
Dietary Intervention with Blackcurrant Pomace Protects Rats from Testicular Oxidative Stress Induced by Exposition to Biodiesel Exhaust

 , , , and
, , , and 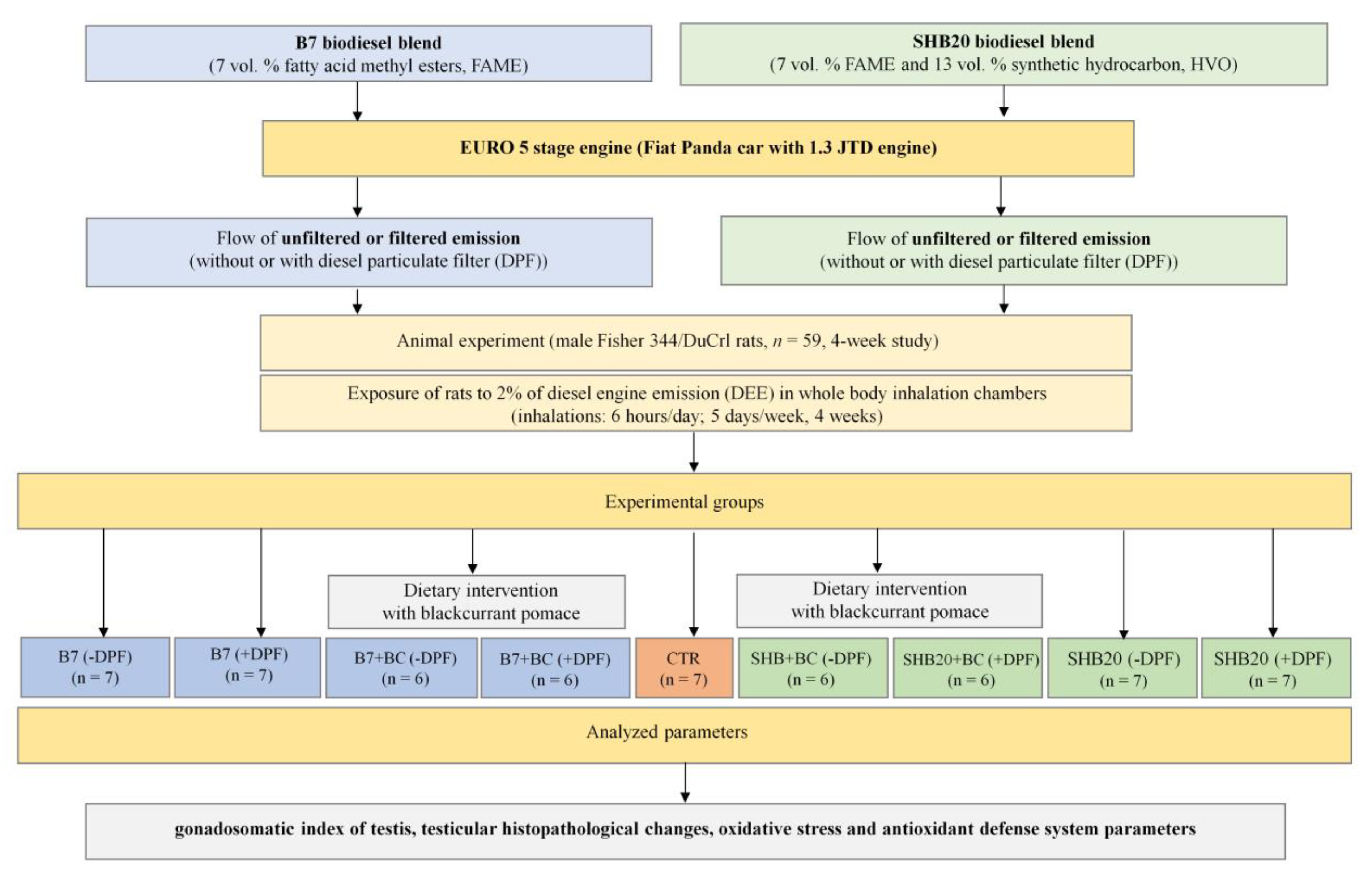{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Animal Study
2.3. Exposure of Animals to DEE
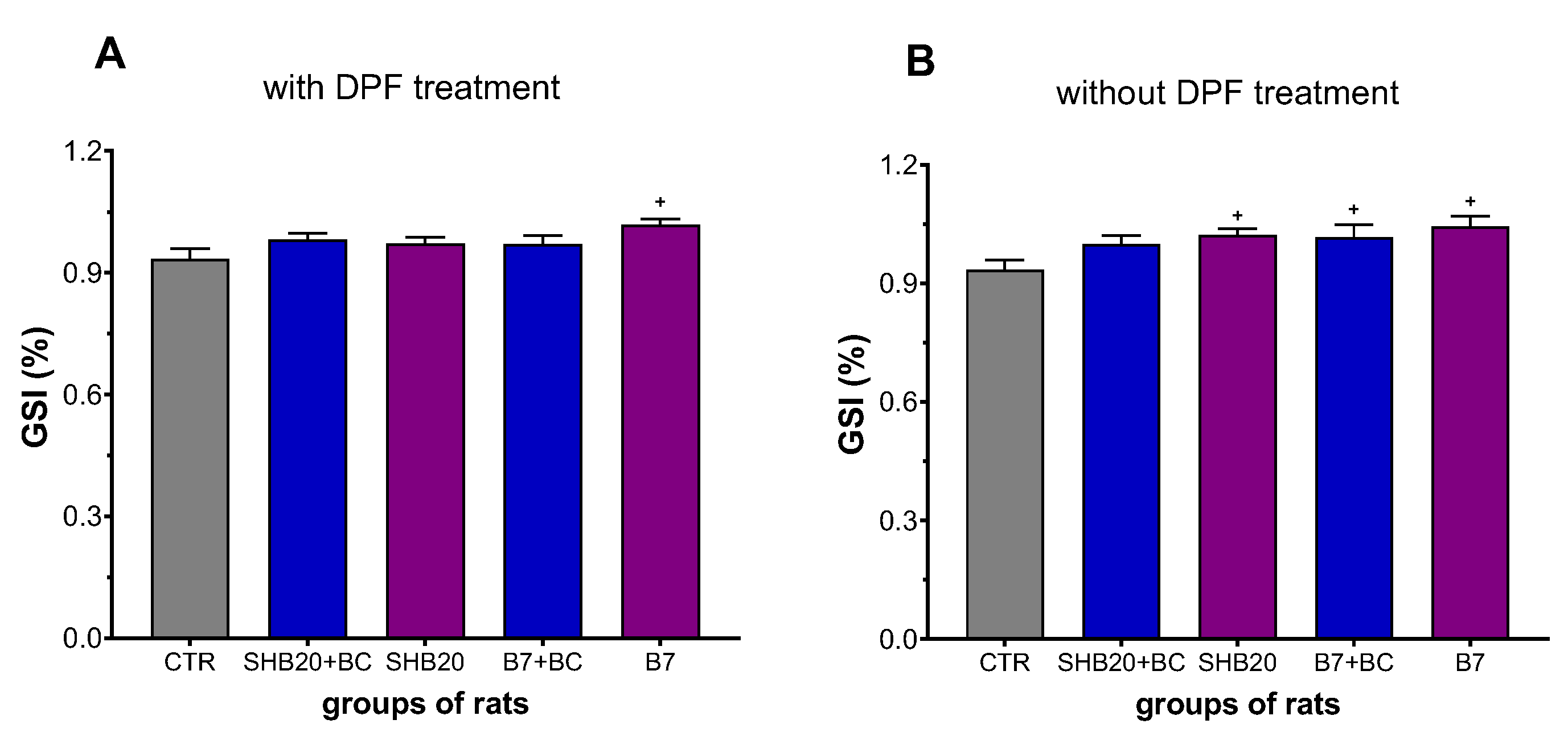
2.4. Gonadosomatic Index
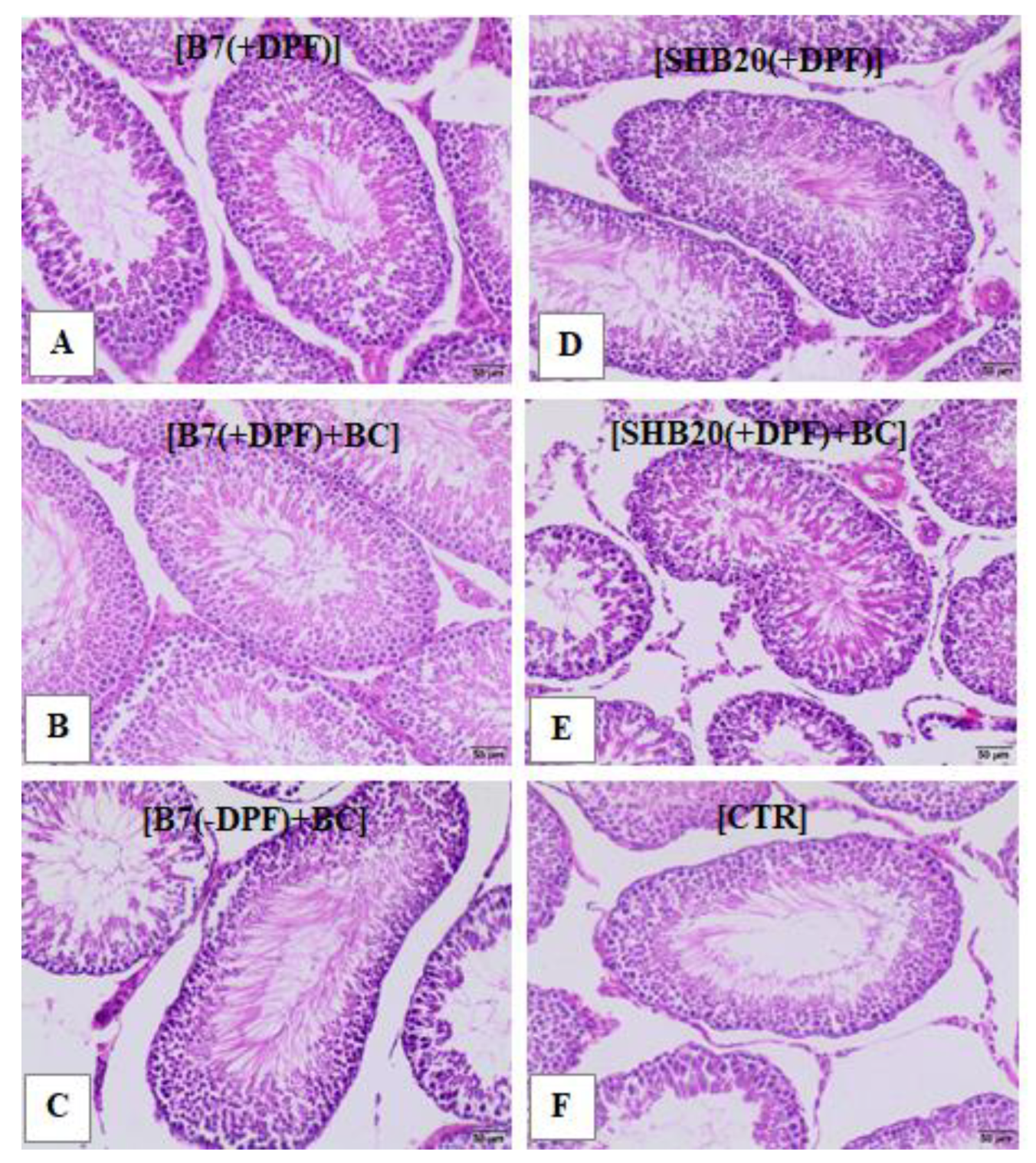
2.5. Histological Assessment of Testis
2.6. Oxidative Stress Parameters in Testis
2.7. Anti-Oxidative Defense System Parameters in Testis
2.8. Statistical Analysis
3. Results
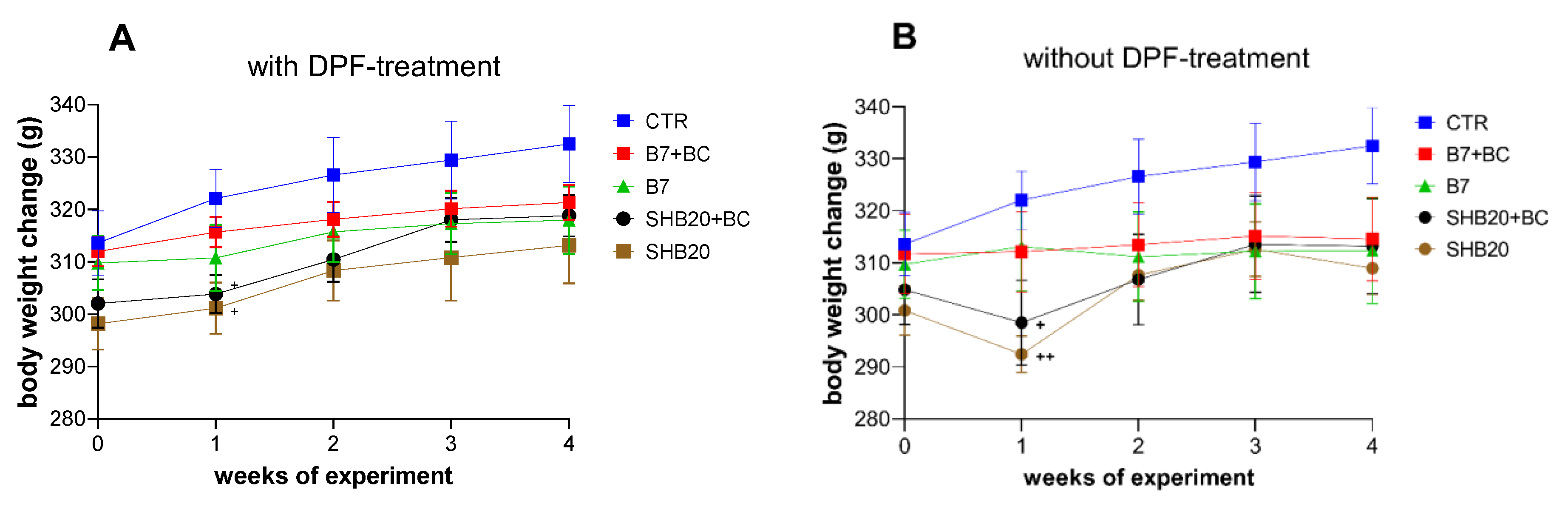
3.1. Animal Observation, Weight Changes, Gonadosomatic Index, and Microscopic Evaluation of Rat Testis
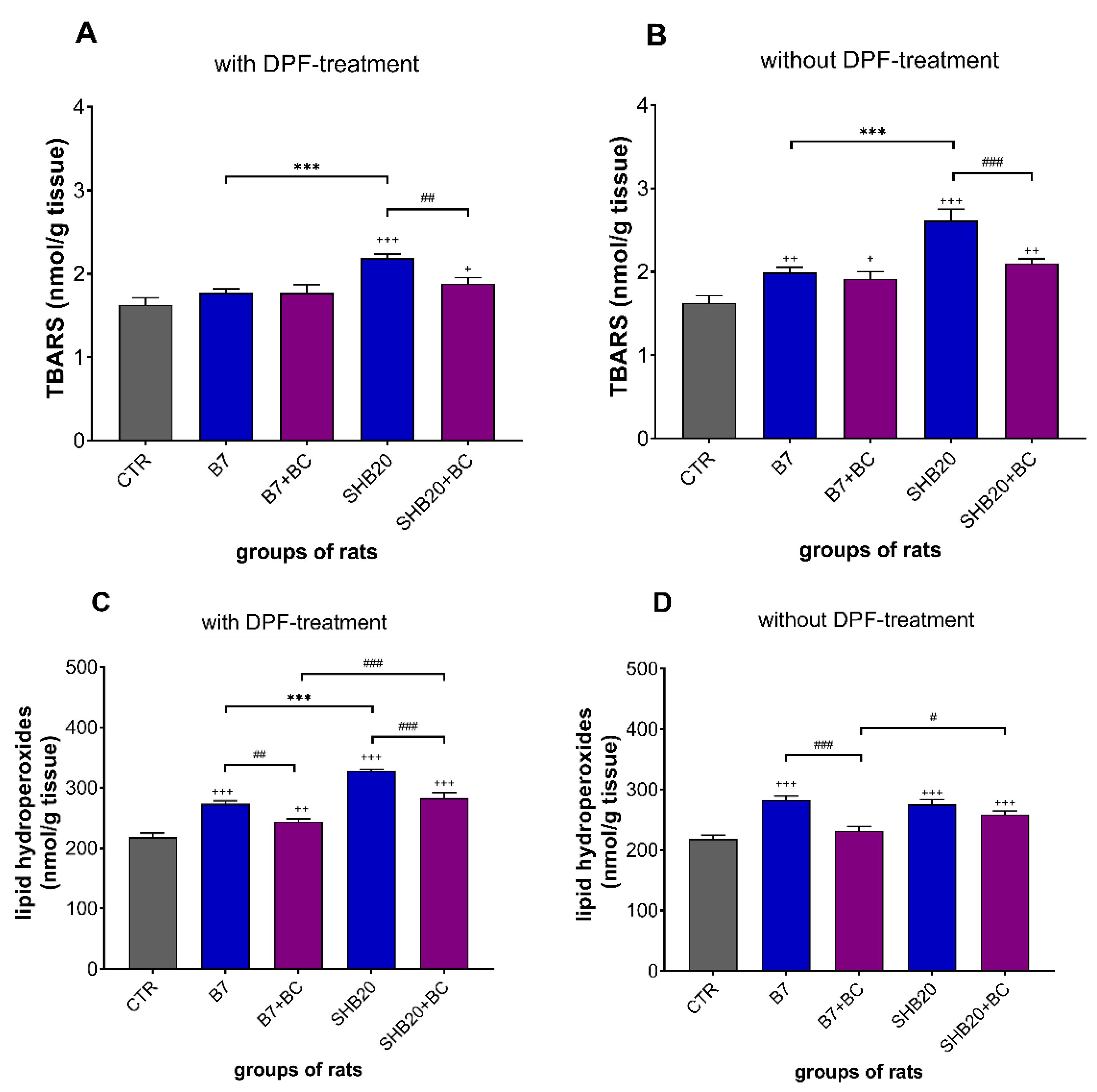
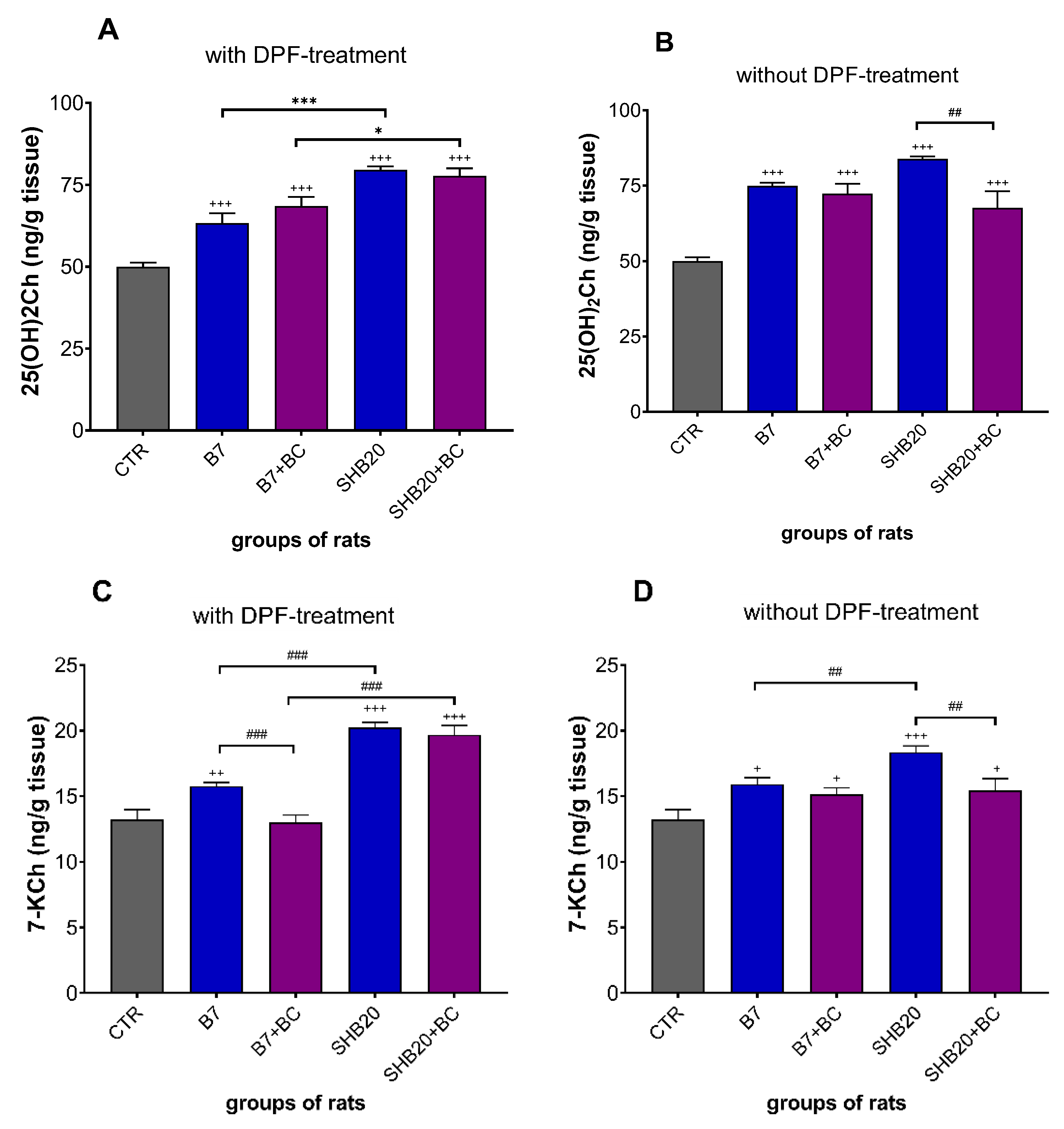
3.2. Oxidative Stress Markers
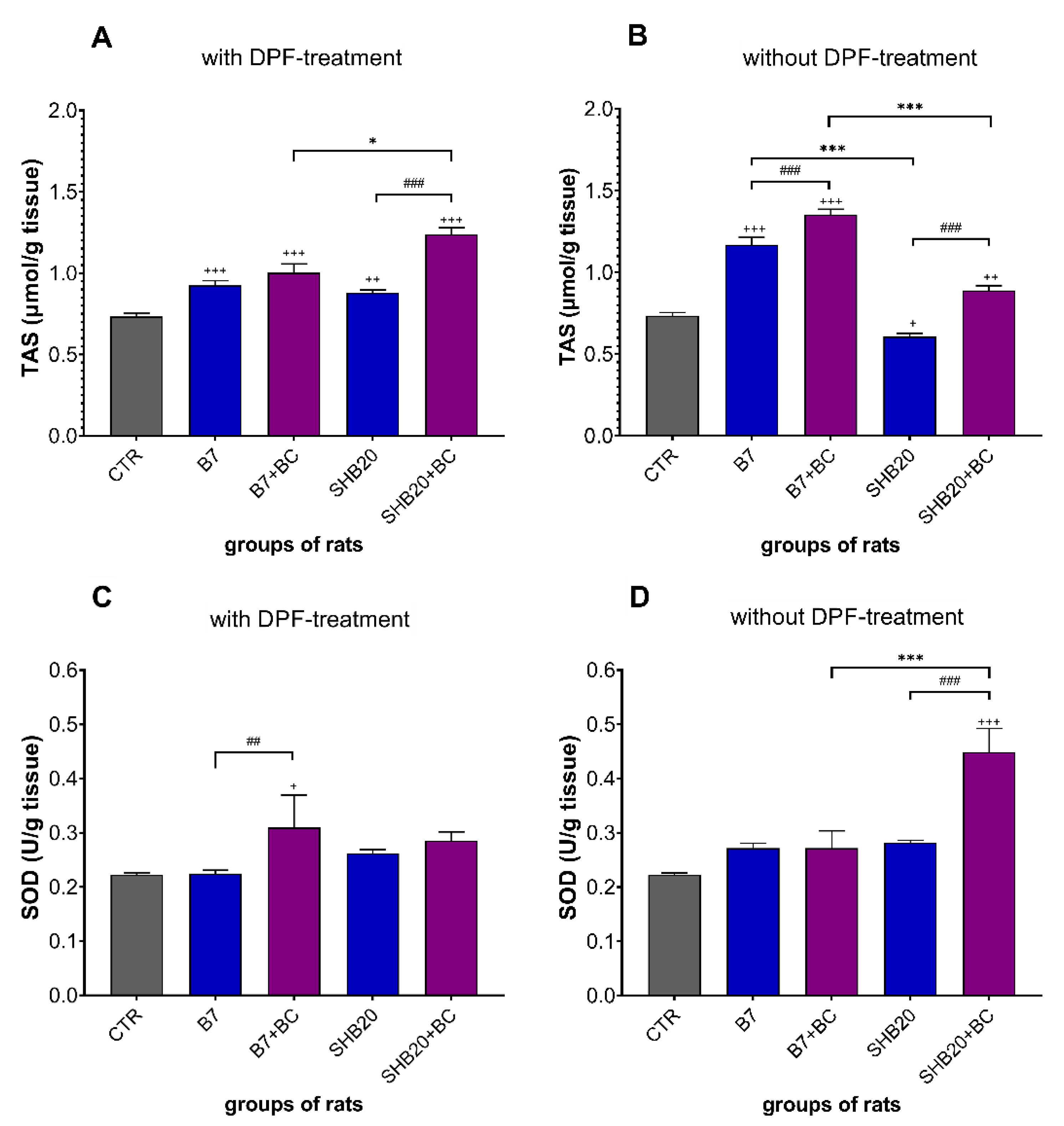
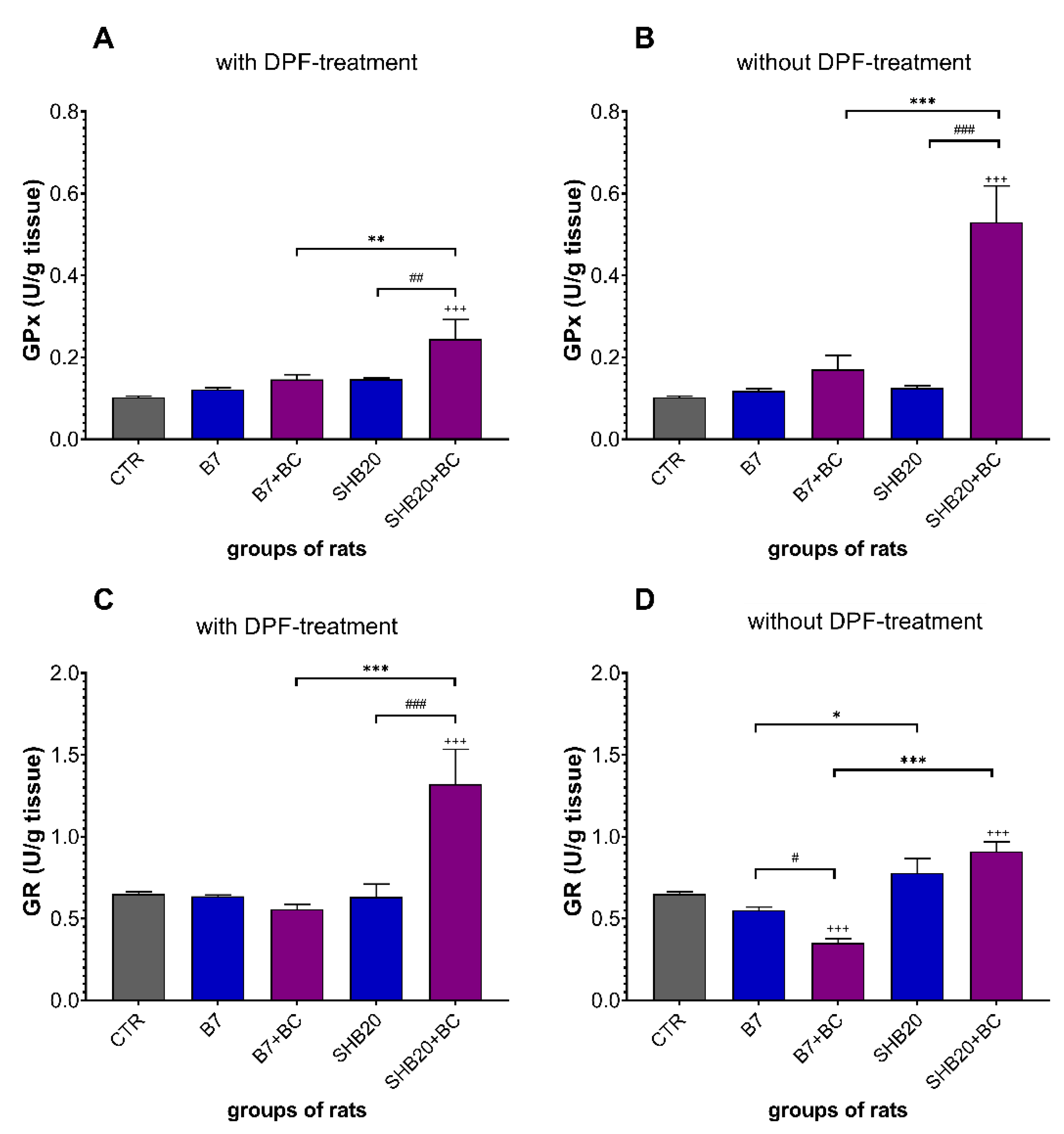
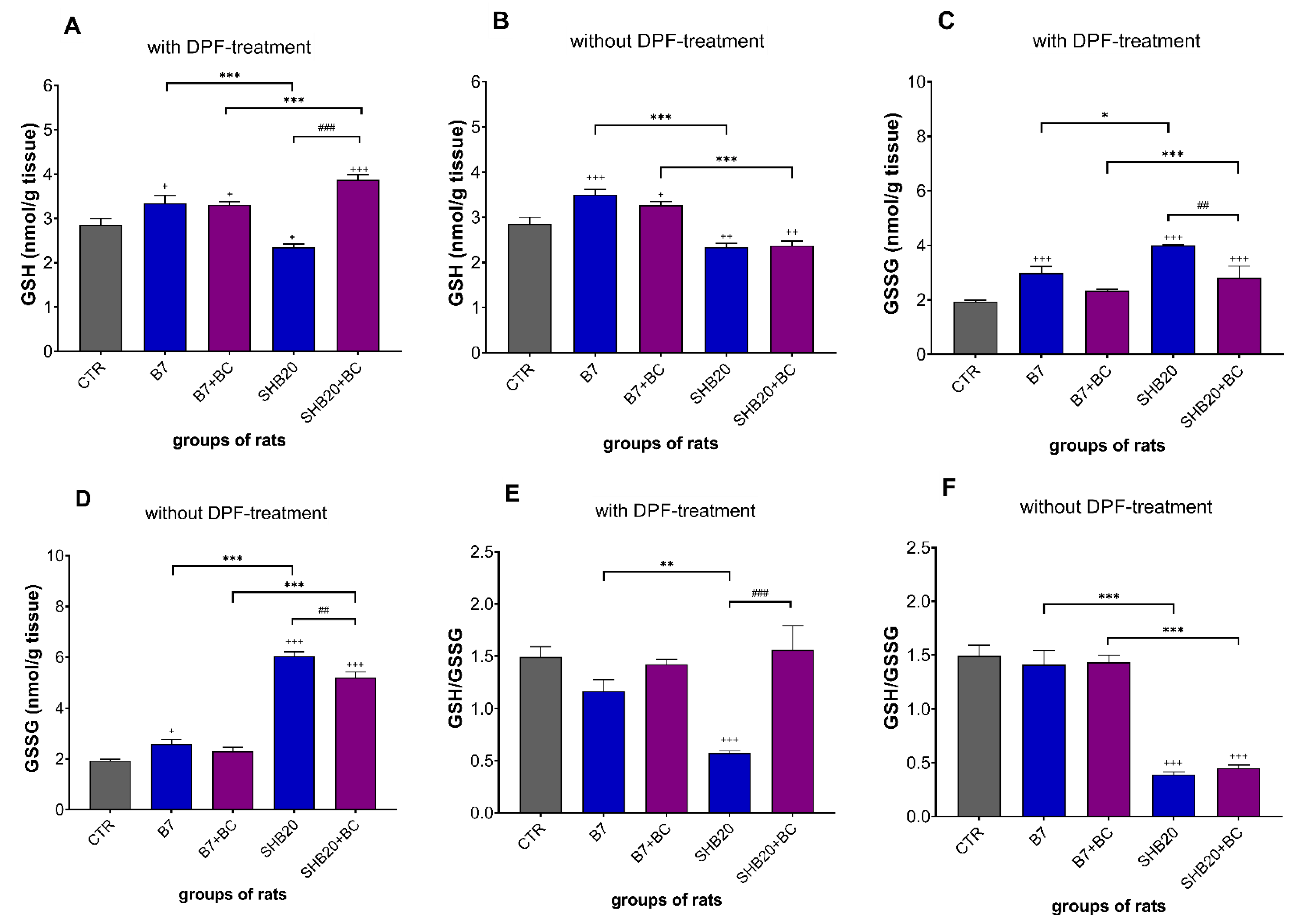
3.3. Antioxidant Defense Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lelieveld, J.; Evans, J.S.; Fnais, M.; Giannadaki, D.; Pozzer, A. The Contribution of Outdoor Air Pollution Sources to Premature Mortality on a Global Scale. Nature 2015, 525, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.J.; Brauer, M.; Burnett, R.; Anderson, H.R.; Frostad, J.; Estep, K.; Balakrishnan, K.; Brunekreef, B.; Dandona, L.; Dandona, R.; et al. Estimates and 25-Year Trends of the Global Burden of Disease Attributable to Ambient Air Pollution: An Analysis of Data from the Global Burden of Diseases Study 2015. Lancet 2017, 389, 1907–1918. [Google Scholar] [CrossRef]
- Weitekamp, C.A.; Kerr, L.B.; Dishaw, L.; Nichols, J.; Lein, M.; Stewart, M.J. A Systematic Review of the Health Effects Associated with the Inhalation of Particle-Filtered and Whole Diesel Exhaust. Inhal. Toxicol. 2020, 32, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Verma, M.K.; Srivastava, A.K. Ultrafine Particles in Urban Ambient Air and Their Health Perspectives. Rev. Environ. Health 2013, 28, 117–128. [Google Scholar] [CrossRef]
- Gren, L.; Dierschke, K.; Mattsson, F.; Assarsson, E.; Krais, A.M.; Kåredal, M.; Lovén, K.; Löndahl, J.; Pagels, J.; Strandberg, B.; et al. Lung Function and Self-Rated Symptoms in Healthy Volunteers after Exposure to Hydrotreated Vegetable Oil (HVO) Exhaust with and without Particles. Part. Fibre Toxicol. 2022, 19, 9. [Google Scholar] [CrossRef]
- Burki, T.K. Diesel Emissions from Cars Much Worse than Trucks and Buses. Lancet Respir. Med. 2017, 5, 98. [Google Scholar] [CrossRef]
- Ayodhya, A.S.; Narayanappa, K.G. An Overview of After-Treatment Systems for Diesel Engines. Environ. Sci. Pollut. Res. 2018, 25, 35034–35047. [Google Scholar] [CrossRef]
- O’Driscoll, C.A.; Owens, L.A.; Gallo, M.E.; Hoffmann, E.J.; Afrazi, A.; Han, M.; Fechner, J.H.; Schauer, J.J.; Bradfield, C.A.; Mezrich, J.D. Differential Effects of Diesel Exhaust Particles on T Cell Differentiation and Autoimmune Disease. Part. Fibre Toxicol. 2018, 15, 35. [Google Scholar] [CrossRef]
- Bengalli, R.; Zerboni, A.; Marchetti, S.; Longhin, E.; Priola, M.; Camatini, M.; Mantecca, P. In Vitro Pulmonary and Vascular Effects Induced by Different Diesel Exhaust Particles. Toxicol. Lett. 2019, 306, 13–24. [Google Scholar] [CrossRef]
- Risom, L.; Dybdahl, M.; Bornholdt, J.; Vogel, U.; Wallin, H.; Møller, P.; Loft, S. Oxidative DNA Damage and Defense Gene Expression in the Mouse Lung after Short-Term Exposure to Diesel Exhaust Particles by Inhalation. Carcinogenesis 2003, 24, 1847–1852. [Google Scholar] [CrossRef]
- Kisin, E.R.; Yanamala, N.; Farcas, M.T.; Gutkin, D.W.; Shurin, M.R.; Kagan, V.E.; Bugarski, A.D.; Shvedova, A.A. Abnormalities in the Male Reproductive System after Exposure to Diesel and Biodiesel Blend. Environ. Mol. Mutagenesis 2015, 56, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, X.; Jigami, J.; Hasegawa, C.; Suzuki, A.K.; Zhang, Y.; Fujitani, Y.; Nagaoka, K.; Watanabe, G.; Taya, K. Effect of Nanoparticle-Rich Diesel Exhaust on Testosterone Biosynthesis in Adult Male Mice. Inhal. Toxicol. 2012, 24, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Xu, Y.; Pan, H.; Tian, F.; Wang, Y.; Xia, M.; Hu, J.; Yang, M.; Tao, S.; Sun, S.; et al. Chronic Exposure to Diesel Exhaust Particulate Matter Impairs Meiotic Progression during Spermatogenesis in a Mouse Model. Ecotoxicol. Environ. Saf. 2020, 202, 110881. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Li, D.; Chen, J. Mechanistic Toxicity Assessment of Fine Particulate Matter Emitted from Fuel Combustion via Pathway-Based Approaches in Human Cells. Sci. Total Environ. 2022, 806, 150214. [Google Scholar] [CrossRef] [PubMed]
- Oczkowski, M. Health-Promoting Effects of Bioactive Compounds in Blackcurrant (Ribes nigrum L.) Berries. Rocz. Panstw. Zakl. Hig. (Ann. Natl. Inst. Hyg.) 2021, 72, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A.; Mbimba, T.; Thoppil, R.J.; Háznagy-Radnai, E.; Sipos, P.; Darvesh, A.S.; Folkesson, H.G.; Hohmann, J. Anthocyanin-Rich Black Currant (Ribes nigrum L.) Extract Affords Chemoprevention against Diethylnitrosamine-Induced Hepatocellular Carcinogenesis in Rats. J. Nutr. Biochem. 2011, 22, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Pham, T.X.; Bae, M.; Hu, S.; O’Neill, E.; Chun, O.K.; Han, M.J.; Koo, S.I.; Park, Y.K.; Lee, J.Y. Blackcurrant (Ribes Nigrum) Prevents Obesity-Induced Nonalcoholic Steatohepatitis in Mice. Obesity 2019, 27, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Huebbe, P.; Giller, K.; de Pascual-Teresa, S.; Arkenau, A.; Adolphi, B.; Portius, S.; Arkenau, C.N.; Rimbach, G. Effects of Blackcurrant-Based Juice on Atherosclerosis-Related Biomarkers in Cultured Macrophages and in Human Subjects after Consumption of a High-Energy Meal. Br. J. Nutr. 2012, 108, 234–244. [Google Scholar] [CrossRef]
- Li, L.; Hwang, E.; Ngo, H.T.T.; Seo, S.A.; Lin, P.; Gao, W.; Liu, Y.; Yi, T.H. Ribes nigrum L. Prevents UVB-Mediated Photoaging in Human Dermal Fibroblasts: Potential Antioxidant and Antiinflammatory Activity. Photochem. Photobiol. 2018, 94, 1032–1039. [Google Scholar] [CrossRef]
- Kranz, S.; Guellmar, A.; Olschowsky, P.; Tonndorf-Martini, S.; Heyder, M.; Pfister, W.; Reise, M.; Sigusch, B. Antimicrobial Effect of Natural Berry Juices on Common Oral Pathogenic Bacteria. Antibiotics 2020, 9, 533. [Google Scholar] [CrossRef]
- Lyall, K.A.; Hurst, S.M.; Cooney, J.; Jensen, D.; Lo, K.; Hurst, R.D.; Stevenson, L.M. Short-Term Blackcurrant Extract Consumption Modulates Exercise-Induced Oxidative Stress and Lipopolysaccharide-Stimulated Inflammatory Responses. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2009, 297, R70–R81. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.C.; Nunes, A.R.; Falcão, A.; Alves, G.; Silva, L.R. Dietary Effects of Anthocyanins in Human Health: A Comprehensive Review. Pharmaceuticals 2021, 14, 690. [Google Scholar] [CrossRef] [PubMed]
- Dziendzikowska, K.; Gajewska, M.; Wilczak, J.; Mruk, R.; Oczkowski, M.; Żyła, E.; Królikowski, T.; Stachoń, M.; Øvrevik, J.; Myhre, O.; et al. The Effects of 1st and 2nd Generation Biodiesel Exhaust Exposure on Hematological and Biochemical Blood Indices of Fisher344 Male Rats—The FuelHealth Project. Environ. Toxicol. Pharmacol. 2018, 63, 34–47. [Google Scholar] [CrossRef]
- Magnusson, P.; Oczkowski, M.; Øvrevik, J.; Gajewska, M.; Wilczak, J.; Biedrzycki, J.; Dziendzikowska, K.; Kamola, D.; Królikowski, T.; Kruszewski, M.; et al. No Adverse Lung Effects of 7- and 28-Day Inhalation Exposure of Rats to Emissions from Petrodiesel Fuel Containing 20% Rapeseed Methyl Esters (B20) with and without Particulate Filter—The FuelHealth Project. Inhal. Toxicol. 2017, 29, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Valand, R.; Magnusson, P.; Dziendzikowska, K.; Gajewska, M.; Wilczak, J.; Oczkowski, M.; Kamola, D.; Królikowski, T.; Kruszewski, M.; Lankoff, A.; et al. Gene Expression Changes in Rat Brain Regions after 7- and 28 Days Inhalation Exposure to Exhaust Emissions from 1st and 2nd Generation Biodiesel Fuels—The FuelHealth Project. Inhal. Toxicol. 2018, 30, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Odziemkowska, M.; Czarnocka, J.; Frankiewicz, A.; Szewczyńska, M.; Lankoff, A.; Gromadzka-Ostrowska, J.; Mruk, R. Chemical Characterization of Exhaust Gases from Compression Ignition Engine Fuelled with Various Biofuels. Pol. J. Environ. Stud. 2017, 26, 1183–1190. [Google Scholar] [CrossRef]
- Magnusson, P.; Dziendzikowska, K.; Oczkowski, M.; Øvrevik, J.; Eide, D.M.; Brunborg, G.; Gutzkow, K.B.; Instanes, C.; Gajewska, M.; Wilczak, J.; et al. Lung Effects of 7- and 28-Day Inhalation Exposure of Rats to Emissions from 1st and 2nd Generation Biodiesel Fuels with and without Particle Filter—The FuelHealth Project. Environ. Toxicol. Pharmacol. 2019, 67, 8–20. [Google Scholar] [CrossRef] [PubMed]
- OECD/OCDE. OECD Guidelines on the Testing of Chemicals No 412. Subacute Inhalation Toxicity: 28-day Study; OECD/OCDE: Paris, France, 2009.
- Khan, I.A.; Reddy, B.V.; Mahboob, M.; Rahman, M.F.; Jamil, K. Effects of Phosphorothionate on the Reproductive System of Male Rats. J. Environ. Sci. Health Part B 2001, 36, 445–456. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for Lipid Peroxides in Animal Tissues by Thiobarbituric Acid Reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Yagi, K. Simple Procedure for Specific Assay of Lipid Hydroperoxides in Serum or Plasma. In Free Radical and Antioxidant Protocols; Humana Press: Totowa, NJ, USA, 1998; pp. 107–110. [Google Scholar]
- Suchecka, D.; Harasym, J.P.; Wilczak, J.; Gajewska, M.; Oczkowski, M.; Gudej, S.; Błaszczyk, K.; Kamola, D.; Filip, R.; Gromadzka-Ostrowska, J. Antioxidative and Anti-Inflammatory Effects of High Beta-Glucan Concentration Purified Aqueous Extract from Oat in Experimental Model of LPS-Induced Chronic Enteritis. J. Funct. Foods 2015, 14, 244–254. [Google Scholar] [CrossRef]
- Han, D.; Hanawa, N.; Saberi, B.; Kaplowitz, N. Mechanisms of Liver Injury. III. Role of Glutathione Redox Status in Liver Injury. Am. J. Physiol. -Gastrointest. Liver Physiol. 2006, 291, G1–G7. [Google Scholar] [CrossRef] [PubMed]
- Rebrin, I.; Forster, M.J.; Sohal, R.S. Effects of Age and Caloric Intake on Glutathione Redox State in Different Brain Regions of C57BL/6 and DBA/2 Mice. Brain Res. 2007, 1127, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Wilczak, J.; Błaszczyk, K.; Kamola, D.; Gajewska, M.; Harasym, J.P.; Jałosińska, M.; Gudej, S.; Suchecka, D.; Oczkowski, M.; Gromadzka-Ostrowska, J. The Effect of Low or High Molecular Weight Oat Beta-Glucans on the Inflammatory and Oxidative Stress Status in the Colon of Rats with LPS-Induced Enteritis. Food Funct. 2015, 6, 590–603. [Google Scholar] [CrossRef] [PubMed]
- TIBCO Software Inc. STATISTICA (Data Analysis Software System), Version 13.3; StatSoft Polska: Krakow, Poland, 2017. [Google Scholar]
- Prism—GraphPad. Available online: https://www.graphpad.com/scientific-software/prism/ (accessed on 6 May 2022).
- Cassee, F.R.; Campbell, A.; Boere, A.J.F.; McLean, S.G.; Duffin, R.; Krystek, P.; Gosens, I.; Miller, M.R. The Biological Effects of Subacute Inhalation of Diesel Exhaust Following Addition of Cerium Oxide Nanoparticles in Atherosclerosis-Prone Mice. Environ. Res. 2012, 115, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pires, A.; de Melo, E.N.; Mauad, T.; Nascimento Saldiva, P.H.; de Siqueira Bueno, H.M. Pre- and Postnatal Exposure to Ambient Levels of Urban Particulate Matter (PM2.5) Affects Mice Spermatogenesis. Inhal. Toxicol. 2011, 23, 237–245. [Google Scholar] [CrossRef]
- Watanabe, N.; Oonuki, Y. Inhalation of Diesel Engine Exhaust Affects Spermatogenesis in Growing Male Rats. Environ. Health Perspect. 1999, 107, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Ono, N.; Oshio, S.; Niwata, Y.; Yoshida, S.; Tsukue, N.; Sugawara, I.; Takano, H.; Takeda, K. Prenatal Exposure to Diesel Exhaust Impairs Mouse Spermatogenesis. Inhal. Toxicol. 2007, 19, 275–281. [Google Scholar] [CrossRef]
- Kubo-Irie, M.; Oshio, S.; Niwata, Y.; Ishihara, A.; Sugawara, I.; Takeda, K. Pre- and Postnatal Exposure to Low-Dose Diesel Exhaust Impairs Murine Spermatogenesis. Inhal. Toxicol. 2011, 23, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Aatola, H.; Larmi, M.; Sarjovaara, T.; Mikkonen, S. Hydrotreated vegetable oil (HVO) as a renewable diesel fuel: Trade-off between NOx, particulate emission, and fuel consumption of a heavy duty engine. SAE Int. J. Engines 2009, 1, 1251–1262. [Google Scholar] [CrossRef]
- Karavalakis, G.; Fontaras, G.; Ampatzoglou, D.; Kousoulidou, M.; Stournas, S.; Samaras, Z.; Bakeas, E. Effects of Low Concentration Biodiesel Blends Application on Modern Passenger Cars. Part 3: Impact on PAH, Nitro-PAH, and Oxy-PAH Emissions. Environ. Pollut. 2010, 158, 1584–1594. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Ge, J.C.; Choi, N.J. A Comprehensive Review of the Application Characteristics of Biodiesel Blends in Diesel Engines. Appl. Sci. 2020, 10, 8015. [Google Scholar] [CrossRef]
- Asadi, N.; Bahmani, M.; Kheradmand, A.; Rafieian-Kopaei, M. The Impact of Oxidative Stress on Testicular Function and the Role of Antioxidants in Improving It: A Review. J. Clin. Diagn. Res. 2017, 11, IE01–IE05. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Sengupta, P.; Slama, P.; Roychoudhury, S. Oxidative Stress, Testicular Inflammatory Pathways, and Male Reproduction. Int. J. Mol. Sci. 2021, 22, 10043. [Google Scholar] [CrossRef] [PubMed]
- Landau, G.; Kodali, V.K.; Malhotra, J.D.; Kaufman, R.J. Detection of Oxidative Damage in Response to Protein Misfolding in the Endoplasmic Reticulum. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2013; Volume 526, pp. 231–250. ISBN 9780124058835. [Google Scholar]
- Ghani, M.A.; Barril, C.; Bedgood, D.R.; Prenzler, P.D. Measurement of Antioxidant Activity with the Thiobarbituric Acid Reactive Substances Assay. Food Chem. 2017, 230, 195–207. [Google Scholar] [CrossRef]
- Liu, B.; Wu, S.D.; Shen, L.J.; Zhao, T.X.; Wei, Y.; Tang, X.L.; Long, C.L.; Zhou, Y.; He, D.W.; Lin, T.; et al. Spermatogenesis Dysfunction Induced by PM2.5 from Automobile Exhaust via the ROS-Mediated MAPK Signaling Pathway. Ecotoxicol. Environ. Saf. 2019, 167, 161–168. [Google Scholar] [CrossRef]
- Liu, B.; Shen, L.-J.; Zhao, T.-X.; Sun, M.; Wang, J.-K.; Long, C.-L.; He, D.-W.; Lin, T.; Wu, S.-D.; Wei, G.-H. Automobile Exhaust-Derived PM2.5 Induces Blood-Testis Barrier Damage through ROS-MAPK-Nrf2 Pathway in Sertoli Cells of Rats. Ecotoxicol. Environ. Saf. 2020, 189, 110053. [Google Scholar] [CrossRef] [PubMed]
- Bu, T.L.; Jia, Y.D.; Lin, J.X.; Mi, Y.L.; Zhang, C.Q. Alleviative Effect of Quercetin on Germ Cells Intoxicated by 3-Methyl-4-Nitrophenol from Diesel Exhaust Particles. J. Zhejiang Univ. Sci. B 2012, 13, 318–326. [Google Scholar] [CrossRef]
- Girotti, A.W. Lipid Hydroperoxide Generation, Turnover, and Effector Action in Biological Systems. J. Lipid Res. 1998, 39, 1529–1542. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Olkkonen, V.M.; Hynynen, R. Interactions of Oxysterols with Membranes and Proteins. Mol. Asp. Med. 2009, 30, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.; Campo, A.; Fulton, E.; Corwin, A.; Jerome, W.G.; O’Connor, M.S. 7-Ketocholesterol in Disease and Aging. Redox Biol. 2020, 29, 101380. [Google Scholar] [CrossRef]
- Choi, C.; Finlay, D.K. Diverse Immunoregulatory Roles of Oxysterols—The Oxidized Cholesterol Metabolites. Metabolites 2020, 10, 384. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Zhong, J.; Maiseyeu, A.; Gopalakrishnan, B.; Villamena, F.A.; Chen, L.C.; Harkema, J.R.; Sun, Q.; Rajagopalan, S. CD36-Dependent 7-Ketocholesterol Accumulation in Macrophages Mediates Progression of Atherosclerosis in Response to Chronic Air Pollution Exposure. Circ. Res. 2014, 115, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Hesterberg, T.W.; Long, C.M.; Bunn, W.B.; Sax, S.N.; Lapin, C.A.; Valberg, P.A. Non-Cancer Health Effects of Diesel Exhaust: A Critical Assessment of Recent Human and Animal Toxicological Literature. Crit. Rev. Toxicol. 2009, 39, 195–227. [Google Scholar] [CrossRef]
- Mokrzyński, K.; Krzysztyńska-Kuleta, O.; Zawrotniak, M.; Sarna, M.; Sarna, T. Fine Particulate Matter-induced Oxidative Stress Mediated by Uva-visible Light Leads to Keratinocyte Damage. Int. J. Mol. Sci. 2021, 22, 10645. [Google Scholar] [CrossRef] [PubMed]
- Czarnocka, J.; Odziemkowska, M. Characterization of the Polycyclic Aromatic Hydrocarbons Emitted from a Compression Ignition Engine Powered with Biofuels of the 1st and 2nd Generation. Chemik 2016, 70, 419–425. [Google Scholar]
- Lankoff, A.; Brzoska, K.; Czarnocka, J.; Kowalska, M.; Lisowska, H.; Mruk, R.; Øvrevik, J.; Wegierek-Ciuk, A.; Zuberek, M.; Kruszewski, M. A Comparative Analysis of in Vitro Toxicity of Diesel Exhaust Particles from Combustion of 1st- and 2nd-Generation Biodiesel Fuels in Relation to Their Physicochemical Properties—The FuelHealth Project. Environ. Sci. Pollut. Res. 2017, 24, 19357–19374. [Google Scholar] [CrossRef]
- Øvrevik, J.; Refsnes, M.; Låg, M.; Holme, J.A.; Schwarze, P.E. Activation of Proinflammatory Responses in Cells of the Airway Mucosa by Particulate Matter: Oxidant- and Non-Oxidant-Mediated Triggering Mechanisms. Biomolecules 2015, 5, 1399–1440. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Zhu, T.; Kipen, H.; Wang, G.; Hu, M.; Guo, Q.; Ohman-Strickland, P.; Lu, S.E.; Wang, Y.; Zhu, P.; et al. Comparisons of Ultrafine and Fine Particles in Their Associations with Biomarkers Reflecting Physiological Pathways. Environ. Sci. Technol. 2014, 48, 5264–5273. [Google Scholar] [CrossRef]
- Vara, D.; Pula, G. Reactive Oxygen Species: Physiological Roles in the Regulation of Vascular Cells. Curr. Mol. Med. 2014, 14, 1103–1125. [Google Scholar] [CrossRef] [PubMed]
- Kanti Das, T.; Wati, M.R.; Fatima-Shad, K. Oxidative Stress Gated by Fenton and Haber Weiss Reactions and Its Association with Alzheimer’s Disease. Arch. Neurosci. 2014, 2, 20078. [Google Scholar] [CrossRef]
- Siti, H.N.; Kamisah, Y.; Kamsiah, J. The Role of Oxidative Stress, Antioxidants and Vascular Inflammation in Cardiovascular Disease (A Review). Vasc. Pharmacol. 2015, 71, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Kuciel-Lewandowska, J.; Gnus, J.; Pawlik-Sobecka, L.; Płaczkowska, S.; Kokot, I.; Kasperczak, M.; Paprocka-Borowicz, M. The Assessment of the Integrated Antioxidant System of the Body in the Course of Radon Therapy: A Pilot Study. BioMed Res. Int. 2018, 2018, 6038106. [Google Scholar] [CrossRef]
- Sies, H. Oxidative Stress: Concept and Some Practical Aspects. Antioxidants 2020, 9, 852. [Google Scholar] [CrossRef] [PubMed]
- Banjac, L.; Banjac, G.; Kotur-Stevuljević, J.; Spasojević-Kalimanovska, V.; Gojković, T.; Bogavac-Stanojević, N.; Jelić-Ivanović, Z.; Banjac, G. Pro-Oxidants and Antioxidants in Retinopathy of Prematurity. Acta Clin. Croat. 2018, 57, 458–463. [Google Scholar] [CrossRef]
- Nemmar, A.; Zia, S.; Subramaniyan, D.; Al-Amri, I.; Al Kindi, M.A.; Ali, B.H. Interaction of Diesel Exhaust Particles with Human, Rat and Mouse Erythrocytes in Vitro. Cell. Physiol. Biochem. 2012, 29, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-Y.; Liao, J.-W.; Liu, Y.-C.; Lu, S.-Y.; Chou, C.-P.; Chan, W.-H.; Chen, S.-U.; Ueng, T.-H. Motorcycle Exhaust Induces Reproductive Toxicity and Testicular Interleukin-6 in Male Rats. Toxicol. Sci. 2008, 103, 137–148. [Google Scholar] [CrossRef]
- Cao, X.-N.; Shen, L.-J.; Wu, S.-D.; Yan, C.; Zhou, Y.; Xiong, G.; Wang, Y.-C.; Liu, Y.; Liu, B.; Tang, X.-L.; et al. Urban Fine Particulate Matter Exposure Causes Male Reproductive Injury through Destroying Blood-Testis Barrier (BTB) Integrity. Toxicol. Lett. 2017, 266, 1–12. [Google Scholar] [CrossRef]
- Aquilano, K.; Baldelli, S.; Ciriolo, M.R. Glutathione: New Roles in Redox Signalling for an Old Antioxidant. Front. Pharmacol. 2014, 5, 196. [Google Scholar] [CrossRef]
- Nur, E.; Verwijs, M.; de Waart, D.R.; Schnog, J.J.B.; Otten, H.M.; Brandjes, D.P.; Biemond, B.J.; Elferink, R.P.J.O. Increased Efflux of Oxidized Glutathione (GSSG) Causes Glutathione Depletion and Potentially Diminishes Antioxidant Defense in Sickle Erythrocytes. Biochim. Biophys. Acta Mol. Basis Dis. 2011, 1812, 1412–1417. [Google Scholar] [CrossRef]
- Zheng, S.; Yumei, F.; Chen, A. De Novo Synthesis of Glutathione Is a Prerequisite for Curcumin to Inhibit Hepatic Stellate Cell (HSC) Activation. Free. Radic. Biol. Med. 2007, 43, 444–453. [Google Scholar] [CrossRef]
- Li, J.; Gao, L.; Zhu, B.B.; Lin, Z.J.; Chen, J.; Lu, X.; Wang, H.; Zhang, C.; Chen, Y.H.; Xu, D.X. Long-Term 1-Nitropyrene Exposure Induces Endoplasmic Reticulum Stress and Inhibits Steroidogenesis in Mice Testes. Chemosphere 2020, 251, 126336. [Google Scholar] [CrossRef]
- Lee, S.G.; Vance, T.M.; Nam, T.G.; Kim, D.O.; Koo, S.I.; Chun, O.K. Contribution of Anthocyanin Composition to Total Antioxidant Capacity of Berries. Plant Foods Hum. Nutr. 2015, 70, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Sivasinprasasn, S.; Pantan, R.; Thummayot, S.; Tocharus, J.; Suksamrarn, A.; Tocharus, C. Cyanidin-3-Glucoside Attenuates Angiotensin II-Induced Oxidative Stress and Inflammation in Vascular Endothelial Cells. Chem. -Biol. Interact. 2016, 260, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Chen, W.; Tu, P.; Jia, R.; Liu, Y.; Li, Y.; Tang, Q.; Zheng, X.; Chu, Q. Food-Derived Cyanidin-3-O-Glucoside Alleviates Oxidative Stress: Evidence from the Islet Cell Line and Diabetic Db/Db Mice. Food Funct. 2021, 12, 11599–11610. [Google Scholar] [CrossRef] [PubMed]
- Sukprasansap, M.; Chanvorachote, P.; Tencomnao, T. Cyanidin-3-Glucoside Activates Nrf2-Antioxidant Response Element and Protects against Glutamate-Induced Oxidative and Endoplasmic Reticulum Stress in HT22 Hippocampal Neuronal Cells. BMC Complementary Med. Ther. 2020, 20, 46. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Jia, Q.; Wang, Y.; Zhang, Y.; Xia, M. The Anthocyanin Cyanidin-3-O-β-Glucoside, a Flavonoid, Increases Hepatic Glutathione Synthesis and Protects Hepatocytes against Reactive Oxygen Species during Hyperglycemia: Involvement of a CAMP-PKA-Dependent Signaling Pathway. Free. Radic. Biol. Med. 2012, 52, 314–327. [Google Scholar] [CrossRef]
- Casati, L.; Pagani, F.; Fibiani, M.; Scalzo, R.L.; Sibilia, V. Potential of Delphinidin-3-Rutinoside Extracted from Solanum melongena L. as Promoter of Osteoblastic MC3T3-E1 Function and Antagonist of Oxidative Damage. Eur. J. Nutr. 2019, 58, 1019–1032. [Google Scholar] [CrossRef] [PubMed]
- Šimerdová, B.; Bobríková, M.; Lhotská, I.; Kaplan, J.; Křenová, A.; Šatínský, D. Evaluation of Anthocyanin Profiles in Various Blackcurrant Cultivars over a Three-Year Period Using a Fast Hplc-Dad Method. Foods 2021, 10, 1745. [Google Scholar] [CrossRef]
- Tsuda, T.; Watanabe, M.; Ohshima, K.; Norinobu, S.; Choi, S.W.; Kawakishi, S.; Osawa, T. Antioxidative Activity of the Anthocyanin Pigments Cyanidin 3-O-β-d-Glucoside and Cyanidin. J. Agric. Food Chem. 1994, 42, 2407–2410. [Google Scholar] [CrossRef]
- Tsuda, T.; Ohshima, K.; Kawakishi, S.; Osawa, T. Oxidation Products of Cyanidin 3-O-β-D-Glucoside with a Free Radical Initiator. Lipids 1996, 31, 1259–1263. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, T.; Horio, F.; Kitoh, J.; Osawa, T. Protective Effects of Dietary Cyanidin 3-O-β-D-Glucoside on Liver Ischemia-Reperfusion Injury in Rats. Arch. Biochem. Biophys. 1999, 368, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Nowak, D.; Gośliński, M.; Szwengiel, A. Multidimensional Comparative Analysis of Phenolic Compounds in Organic Juices with High Antioxidant Capacity. J. Sci. Food Agric. 2017, 97, 2657–2663. [Google Scholar] [CrossRef]
- Tan, C.; Li, D.; Wang, H.; Tong, Y.; Zhao, Y.; Deng, H.; Kong, Y.; Shu, C.; Yan, T.; Meng, X. Effects of High Hydrostatic Pressure on the Binding Capacity, Interaction, and Antioxidant Activity of the Binding Products of Cyanidin-3-Glucoside and Blueberry Pectin. Food Chem. 2021, 344, 128731. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No 152/2009 of 27 January 2009 laying down the methods of sampling and analysis for the official control of feed (Text with EEA relevance). Off. J. Eur. Union 2009, L54/1.
- PN-EN ISO 6865:2002; Animal Feeding Stuffs—Determination of Crude Fibre Content-Method with Intermediate Filtration. Polish Committee of Standardization: Warsaw, Poland, 2002.
- Hallmann, E.; Kazimierczak, R.; Marszałek, K.; Drela, N.; Kiernozek, E.; Toomik, P.; Matt, D.; Luik, A.; Rembiałkowska, E. The Nutritive Value of Organic and Conventional White Cabbage (Brassica oleracea L. Var. Capitata) and Anti-Apoptotic Activity in Gastric Adenocarcinoma Cells of Sauerkraut Juice Produced Therof. J. Agric. Food Chem 2017, 65, 8171–8183. [Google Scholar] [CrossRef] [PubMed]
- Dóka, O.; Ficzek, G.; Bicanic, D.; Spruijt, R.; Luterotti, S.; Tóth, M.; Buijnsters, J.G.; Végvári, G. Direct photothermal techniques for rapid quantification of total anthocyanin content in sour cherry cultivars. Talanta 2011, 84, 341–346. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oczkowski, M.; Wilczak, J.; Dziendzikowska, K.; Øvrevik, J.; Myhre, O.; Lankoff, A.; Kruszewski, M.; Gromadzka-Ostrowska, J. Dietary Intervention with Blackcurrant Pomace Protects Rats from Testicular Oxidative Stress Induced by Exposition to Biodiesel Exhaust. Antioxidants 2022, 11, 1562. https://doi.org/10.3390/antiox11081562
Oczkowski M, Wilczak J, Dziendzikowska K, Øvrevik J, Myhre O, Lankoff A, Kruszewski M, Gromadzka-Ostrowska J. Dietary Intervention with Blackcurrant Pomace Protects Rats from Testicular Oxidative Stress Induced by Exposition to Biodiesel Exhaust. Antioxidants. 2022; 11(8):1562. https://doi.org/10.3390/antiox11081562
Chicago/Turabian StyleOczkowski, Michał, Jacek Wilczak, Katarzyna Dziendzikowska, Johan Øvrevik, Oddvar Myhre, Anna Lankoff, Marcin Kruszewski, and Joanna Gromadzka-Ostrowska. 2022. "Dietary Intervention with Blackcurrant Pomace Protects Rats from Testicular Oxidative Stress Induced by Exposition to Biodiesel Exhaust" Antioxidants 11, no. 8: 1562. https://doi.org/10.3390/antiox11081562