
Evaluation of Plant Protein Hydrolysates as Natural Antioxidants in Fish Oil-In-Water Emulsions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Meals and Enzymes
2.2. Enzymatic Hydrolysis
2.3. Plant Meal Solubilization and Protein Recovery
2.4. Characterization of the Hydrolysates
2.4.1. Proximate Composition of the Hydrolysates
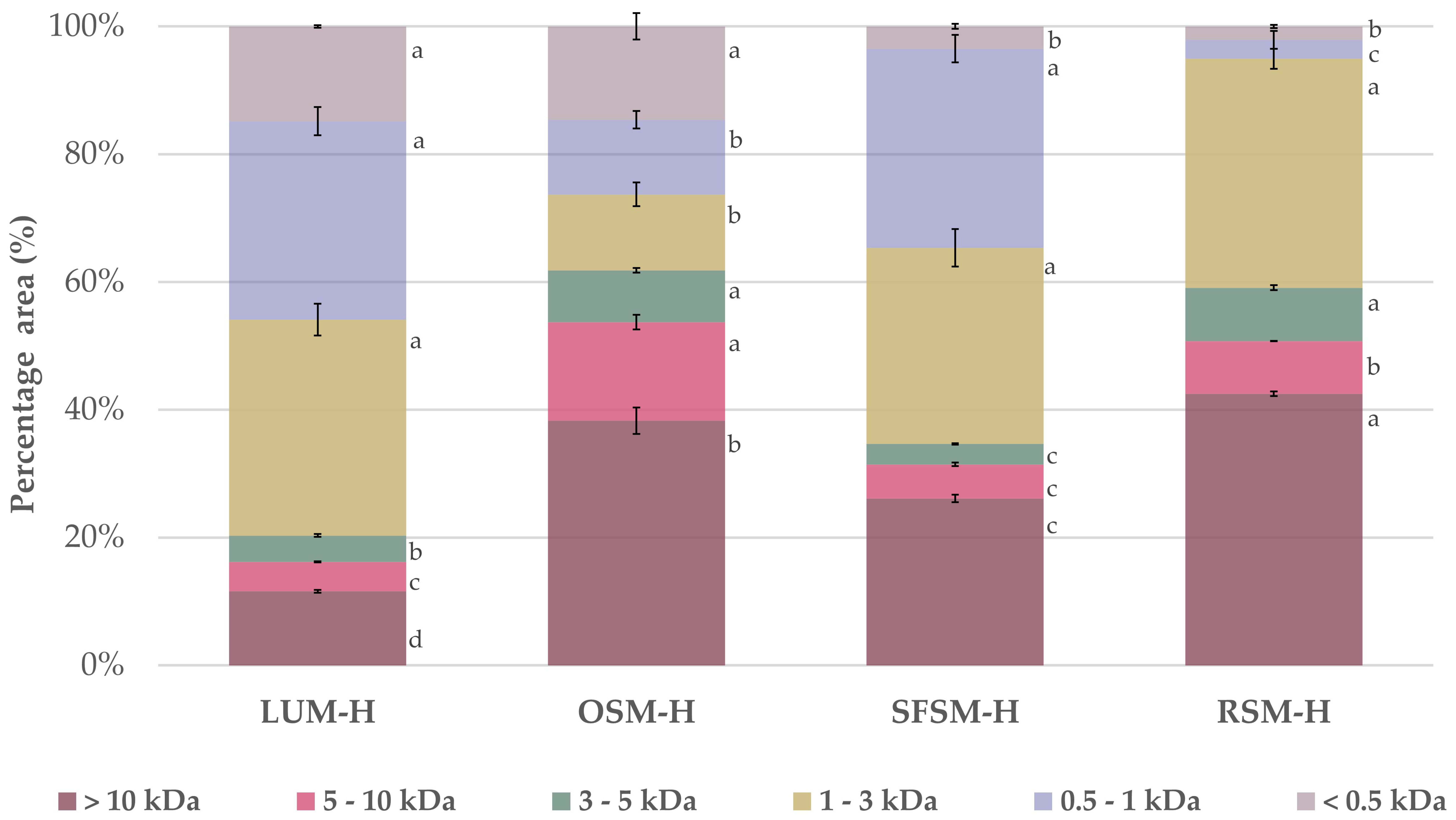
2.4.2. Molecular Weight Distribution of the Hydrolysates
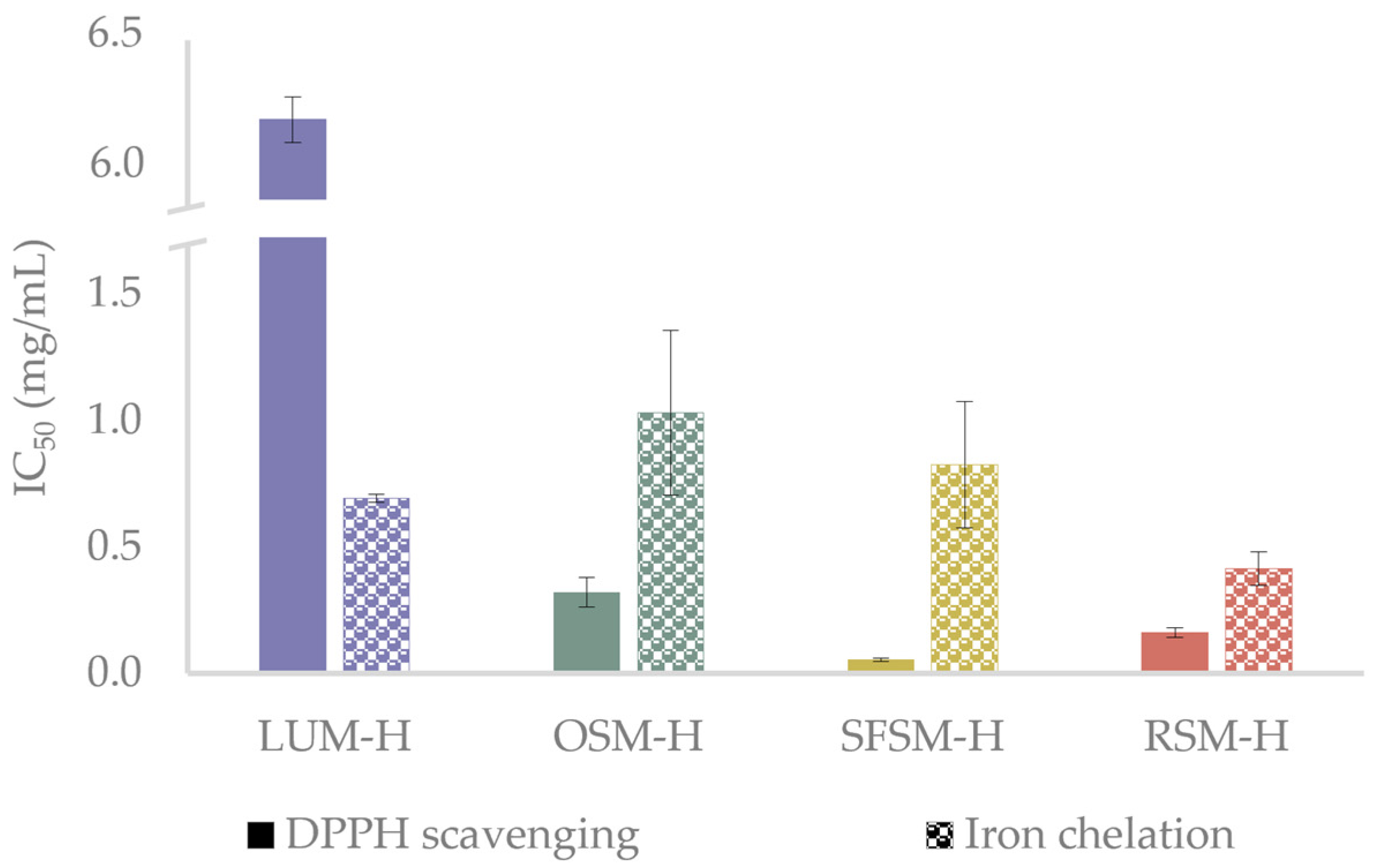
2.4.3. Antioxidant In Vitro Activity of Hydrolysates
2.5. Preparation of Emulsions
2.6. Physical Stability of the Emulsions
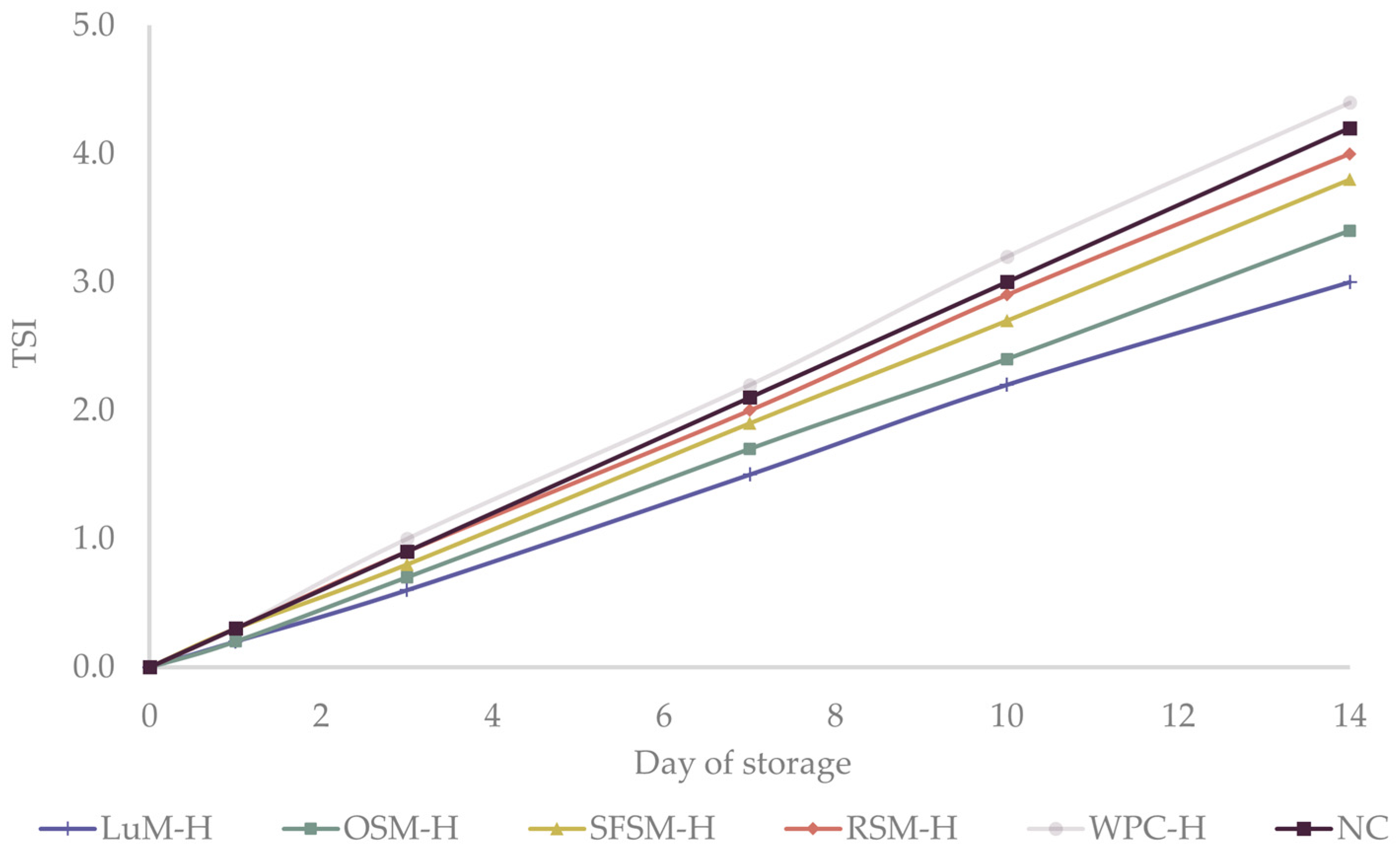
2.7. Oxidative Stability of the Emulsions
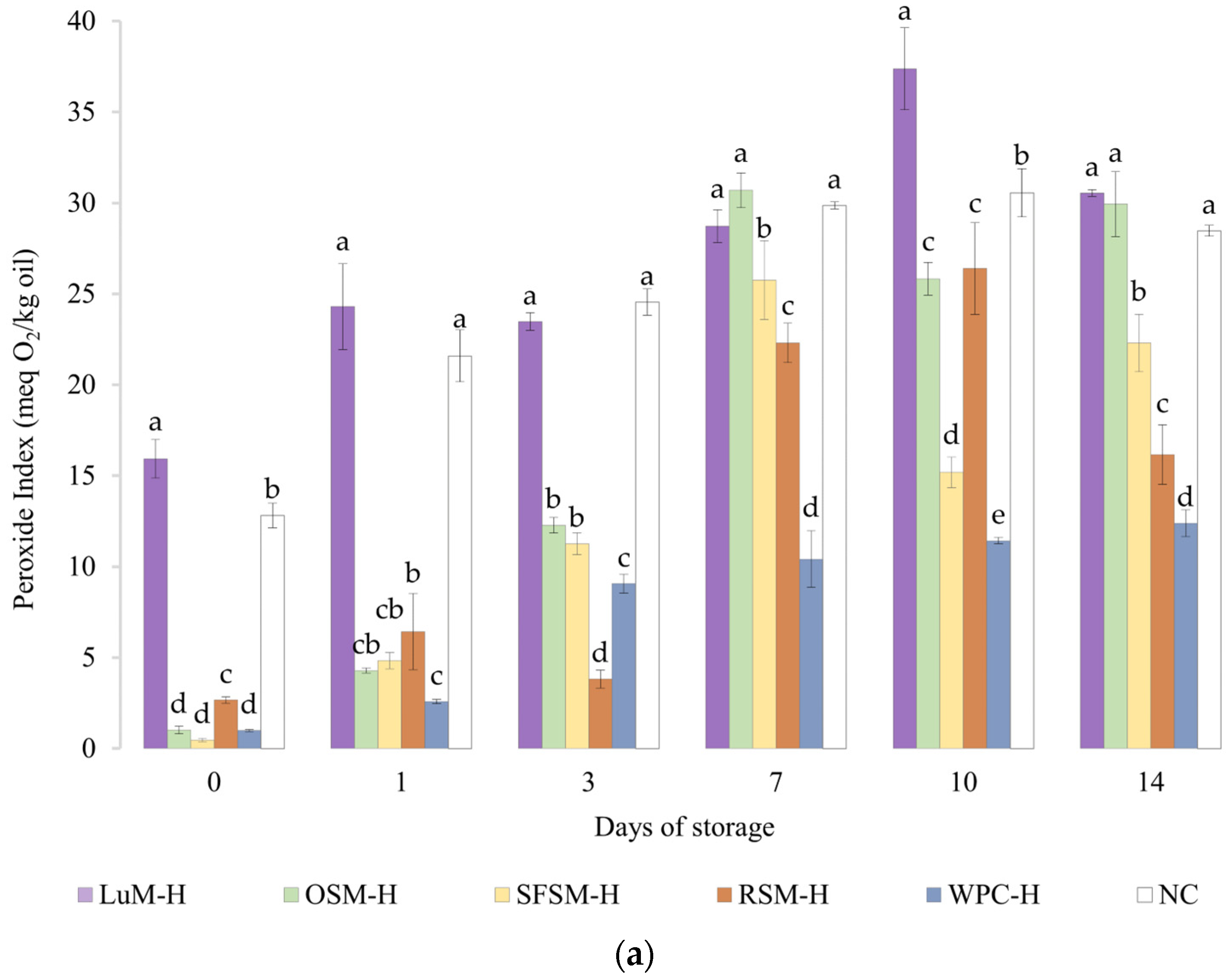
2.7.1. Determination of the Peroxide Value
2.7.2. Anisidine Value Assay
2.8. Statistical Analysis
3. Results and Discussion
3.1. Characterization of the Hydrolysates
3.2. Antioxidant In Vitro Activity of Hydrolysates
3.3. Physical Stability of Emulsions
3.4. Oxidative Stability of Emulsions
3.4.1. Peroxide Value
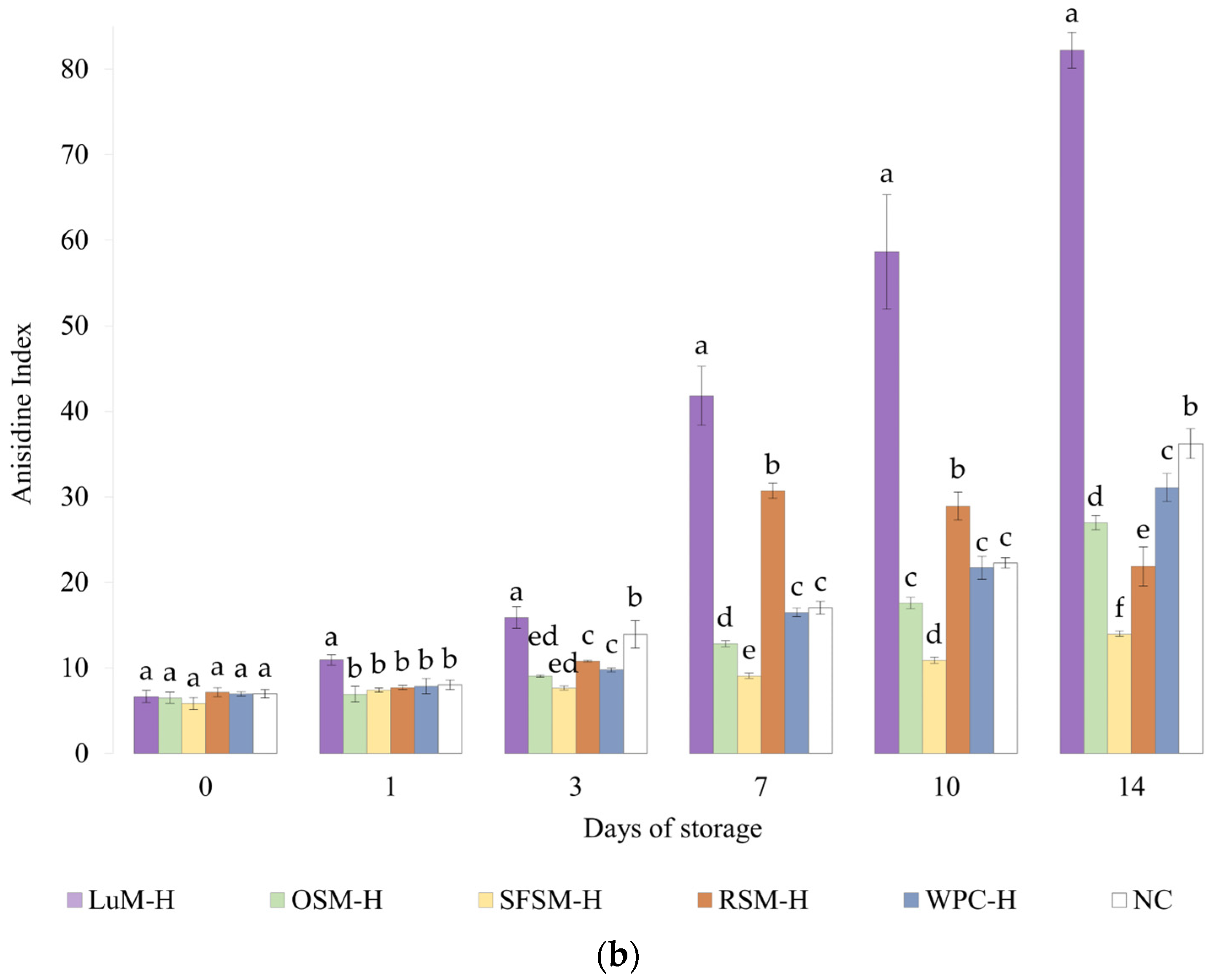
3.4.2. Anisidine Value
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elagizi, A.; Lavie, C.J.; Marshall, K.; DiNicolantonio, J.J.; O’Keefe, J.H.; Milani, R.V. Omega-3 Polyunsaturated Fatty Acids and Cardiovascular Health: A Comprehensive Review. Prog. Cardiovasc. Dis. 2018, 61, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Fang, Z.; Zhang, T.; Chen, Y. Polyunsaturated Fatty Acid Intake and Incidence of Type 2 Diabetes in Adults: A Dose Response Meta-Analysis of Cohort Studies. Diabetol. Metab. Syndr. 2022, 14, 34. [Google Scholar] [CrossRef] [PubMed]
- Sigaux, J.; Mathieu, S.; Nguyen, Y.; Sanchez, P.; Letarouilly, J.G.; Soubrier, M.; Czernichow, S.; Flipo, R.M.; Sellam, J.; Daïen, C. Impact of Type and Dose of Oral Polyunsaturated Fatty Acid Supplementation on Disease Activity in Inflammatory Rheumatic Diseases: A Systematic Literature Review and Meta-Analysis. Arthritis Res. Ther. 2022, 24, 100. [Google Scholar] [CrossRef] [PubMed]
- Wood, A.H.R.; Chappell, H.F.; Zulyniak, M.A. Dietary and Supplemental Long-Chain Omega-3 Fatty Acids as Moderators of Cognitive Impairment and Alzheimer’s Disease. Eur. J. Nutr. 2022, 61, 589–604. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Calder, P.C. Dietary α-Linolenic Acid and Health-Related Outcomes: A Metabolic Perspective. Nutr. Res. Rev. 2006, 19, 26–52. [Google Scholar] [CrossRef] [PubMed]
- Agostoni, C.; Bresson, J.-L.; Fairweather-Tait, S.; Flynn, A.; Golly, I.; Korhonen, H.; Lagiou, P.; Løvik, M.; Marchelli, R.; Martin, A.; et al. Scientific Opinion on the Tolerable Upper Intake Level of Eicosapentaenoic Acid (EPA), Docosahexaenoic Acid (DHA) and Docosapentaenoic Acid (DPA). EFSA J. 2012, 10, 2815. [Google Scholar] [CrossRef]
- Neves, M.A.; Wang, Z.; Kobayashi, I.; Nakajima, M. Assessment of Oxidative Stability in Fish Oil-in-Water Emulsions: Effect of Emulsification Process, Droplet Size and Storage Temperature. J. Food Process Eng. 2017, 40, e12316. [Google Scholar] [CrossRef]
- Jamshidi, A.; Cao, H.; Xiao, J.; Simal-Gandara, J. Advantages of Techniques to Fortify Food Products with the Benefits of Fish Oil. Food Res. Int. 2020, 137, 109353. [Google Scholar] [CrossRef]
- Ye, A.; Cui, J.; Taneja, A.; Zhu, X.; Singh, H. Evaluation of Processed Cheese Fortified with Fish Oil Emulsion. Food Res. Int. 2009, 42, 1093–1098. [Google Scholar] [CrossRef]
- Pourashouri, P.; Shabanpour, B.; Kordjazi, M.; Jamshidi, A. Characteristic and Shelf Life of Fish Sausage: Fortification with Fish Oil through Emulsion and Gelled Emulsion Incorporated with Green Tea Extract. J. Sci. Food Agric. 2020, 100, 4474–4482. [Google Scholar] [CrossRef]
- Gumus, C.E.; Gharibzahedi, S.M.T. Yogurts Supplemented with Lipid Emulsions Rich in Omega-3 Fatty Acids: New Insights into the Fortification, Microencapsulation, Quality Properties, and Health-Promoting Effects. Trends Food Sci. Technol. 2021, 110, 267–279. [Google Scholar] [CrossRef]
- Lourenço, S.C.; Moldão-Martins, M.; Alves, V.D. Antioxidants of Natural Plant Origins: From Sources to Food Industry Applications. Molecules 2019, 24, 4132. [Google Scholar] [CrossRef] [PubMed]
- Wong, F.-C.; Xiao, J.; Wang, S.; Ee, K.-Y.; Chai, T.-T. Advances on the Antioxidant Peptides from Edible Plant Sources. Trends Food Sci. Technol. 2020, 99, 44–57. [Google Scholar] [CrossRef]
- Görgüç, A.; Gençdağ, E.; Yılmaz, F.M. Bioactive Peptides Derived from Plant Origin by-Products: Biological Activities and Techno-Functional Utilizations in Food Developments—A Review. Food Res. Int. 2020, 136, 109504. [Google Scholar] [CrossRef] [PubMed]
- Aondona, M.M.; Ikya, J.K.; Ukeyima, M.T.; Gborigo, T.-W.J.A.; Aluko, R.E.; Girgih, A.T. In Vitro Antioxidant and Antihypertensive Properties of Sesame Seed Enzymatic Protein Hydrolysate and Ultrafiltration Peptide Fractions. J. Food Biochem. 2021, 45, e13587. [Google Scholar] [CrossRef]
- Ozón, B.; Cotabarren, J.; Valicenti, T.; Parisi, M.G.; Obregón, W.D. Chia Expeller: A Promising Source of Antioxidant, Antihypertensive and Antithrombotic Peptides Produced by Enzymatic Hydrolysis with Alcalase and Flavourzyme. Food Chem. 2022, 380, 132185. [Google Scholar] [CrossRef]
- Ohara, A.; Cason, V.G.; Nishide, T.G.; Miranda de Matos, F.; de Castro, R.J.S. Improving the Antioxidant and Antidiabetic Properties of Common Bean Proteins by Enzymatic Hydrolysis Using a Blend of Proteases. Biocatal. Biotransform. 2021, 39, 100–108. [Google Scholar] [CrossRef]
- Aydemir, L.Y.; Diblan, S.; Aktas, H.; Cakitli, G. Changes in Bioactive Properties of Dry Bean Extracts during Enzymatic Hydrolysis and in Vitro Digestion Steps. J. Food Meas. Charact. 2022, 2022, 1–17. [Google Scholar] [CrossRef]
- Suarez, L.M.; Fan, H.; Zapata, J.E.; Wu, J. Optimization of Enzymatic Hydrolysis for Preparing Cassava Leaf Hydrolysate with Antioxidant Activity. Food Bioprocess Technol. 2021, 14, 2181–2194. [Google Scholar] [CrossRef]
- Famuwagun, A.A.; Alashi, A.M.; Gbadamosi, O.S.; Taiwo, K.A.; Oyedele, D.; Adebooye, O.C.; Aluko, R.E. Antioxidant and Enzymes Inhibitory Properties of Amaranth Leaf Protein Hydrolyzates and Ultrafiltration Peptide Fractions. J. Food Biochem. 2021, 45, e13396. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Enzymic Hydrolysis of Food Proteins, 1st ed.; Elsevier Applied Science Publishers LTD: London, UK, 1986. [Google Scholar]
- Picot, L.; Ravallec, R.; Fouchereau-Péron, M.; Vandanjon, L.; Jaouen, P.; Chaplain-Derouiniot, M.; Guérard, F.; Chabeaud, A.; Legal, Y.; Alvarez, O.M.; et al. Impact of Ultrafiltration and Nanofiltration of an Industrial Fish Protein Hydrolysate on Its Bioactive Properties. J. Sci. Food Agric. 2010, 90, 1819–1826. [Google Scholar] [CrossRef] [PubMed]
- Decker, E.A.; Welch, B. Role of Ferritin as a Lipid Oxidation Catalyst in Muscle Food. J. Agric. Food Chem. 1990, 38, 674–677. [Google Scholar] [CrossRef]
- Padial-Domínguez, M.; Espejo-Carpio, F.J.; García-Moreno, P.J.; Jacobsen, C.; Guadix, E.M. Protein Derived Emulsifiers with Antioxidant Activity for Stabilization of Omega-3 Emulsions. Food Chem. 2020, 329, 127148. [Google Scholar] [CrossRef]
- Padial-Domínguez, M.; Espejo-Carpio, F.J.; Pérez-Gálvez, R.; Guadix, A.; Guadix, E.M. Optimization of the Emulsifying Properties of Food Protein Hydrolysates for the Production of Fish Oil-in-Water Emulsions. Foods 2020, 9, 636. [Google Scholar] [CrossRef]
- Petursson, S.; Decker, E.A.; McClements, D.J. Stabilization of Oil-in-Water Emulsions by Cod Protein Extracts. J. Agric. Food Chem. 2004, 52, 3996–4001. [Google Scholar] [CrossRef] [PubMed]
- Shantha, N.C.; Decker, E.A. Rapid, Sensitive, Iron-Based Spectrophotometric Methods for Determination of Peroxide Values of Food Lipids. J. AOAC Int. 1994, 77, 421–424. [Google Scholar] [CrossRef]
- ISO 6885:2006; Animal and Vegetable Fats and Oils—Determination of Anisidine Value. ISO: Geneva, Switzerland, 2006. Available online: https://www.iso.org/standard/40052.html (accessed on 17 June 2022).
- Zafeiriou, I.; Polidoros, A.N.; Baira, E.; Kasiotis, K.M.; Machera, K.; Mylona, P.V. Mediterranean White Lupin Landraces as a Valuable Genetic Reserve for Breeding. Plants 2021, 10, 2403. [Google Scholar] [CrossRef]
- Garmidolova, A.; Desseva, I.; Mihaylova, D.; Lante, A. Bioactive Peptides from Lupinus spp. Seed Proteins-State-of-the-Art and Perspectives. Appl. Sci. 2022, 12, 3766. [Google Scholar] [CrossRef]
- Kotecka-Majchrzak, K.; Sumara, A.; Fornal, E.; Montowska, M. Oilseed Proteins—Properties and Application as a Food Ingredient. Trends Food Sci. Technol. 2020, 106, 160–170. [Google Scholar] [CrossRef]
- Okagu, I.U.; Ndefo, J.C.; Aham, E.C.; Obeme-Nmom, J.I.; Agboinghale, P.E.; Aguchem, R.N.; Nechi, R.N.; Lammi, C. Lupin-Derived Bioactive Peptides: Intestinal Transport, Bioavailability and Health Benefits. Nutrients 2021, 13, 3266. [Google Scholar] [CrossRef]
- Maestri, D.; Barrionuevo, D.; Bodoira, R.; Zafra, A.; Jiménez-López, J.; Alché, J.d.D. Nutritional Profile and Nutraceutical Components of Olive (Olea europaea L.) Seeds. J. Food Sci. Technol. 2019, 56, 4359–4370. [Google Scholar] [CrossRef]
- Khanpit, V.V.; Tajane, S.P.; Mandavgane, S.A. Dietary Fibers from Fruit and Vegetable Waste: Methods of Extraction and Processes of Value Addition. Biomass Convers. Biorefinery 2021. [Google Scholar] [CrossRef]
- Sergio, J.-C.; Serna-Saldívar, O.; Campanella, O.; Editors, V.-O. Science and Technology of Fibers in Food Systems; Springer Nature Switzerland AG 2020: Cham, Switzerland, 2020. [Google Scholar]
- Rodrigues, I.M.; Coelho, J.F.J.; Carvalho, M.G.V.S. Isolation and Valorisation of Vegetable Proteins from Oilseed Plants: Methods, Limitations and Potential. J. Food Eng. 2012, 109, 337–346. [Google Scholar] [CrossRef]
- Ghribi, A.M.; Gafsi, I.M.; Sila, A.; Blecker, C.; Danthine, S.; Attia, H.; Bougatef, A.; Besbes, S. Effects of Enzymatic Hydrolysis on Conformational and Functional Properties of Chickpea Protein Isolate. Food Chem. 2015, 187, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.V.; Ong, S.-E.; Mann, M. Trypsin Cleaves Exclusively C-Terminal to Arginine and Lysine Residues. Mol. Cell. Proteom. 2004, 3, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Berraquero-García, C.; Almécija, M.C.; Guadix, E.M.; Pérez-Gálvez, R. Valorisation of Blood Protein from Livestock to Produce Haem Iron-Fortified Hydrolysates with Antioxidant Activity. Int. J. Food Sci. Technol. 2022, 57, 2479–2486. [Google Scholar] [CrossRef]
- Vioque, J.; Sánchez-Vioque, R.; Clemente, A.; Pedroche, J.; Millán, F. Partially Hydrolyzed Rapeseed Protein Isolates with Improved Functional Properties. J. Am. Oil Chem. Soc. 2000, 77, 447–450. [Google Scholar] [CrossRef]
- Morales-Medina, R.; Tamm, F.; Guadix, A.M.; Guadix, E.M.; Drusch, S. Functional and Antioxidant Properties of Hydrolysates of Sardine (S. pilchardus) and Horse Mackerel (T. mediterraneus) for the Microencapsulation of Fish Oil by Spray-Drying. Food Chem. 2016, 194, 1208–1216. [Google Scholar] [CrossRef]
- Tacias-Pascacio, V.G.; Morellon-Sterling, R.; Siar, E.H.; Tavano, O.; Berenguer-Murcia, Á.; Fernandez-Lafuente, R. Use of Alcalase in the Production of Bioactive Peptides: A Review. Int. J. Biol. Macromol. 2020, 165, 2143–2196. [Google Scholar] [CrossRef]
- Ying, X.; Agyei, D.; Udenigwe, C.; Adhikari, B.; Wang, B. Manufacturing of Plant-Based Bioactive Peptides Using Enzymatic Methods to Meet Health and Sustainability Targets of the Sustainable Development Goals. Front. Sustain. Food Syst. 2021, 5, 769028. [Google Scholar] [CrossRef]
- Manzoor, M.; Singh, J.; Gani, A. Exploration of Bioactive Peptides from Various Origin as Promising Nutraceutical Treasures: In Vitro, in Silico and in Vivo Studies. Food Chem. 2022, 373, 131395. [Google Scholar] [CrossRef] [PubMed]
- López-García, G.; Dublan-García, O.; Arizmendi-Cotero, D.; Oliván, L.M.G. Antioxidant and Antimicrobial Peptides Derived from Food Proteins. Molecules 2022, 27, 1343. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Xiong, Y.L.; Kong, B. Antioxidant Activity of Peptide Fractions from Whey Protein Hydrolysates as Measured by Electron Spin Resonance. Food Chem. 2009, 113, 196–201. [Google Scholar] [CrossRef]
- Rival, S.G.; Boeriu, C.G.; Wichers, H.J. Caseins and Casein Hydrolysates. 2. Antioxidative Properties and Relevance to Lipoxygenase Inhibition. J. Agric. Food Chem. 2001, 49, 295–302. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, H.; Han, B.; Li, H.Y.; Liu, X.L. Improvement of Nutritional Value, Molecular Weight Patterns (Soluble Peptides), Free Amino Acid Patterns, Total Phenolics and Antioxidant Activity of Fermented Extrusion Pretreatment Rapeseed Meal with Bacillus subtilis YY-1 and Saccharomyces cerevisiae YY-2. LWT 2022, 160, 113280. [Google Scholar] [CrossRef]
- Bougatef, A.; Hajji, M.; Balti, R.; Lassoued, I.; Triki-Ellouz, Y.; Nasri, M. Antioxidant and Free Radical-Scavenging Activities of Smooth Hound (Mustelus mustelus) Muscle Protein Hydrolysates Obtained by Gastrointestinal Proteases. Food Chem. 2009, 114, 1198–1205. [Google Scholar] [CrossRef]
- Fathi, M.; Hosseini, F.S.; Reyhane, R.; Rashidi, L. Optimized Enzymatic Hydrolysis of Olive Pomace Proteins Using Response Surface Methodology. Appl. Food Biotechnol. 2022, 9, 79–90. [Google Scholar] [CrossRef]
- Ibagon, J.A.; Lee, S.A.; Stein, H.H. Sunflower Expellers Have Greater Ileal Digestibility of Amino Acids than Sunflower Meal, but There Are Only Minor Variations among Different Sources of Sunflower Meal When Fed to Growing Pigs. J. Anim. Sci. 2021, 99, skab198. [Google Scholar] [CrossRef]
- Vioque, J.; Sánchez-Vioque, R.; Clemente, A.; Pedroche, J.; Bautista, J.; Millan, F. Production and Characterization of an Extensive Rapeseed Protein Hydrolysate. JAOCS J. Am. Oil Chem. Soc. 1999, 76, 819–823. [Google Scholar] [CrossRef]
- Mierlita, D.; Simeanu, D.; Pop, I.M.; Criste, F.; Pop, C.; Simeanu, C.; Lup, F. Chemical Composition and Nutritional Evaluation of the Lupine Seeds (Lupinus albus L.) from Low-Alkaloid Varieties. Rev. Chim. 2018, 69, 453–458. [Google Scholar] [CrossRef]
- Vogelsang-O’Dwyer, M.; Bez, J.; Petersen, I.L.; Joehnke, M.S.; Detzel, A.; Busch, M.; Krueger, M.; Ispiryan, L.; O’Mahony, J.A.; Arendt, E.K.; et al. Techno-Functional, Nutritional and Environmental Performance of Protein Isolates from Blue Lupin and White Lupin. Foods 2020, 9, 230. [Google Scholar] [CrossRef] [PubMed]
- Farvin, K.H.S.; Andersen, L.L.; Otte, J.; Nielsen, H.H.; Jessen, F.; Jacobsen, C. Antioxidant Activity of Cod (Gadus morhua) Protein Hydrolysates: Fractionation and Characterisation of Peptide Fractions. Food Chem. 2016, 204, 409–419. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Girgih, A.T.; Malomo, S.A.; Ju, X.; Aluko, R.E. Antioxidant Activities of Enzymatic Rapeseed Protein Hydrolysates and the Membrane Ultrafiltration Fractions. J. Funct. Foods 2013, 5, 219–227. [Google Scholar] [CrossRef]
- González-Hidalgo, I.; Bañón, S.; Ros, J.M. Evaluation of Table Olive by-Product as a Source of Natural Antioxidants. Int. J. Food Sci. Technol. 2012, 47, 674–681. [Google Scholar] [CrossRef]
- Farvin, K.H.S.; Andersen, L.L.; Nielsen, H.H.; Jacobsen, C.; Jakobsen, G.; Johansson, I.; Jessen, F. Antioxidant Activity of Cod (Gadus morhua) Protein Hydrolysates: In Vitro Assays and Evaluation in 5% Fish Oil-in-Water Emulsion. Food Chem. 2014, 149, 326–334. [Google Scholar] [CrossRef]
- Zhang, T.; Li, Y.; Miao, M.; Jiang, B. Purification and Characterisation of a New Antioxidant Peptide from Chickpea (Cicer arietium L.) Protein Hydrolysates. Food Chem. 2011, 128, 28–33. [Google Scholar] [CrossRef]
- Carrasco-Castilla, J.; Hernández-Álvarez, A.J.; Jiménez-Martínez, C.; Jacinto-Hernández, C.; Alaiz, M.; Girón-Calle, J.; Vioque, J.; Dávila-Ortiz, G. Antioxidant and Metal Chelating Activities of Peptide Fractions from Phaseolin and Bean Protein Hydrolysates. Food Chem. 2012, 135, 1789–1795. [Google Scholar] [CrossRef]
- Yesiltas, B.; García-Moreno, P.J.; Gregersen, S.; Olsen, T.H.; Jones, N.C.; Hoffmann, S.V.; Marcatili, P.; Overgaard, M.T.; Hansen, E.B.; Jacobsen, C. Antioxidant Peptides Derived from Potato, Seaweed, Microbial and Spinach Proteins: Oxidative Stability of 5% Fish Oil-in-Water Emulsions. Food Chem. 2022, 385, 132699. [Google Scholar] [CrossRef]
- Esmaeilzadeh Kenari, R.; Razavi, R. Phenolic Profile and Antioxidant Activity of Free/Bound Phenolic Compounds of Sesame and Properties of Encapsulated Nanoparticles in Different Wall Materials. Food Sci. Nutr. 2022, 10, 525–535. [Google Scholar] [CrossRef]
- Cheng, Y.; Xiong, Y.L.; Chen, J. Antioxidant and Emulsifying Properties of Potato Protein Hydrolysate in Soybean Oil-in-Water Emulsions. Food Chem. 2010, 120, 101–108. [Google Scholar] [CrossRef]
- García-Moreno, P.J.; Yang, J.; Gregersen, S.; Jones, N.C.; Berton-Carabin, C.C.; Sagis, L.M.C.; Hoffmann, S.V.; Marcatili, P.; Overgaard, M.T.; Hansen, E.B.; et al. The Structure, Viscoelasticity and Charge of Potato Peptides Adsorbed at the Oil-Water Interface Determine the Physicochemical Stability of Fish Oil-in-Water Emulsions. Food Hydrocoll. 2021, 115, 106605. [Google Scholar] [CrossRef]
- Wang, B.; Li, L.; Chi, C.F.; Ma, J.H.; Luo, H.Y.; Xu, Y.F. Purification and Characterisation of a Novel Antioxidant Peptide Derived from Blue Mussel (Mytilus edulis) Protein Hydrolysate. Food Chem. 2013, 138, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Ka, H.; Yi, B.; Kim, M.J.; Lee, J. Evaluation of Antioxidant or Prooxidant Properties of Selected Amino Acids Using In Vitro Assays and in Oil-in-Water Emulsions Under Riboflavin Sensitization. J. Food Sci. 2016, 81, C1118–C1123. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Liu, M.; Zhao, H.; Lv, Z.; Liang, L.; Zhang, B. A Novel Insight into Screening for Antioxidant Peptides from Hazelnut Protein: Based on the Properties of Amino Acid Residues. Antioxidants 2022, 11, 127. [Google Scholar] [CrossRef] [PubMed]
- Petraru, A.; Ursachi, F.; Amariei, S. Nutritional Characteristics Assessment of Sunflower Seeds, Oil and Cake. Perspective of Using Sunflower Oilcakes as a Functional Ingredient. Plants 2021, 10, 2487. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Protein Hydrolysate | LUM–H | OSM–H | SFSM–H | RSM–H |
|---|---|---|---|---|
| Plant meal solubilization (%) | 57.9 ± 2.5 b | 60.4 ± 1.6 b | 67.0 ± 2.8 b | 75.5 ± 1.4 a |
| Protein recovery (%) | 56.8 ± 7.0 b | 56.6 ± 6.6 b | 82.2 ± 11.2 a | 79.6 ± 8.9 a |
| Protein content (%wt) | 45.1 ± 3.1 a | 25.0 ± 1.6 b | 44.7 ± 3.5 a | 50.6 ± 3.6 a |
| Crude fat (%wt) | 5.9 ± 0.5 b | 8.2 ± 0.8 a | 1.7 ± 0.2 b | 2.1 ± 0.2 b |
| Moisture (%wt) | 6.9 ± 0.5 c | 9.8 ± 0.7 a | 9.3 ± 0.6 ab | 8.0 ± 0.6 bc |
| Emulsion | ζ-Potential (mV) | Surface Mean Diameter D[3,2] (µm) | Volume Mean Diameter D[4,3] (µm) |
|---|---|---|---|
| LUM–H | −48.5 ± 2.70 c | 0.316 ± 0.002 a | 0.417 ± 0.001 ab |
| OSM–H | −52.3 ± 2.48 c | 0.315 ± 0.005 a | 0.393 ± 0.009 b |
| SFSM–H | −42.7 ± 0.58 b | 0.326 ± 0.009 a | 0.425 ± 0.021 a |
| RSM–H | −42.4 ± 0.88 b | 0.329 ± 0.009 a | 0.418 ± 0.007 ab |
| WPC–H | −40.1 ± 2.26 ab | 0.312 ± 0.007 a | 0.399 ± 0.005 ab |
| NC | −36.1 ± 1.12 a | 0.324 ± 0.006 a | 0.417 ± 0.005 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ospina-Quiroga, J.L.; García-Moreno, P.J.; Guadix, A.; Guadix, E.M.; Almécija-Rodríguez, M.d.C.; Pérez-Gálvez, R. Evaluation of Plant Protein Hydrolysates as Natural Antioxidants in Fish Oil-In-Water Emulsions. Antioxidants 2022, 11, 1612. https://doi.org/10.3390/antiox11081612
Ospina-Quiroga JL, García-Moreno PJ, Guadix A, Guadix EM, Almécija-Rodríguez MdC, Pérez-Gálvez R. Evaluation of Plant Protein Hydrolysates as Natural Antioxidants in Fish Oil-In-Water Emulsions. Antioxidants. 2022; 11(8):1612. https://doi.org/10.3390/antiox11081612
Chicago/Turabian StyleOspina-Quiroga, Jeimmy Lizeth, Pedro J. García-Moreno, Antonio Guadix, Emilia M. Guadix, María del Carmen Almécija-Rodríguez, and Raúl Pérez-Gálvez. 2022. "Evaluation of Plant Protein Hydrolysates as Natural Antioxidants in Fish Oil-In-Water Emulsions" Antioxidants 11, no. 8: 1612. https://doi.org/10.3390/antiox11081612