
Changes in Content of Bioactive Compounds and Antioxidant Activity Induced in Needles of Different Half-Sib Families of Norway Spruce (Picea abies (L.) H. Karst) by Seed Treatment with Cold Plasma

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Collection
2.2. Seeds Treatment with Cold Plasma
2.3. Cultivation of Seedlings and Sample Collection for Experiments
2.4. Sample Preparation
2.5. Quantification of Total Flavonoid Content
2.6. Detection of Radical Scavenging Activity Using ABTS and DPPH Assays
2.7. Quantification of Organic Acids by HPLC
2.8. Statistical Data Analysis
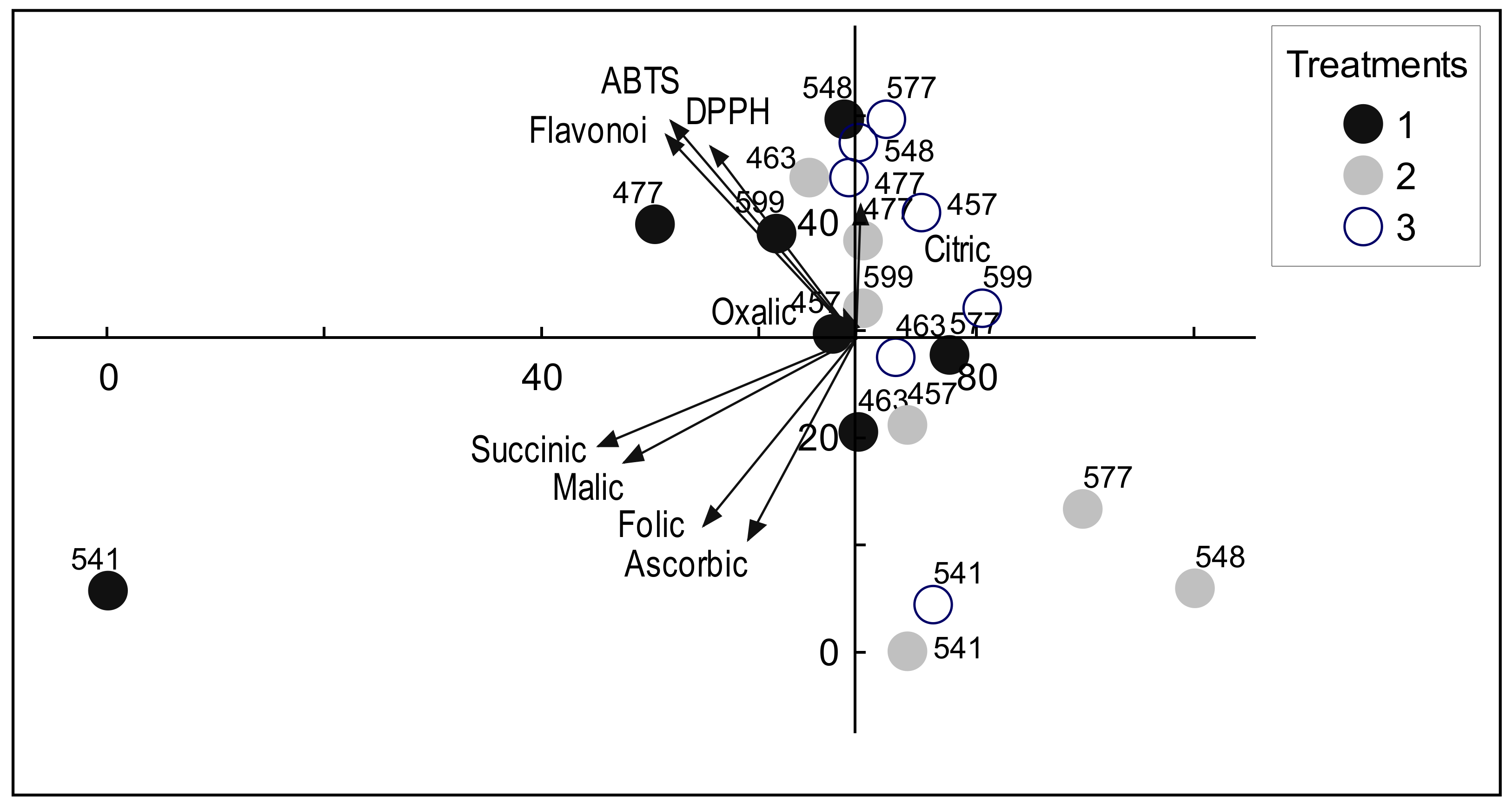
3. Results
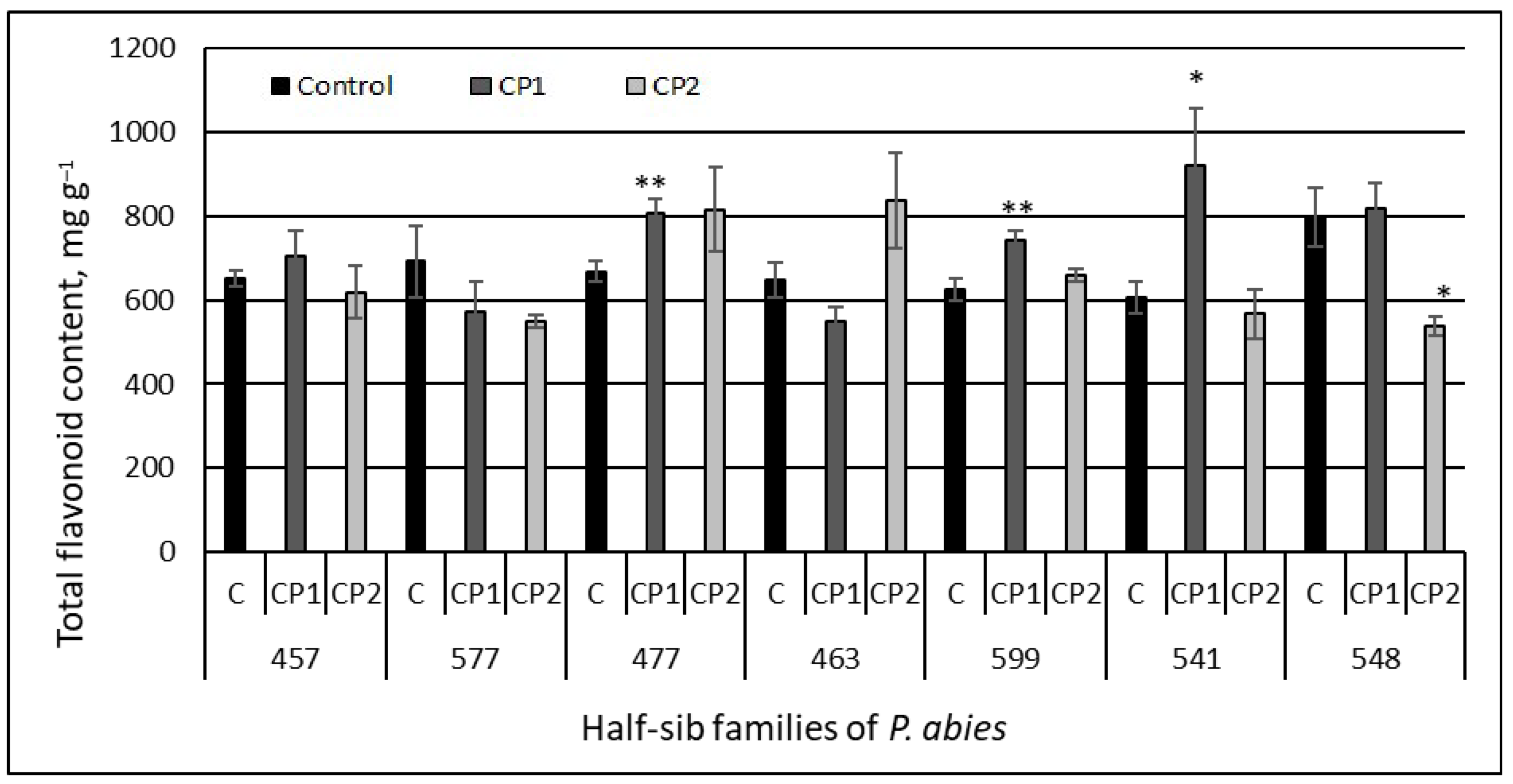
3.1. Total Flavonoid Content
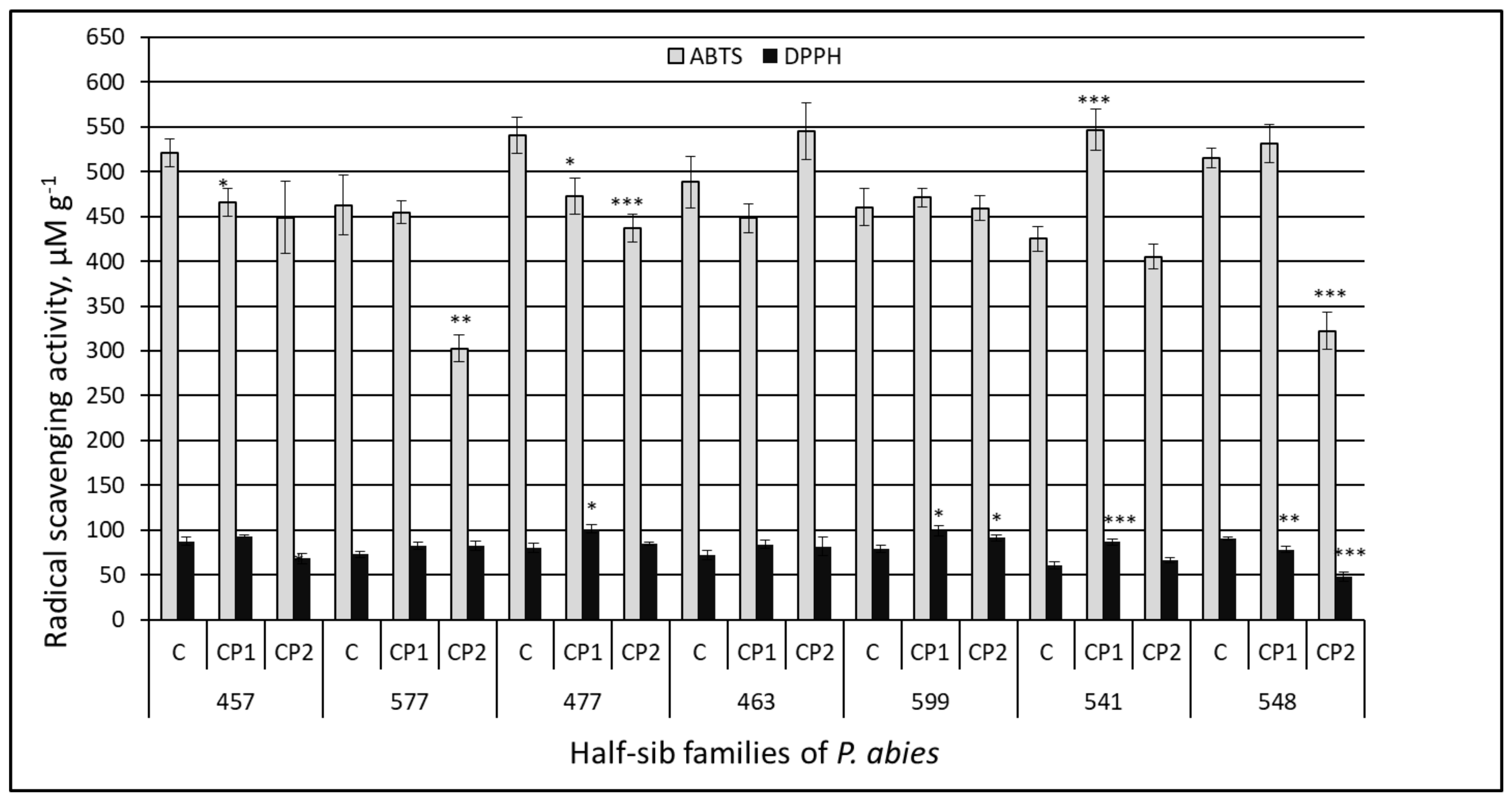
3.2. Antiradical Activity
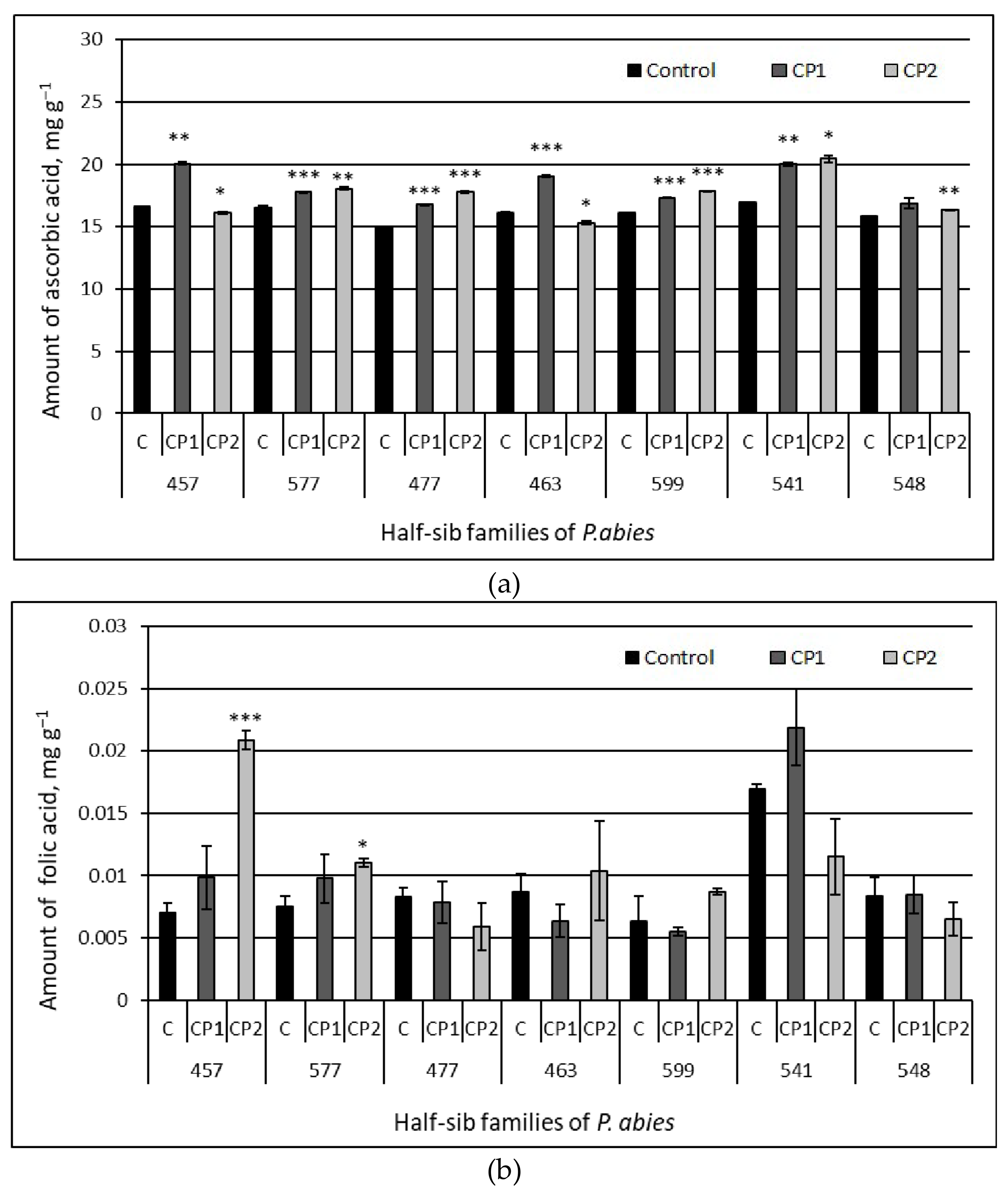
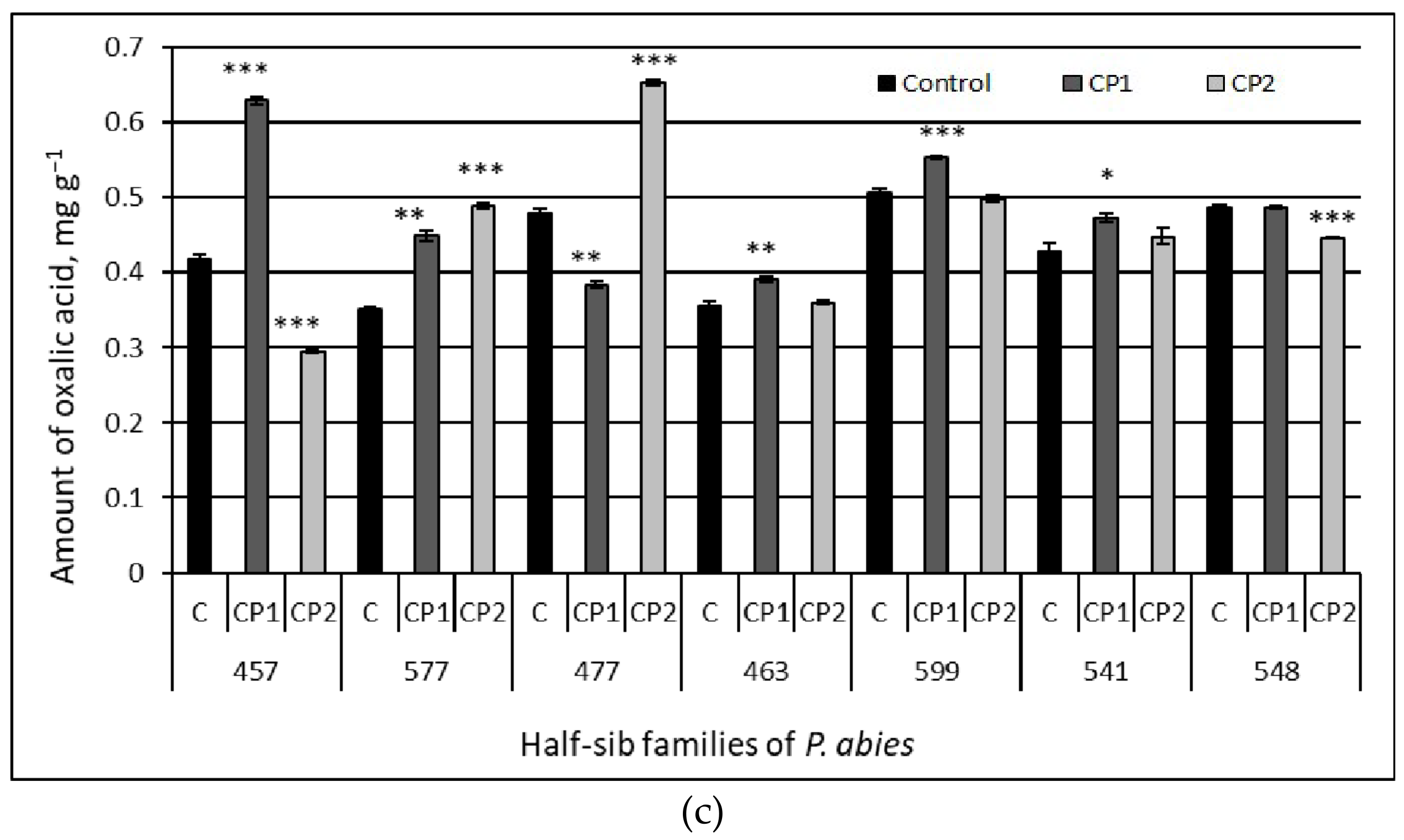
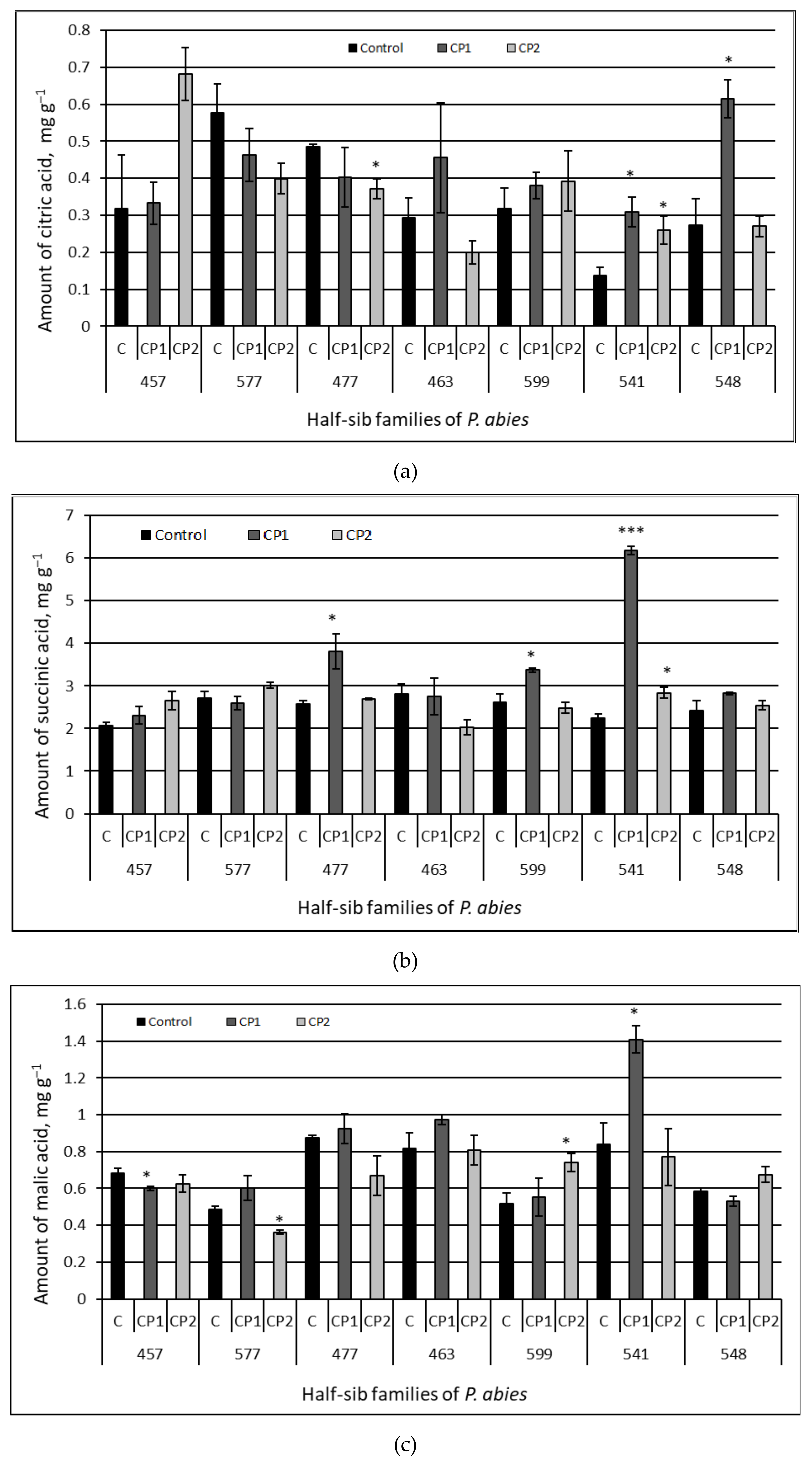
3.3. Amount of Organic Acids
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviation
| ABTS | 2:2’-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) |
| CP | cold plasma |
| DBD | dielectric barrier discharge |
| DPPH | 2,2-diphenyl-1-picryl-hydrazyl-hydrate |
| NO | nitric oxide |
| TFC | total flavonoid content |
References
- Roitto, M.; Siwale, W.; Tanner, J.; Ilvesniemi, H.; Julkunen-Tiitto, R.; Verkasalo, E. Characterization of extractives in tree biomass and by-products of plywood and saw mills from Finnish birch in different climatic regions for value-added chemical products. In Proceedings of the 5th International scientific conference on hardwood processing, Quebec City, QC, Canada, 15–17 September 2015; pp. 174–181. [Google Scholar]
- Radulescu, V.; Ilies, D.C.; Voiculescu, I.; Iovu-Adrian, B.; Craciunescu, A. Determination of ascorbic acid in shoots from different coniferous species by HPLC. Farmacia 2013, 61, 1158–1166. [Google Scholar]
- Virjamo, V.; Julkunen-Tiitto, R. Shoot development of Norway spruce (Picea abies) involves changes in piperidine alkaloids and condensed tannins. Trees 2014, 28, 427–437. [Google Scholar] [CrossRef]
- Metsämuuronen, S.; Sirén, H. Bioactive phenolic compounds, metabolism and properties: A review on valuable chemical compounds in Scots pine and Norway spruce. Phytochem. Rev. 2019, 18, 623–664. [Google Scholar] [CrossRef]
- Zulaica-Villagomez, H.; Peterson, D.M.; Herrin, L.; Young, R.A. Antioxidant activity of different components of pine species. Holzforschung 2005, 59, 156–162. [Google Scholar] [CrossRef]
- Jyske, T.; Järvenpää, E.; Kunnas, S.; Sarjala, T.; Raitanen, J.E.; Mäki, M.; Pastell, H.; Korpinen, R.; Kaseva, J.; Tupasela, T. Sprouts and Needles of Norway Spruce (Picea abies (L.) Karst.) as Nordic Specialty—Consumer Acceptance, Stability of Nutrients, and Bioactivities during Storage. Molecules 2020, 25, 4187. [Google Scholar] [CrossRef] [PubMed]
- Ganthaler, A.; Stoggl, Q.; Kranner, I.; Mayr, S. Foliar Phenolic Compounds in Norway Spruce with Varying Susceptibility to Chrysomyxa rhododendri: Analyses of Seasonal and Infection-Induced Accumulation Patterns. Front. Plant Sci. 2017, 8, 1173. [Google Scholar] [CrossRef] [PubMed]
- Lalhminghlui, K.; Jagetia, G.C. Evaluation of the free-radical scavenging and antioxidant activities of Chilauni, Schima wallichii Korth in vitro. Future Sci. 2018, 4, FSO272. [Google Scholar] [CrossRef]
- Tanase, C.; Cosarca, S.; Muntean, D. A critical review of phenolic compounds extracted from the bark of woody vascular plants and their potential biological activity. Molecules 2019, 24, 1182. [Google Scholar] [CrossRef]
- Kopaczyk, J.M.; Warguła, J.; Jelonek, T. The variability of terpenes in conifers under developmental and environmental stimuli. Environ. Exp. Bot. 2020, 180. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Neha, K.; Haider, R.; Pathak, A.; Yra, S. Medicinal prospects of antioxidants: A review. Eur. J. Med. Chem. 2019, 178, 687–704. [Google Scholar] [CrossRef]
- Dable-Tupas, G.; Otero, M.C.B.; Bernolo, L. Functional Foods and Health Benefits. In Functional Foods and Nutraceutical; Egbuna, C., Dable-Tupas, G., Eds.; Springer: New York, NY, USA, 2020; pp. 1–11. [Google Scholar]
- Kurlbaum, M.; Högger, P. Plasma protein binding of polyphenols from maritime pine bark extract (USP). J. Pharm. Biomed. Ana. 2011, 54, 127–132. [Google Scholar] [CrossRef]
- Samieri, C.; Sun, Q.; Townsend, M.K.; Rimm, E.B.; Grodstein, F. Dietary flavonoid intake at midlife and healthy aging in women. AJCN 2014, 100, 1489–1497. [Google Scholar] [CrossRef]
- Werkelin, J.; Lindberg, D.; Boström, D.; Skrifvars, B.J.; Hupa, M. Ash-forming elements in four Scandinavian wood species part 3: Combustion of five spruce samples. Biomass Bioenergy 2011, 35, 725–733. [Google Scholar] [CrossRef]
- Tomášková, I.; Pastierovič, F.; Krejzková, A.; Čepl, J.; Hradecký, J. Norway spruce ecotypes distinguished by chlorophyll a fuorescence kinetics. Acta. Physiol. Plant. 2021, 43, 24. [Google Scholar] [CrossRef]
- OuYang, F.; Ou, Y.; Zhu, T.; Ma, J.; An, S.; Zhao, J.; Wang, J.; Kong, L.; Zhang, H.; Tigabu, M. Growth and Physiological Responses of Norway Spruce (Picea abies (L.) H. Karst) Supplemented with Monochromatic Red, Blue and Far-Red Light. Forests 2012, 12, 164. [Google Scholar] [CrossRef]
- Weltmann, K.; Kolb, J.F.; Holub, M.; Uhrlandt, D.; Šimek, M.; Ostrikov, K.; Hamaguchi, S.; Cvelbar, U.; Cernák, M.; Locke, B.; et al. The future for plasma science and technology. Plasma Proc. Polym. 2018, 16, 118. [Google Scholar] [CrossRef]
- Waskow, A.; Butscher, D.; Oberbossel, G.; Klöti, D.; Rudolf von Rohr, P.; Büttner-Mainik, A.; Drissner, D.; Schuppler, M. Low-energy electron beam has severe impact on seedling development compared to cold atmospheric pressure plasma. Sci. Rep. 2021, 11, 16373. [Google Scholar] [CrossRef]
- Waskow, A.; Howling, A.; Furno, I. Mechanisms of Plasma-Seed Treatments as a Potential Seed Processing Technology. Front. Phys. 2021, 14, 617345. [Google Scholar] [CrossRef]
- Attri, P.; Ishikawa, K.; Okumura, T.; Kazunori, K.; Shiratani, M. Plasma Agriculture from Laboratory to Farm: A Review. Processes 2020, 8, 1002. [Google Scholar] [CrossRef]
- Starič, P.; Vogel-Mikuš, K.; Mozetič, M.; Junkar, I. Effects of Nonthermal Plasma on Morphology, Genetics and Physiology of Seeds: A Review. Plants 2020, 9, 1736. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, B.; Adhikari, M.; Park, G. The Effects of Plasma on Plant Growth, Development, and Sustainability. Appl. Sci. 2020, 10, 6045. [Google Scholar] [CrossRef]
- Holubová, L.; Kyzek, S.; Ďurovcová, I.; Fabová, J.; Horváthová, E.; Ševčovičová, A.; Gálová, E. Non-Thermal Plasma—A New Green Priming Agent for Plants? Int. J. Mol. Sci. 2020, 21, 9466. [Google Scholar] [CrossRef] [PubMed]
- Saberi, M.; Modarres-Sanavy, S.A.M.; Zare, R.; Ghomi, H. Amelioration of Photosynthesis and Quality of Wheat under Non-thermal Radio Frequency Plasma Treatment. Sci. Rep. 2018, 8, 11655. [Google Scholar] [CrossRef]
- Ivankov, A.; Naučienė, Z.; Degutytė-Fomins, L.; Žūkienė, R.; Januškaitienė, I.; Malakauskienė, A.; Jakštas, V.; Ivanauskas, L.; Lyushkevich, V.; Mildažienė, V. Changes in Agricultural Performance of Common Buckwheat Induced by Seed Treatment with Cold Plasma and Electromagnetic Field. Appl. Sci. 2021, 11, 4391. [Google Scholar] [CrossRef]
- Mildažienė, V.; Paužaitė, G.; Naučienė, Z.; Žūkienė, R.; Malakauskienė, A.; Norkevičienė, E.; Šlepetienė, A.; Stukonis, V.; Olšauskaitė, V. Effect of seed treatment with cold plasma and electromagnetic field on red clover germination, growth and content of major isoflavones. J. Phys. D Appl. Phys. 2020, 53, 26. [Google Scholar] [CrossRef]
- Mildaziene, V.; Ivankov, A.; Sera, B.; Baniulis, D. Biochemical and Physiological Plant Processes Affected by Seed Treatment with Non-Thermal Plasma. Plants 2022, 11, 856. [Google Scholar] [CrossRef]
- Ivankov, A.; Nauciene, Z.; Zukiene, R.; Degutyte-Fomins, L.; Malakauskiene, A.; Kraujalis, P.; Lyushkevich, V.; Mildaziene, V. Changes in Growth and Production of Non-Psychotropic Cannabinoids Induced by Pre-Sowing Treatment of Hemp Seeds with Cold Plasma, Vacuum and Electromagnetic Field. Appl. Sci. 2020, 11, 8591. [Google Scholar] [CrossRef]
- Sarinont, T.; Amano, T.; Koga, K.; Shiratani, M.; Hayashi, N. Effects of Atmospheric Air Plasma Irradiation to Seeds of Radish Sprouts on Chlorophyll and Carotenoids Concentrations in their Leaves. MRS Proc. 2014, 1723, 34–38. [Google Scholar] [CrossRef]
- Ji, S.H.; Choi, K.H.; Pengkit, A.; Im, J.S.; Kim, J.S.; Kim, Y.H.; Park, Y.; Hong, E.J.; Jung, S.K.; Choi, E.H.; et al. Effects of high voltage nanosecond pulsed plasma and micro DBD plasma on seed germination, growth development and physiological activities in spinach. Arch. Biochem. Biophys. 2016, 605, 117–128. [Google Scholar] [CrossRef]
- Guo, Q.; Wang, Y.; Zhang, H.; Qu, G.; Wang, T.; Sun, Q.; Liang, D. Alleviation of adverse effects of drought stress on wheat seed germination using atmospheric dielectric barrier discharge plasma treatment. Sci. Rep. 2017, 7, 3893. [Google Scholar] [CrossRef]
- Thor, W.; Stenlid, J. Heterobasidion annosum infection of Picea abies following manual or mechanized stump treatment. Scand. J. For. Res. 2007, 20, 154–164. [Google Scholar] [CrossRef]
- Oliva, J.; Romeralo, C.; Stenlid, J.; Thor, W.; Oliva. Accuracy of the Rotfnder instrument in detecting decay on Norway spruce (Picea abies) trees. Ecol. Manag. 2011, 262, 1378–1386. [Google Scholar] [CrossRef]
- Marčiulynas, A.; Sirgedaitė-Šėžienė, V.; Žemaitis, P.; Baliuckas, V. The resistance of Scots pine (Pinus sylvestris L.) half-sib families to Heterobasidion annosum. Forests 2019, 10, 287. [Google Scholar] [CrossRef]
- Sirgedaitė-Šėžienė, V.; Mildažienė, V.; Žemaitis, P.; Ivankov, A.; Koga, K.; Shiratani, M.; Baliuckas, V. Long-term response of Norway spruce to seed treatment with cold plasma: Dependence of the effects on the genotype. Plasma Process Polym. 2021, 18, 2000159. [Google Scholar] [CrossRef]
- Lučinskaitė, I.; Laužikė, K.; Žiauka, J.; Baliuckas, V.; Čėsna, V.; Sirgedaitė-Šėžienė, V. Assessment of biologically active compounds, organic acids and antioxidant activity in needle extracts of different Norway spruce (Picea abies (L.) H. Karst) half-sib families. Wood Sci. Technol. 2021, 55, 1221–1235. [Google Scholar] [CrossRef]
- Sarinont, T.; Amano, T.; Attri, P.; Koga, K.; Hayashi, N.; Shiratani, M. Effects of plasma irradiation using various feeding gases on growth of Raphanus sativus L. Arch. Biochem. Biophys. 2016, 60, 129–140. [Google Scholar] [CrossRef]
- Re, R.; Pallergini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 22, 1231–1237. [Google Scholar] [CrossRef]
- Ragaee, S.; Abdel-Aal, E.S.; Noaman, M. Antioxidant activity and nutrient composition of selected cereals for food use. Food Chem. 2006, 98, 32–38. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.; Chang, W.; Zhao, Z.; Cao, J. HPLC method for the simultaneous quantification of the major organic acids in Angeleno plum fruit. IOP Conf. Ser. Mater. Sci. Eng. 2014, 62, 012035. [Google Scholar] [CrossRef]
- Ramakrishna, A.; Ravishankar, G.A. Infuences of abiotic stress signals on secondary metabolites in plants. Plant Signal Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef]
- Rejeb, I.B.; Pastor, V.; Mauch-Mani, B. Plant responses to simultaneous biotic and abiotic stress: Molecular mechanisms. Plants 2017, 3, 458–475. [Google Scholar] [CrossRef]
- Carretto, S.; Linsalata, V.; Colella, G.; Mita, G.; Lattanzio, V. Carbon fuxes between primary metabolism and phenolic pathway in plant tissues under stress. Int. J. Mol. Sci. 2015, 16, 26378–26394. [Google Scholar] [CrossRef]
- Narayani, M.; Srivastava, S. Elicitation: A stimulation of stress in in vitro plant cell/tissue cultures for enhancement of secondary metabolite production. Phytochem Rev. 2018, 16, 1227–1252. [Google Scholar] [CrossRef]
- De Luca, V.; Salim, V.; Atsumi, S.M.; Yu, F. Mining the biodiversity of plants: A revolution in the making. Science 2012, 336, 1658–1661. [Google Scholar] [CrossRef]
- Wurtzel, E.T.; Kutchan, T.M. Plant metabolism, the diverse chemistry set of the future. Science 2016, 353, 1232–1236. [Google Scholar] [CrossRef]
- Soukupová, J.; Rock, B.; Albrechrova, J. Comparative study of two spruce species in a polluted mountainous region. New Phytol. 2002, 150, 133–145. [Google Scholar] [CrossRef]
- Raskin, I.; Ribnicky, D.M.; Komarnytsky, S.; Ilic, N.; Poulev, A.; Borisijuk, N.; Brinker, A.; Moreo, A.; Ripoll, C.; Yakoby, N.; et al. Plants and human health in the twenty-frst century. Trends Biotechnol. 2002, 20, 412–420. [Google Scholar] [CrossRef]
- Hall, R.D.; Brouwer, I.D.; Fitzgerald, M.A. Plant metabolomics and its potential application for human nutrition. Physiol. Plant. 2008, 132, 162–175. [Google Scholar] [CrossRef]
- Clerici, M.T.P.S.; Carvalho-Silva, L.B. Nutritional bioactive compounds and technological aspects of minor fruits grown in Brazil. Food Res. Int. 2011, 44, 1658–1670. [Google Scholar] [CrossRef]
- Cifuentes, A. Food analysis: Present, future, and foodomics. ISRN Anal. Chem. 2012, 2012, 801607. [Google Scholar] [CrossRef]
- Gismondi, A.; Di Marco, G.; Canuti, L.; Canini, A. Antiradical activity of phenolic metabolites extracted from grapes of white and red Vitis vinifera L. cultivars. Vitis 2017, 56, 19–26. [Google Scholar] [CrossRef]
- Rahman, M.M.; Rahaman, M.S.; Islam, M.R.; Rahman, F.; Mithi, F.M.; Alqahtani, T.; Almikhlafi, M.A.; Alghamdi, S.Q.; Alruwaili, A.S.; Hossain, M.S.; et al. Role of Phenolic Compounds in Human Disease: Current Knowledge and Future Prospects. Molecules 2022, 27, 233. [Google Scholar] [CrossRef] [PubMed]
- Carbonneau, M.A.; Leger, C.L.; Descomps, B.; Michel Fand Monnier, L. Improvement in the antioxidant status of plasma and low-density lipoprotein in subjects receiving a redwine phenolics mixture. JAOCS 1998, 75, 235–240. [Google Scholar] [CrossRef]
- Impei, S.; Gismondi, A.; Canuti, L.; Canini, A. Metabolic and biological profle of autochthonous Vitis vinifera L. ecotypes. Food Funct. 2015, 6, 1526–1538. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, A.C.; Kostoudi, C.; Sorotos, I.; Varsamis, G.; Korakis, G.; Drouzas, A.D. Diversity in needle morphology and genetic markers in a marginal Abies cephalonica (Pinaceae) population. Ann. For. Sci. 2015, 58, 217–234. [Google Scholar] [CrossRef]
- Karapandzova, M.; Stefkova, G.; Cvetkovikj, I.; Stanoevab, J.P.; Stefovab, M.; Kulevanova, S. Flavonoids and Other Phenolic Compounds in Needles of Pinus peuce and Other Pine Species from the Macedonian Flora. Nature 2015, 10, 987–990. [Google Scholar] [CrossRef]
- Ghaffari, Z.; Rahimmalek, M.; Sabzalian, M.R. Variation in the primary and secondary metabolites derived from the isoprenoid pathway in the Perovskia species in response to different wavelengths generated by light emitting diodes (LEDs). Ind. Crops. Prod. 2019, 140, 111592. [Google Scholar] [CrossRef]
- Karapandzova, M.; Stefkov, G.; Karanflova, I.; Penovska, T.; Stanoeva, J.; Stefova, M.; Kulenova, S. Chemical Characterization and Antioxidant Activity of Mountain Pine (Pinus mugo Turra, Pinaceae) from Republic of Macedonia. Rec. Ant. Prod. 2018, 13, 50–63. [Google Scholar] [CrossRef]
- Speisky, H.; Shahidi, F.; Costa de Camargo, A.; Fuentes, J. Revisiting the Oxidation of Flavonoids: Loss, Conservation or Enhancement of Their Antioxidant Properties. Antioxidants 2022, 11, 133. [Google Scholar] [CrossRef]
- Vauzour, D. Dietary polyphenols as modulators of brain functions: Biological actions and molecular mechanisms underpinning their beneficial effects. Oxid. Med. Cell. Longev. 2012, 212, 1–16. [Google Scholar] [CrossRef]
- Ribeiro, D.; Proenca, C.; Rocha, S.; Lima, J.L.; Carvalho, F.; Fernandes, E.; Freitas, M. Immunomodulatory effects of flavonoids in the prophylaxis and treatment of inflammatory bowel diseases: A comprehensive review. Curr. Med. Chem. 2018, 25, 3374–3412. [Google Scholar] [CrossRef]
- Mildaziene, V.; Pauzaite, G.; Nauciene, Z.; Malakauskiene, A.; Zukiene, R.; Januskaitiene, I.; Jakstas, V.; Ivanauskas, L.; Filatova, I.; Lyushkevich, V. Pre-sowing seed treatment with cold plasma and electromagnetic field increases secondary metabolite content in purple coneflower (Echinacea purpurea) leaves. Plasma Process 2017, 5, 1700059. [Google Scholar] [CrossRef]
- Prior, R.; Wu, X.; Schaich, K. Standardized Methods for the Determination of Antioxidant Capacity and Phenolics in Foods and Dietary Supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Measurement of antioxidant activity. J. Funct. Foods 2015, 18, 757–781. [Google Scholar] [CrossRef]
- Valu, M.V.; Ducu, C.; Moga, S.; Negrea, D.; Hritcu, L.; Boiangiu, R.S.; Vamanu, E.; Balseanu, T.A.; Carradori, S.; Soare, L.C. Effects of the Hydroethanolic Extract of Lycopodium selago L. on Scopolamine-Induced Memory Deficits in Zebrafish. Pharmaceuticals 2021, 14, 568. [Google Scholar] [CrossRef]
- Plank, D.W.; Szpylka, J.; Sapirstein, H.; Woollard, D.; Zapf, C.M.; Lee, V.; Chen, C.Y.; Liu., R.H.; Tsao, R.; Düsterloh, A.; et al. Determination of antioxidant activity in foods and beverages by reaction with 2,2′-diphenyl-1-picrylhydrazyl (DPPH): Collaborative study First Action 2012.04. J. AOAC Int. 2012, 95, 1562–1569. [Google Scholar] [CrossRef]
- French, D. Organic acid analysis. Adv. Clin. Chem. 2017, 79, 153–198. [Google Scholar] [CrossRef]
- Ivanova Petropulos, V.; Ricci, A.; Nedelkovski, D.; Dimovska, V.; Parpinello, G.P.; Versari, A. Targeted analysis of bioactive phenolic compounds and antioxidant activity of Macedonian red wines. Food Chem. 2015, 171, 412–420. [Google Scholar] [CrossRef]
- Brul, S.; Coote, P. Preservative agents in foods. Mode of action and microbial resistance mechanisms. Int. J. Food Microbiol. 1999, 50, 1–17. [Google Scholar] [CrossRef]
- Theron, M.M.; Lues, J.F.R. Organic acid and meat preservation: A review. Food Rev. Int. 2007, 23, 141–158. [Google Scholar] [CrossRef]
- Fiume, M.M.; Heldreth, B.A.; Bergfeld, W.F.; Belsito, D.V.; Hill, R.A.; Klaassen, C.D.; Liebler, D.C.; Marks, J.G.; Shank, R.C.; Slaga, T.J.; et al. Safety Assessment of Citric Acid, Inorganic Citrate Salts, and Alkyl Citrate Esters as Used in Cosmetics. Int. J. Toxicol. 2014, 33, 16–46. [Google Scholar] [CrossRef]
- Nghiem, N.P.; Kleff, S.; Schwegmann, S. Succinic Acid: Technology Development and Commercialization. Fermentation 2017, 3, 26. [Google Scholar] [CrossRef]
- Rodgers, A.L.; Webber, D.; de Charmoy, R.; Jackson, G.E.; Ravenscroft, N. Malic acid supplementation increases urinary citrate excretion and urinary pH: Implications for the potential treatment of calcium oxalate stone disease. J. Endourol. 2014, 28, 229–236. [Google Scholar] [CrossRef]
- Putchala, M.C.; Ramani, P.; Sherlin, H.J.; Premkumar, P.; Natesan, A. Ascorbic acid and its pro-oxidant activity as a therapy for tumours of oral cavity—A systematic review. Arch. Oral Biol. 2013, 58, 563–574. [Google Scholar] [CrossRef]
- Das, U.N. Folic acid says NO to vascular diseases. Nutrients 2003, 19, 686–692. [Google Scholar] [CrossRef]
- Ricke, S.C. Perspectives on the use of organic acids and short chain fatty acids as antimicrobials. Poult. Sci. 2003, 82, 632–639. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flavonoids and antioxidant activity | 7 half-sib families × 3 different treatment | 3 biological replicates | n = 63 |
| Organic acids | 7 half-sib families × 3 different treatment | 1 biological replicates | n = 21 |
| Trait | 1st Component | 2nd Component |
|---|---|---|
| Flavonoids | 14.63 | 16.92 |
| DPPH | 8.48 | 14.99 |
| ABTS | 14.00 | 19.09 |
| Oxalic | 0.07 | 0.31 |
| Malic | 21.90 | 6.19 |
| Ascorbic | 4.67 | 16.31 |
| Folic | 9.38 | 14.30 |
| Citric | 0.01 | 7.19 |
| Succinic | 26.86 | 4.70 |
| Trait | Compared Variants | t Value | p Value |
|---|---|---|---|
| Flavonoids | CP1 vs. control | 0.41 | 0.6802 |
| CP2 vs. control | −1.25 | 0.2150 | |
| CP1 vs. CP2 | 1.86 | 0.0654 | |
| DPPH | CP1 vs. control | 4.55 | <0.0001 |
| CP2 vs. control | −0.85 | 0.3959 | |
| CP1 vs. CP2 | 4.58 | <0.0001 | |
| ABTS | CP1 vs. control | −0.28 | 0.7824 |
| CP2 vs. control | −4.28 | <0.0001 | |
| CP1 vs. CP2 | 4.27 | <0.0001 | |
| Oxalic | CP1 vs. control | 2.17 | 0.0364 |
| CP2 vs. control | 0.87 | 0.3923 | |
| CP1 vs. CP2 | 0.84 | 0.4043 | |
| Malic | CP1 vs. control | 1.42 | 0.1644 |
| CP2 vs. control | −0.38 | 0.7025 | |
| CP1 vs. CP2 | 1.68 | 0.1037 | |
| Ascorbic | CP1 vs. control | 6.37 | <0.0001 |
| CP2 vs. control | 3.34 | 0.0026 | |
| CP1 vs. CP2 | 1.82 | 0.0766 | |
| Folic | CP1 vs. control | 0.71 | 0.4840 |
| CP2 vs. control | 0.81 | 0.4234 | |
| CP1 vs. CP2 | −0.04 | 0.9710 | |
| Citric | CP1 vs. control | 1.59 | 0.1200 |
| CP2 vs. control | 0.46 | 0.6476 | |
| CP1 vs. CP2 | 1.13 | 0.2636 | |
| Succinic | CP1 vs. control | 3.09 | 0.0052 |
| CP2 vs. control | 1.08 | 0.2873 | |
| CP1 vs. CP2 | 2.69 | 0.0131 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sirgedaitė-Šėžienė, V.; Lučinskaitė, I.; Mildažienė, V.; Ivankov, A.; Koga, K.; Shiratani, M.; Laužikė, K.; Baliuckas, V. Changes in Content of Bioactive Compounds and Antioxidant Activity Induced in Needles of Different Half-Sib Families of Norway Spruce (Picea abies (L.) H. Karst) by Seed Treatment with Cold Plasma. Antioxidants 2022, 11, 1558. https://doi.org/10.3390/antiox11081558
Sirgedaitė-Šėžienė V, Lučinskaitė I, Mildažienė V, Ivankov A, Koga K, Shiratani M, Laužikė K, Baliuckas V. Changes in Content of Bioactive Compounds and Antioxidant Activity Induced in Needles of Different Half-Sib Families of Norway Spruce (Picea abies (L.) H. Karst) by Seed Treatment with Cold Plasma. Antioxidants. 2022; 11(8):1558. https://doi.org/10.3390/antiox11081558
Chicago/Turabian StyleSirgedaitė-Šėžienė, Vaida, Ieva Lučinskaitė, Vida Mildažienė, Anatolii Ivankov, Kazunori Koga, Masaharu Shiratani, Kristina Laužikė, and Virgilijus Baliuckas. 2022. "Changes in Content of Bioactive Compounds and Antioxidant Activity Induced in Needles of Different Half-Sib Families of Norway Spruce (Picea abies (L.) H. Karst) by Seed Treatment with Cold Plasma" Antioxidants 11, no. 8: 1558. https://doi.org/10.3390/antiox11081558