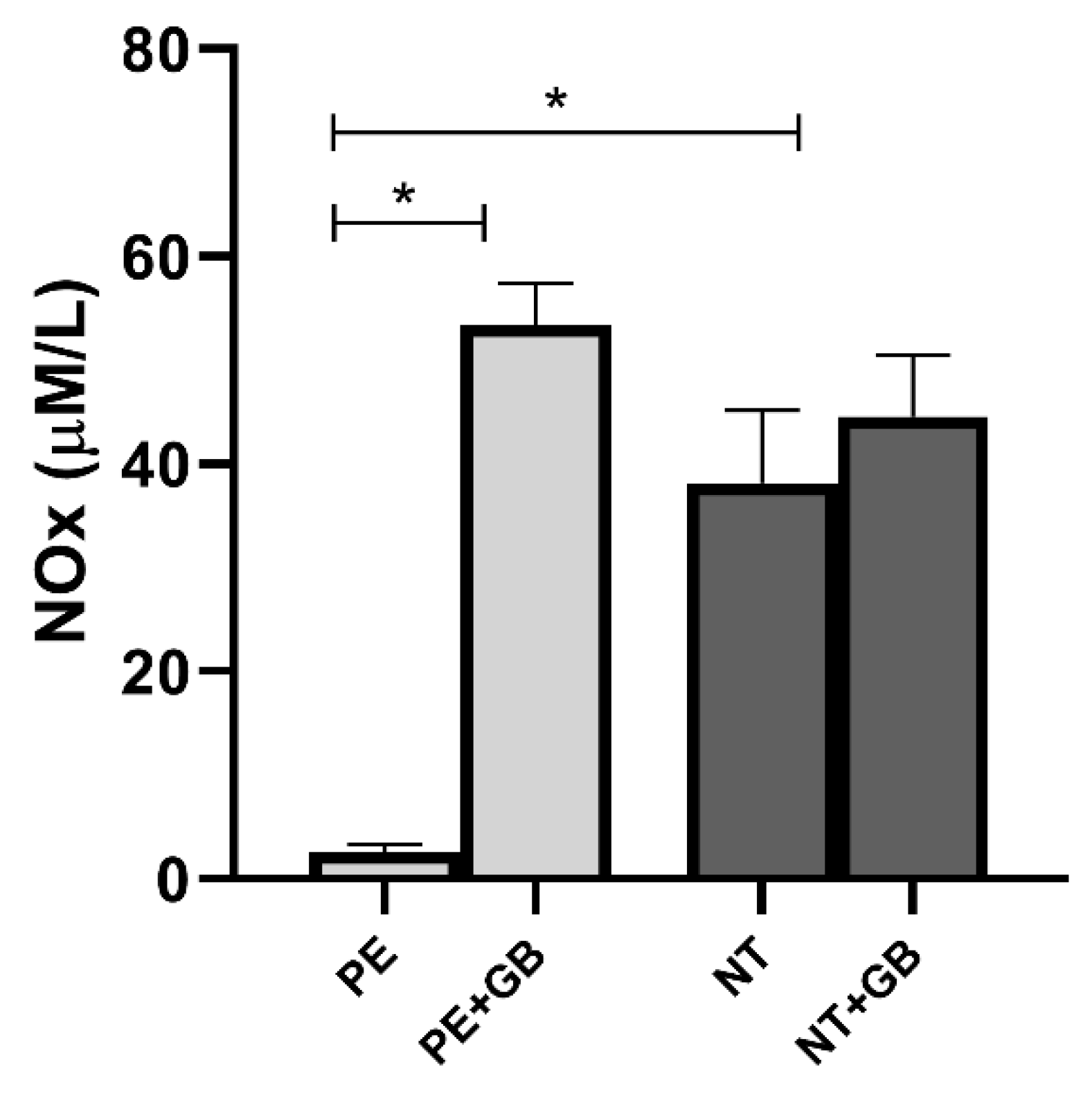
3.3. GB Increases NO Production in HUVEC Incubated with Plasma from PE and NT Pregnant Women and Nox in Cell Supernatant
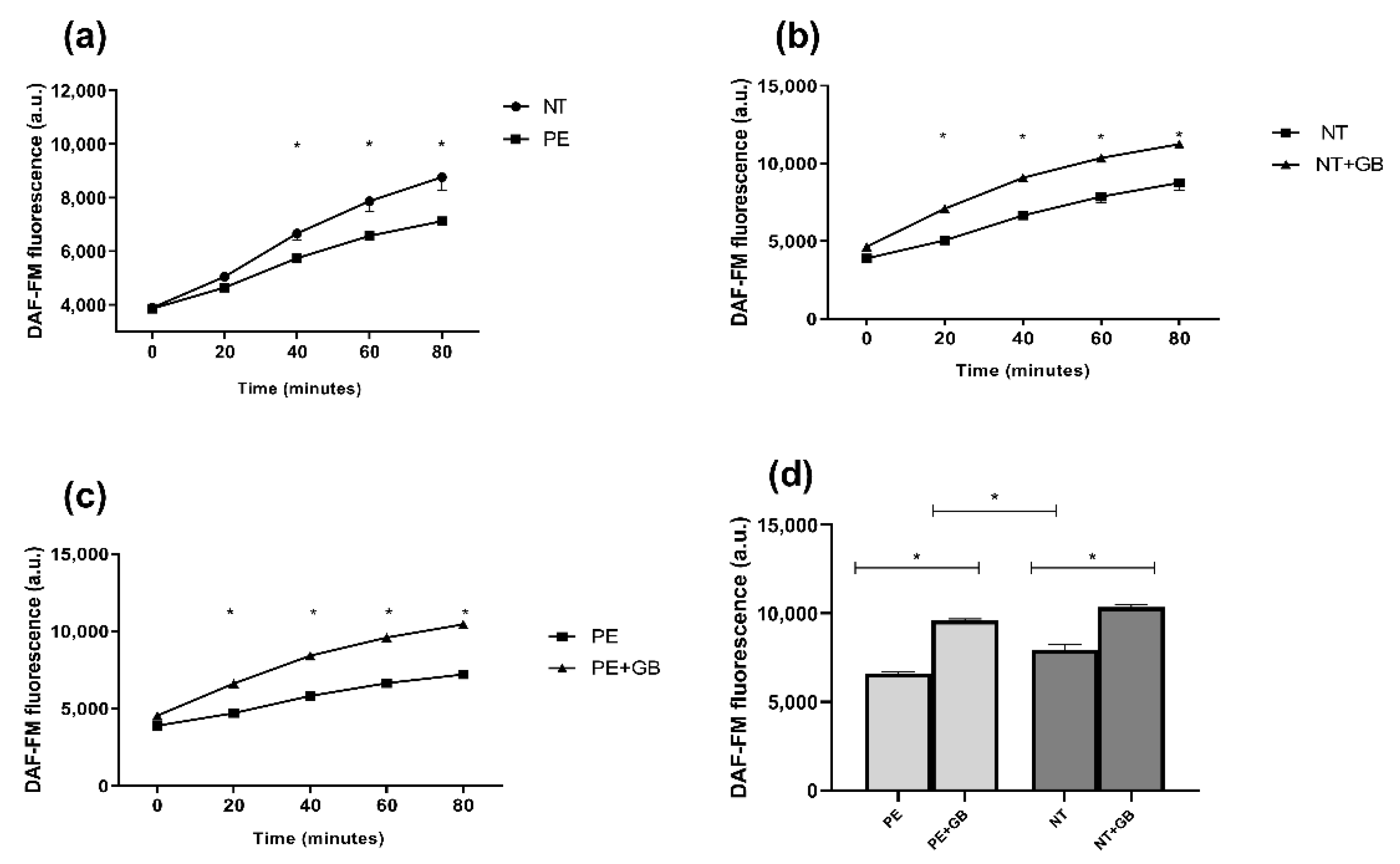
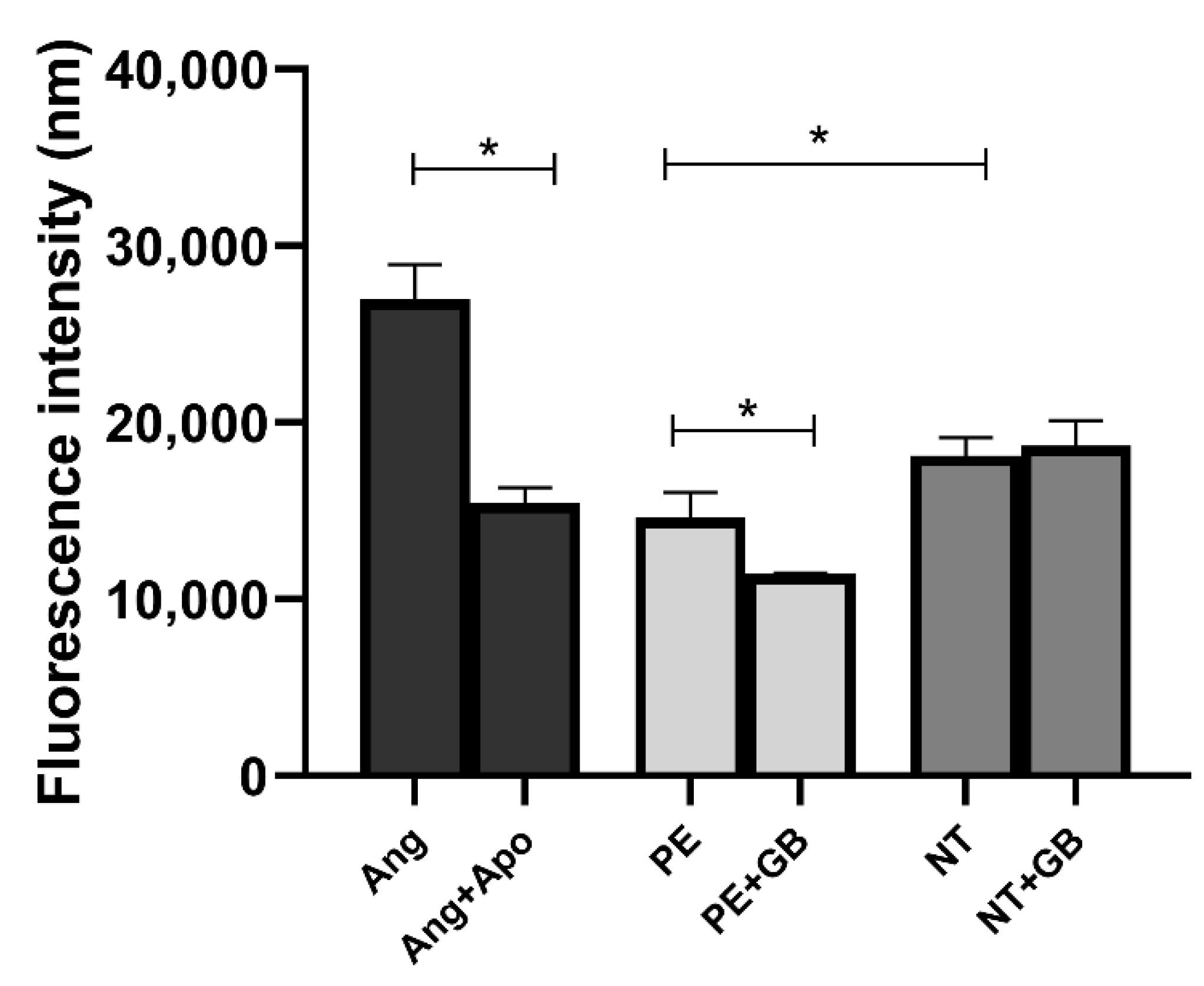
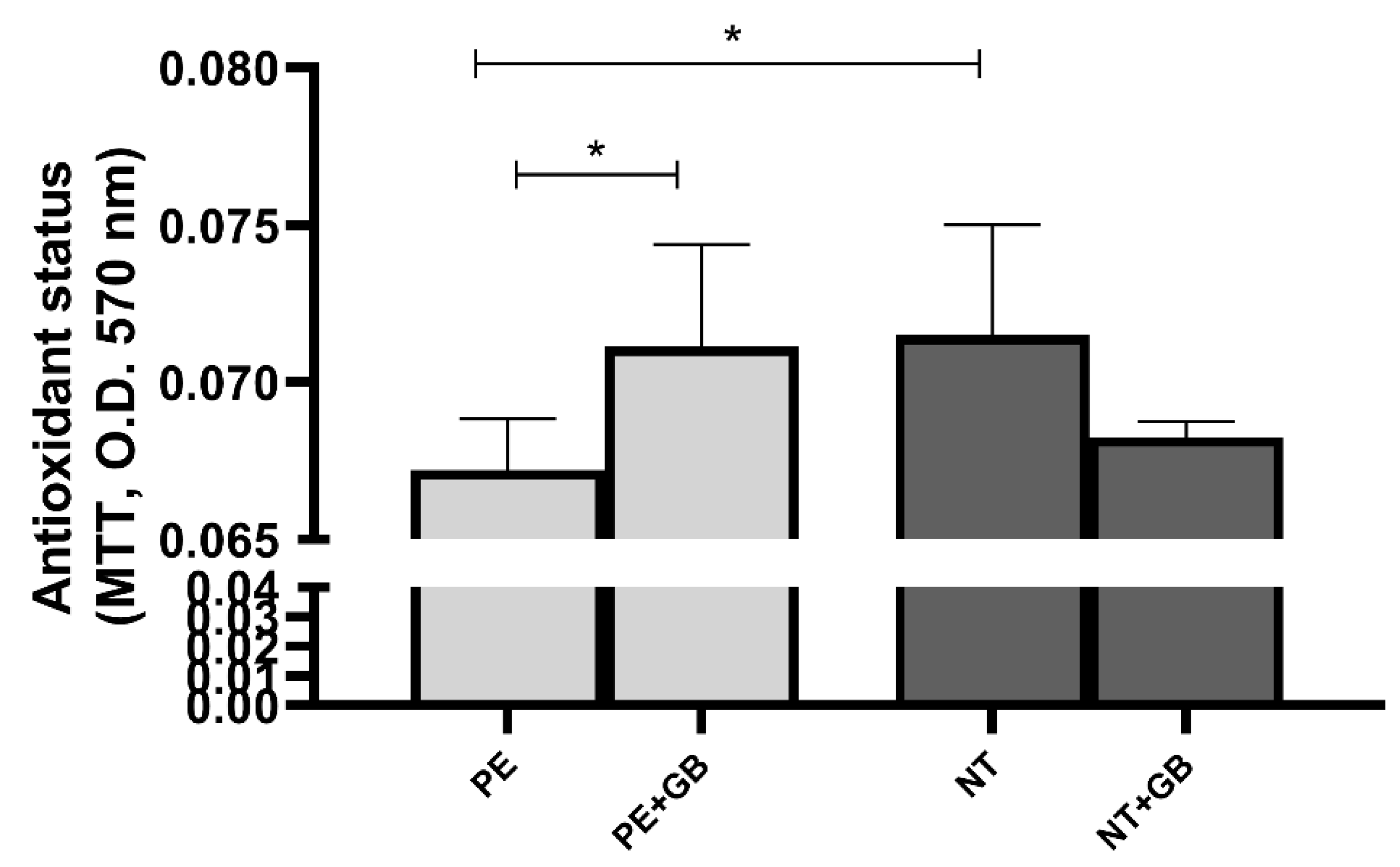
3.4. ROS Levels Are Elevated in Cells Treated with Plasma from NT, and GB Can Reduce Oxidative Stress in Cells Incubated for 30 min with PE Plasma
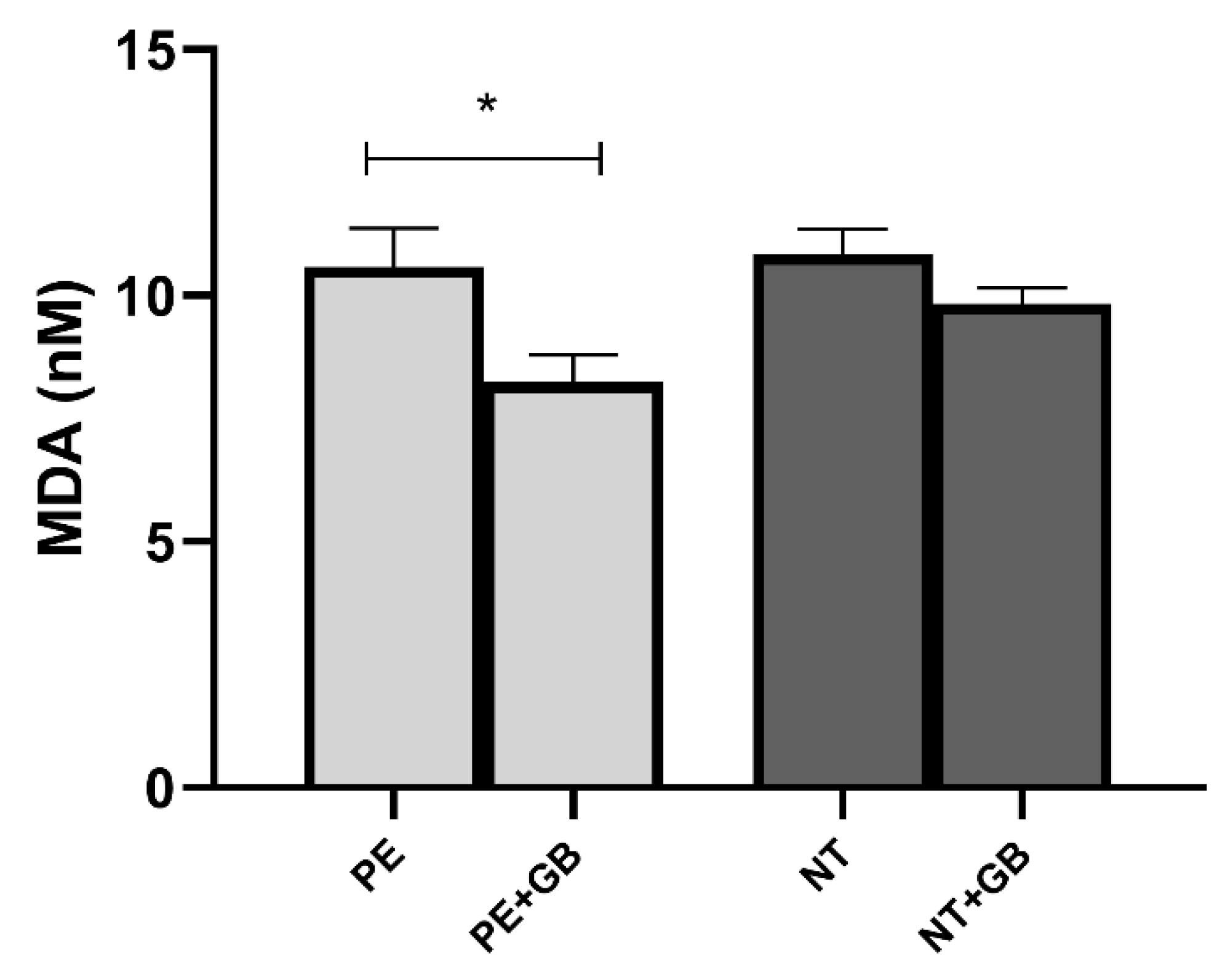
3.5. Supernatant Levels of MDA Indicate a Decrease in Lipid Peroxidation in GB-Treated Cells of the PE Group
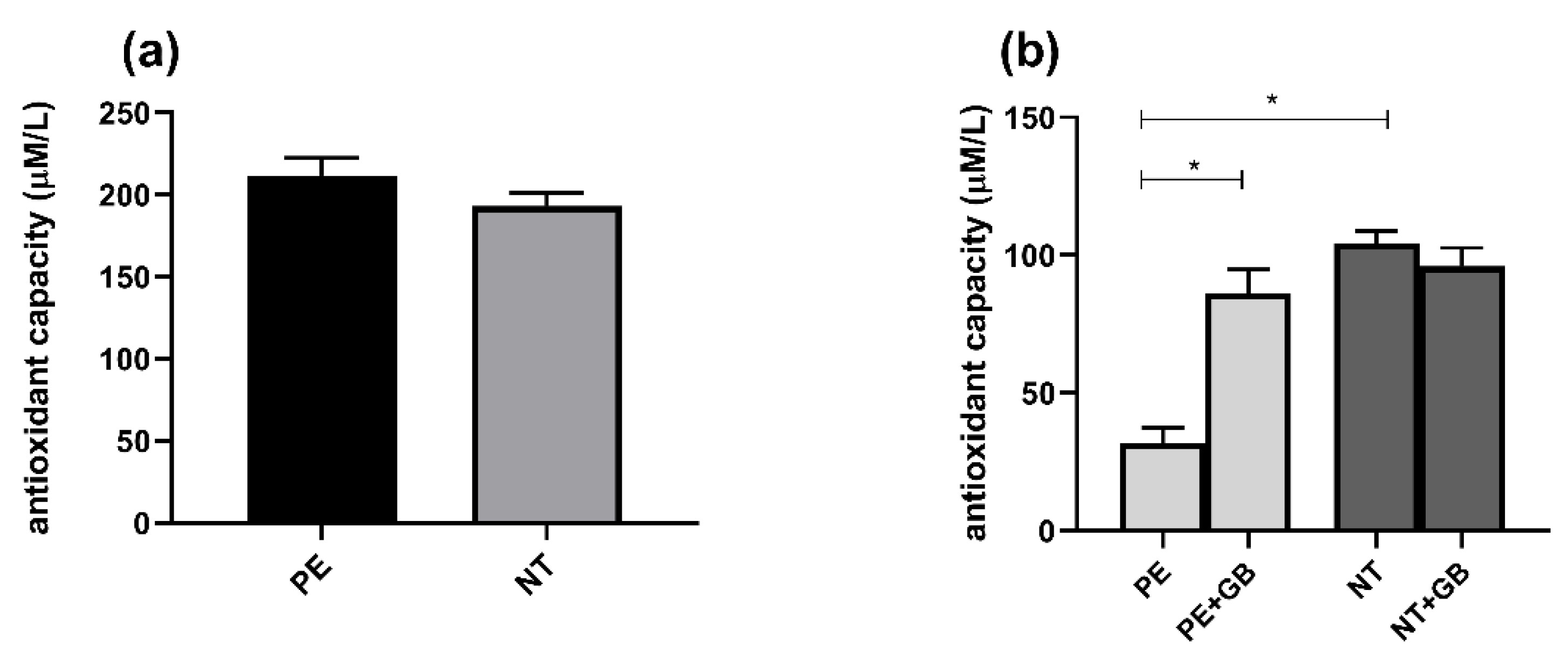
3.6. Plasma from Pregnant Women did Not Differ in FRAP Activity, but Supernatant from PE Women Showed Higher Antioxidant Power, while GB Decreased this Response
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American College of Obstetricians and Gynecologists. ACOG Practice Bulletins Clinical Management Guidelines for Obstetrician—Gynecologists. Obstet. Gynecol. 2020, 133, 168–186. [Google Scholar]
- Redman, C.W.; Sargent, I.L. Latest advances in understanding preeclampsia. Science 2005, 308, 1592–1594. [Google Scholar] [CrossRef] [PubMed]
- Fantone, S.; Mazzucchelli, R.; Giannubilo, S.R.; Ciavattini, A.; Marzioni, D.; Tossetta, G. AT-rich interactive domain 1A protein expression in normal and pathological pregnancies complicated by preeclampsia. Histochem. Cell Biol. 2020, 154, 339–346. [Google Scholar] [CrossRef]
- Brennan, L.J.; Morton, J.S.; Davidge, S.T. Vascular dysfunction in preeclampsia. Microcirculation 2014, 21, 4–14. [Google Scholar] [CrossRef]
- Goulopoulou, S.; Davidge, S.T. Molecular mechanisms of maternal vascular dysfunction in preeclampsia. Trends Mol. Med. 2015, 21, 88–97. [Google Scholar] [CrossRef]
- Roberts, J.M. Endothelial dysfunction in preeclampsia. In Seminars in Reproductive Endocrinology; Thieme Medical Publishers, Inc.: New York, NY, USA, 1998; Volume 16, pp. 5–15. [Google Scholar]
- Khalil, R.A. Endothelium; Academic Press: Cambridge, UK, 2016; ISBN 0128044152. [Google Scholar]
- Echeverri, I.; Ortega-Ávila, J.G.; Mosquera, M.; Castillo, A.; Jiménez, E.; Suárez-Ortegon, M.F.; Mateus, J.C.; Aguilar-de Plata, C. Relationship between maternal and newborn endothelial function and oxidative stress. Am. J. Hum. Biol. 2015, 27, 822–831. [Google Scholar] [CrossRef]
- He, Y.; Hara, H.; Núñez, G. Mechanism and regulation of NLRP3 inflammasome activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abais, J.M.; Xia, M.; Zhang, Y.; Boini, K.M.; Li, P.-L. Redox regulation of NLRP3 inflammasomes: ROS as trigger or effector? Antioxid. Redox Signal. 2015, 22, 1111–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shihata, W.A.; Michell, D.L.; Andrews, K.L.; Chin-Dusting, J.P.F. Caveolae: A role in endothelial inflammation and mechanotransduction? Front. Physiol. 2016, 7, 628. [Google Scholar] [CrossRef] [Green Version]
- Lamkanfi, M.; Kanneganti, T.; Franchi, L.; Núñez, G. Caspase-1 inflammasomes in infection and inflammation. J. Leukoc. Biol. 2007, 82, 220–225. [Google Scholar] [CrossRef] [Green Version]
- Zahid, A.; Li, B.; Kombe, A.J.K.; Jin, T.; Tao, J. Pharmacological inhibitors of the nlrp3 inflammasome. Front. Immunol. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.; Dong, Q.; Song, Z.; Shen, F.; Shi, J.; Li, Y. NLRP3 inflammasome: A promising target in ischemic stroke. Inflamm. Res. 2017, 66, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Satoh, T.; Kambe, N.; Matsue, H. NLRP3 activation induces ASC-dependent programmed necrotic cell death, which leads to neutrophilic inflammation. Cell Death Dis. 2013, 4, e644. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Shen, G.; Su, Z.; He, Z.; Yuan, L. Glibenclamide ameliorates the disrupted blood–brain barrier in experimental intracerebral hemorrhage by inhibiting the activation of NLRP3 inflammasome. Brain Behav. 2019, 9, e01254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Lin, X.; Zhang, S.; Xiu, H.; Pan, C.; Cui, W. A protective role of glibenclamide in inflammation-associated injury. Mediators Inflamm. 2017, 2017, 3578702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.-J.; Jin, Y.; Zhang, W.-Y.; Li, L.; Liu, X.-W. Role of AKR1C3 in renal injury and glibenclamide is anti-inflammatory in preeclamptic rats. Gene 2018, 662, 1–9. [Google Scholar] [CrossRef]
- Langer, O.; Yogev, Y.; Xenakis, E.M.J.; Rosenn, B. Insulin and glyburide therapy: Dosage, severity level of gestational diabetes, and pregnancy outcome. Am. J. Obstet. Gynecol. 2005, 192, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.; Yu, G.; Smith, L. 993: Adverse pregnancy outcomes with glyburide vs insulin among patients with gestational diabetes established by the International Association of Diabetes and Pregnancy Study Group (IADPSG). Am. J. Obstet. Gynecol. 2018, 218, S586. [Google Scholar] [CrossRef]
- Rocha-Penha, L.; Caldeira-Dias, M.; Tanus-Santos, J.E.; de Carvalho Cavalli, R.; Sandrim, V.C. Myeloperoxidase in hypertensive disorders of pregnancy and its relation with nitric oxide. Hypertension 2017, 69, 1173–1180. [Google Scholar] [CrossRef]
- Uchiyama, M.; Mihara, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moncada, S. Nitric oxide: Physiology, pathophysiology and pharmacology. Pharmacol. Rev. 1991, 43, 109–142. [Google Scholar]
- Krause, B.J.; Hanson, M.A.; Casanello, P. Role of nitric oxide in placental vascular development and function. Placenta 2011, 32, 797–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poniedzialek-Czajkowska, E.; Marciniak, B.; Kimber-Trojnar, Z.; Leszczynska-Gorzelak, B.; Oleszczuk, J. Nitric oxide in normal and preeclamptic pregnancy. Curr. Pharm. Biotechnol. 2011, 12, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Farrow, K.N.; Lakshminrusimha, S.; Reda, W.J.; Wedgwood, S.; Czech, L.; Gugino, S.F.; Davis, J.M.; Russell, J.A.; Steinhorn, R.H. Superoxide dismutase restores eNOS expression and function in resistance pulmonary arteries from neonatal lambs with persistent pulmonary hypertension. Am. J. Physiol. Cell. Mol. Physiol. 2008, 295, L979–L987. [Google Scholar] [CrossRef] [Green Version]
- Toescu, V.; Nuttall, S.L.; Martin, U.; Kendall, M.J.; Dunne, F. Oxidative stress and normal pregnancy. Clin. Endocrinol. 2002, 57, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Duhig, K.; Chappell, L.C.; Shennan, A.H. Oxidative stress in pregnancy and reproduction. Obstet. Med. 2016, 9, 113–116. [Google Scholar] [CrossRef] [Green Version]
- Dennery, P.A. Oxidative stress in development: Nature or nurture? Free Radic. Biol. Med. 2010, 49, 1147–1151. [Google Scholar] [CrossRef]
- Gomes, H.F.; Palei, A.C.T.; Machado, J.S.R.; Da Silva, L.M.; Montenegro, M.F.; Jordão, A.A.; Duarte, G.; Tanus-Santos, J.E.; Cavalli, R.D.C.; Sandrim, V.C. Assessment of oxidative status markers and NO bioavailability in hypertensive disorders of pregnancy. J. Hum. Hypertens. 2013, 27, 345–348. [Google Scholar] [CrossRef]
- Erejuwa, O.O.; Sulaiman, S.A.; Wahab, M.S.A.; Salam, S.K.N.; Salleh, M.S.M.; Gurtu, S. Antioxidant protective effect of glibenclamide and metformin in combination with honey in pancreas of streptozotocin-induced diabetic rats. Int. J. Mol. Sci. 2010, 11, 2056–2066. [Google Scholar] [CrossRef] [Green Version]
- Chukwunonso Obi, B.; Chinwuba Okoye, T.; Okpashi, V.E.; Nonye Igwe, C.; Olisah Alumanah, E. Comparative study of the antioxidant effects of metformin, glibenclamide, and repaglinide in alloxan-induced diabetic rats. J. Diabetes Res. 2016, 2016, 1635361. [Google Scholar] [CrossRef] [Green Version]
- Okayama, N.; Coe, L.; Itoh, M.; Alexander, J.S. Intracellular mechanisms of nitric oxide plus hydrogen peroxide-mediated neutrophil adherence to cultured human endothelial cells. Inflamm. Res. 1998, 47, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Murugan, D.D.; Md Zain, Z.; Choy, K.W.; Zamakshshari, N.H.; Choong, M.J.; Lim, Y.M.; Mustafa, M.R. Edible bird’s nest protects against hyperglycemia-induced oxidative stress and endothelial dysfunction. Front. Pharmacol. 2020, 10, 1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, B.; Cui, Y.; Zhao, J.; Yu, F.; Zhu, Y.; Xu, G.; Zhang, Z.; Tang, C.; Du, J. Hydrogen sulfide downregulates the aortic L-arginine/nitric oxide pathway in rats. Am. J. Physiol. Integr. Comp. Physiol. 2007, 293, R1608–R1618. [Google Scholar] [CrossRef]
- Li, C.; Gao, C.; Zhang, X.; Zhang, L.; Shi, H.; Jia, X. Comparison of the effectiveness and safety of insulin and oral hypoglycemic drugs in the treatment of gestational diabetes mellitus: A meta-analysis of 26 randomized controlled trials. Gynecol. Endocrinol. 2022, 38, 303–309. [Google Scholar] [CrossRef]
- Caritis, S.N.; Hebert, M.F. A pharmacologic approach to the use of glyburide in pregnancy. Obstet. Gynecol. 2013, 121, 1309–1312. [Google Scholar] [CrossRef] [Green Version]
- Moretti, M.E.; Rezvani, M.; Koren, G. Safety of glyburide for gestational diabetes: A meta-analysis of pregnancy outcomes. Ann. Pharmacother. 2008, 42, 483–490. [Google Scholar] [CrossRef]
- Sarkhail, P.; Abdollahi, M.; Fadayevatan, S.; Shafiee, A.; Mohammadirad, A.; Dehghan, G.; Esmaily, H.; Amin, G. Effect of Phlomis persica on glucose levels and hepatic enzymatic antioxidants in streptozotocin-induced diabetic rats. Pharmacogn. Mag. 2010, 6, 219–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moniruzzaman, M.; Rokeya, B.; Ahmed, S.; Bhowmik, A.; Khalil, M.I.; Gan, S.H. In vitro antioxidant effects of Aloe barbadensis Miller extracts and the potential role of these extracts as antidiabetic and antilipidemic agents on streptozotocin-induced type 2 diabetic model rats. Molecules 2012, 17, 12851–12867. [Google Scholar] [CrossRef] [PubMed]
- Alabi, T.D.; Chegou, N.N.; Brooks, N.L.; Oguntibeju, O.O. Effects of anchomanes difformis on inflammation, apoptosis, and organ toxicity in STZ-induced diabetic cardiomyopathy. Biomedicines 2020, 8, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qasem, M.A.; Noordin, M.I.; Arya, A.; Alsalahi, A.; Jayash, S.N. Evaluation of the glycemic effect of Ceratonia siliqua pods (Carob) on a streptozotocin-nicotinamide induced diabetic rat model. PeerJ 2018, 6, e4788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, A.S.; Schaefer, M.; Korte, T.; Herrmann, A.; Tannert, A. HaCaT keratinocytes exhibit a cholesterol and plasma membrane viscosity gradient during directed migration. Exp. Cell Res. 2012, 318, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-P.; Jiang, Y.; Yang, H.; Peng, C.; Zhang, C.; Tao, X.; Xie, H.-H. Combination therapy of nifedipine and sulphonylureas exhibits a mutual antagonistic effect on the endothelial cell dysfunction induced by hyperglycemia linked to vascular disease. Cell. Physiol. Biochem. 2016, 38, 2337–2347. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Normotensive Pregnant | Preeclampsia | P |
|---|---|---|---|
| (n = 10) | (n = 10) | (<0.05) | |
| Age (years) | 26.72 ± 0.71 | 27.32 ± 0.58 | 0.5177 |
| Race Caucasian | 8 | 9 | >0.9999 |
| Non-caucasian | 2 | 1 | >0.9999 |
| GAS (weeks) | 34.33 ± 0.35 | 31.87 ± 0.52 | 0.0001 |
| SBP (mm/Hg) | 115.70 ± 0.70 | 155.80 ± 0.20 | <0.0001 |
| DPB (mm/Hg) | 73.30 ± 0.10 | 103.30 ± 0.20 | <0.0001 |
| 24-h Pr (mg) | ND | 1857 ± 539.40 | - |
| Uric acid (mg/dL) | ND | 5.15 ± 0.20 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nunes, P.R.; Bueno Pereira, T.O.; Bertozzi Matheus, M.; Grandini, N.A.; Siqueira, J.S.; Correa, C.R.; Abbade, J.F.; Sandrim, V.C. Glibenclamide Increases Nitric Oxide Levels and Decreases Oxidative Stress in an In Vitro Model of Preeclampsia. Antioxidants 2022, 11, 1620. https://doi.org/10.3390/antiox11081620
Nunes PR, Bueno Pereira TO, Bertozzi Matheus M, Grandini NA, Siqueira JS, Correa CR, Abbade JF, Sandrim VC. Glibenclamide Increases Nitric Oxide Levels and Decreases Oxidative Stress in an In Vitro Model of Preeclampsia. Antioxidants. 2022; 11(8):1620. https://doi.org/10.3390/antiox11081620
Chicago/Turabian StyleNunes, Priscila Rezeck, Thaina Omia Bueno Pereira, Mariana Bertozzi Matheus, Nubia Alves Grandini, Juliana Silva Siqueira, Camila Renata Correa, Joelcio Francisco Abbade, and Valeria Cristina Sandrim. 2022. "Glibenclamide Increases Nitric Oxide Levels and Decreases Oxidative Stress in an In Vitro Model of Preeclampsia" Antioxidants 11, no. 8: 1620. https://doi.org/10.3390/antiox11081620