
Role of Oxidative Stress and Antioxidants in Acquired Inner Ear Disorders



Abstract
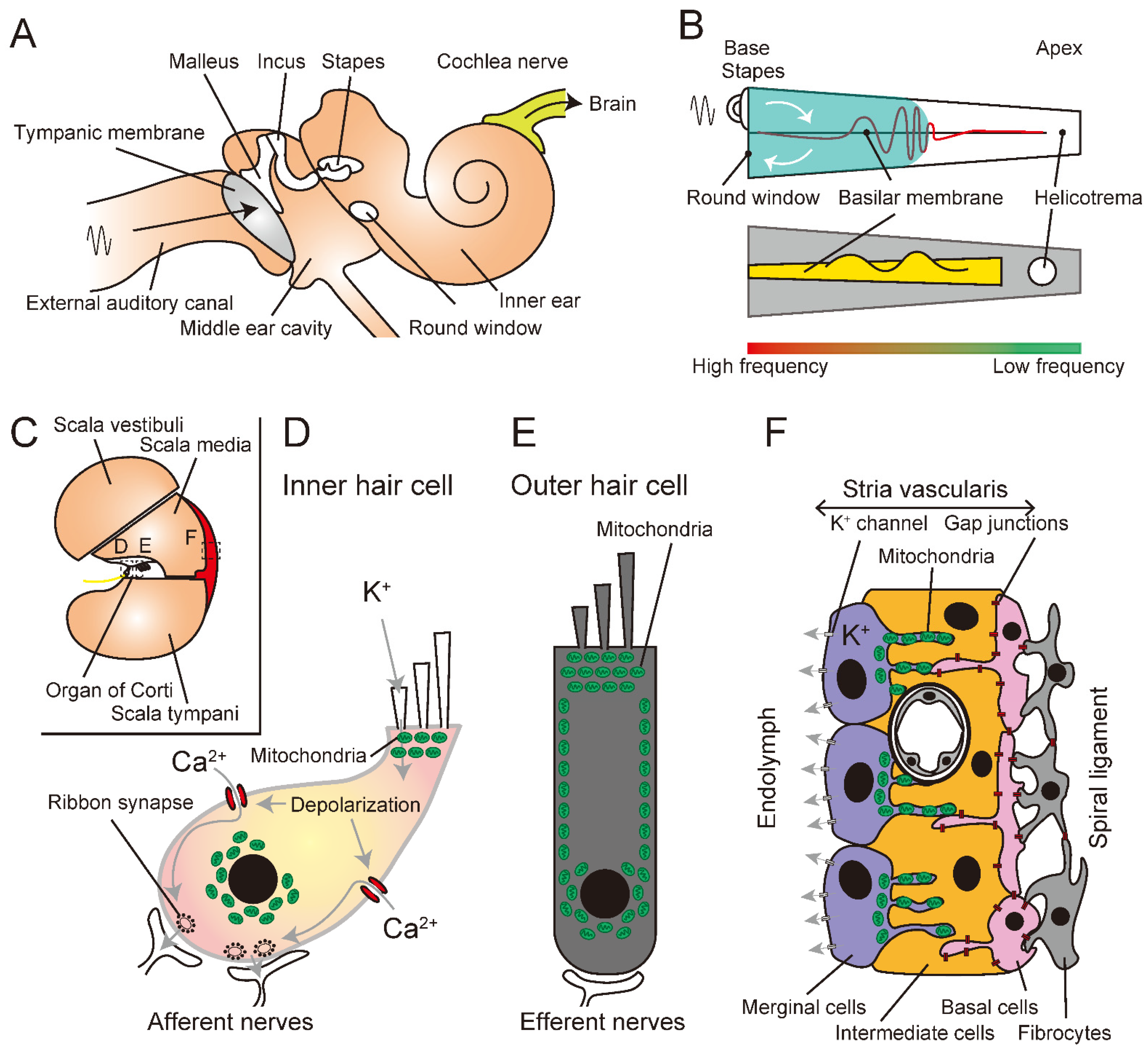
:1. Introduction
2. Auditory Processing
3. Sensorineural Hearing Loss
3.1. Drug-Induced Hearing Loss
3.2. Age-Related Hearing Loss
3.3. Noise-Induced Hearing Loss
4. Role of Oxidative Stress
4.1. Reactive Oxygen Species
4.2. Mitochondrial Function
5. Antioxidant Treatments
5.1. Intrinsic Antioxidants
5.2. Extrinsic Antioxidants
5.2.1. Water-Soluble Antioxidants
5.2.2. Lipid-Soluble Antioxidants
5.2.3. Amphoteric Antioxidants
5.3. Antioxidants in Acquired Inner Ear Disorders
5.3.1. Medicines
Drug-Induced Hearing Loss
Age-Related Hearing Loss
Noise-Induced Hearing Loss
5.3.2. Vitamins
Neurodegenerative Disorders
Inner Ear Disorder
5.3.3. Natural Foods
Green Tea
Brown Seaweeds
5.3.4. New Antioxidants
5.3.5. Clinical Studies
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Beckhauser, T.F.; Francis-Oliveira, J.; De Pasquale, R. Reactive Oxygen Species: Physiological and Physiopathological Effects on Synaptic Plasticity. J. Exp. Neurosci. 2016, 10, 23–48. [Google Scholar] [CrossRef] [Green Version]
- Beltrán González, A.N.; López Pazos, M.I.; Calvo, D.J. Reactive Oxygen Species in the Regulation of the GABA Mediated Inhibitory Neurotransmission. Neuroscience 2020, 439, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Sena, L.A.; Chandel, N.S. Physiological roles of mitochondrial reactive oxygen species. Mol. Cell 2012, 48, 158–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-mediated cellular signaling. Oxid. Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef] [Green Version]
- Marchi, S.; Giorgi, C.; Suski, J.M.; Agnoletto, C.; Bononi, A.; Bonora, M.; De Marchi, E.; Missiroli, S.; Patergnani, S.; Poletti, F. Mitochondria-ros crosstalk in the control of cell death and aging. J. Signal Transduct. 2012, 2012, 329635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, H.-U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Chompoosor, A.; Saha, K.; Ghosh, P.S.; Macarthy, D.J.; Miranda, O.R.; Zhu, Z.; Arcaro, K.F.; Rotello, V.M. The role of surface functionality on acute cytotoxicity, ROS generation and DNA damage by cationic gold nanoparticles. Small 2010, 6, 2246–2249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalska, P.; León, R. When it comes to an end: Oxidative stress crosstalk with protein aggregation and neuroinflammation induce neurodegeneration. Antioxidants 2020, 9, 740. [Google Scholar] [CrossRef]
- Wu, J.; Ye, J.; Kong, W.; Zhang, S.; Zheng, Y. Programmed cell death pathways in hearing loss: A review of apoptosis, autophagy and programmed necrosis. Cell Prolif. 2020, 53, e12915. [Google Scholar] [CrossRef] [PubMed]
- Fettiplace, R. Diverse Mechanisms of Sound Frequency Discrimination in the Vertebrate Cochlea. Trends Neurosci. 2020, 43, 88–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Békésy, G.; Peake, W.T. Experiments in hearing. J. Acoust. Soc. Am. 1990, 88, 2905. [Google Scholar] [CrossRef]
- Hudspeth, A.J. Integrating the active process of hair cells with cochlear function. Nat. Rev. Neurosci. 2014, 15, 600–614. [Google Scholar] [CrossRef] [PubMed]
- Davis, H. Transmission and transduction in the cochlea. Laryngoscope 1958, 68, 359–382. [Google Scholar] [CrossRef]
- Ren, T. Longitudinal pattern of basilar membrane vibration in the sensitive cochlea. Proc. Natl. Acad. Sci. USA 2002, 99, 17101–17106. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.Y.; Raphael, P.D.; Park, J.; Ellerbee, A.K.; Applegate, B.E.; Oghalai, J.S. Noninvasive in vivo imaging reveals differences between tectorial membrane and basilar membrane traveling waves in the mouse cochlea. Proc. Natl. Acad. Sci. USA 2015, 112, 3128–3133. [Google Scholar] [CrossRef] [Green Version]
- Tsukano, H.; Horie, M.; Ohga, S.; Takahashi, K.; Kubota, Y.; Hishida, R.; Takebayashi, H.; Shibuki, K. Reconsidering Tonotopic Maps in the Auditory Cortex and Lemniscal Auditory Thalamus in Mice. Front. Neural Circuits 2017, 11, 14. [Google Scholar] [PubMed] [Green Version]
- Xiong, W.; Grillet, N.; Elledge, H.M.; Wagner, T.F.J.; Zhao, B.; Johnson, K.R.; Kazmierczak, P.; Müller, U. TMHS Is an Integral Component of the Mechanotransduction Machinery of Cochlear Hair Cells. Cell 2012, 151, 1283–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.C.; Lin, C.-C.; Cheung, R.; Zhang-Hooks, Y.; Agarwal, A.; Ellis-Davies, G.; Rock, J.; Bergles, D.E. Spontaneous Activity of Cochlear Hair Cells Triggered by Fluid Secretion Mechanism in Adjacent Support Cells. Cell 2015, 163, 1348–1359. [Google Scholar] [CrossRef] [Green Version]
- Ashmore, J. Cochlear outer hair cell motility. Physiol. Rev. 2008, 88, 173–210. [Google Scholar] [CrossRef] [Green Version]
- Blagosklonny, M. V Aging: Ros or tor. Cell Cycle 2008, 7, 3344–3354. [Google Scholar] [CrossRef]
- Huth, M.E.; Ricci, A.J.; Cheng, A. Mechanisms of aminoglycoside ototoxicity and targets of hair cell protection. Int. J. Otolaryngol. 2011, 2011, 937861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroese, A.B.A.; van den Bercken, J. Dual action of ototoxic antibiotics on sensory hair cells. Nature 1980, 283, 395–397. [Google Scholar] [CrossRef] [PubMed]
- Schacht, J.; Talaska, A.E.; Rybak, L.P. Cisplatin and aminoglycoside antibiotics: Hearing loss and its prevention. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2012, 295, 1837–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffield, J.S.; Bonventre, J. V Kidney tubular epithelium is restored without replacement with bone marrow–derived cells during repair after ischemic injury. Kidney Int. 2005, 68, 1956–1961. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.G.; Cunningham, L.L.; Rubel, E.W. Hair cell death in the avian basilar papilla: Characterization of the in vitro model and caspase activation. J. Assoc. Res. Otolaryngol. 2003, 4, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Jensen-Smith, H.C.; Hallworth, R.; Nichols, M.G. Gentamicin rapidly inhibits mitochondrial metabolism in high-frequency cochlear outer hair cells. PLoS ONE 2012, 7, e38471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehne, N.; Rauen, U.; de Groot, H.; Lautermann, J. Involvement of the mitochondrial permeability transition in gentamicin ototoxicity. Hear. Res. 2002, 169, 47–55. [Google Scholar] [CrossRef]
- Suzuki, M.; Ushio, M.; Yamasoba, T. Time course of apoptotic cell death in guinea pig cochlea following intratympanic gentamicin application. Acta Otolaryngol. 2008, 128, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Marcotti, W.; Van Netten, S.M.; Kros, C.J. The aminoglycoside antibiotic dihydrostreptomycin rapidly enters mouse outer hair cells through the mechano-electrical transducer channels. J. Physiol. 2005, 567, 505–521. [Google Scholar] [CrossRef] [PubMed]
- Abi-Hachem, R.N.; Zine, A.; Van De Water, T.R. The injured cochlea as a target for inflammatory processes, initiation of cell death pathways and application of related otoprotective strategies. Recent Patents CNS Drug Discov. 2010, 5, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Hobbie, S.N.; Akshay, S.; Kalapala, S.K.; Bruell, C.M.; Shcherbakov, D.; Böttger, E.C. Genetic analysis of interactions with eukaryotic rRNA identify the mitoribosome as target in aminoglycoside ototoxicity. Proc. Natl. Acad. Sci. USA 2008, 105, 20888–20893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halestrap, A.P.; Clarke, S.J.; Javadov, S.A. Mitochondrial permeability transition pore opening during myocardial reperfusion—a target for cardioprotection. Cardiovasc. Res. 2004, 61, 372–385. [Google Scholar] [CrossRef] [Green Version]
- Halestrap, A.P. What is the mitochondrial permeability transition pore? J. Mol. Cell. Cardiol. 2009, 46, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Tono, T.; Kiyomizu, K.; Matsuda, K.; Komune, S.; Usami, S.; Abe, S.; Shinkawa, H. Different clinical characteristics of aminoglycoside-induced profound deafness with and without the 1555 A-->G mitochondrial mutation. ORL J. Otorhinolaryngol. Relat. Spec. 2001, 63, 25–30. [Google Scholar] [CrossRef]
- Dowlati, M.A.; Derakhshandeh-Peykar, P.; Houshmand, M.; Farhadi, M.; Shojaei, A.; Fallah, M.; Mohammadi, E.; Tajdini, A.; Arastoo, S.; Tavakkoly-Bazzaz, J. Novel nucleotide changes in mutational analysis of mitochondrial 12SrRNA gene in patients with nonsyndromic and aminoglycoside-induced hearing loss. Mol. Biol. Rep. 2013, 40, 2689–2695. [Google Scholar] [CrossRef]
- Böttger, E.C. Mutant A1555G mitochondrial 12S rRNA and aminoglycoside susceptibility. Antimicrob. Agents Chemother. 2010, 54, 3073–3075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vokes, E.E. Induction chemotherapy for head and neck cancer: Recent data. Oncologist 2010, 15 (Suppl. 3), 3–7. [Google Scholar] [CrossRef] [PubMed]
- Damsma, G.E.; Alt, A.; Brueckner, F.; Carell, T.; Cramer, P. Mechanism of transcriptional stalling at cisplatin-damaged DNA. Nat. Struct. Mol. Biol. 2007, 14, 1127–1133. [Google Scholar] [CrossRef]
- Chu, G. Cellular responses to cisplatin. The roles of DNA-binding proteins and DNA repair. J. Biol. Chem. 1994, 269, 787–790. [Google Scholar] [CrossRef]
- Rybak, L.P.; Ramkumar, V. Ototoxicity. Kidney Int. 2007, 72, 931–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rybak, L.P.; Whitworth, C.A.; Mukherjea, D.; Ramkumar, V. Mechanisms of cisplatin-induced ototoxicity and prevention. Hear. Res. 2007, 226, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Kimitsuki, T.; Nakagawa, T.; Hisashi, K.; Komune, S.; Komiyama, S. Cisplatin blocks mechano-electric transducer current in chick cochlear hair cells. Hear. Res. 1993, 71, 64–68. [Google Scholar] [CrossRef]
- García-Berrocal, J.R.; Nevado, J.; Ramírez-Camacho, R.; Sanz, R.; González-García, J.A.; Sánchez-Rodríguez, C.; Cantos, B.; España, P.; Verdaguer, J.M.; Trinidad Cabezas, A. The anticancer drug cisplatin induces an intrinsic apoptotic pathway inside the inner ear. Br. J. Pharmacol. 2007, 152, 1012–1020. [Google Scholar] [CrossRef] [PubMed]
- Kaltenbach, J.A.; Rachel, J.D.; Mathog, T.A.; Zhang, J.; Falzarano, P.R.; Lewandowski, M. Cisplatin-induced hyperactivity in the dorsal cochlear nucleus and its relation to outer hair cell loss: Relevance to tinnitus. J. Neurophysiol. 2002, 88, 699–714. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.A.; Ikeda, K.; Oshima, T.; Suzuki, M.; Kawase, T.; Kikuchi, T.; Takasaka, T. Cisplatin-induced apoptotic cell death in Mongolian gerbil cochlea. Hear. Res. 2000, 141, 28–38. [Google Scholar] [CrossRef]
- Lee, J.E.; Nakagawa, T.; Kita, T.; Kim, T.S.; Iguchi, F.; Endo, T.; Shiga, A.; Lee, S.H.; Ito, J. Mechanisms of apoptosis induced by cisplatin in marginal cells in mouse stria vascularis. ORL J. Otorhinolaryngol. Relat. Spec. 2004, 66, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Meech, R.P.; Campbell, K.C.M.; Hughes, L.P.; Rybak, L.P. A semiquantitative analysis of the effects of cisplatin on the rat stria vascularis. Hear. Res. 1998, 124, 44–59. [Google Scholar] [CrossRef]
- Lee, J.E.; Nakagawa, T.; Kim, T.S.; Iguchi, F.; Endo, T.; Dong, Y.; Yuki, K.; Naito, Y.; Lee, S.H.; Ito, J. A novel model for rapid induction of apoptosis in spiral ganglions of mice. Laryngoscope 2003, 113, 994–999. [Google Scholar] [CrossRef] [PubMed]
- Bowers, W.J.; Chen, X.; Guo, H.; Frisina, D.R.; Federoff, H.J.; Frisina, R.D. Neurotrophin-3 transduction attenuates cisplatin spiral ganglion neuron ototoxicity in the cochlea. Mol. Ther. 2002, 6, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Anniko, M.; Sobin, A. Cisplatin: Evaluation of its ototoxic potential. Am. J. Otolaryngol. 1986, 7, 276–293. [Google Scholar] [CrossRef]
- Bánfi, B.; Malgrange, B.; Knisz, J.; Steger, K.; Dubois-Dauphin, M.; Krause, K.-H. NOX3, a superoxide-generating NADPH oxidase of the inner ear. J. Biol. Chem. 2004, 279, 46065–46072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohri, H.; Ninoyu, Y.; Sakaguchi, H.; Hirano, S.; Saito, N.; Ueyama, T. Nox3-derived superoxide in cochleae induces sensorineural hearing loss. J. Neurosci. 2021, 41, 4716–4731. [Google Scholar] [CrossRef]
- Ikeda, K.; Sunose, H.; Takasaka, T. Effects of free radicals on the intracellular calcium concentration in the isolated outer hair cell of the guinea pig cochlea. Acta Otolaryngol. 1993, 113, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Kopke, R.D.; Liu, W.; Gabaizadeh, R.; Jacono, A.; Feghali, J. Use of Organotypic Cultures of Corti’s Organ to. Ann. J. Otol. 1997, 18, 559–571. [Google Scholar]
- Casares, C.; Ramírez-Camacho, R.; Trinidad, A.; Roldán, A.; Jorge, E.; García-Berrocal, J.R. Reactive oxygen species in apoptosis induced by cisplatin: Review of physiopathological mechanisms in animal models. Eur. Arch. Oto-Rhino-Laryngol. 2012, 269, 2455–2459. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Inai, S.; Jinnouchi, K.; Baba, S.; Yagi, T. Expression of caspase-activated deoxyribonuclease (CAD) and caspase 3 (CPP32) in the cochlea of cisplatin (CDDP)-treated guinea pigs. Auris. Nasus. Larynx 2003, 30, 219–225. [Google Scholar] [CrossRef]
- Sheth, S.; Mukherjea, D.; Rybak, L.P.; Ramkumar, V. Mechanisms of cisplatin-induced ototoxicity and otoprotection. Front. Cell. Neurosci. 2017, 11, 338. [Google Scholar] [CrossRef] [PubMed]
- Rybak, L.P.; Mukherjea, D.; Jajoo, S.; Ramkumar, V. Cisplatin ototoxicity and protection: Clinical and experimental studies. Tohoku J. Exp. Med. 2009, 219, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Yamasoba, T.; Lin, F.R.; Someya, S.; Kashio, A.; Sakamoto, T.; Kondo, K. Current concepts in age-related hearing loss: Epidemiology and mechanistic pathways. Hear. Res. 2013, 303, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Cruickshanks, K.J.; Zhan, W.; Zhong, W. Epidemiology of age-related hearing impairment. In The Aging Auditory System; Springer: Berlin/Heidelberg, Germany, 2010; pp. 259–274. [Google Scholar]
- Lee, K.-Y. Pathophysiology of age-related hearing loss (peripheral and central). Korean J. Audiol. 2013, 17, 45. [Google Scholar] [CrossRef] [Green Version]
- Gates, G.A.; Mills, J.H. Presbycusis. Lancet 2005, 366, 1111–1120. [Google Scholar] [CrossRef]
- Schuknecht, H.F. Presbycusis. Laryngoscope 1955, 65, 402–419. [Google Scholar] [CrossRef] [PubMed]
- Schuknecht, H.F.; Gacek, M.R. Cochlear pathology in presbycusis. Ann. Otol. Rhinol. Laryngol. 1993, 102, 1–16. [Google Scholar] [CrossRef]
- Grivell, L.A. Mitochondrial DNA. Small, beautiful and essential. Nature 1989, 341, 569–571. [Google Scholar] [CrossRef]
- Linnane, A.W.; Marzuki, S.; Ozawa, T.; Tanaka, M. Mitochondrial DNA mutations as an important contributor to ageing and degenerative diseases. Lancet 1989, 1, 642–645. [Google Scholar] [CrossRef]
- Anderson, S.; Bankier, A.T.; Barrell, B.G.; de Bruijn, M.H.L.; Coulson, A.R.; Drouin, J.; Eperon, I.C.; Nierlich, D.P.; Roe, B.A.; Sanger, F. Sequence and organization of the human mitochondrial genome. Nature 1981, 290, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Clayton, D.A.; Doda, J.N.; Friedberg, E.C. The absence of a pyrimidine dimer repair mechanism in mammalian mitochondria. Proc. Natl. Acad. Sci. USA 1974, 71, 2777–2781. [Google Scholar] [CrossRef] [Green Version]
- Bai, U.; Seidman, M.D.; Hinojosa, R.; Quirk, W.S. Mitochondrial DNA deletions associated with aging and possibly presbycusis: A human archival temporal bone study. Am. J. Otol. 1997, 18, 449–453. [Google Scholar] [PubMed]
- Ozawa, T. Mechanism of somatic mitochondrial DNA mutations associated with age and diseases. Biochim. Biophys. Acta (BBA)-Molecular Basis Dis. 1995, 1271, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Menardo, J.; Tang, Y.; Ladrech, S.; Lenoir, M.; Casas, F.; Michel, C.; Bourien, J.; Ruel, J.; Rebillard, G.; Maurice, T.; et al. Oxidative stress, inflammation, and autophagic stress as the key mechanisms of premature age-related hearing loss in SAMP8 mouse Cochlea. Antioxid. Redox Signal. 2012, 16, 263–274. [Google Scholar] [CrossRef] [PubMed]
- McFadden, S.L.; Ding, D.; Reaume, A.G.; Flood, D.G.; Salvi, R.J. Age-related cochlear hair cell loss is enhanced in mice lacking copper/zinc superoxide dismutase. Neurobiol. Aging 1999, 20, 1–8. [Google Scholar] [CrossRef]
- Darrat, I.; Ahmad, N.; Seidman, K.; Seidman, M.D. Auditory research involving antioxidants. Curr. Opin. Otolaryngol. Head Neck Surg. 2007, 15, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Seidman, M.D. Effects of dietary restriction and antioxidants on presbyacusis. Laryngoscope 2000, 110, 727–738. [Google Scholar] [CrossRef] [Green Version]
- Heman-Ackah, S.E.; Juhn, S.K.; Huang, T.C.; Wiedmann, T.S. A combination antioxidant therapy prevents age-related hearing loss in C57BL/6 mice. Otolaryngol. neck Surg. Off. J. Am. Acad. Otolaryngol. Neck Surg. 2010, 143, 429–434. [Google Scholar] [CrossRef]
- Someya, S.; Xu, J.; Kondo, K.; Ding, D.; Salvi, R.J.; Yamasoba, T.; Rabinovitch, P.S.; Weindruch, R.; Leeuwenburgh, C.; Tanokura, M.; et al. Age-related hearing loss in C57BL/6J mice is mediated by Bak-dependent mitochondrial apoptosis. Proc. Natl. Acad. Sci. USA 2009, 106, 19432–19437. [Google Scholar] [CrossRef] [Green Version]
- Sha, S.-H.; Kanicki, A.; Halsey, K.; Wearne, K.A.; Schacht, J. Antioxidant-enriched diet does not delay the progression of age-related hearing loss. Neurobiol. Aging 2012, 33, 1010.e15–1010.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, G.M. Caspases: The executioners of apoptosis. Biochem. J. 1997, 326, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsten, T.; Ross, A.J.; King, A.; Zong, W.-X.; Rathmell, J.C.; Shiels, H.A.; Ulrich, E.; Waymire, K.G.; Mahar, P.; Frauwirth, K. The combined functions of proapoptotic Bcl-2 family members bak and bax are essential for normal development of multiple tissues. Mol. Cell 2000, 6, 1389–1399. [Google Scholar] [CrossRef] [Green Version]
- Youle, R.J.; Strasser, A. The BCL-2 protein family: Opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Spoendlin, H. Primary structural changes in the organ of Corti after acoustic overstimulation. Acta Otolaryngol. 1971, 71, 166–176. [Google Scholar] [CrossRef]
- Mulroy, M.J.; Henry, W.R.; McNeil, P.L. Noise-induced transient microlesions in the cell membranes of auditory hair cells. Hear. Res. 1998, 115, 93–100. [Google Scholar] [CrossRef]
- Slepecky, N. Overview of mechanical damage to the inner ear: Noise as a tool to probe cochlear function. Hear. Res. 1986, 22, 307–321. [Google Scholar] [CrossRef]
- Lim, D.J.; Melnick, W. Acoustic damage of the cochlea: A scanning and transmission electron microscopic observation. Arch. Otolaryngol. 1971, 94, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Yamane, H.; Nakai, Y.; Takayama, M.; Iguchi, H.; Nakagawa, T.; Kojima, A. Appearance of free radicals in the guinea pig inner ear after noise-induced acoustic trauma. Eur. Arch. Oto-Rhino-Laryngol. 1995, 252, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Hong, O.; Kerr, M.J.; Poling, G.L.; Dhar, S. Understanding and preventing noise-induced hearing loss. Dis Mon 2013, 59, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Henderson, D.; Bielefeld, E.C.; Harris, K.C.; Hu, B.H. The role of oxidative stress in noise-induced hearing loss. Ear Hear. 2006, 27, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.H.; Cai, Q.; Manohar, S.; Jiang, H.; Ding, D.; Coling, D.E.; Zheng, G.; Salvi, R. Differential expression of apoptosis-related genes in the cochlea of noise-exposed rats. Neuroscience 2009, 161, 915–925. [Google Scholar] [CrossRef] [Green Version]
- Nicotera, T.M.; Hu, B.H.; Henderson, D. The caspase pathway in noise-induced apoptosis of the chinchilla cochlea. J. Assoc. Res. Otolaryngol. 2003, 4, 466–477. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, D.; Miller, J.M.; Jiang, H.-Y.; Minami, S.B.; Schacht, J. AIF and EndoG in noise-induced hearing loss. Neuroreport 2004, 15, 2719–2722. [Google Scholar] [PubMed]
- Ohlemiller, K.K.; Wright, J.S.; Dugan, L.L. Early elevation of cochlear reactive oxygen species following noise exposure. Audiol. Neurootol. 1999, 4, 229–236. [Google Scholar] [CrossRef]
- Ohinata, Y.; Miller, J.M.; Altschuler, R.A.; Schacht, J. Intense noise induces formation of vasoactive lipid peroxidation products in the cochlea. Brain Res. 2000, 878, 163–173. [Google Scholar] [CrossRef]
- Pirvola, U.; Xing-Qun, L.; Virkkala, J.; Saarma, M.; Murakata, C.; Camoratto, A.M.; Walton, K.M.; Ylikoski, J. Rescue of hearing, auditory hair cells, and neurons by CEP-1347/KT7515, an inhibitor of c-Jun N-terminal kinase activation. J. Neurosci. 2000, 20, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perlman, H.B.; Kimura, R. Cochlear blood flow in acoustic trauma. Acta Otolaryngol. 1962, 54, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Hawkins Jr, J.E. The role of vasoconstriction in noise-induced hearing loss. Ann. Otol. Rhinol. Laryngol. 1971, 80, 903–913. [Google Scholar] [CrossRef] [PubMed]
- Puel, J.L.; Ruel, J.; Gervais d’Aldin, C.; Pujol, R. Excitotoxicity and repair of cochlear synapses after noise-trauma induced hearing loss. Neuroreport 1998, 9, 2109–2114. [Google Scholar] [CrossRef] [PubMed]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Balaban, R.S.; Nemoto, S.; Finkel, T. Mitochondria, oxidants, and aging. Cell 2005, 120, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, T.; Wood, R.D. Quality control by DNA repair. Science 1999, 286, 1897–1905. [Google Scholar] [CrossRef] [PubMed]
- Hemnani, T.; Parihar, M.S. Reactive oxygen species and oxidative DNA damage. Indian J. Physiol. Pharmacol. 1998, 42, 440–452. [Google Scholar]
- Martinet, W.; Knaapen, M.W.M.; De Meyer, G.R.Y.; Herman, A.G.; Kockx, M.M. Elevated levels of oxidative DNA damage and DNA repair enzymes in human atherosclerotic plaques. Circulation 2002, 106, 927–932. [Google Scholar] [CrossRef] [Green Version]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997, 82, 291–295. [Google Scholar] [CrossRef]
- Vertuani, S.; Angusti, A.; Manfredini, S. The Antioxidants and Pro-Antioxidants Network: An Overview. Curr. Pharm. Des. 2004, 10, 1677–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orr, W.C.; Sohal, R.S. Extension of life-span by overexpression of superoxide dismutase and catalase in Drosophila melanogaster. Science 1994, 263, 1128–1130. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.; Shi, H.; Zhang, C.; Ren, M.; Han, M.; Wei, X.; Zhang, X.; Lou, H. Dimethyl fumarate attenuates 6-OHDA-induced neurotoxicity in SH-SY5Y cells and in animal model of Parkinson’s disease by enhancing Nrf2 activity. Neuroscience 2015, 286, 131–140. [Google Scholar] [CrossRef]
- Sano, M.; Ernesto, C.; Thomas, R.G.; Klauber, M.R.; Schafer, K.; Grundman, M.; Woodbury, P.; Growdon, J.; Cotman, C.W.; Pfeiffer, E. A controlled trial of selegiline, alpha-tocopherol, or both as treatment for Alzheimer’s disease. N. Engl. J. Med. 1997, 336, 1216–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelhart, M.J.; Geerlings, M.I.; Ruitenberg, A.; van Swieten, J.C.; Hofman, A.; Witteman, J.C.M.; Breteler, M.M.B. Dietary intake of antioxidants and risk of Alzheimer disease. JAMA 2002, 287, 3223–3229. [Google Scholar] [CrossRef] [Green Version]
- Pelicano, H.; Carney, D.; Huang, P. ROS stress in cancer cells and therapeutic implications. Drug Resist. Updat. 2004, 7, 97–110. [Google Scholar] [CrossRef]
- Moloney, J.N.; Cotter, T.G. ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef]
- Wu, D.; Cederbaum, A.I. Alcohol, oxidative stress, and free radical damage. Alcohol Res. Health 2003, 27, 277. [Google Scholar] [PubMed]
- Radak, Z.; Zhao, Z.; Koltai, E.; Ohno, H.; Atalay, M. Oxygen consumption and usage during physical exercise: The balance between oxidative stress and ROS-dependent adaptive signaling. Antioxid. Redox Signal. 2013, 18, 1208–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davalli, P.; Mitic, T.; Caporali, A.; Lauriola, A.; D’Arca, D. ROS, cell senescence, and novel molecular mechanisms in aging and age-related diseases. Oxid. Med. Cell. Longev. 2016, 2016, 3565127. [Google Scholar] [CrossRef] [Green Version]
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. Am. J. Physiol. Cell. Mol. Physiol. 2000, 279, L1005–L1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volpe, C.M.O.; Villar-Delfino, P.H.; Dos Anjos, P.M.F.; Nogueira-Machado, J.A. Cellular death, reactive oxygen species (ROS) and diabetic complications. Cell Death Dis. 2018, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Su, L.-J.; Zhang, J.-H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.-Y. Reactive oxygen species-induced lipid peroxidation in apoptosis, autophagy, and ferroptosis. Oxid. Med. Cell. Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef] [Green Version]
- Sinha, K.; Das, J.; Pal, P.B.; Sil, P.C. Oxidative stress: The mitochondria-dependent and mitochondria-independent pathways of apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Noguchi, T.; Homma, K.; Katagiri, K.; Takeda, K.; Matsuzawa, A.; Ichijo, H. Ubiquitin-like Sequence in ASK1 Plays Critical Roles in the Recognition and Stabilization by USP9X and Oxidative Stress-Induced Cell Death. Mol. Cell 2009, 36, 805–818. [Google Scholar] [CrossRef]
- Schon, E.A.; Bonilla, E.; DiMauro, S. Mitochondrial DNA mutations and pathogenesis. J. Bioenerg. Biomembr. 1997, 29, 131–149. [Google Scholar] [CrossRef]
- Chomyn, A. The myoclonic epilepsy and ragged-red fiber mutation provides new insights into human mitochondrial function and genetics. Am. J. Hum. Genet. 1998, 62, 745–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sue, C.M.; Lipsett, L.J.; Crimmins, D.S.; Tsang, C.S.; Boyages, S.C.; Presgrave, C.M.; Gibson, W.P.; Byrne, E.; Morris, J.G. Cochlear origin of hearing loss in MELAS syndrome. Ann. Neurol. 1998, 43, 350–359. [Google Scholar] [CrossRef]
- Kokotas, H.; Petersen, M.B.; Willems, P.J. Mitochondrial deafness. Clin. Genet. 2007, 71, 379–391. [Google Scholar] [CrossRef]
- Carroll, C.J.; Brilhante, V.; Suomalainen, A. Next-generation sequencing for mitochondrial disorders. Br. J. Pharmacol. 2014, 171, 1837–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtake, A.; Murayama, K.; Mori, M.; Harashima, H.; Yamazaki, T.; Tamaru, S.; Yamashita, Y.; Kishita, Y.; Nakachi, Y.; Kohda, M.; et al. Diagnosis and molecular basis of mitochondrial respiratory chain disorders: Exome sequencing for disease gene identification. Biochim. Biophys. Acta 2014, 1840, 1355–1359. [Google Scholar] [CrossRef] [Green Version]
- Tsutomu, S.; Asuteka, N.; Takeo, S. Human mitochondrial diseases caused by lack of taurine modification in mitochondrial tRNAs. Wiley Interdiscip. Rev. RNA 2011, 2, 376–386. [Google Scholar] [CrossRef]
- Kirino, Y.; Yasukawa, T.; Ohta, S.; Akira, S.; Ishihara, K.; Watanabe, K.; Suzuki, T. Codon-specific translational defect caused by a wobble modification deficiency in mutant tRNA from a human mitochondrial disease. Proc. Natl. Acad. Sci. USA 2004, 101, 15070–15075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morscher, R.J.; Ducker, G.S.; Li, S.H.-J.; Mayer, J.A.; Gitai, Z.; Sperl, W.; Rabinowitz, J.D. Mitochondrial translation requires folate-dependent tRNA methylation. Nature 2018, 554, 128–132. [Google Scholar] [CrossRef]
- Umeda, N.; Suzuki, T.; Yukawa, M.; Ohya, Y.; Shindo, H.; Watanabe, K.; Suzuki, T. Mitochondria-specific RNA-modifying enzymes responsible for the biosynthesis of the wobble base in mitochondrial tRNAs: Implications for the molecular pathogenesis of human mitochondrial diseases. J. Biol. Chem. 2005, 280, 1613–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Guan, M.-X. A human mitochondrial GTP binding protein related to tRNA modification may modulate phenotypic expression of the deafness-associated mitochondrial 12S rRNA mutation. Mol. Cell. Biol. 2002, 22, 7701–7711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villarroya, M.; Prado, S.; Esteve, J.M.; Soriano, M.A.; Aguado, C.; Pérez-Martínez, D.; Martínez-Ferrandis, J.I.; Yim, L.; Victor, V.M.; Cebolla, E. Characterization of human GTPBP3, a GTP-binding protein involved in mitochondrial tRNA modification. Mol. Cell. Biol. 2008, 28, 7514–7531. [Google Scholar] [CrossRef] [Green Version]
- Bykhovskaya, Y.; Mengesha, E.; Wang, D.; Yang, H.; Estivill, X.; Shohat, M.; Fischel-Ghodsian, N. Phenotype of non-syndromic deafness associated with the mitochondrial A1555G mutation is modulated by mitochondrial RNA modifying enzymes MTO1 and GTPBP3. Mol. Genet. Metab. 2004, 83, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Takai, D.; Soejima, A.; Isobe, K.; Yamasoba, T.; Oka, Y.; Goto, Y.; Hayashi, J. Mutant mtDNA at 1555 A to G in 12S rRNA gene and hypersusceptibility of mitochondrial translation to streptomycin can be co-transferred to rho 0 HeLa cells. Biochem. Biophys. Res. Commun. 1996, 223, 496–501. [Google Scholar] [CrossRef]
- Yamasoba, T.; Goto, Y.I.; Oka, Y.; Nishino, I.; Tsukuda, K.; Nonaka, I. Atypical muscle pathology and a survey of cis-mutations in deaf patients harboring a 1555 A-to-G point mutation in the mitochondrial ribosomal RNA gene. Neuromuscul. Disord. 2002, 12, 506–512. [Google Scholar] [CrossRef]
- Ghezzi, D.; Baruffini, E.; Haack, T.B.; Invernizzi, F.; Melchionda, L.; Dallabona, C.; Strom, T.M.; Parini, R.; Burlina, A.B.; Meitinger, T. Mutations of the mitochondrial-tRNA modifier MTO1 cause hypertrophic cardiomyopathy and lactic acidosis. Am. J. Hum. Genet. 2012, 90, 1079–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata, H.; Kumahara, K.; Tomemori, T.; Arimoto, Y.; Isoyama, K.; Yoshida, K.; Konno, A. Frequency and clinical features of patients with sensorineural hearing loss associated with the A3243G mutation of the mitochondrial DNA in otorhinolaryngic clinics. J. Hum. Genet. 2001, 46, 595–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadowaki, T.; Kadowaki, H.; Mori, Y.; Tobe, K.; Sakuta, R.; Suzuki, Y.; Tanabe, Y.; Sakura, H.; Awata, T.; Goto, Y. A subtype of diabetes mellitus associated with a mutation of mitochondrial DNA. N. Engl. J. Med. 1994, 330, 962–968. [Google Scholar] [CrossRef]
- Yasukawa, T.; Kirino, Y.; Ishii, N.; Holt, I.J.; Jacobs, H.T.; Makifuchi, T.; Fukuhara, N.; Ohta, S.; Suzuki, T.; Watanabe, K. Wobble modification deficiency in mutant tRNAs in patients with mitochondrial diseases. FEBS Lett. 2005, 579, 2948–2952. [Google Scholar] [CrossRef] [Green Version]
- Zapico, S.C.; Ubelaker, D.H. mtDNA mutations and their role in aging, diseases and forensic sciences. Aging Dis. 2013, 4, 364. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Merchant, S.N.; Miyazawa, T.; Yamaguchi, T.; McKenna, M.J.; Kouda, H.; Iino, Y.; Someya, T.; Tamagawa, Y.; Takiyama, Y. Temporal bone histopathological and quantitative analysis of mitochondrial DNA in MELAS. Laryngoscope 2003, 113, 1362–1368. [Google Scholar] [CrossRef]
- Brown, B.G.; Zhao, X.-Q.; Chait, A.; Fisher, L.D.; Cheung, M.C.; Morse, J.S.; Dowdy, A.A.; Marino, E.K.; Bolson, E.L.; Alaupovic, P. Simvastatin and niacin, antioxidant vitamins, or the combination for the prevention of coronary disease. N. Engl. J. Med. 2001, 345, 1583–1592. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, K.; Akai, H.; Muramatsu, K. Effects of methionine and related compounds on plasma cholesterol level in rats fed a high cholesterol diet. J. Nutr. Sci. Vitaminol. 1986, 32, 537–549. [Google Scholar] [CrossRef]
- Ebadi, M.; Sharma, S.K.; Wanpen, S.; Amornpan, A. Coenzyme Q10 inhibits mitochondrial complex-1 down-regulation and nuclear factor-kappa B activation. J. Cell. Mol. Med. 2004, 8, 213–222. [Google Scholar] [CrossRef]
- Youn, H.-S.; Lim, H.J.; Choi, Y.J.; Lee, J.Y.; Lee, M.-Y.; Ryu, J.-H. Selenium suppresses the activation of transcription factor NF-κB and IRF3 induced by TLR3 or TLR4 agonists. Int. Immunopharmacol. 2008, 8, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Segain, J.P.; De La Blétiere, D.R.; Bourreille, A.; Leray, V.; Gervois, N.; Rosales, C.; Ferrier, L.; Bonnet, C.; Blottiere, H.M.; Galmiche, J.P. Butyrate inhibits inflammatory responses through NFκB inhibition: Implications for Crohn’s disease. Gut 2000, 47, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Lührs, H.; Gerke, T.; Müller, J.G.; Melcher, R.; Schauber, J.; Boxberger, F.; Scheppach, W.; Menzel, T. Butyrate inhibits NF-κB activation in lamina propria macrophages of patients with ulcerative colitis. Scand. J. Gastroenterol. 2002, 37, 458–466. [Google Scholar] [CrossRef]
- Hoshino, T.; Tabuchi, K.; Nishimura, B.; Tanaka, S.; Nakayama, M.; Ishii, T.; Warabi, E.; Yanagawa, T.; Shimizu, R.; Yamamoto, M. Protective role of Nrf2 in age-related hearing loss and gentamicin ototoxicity. Biochem. Biophys. Res. Commun. 2011, 415, 94–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honkura, Y.; Matsuo, H.; Murakami, S.; Sakiyama, M.; Mizutari, K.; Shiotani, A.; Yamamoto, M.; Morita, I.; Shinomiya, N.; Kawase, T. NRF2 is a key target for prevention of noise-induced hearing loss by reducing oxidative damage of cochlea. Sci. Rep. 2016, 6, 19329. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Zhu, Y.; Ye, Y.; Guan, J.; Min, X.; Xiong, H. Nitric oxide protects against cochlear hair cell damage and noise-induced hearing loss through glucose metabolic reprogramming. Free Radic. Biol. Med. 2022, 179, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, U.R.; Schmidtmann, I.; Meuser, R.; Ernst, B.P.; Wünsch, D.; Siemer, S.; Gribko, A.; Stauber, R.H.; Strieth, S. Early Alterations of Endothelial Nitric Oxide Synthase Expression Patterns in the Guinea Pig Cochlea After Noise Exposure. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2019, 67, 845–855. [Google Scholar] [CrossRef]
- Rousset, F.; Nacher-Soler, G.; Coelho, M.; Ilmjarv, S.; Kokje, V.B.C.; Marteyn, A.; Cambet, Y.; Perny, M.; Roccio, M.; Jaquet, V. Redox activation of excitatory pathways in auditory neurons as mechanism of age-related hearing loss. Redox Biol. 2020, 30, 101434. [Google Scholar] [CrossRef] [PubMed]
- Ashoori, M.; Saedisomeolia, A. Riboflavin (vitamin B2) and oxidative stress: A review. Br. J. Nutr. 2014, 111, 1985–1991. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Touyz, R.M.; Park, J.B.; Schiffrin, E.L. Antioxidant effects of vitamins C and E are associated with altered activation of vascular NADPH oxidase and superoxide dismutase in stroke-prone SHR. Hypertension 2001, 38, 606–611. [Google Scholar] [CrossRef] [Green Version]
- Joshi, R.; Adhikari, S.; Patro, B.S.; Chattopadhyay, S.; Mukherjee, T. Free radical scavenging behavior of folic acid: Evidence for possible antioxidant activity. Free Radic. Biol. Med. 2001, 30, 1390–1399. [Google Scholar] [CrossRef]
- Nakagawa, K.; Ninomiya, M.; Okubo, T.; Aoi, N.; Juneja, L.R.; Kim, M.; Yamanaka, K.; Miyazawa, T. Tea catechin supplementation increases antioxidant capacity and prevents phospholipid hydroperoxidation in plasma of humans. J. Agric. Food Chem. 1999, 47, 3967–3973. [Google Scholar] [CrossRef]
- Papucci, L.; Schiavone, N.; Witort, E.; Donnini, M.; Lapucci, A.; Tempestini, A.; Formigli, L.; Zecchi-Orlandini, S.; Orlandini, G.; Carella, G. Coenzyme q10 prevents apoptosis by inhibiting mitochondrial depolarization independently of its free radical scavenging property. J. Biol. Chem. 2003, 278, 28220–28228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillis, J.C.; Benfield, P.; McTavish, D. Idebenone. Drugs Aging 1994, 5, 133–152. [Google Scholar] [CrossRef] [PubMed]
- Marcocci, L.; Packer, L.; Droy-Lefaix, M.-T.; Sekaki, A.; Gardès-Albert, M. Antioxidant action of Ginkgo biloba extract EGb 761. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1994; Volume 234, pp. 462–475. ISBN 0076-6879. [Google Scholar]
- Packer, L.; Witt, E.H.; Tritschler, H.J. Alpha-lipoic acid as a biological antioxidant. Free Radic. Biol. Med. 1995, 19, 227–250. [Google Scholar] [CrossRef]
- Turan, M.; Ciğer, E.; Arslanoğlu, S.; Börekci, H.; Önal, K. Could edaravone prevent gentamicin ototoxicity? An experimental study. Hum. Exp. Toxicol. 2017, 36, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Feldman, L.; Efrati, S.; Eviatar, E.; Abramsohn, R.; Yarovoy, I.; Gersch, E.; Averbukh, Z.; Weissgarten, J. Gentamicin-induced ototoxicity in hemodialysis patients is ameliorated by N-acetylcysteine. Kidney Int. 2007, 72, 359–363. [Google Scholar] [CrossRef] [Green Version]
- Takumida, M.; Popa, R.; Anniko, M. Free radicals in the guinea pig inner ear following gentamicin exposure. ORL 1999, 61, 63–70. [Google Scholar] [CrossRef]
- Fetoni, A.R.; Eramo, S.L.M.; Rolesi, R.; Troiani, D.; Paludetti, G. Antioxidant treatment with coenzyme Q-ter in prevention of gentamycin ototoxicity in an animal model. Acta Otorhinolaryngol. Ital. 2012, 32, 103. [Google Scholar]
- Sha, S.-H.; Schacht, J. Antioxidants attenuate gentamicin-induced free radical formation in vitro and ototoxicity in vivo: D-methionine is a potential protectant. Hear. Res. 2000, 142, 34–40. [Google Scholar] [CrossRef]
- Kalinec, G.M.; Fernandez-Zapico, M.E.; Urrutia, R.; Esteban-Cruciani, N.; Chen, S.; Kalinec, F. Pivotal role of Harakiri in the induction and prevention of gentamicin-induced hearing loss. Proc. Natl. Acad. Sci. USA 2005, 102, 16019–16024. [Google Scholar] [CrossRef] [Green Version]
- Lynch, E.D.; Gu, R.; Pierce, C.; Kil, J. Reduction of acute cisplatin ototoxicity and nephrotoxicity in rats by oral administration of allopurinol and ebselen. Hear. Res. 2005, 201, 81–89. [Google Scholar] [CrossRef]
- Rybak, L.P.; Husain, K.; Morris, C.; Whitworth, C.; Somani, S. Effect of protective agents against cisplatin ototoxicity. Otol. Neurotol. 2000, 21, 513–520. [Google Scholar]
- Kim, J.; Cho, H.-J.; Sagong, B.; Kim, S.-J.; Lee, J.-T.; So, H.-S.; Lee, I.-K.; Kim, U.-K.; Lee, K.-Y.; Choo, Y.-S. Alpha-lipoic acid protects against cisplatin-induced ototoxicity via the regulation of MAPKs and proinflammatory cytokines. Biochem. Biophys. Res. Commun. 2014, 449, 183–189. [Google Scholar] [CrossRef]
- Campbell, K.C.M.; Rybak, L.P.; Meech, R.P.; Hughes, L. D-methionine provides excellent protection from cisplatin ototoxicity in the rat. Hear. Res. 1996, 102, 90–98. [Google Scholar] [CrossRef]
- Dickey, D.T.; Muldoon, L.L.; Kraemer, D.F.; Neuwelt, E.A. Protection against cisplatin-induced ototoxicity by N-acetylcysteine in a rat model. Hear. Res. 2004, 193, 25–30. [Google Scholar] [CrossRef]
- Riga, M.G.; Chelis, L.; Kakolyris, S.; Papadopoulos, S.; Stathakidou, S.; Chamalidou, E.; Xenidis, N.; Amarantidis, K.; Dimopoulos, P.; Danielides, V. Transtympanic injections of N-acetylcysteine for the prevention of cisplatin-induced ototoxicity: A feasible method with promising efficacy. Am. J. Clin. Oncol. 2013, 36, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Takumida, M.; Anniko, M. Radical scavengers for elderly patients with age-related hearing loss. Acta Otolaryngol. 2009, 129, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Salami, A.; Mora, R.; Dellepiane, M.; Manini, G.; Santomauro, V.; Barettini, L.; Guastini, L. Water-soluble coenzyme Q10 formulation (Q-TER®) in the treatment of presbycusis. Acta Otolaryngol. 2010, 130, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Ohinata, Y.; Yamasoba, T.; Schacht, J.; Miller, J.M. Glutathione limits noise-induced hearing loss. Hear. Res. 2000, 146, 28–34. [Google Scholar] [CrossRef]
- Yamasoba, T.; Nuttall, A.L.; Harris, C.; Raphael, Y.; Miller, J.M. Role of glutathione in protection against noise-induced hearing loss. Brain Res. 1998, 784, 82–90. [Google Scholar] [CrossRef]
- Fetoni, A.R.; Piacentini, R.; Fiorita, A.; Paludetti, G.; Troiani, D. Water-soluble Coenzyme Q10 formulation (Q-ter) promotes outer hair cell survival in a guinea pig model of noise induced hearing loss (NIHL). Brain Res. 2009, 1257, 108–116. [Google Scholar] [CrossRef]
- Campbell, K.C.M.; Meech, R.P.; Klemens, J.J.; Gerberi, M.T.; Dyrstad, S.S.W.; Larsen, D.L.; Mitchell, D.L.; El-Azizi, M.; Verhulst, S.J.; Hughes, L.F. Prevention of noise-and drug-induced hearing loss with D-methionine. Hear. Res. 2007, 226, 92–103. [Google Scholar] [CrossRef]
- Seidman, M.; Babu, S.; Tang, W.; Naem, E.; Quirk, W.S. Effects of resveratrol on acoustic trauma. Otolaryngol. Neck Surg. 2003, 129, 463–470. [Google Scholar] [CrossRef]
- Fetoni, A.R.; Mancuso, C.; Eramo, S.L.M.; Ralli, M.; Piacentini, R.; Barone, E.; Paludetti, G.; Troiani, D. In vivo protective effect of ferulic acid against noise-induced hearing loss in the guinea-pig. Neuroscience 2010, 169, 1575–1588. [Google Scholar] [CrossRef] [PubMed]
- Le Prell, C.G.; Yamashita, D.; Minami, S.B.; Yamasoba, T.; Miller, J.M. Mechanisms of noise-induced hearing loss indicate multiple methods of prevention. Hear. Res. 2007, 226, 22–43. [Google Scholar] [CrossRef] [Green Version]
- McFadden, S.L.; Woo, J.M.; Michalak, N.; Ding, D. Dietary vitamin C supplementation reduces noise-induced hearing loss in guinea pigs. Hear. Res. 2005, 202, 200–208. [Google Scholar] [CrossRef]
- Yamasoba, T.; Pourbakht, A.; Sakamoto, T.; Suzuki, M. Ebselen prevents noise-induced excitotoxicity and temporary threshold shift. Neurosci. Lett. 2005, 380, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Pourbakht, A.; Yamasoba, T. Ebselen attenuates cochlear damage caused by acoustic trauma. Hear. Res. 2003, 181, 100–108. [Google Scholar] [CrossRef]
- Wu, F.; Xiong, H.; Sha, S. Noise-induced loss of sensory hair cells is mediated by ROS/AMPKα pathway. Redox Biol. 2020, 29, 101406. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.R.; Custer, D.A.; Krieg, E.; Alagramam, K. N-Acetyl L-Cysteine does not protect mouse ears from the effects of noise. J. Occup. Med. Toxicol. 2010, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Kramer, S.; Dreisbach, L.; Lockwood, J.; Baldwin, K.; Kopke, R.; Scranton, S.; O’Leary, M. Efficacy of the antioxidant N-acetylcysteine (NAC) in protecting ears exposed to loud music. J. Am. Acad. Audiol. 2006, 17, 265–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etminan, M.; Gill, S.S.; Samii, A. Intake of vitamin E, vitamin C, and carotenoids and the risk of Parkinson’s disease: A meta-analysis. Lancet Neurol. 2005, 4, 362–365. [Google Scholar] [CrossRef]
- Lima, L.A.R.; Lopes, M.J.P.; Costa, R.O.; Lima, F.A.V.; Neves, K.R.T.; Calou, I.B.F.; Andrade, G.M.; Viana, G.S.B. Vitamin D protects dopaminergic neurons against neuroinflammation and oxidative stress in hemiparkinsonian rats. J. Neuroinflamm. 2018, 15, 249. [Google Scholar] [CrossRef] [PubMed]
- Knekt, P.; Kilkkinen, A.; Rissanen, H.; Marniemi, J.; Sääksjärvi, K.; Heliövaara, M. Serum vitamin D and the risk of Parkinson disease. Arch. Neurol. 2010, 67, 808–811. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Yoshioka, M.; Hashimoto, M.; Murakami, M.; Noya, M.; Takahashi, D.; Urashima, M. Randomized, double-blind, placebo-controlled trial of vitamin D supplementation in Parkinson disease. Am. J. Clin. Nutr. 2013, 97, 1004–1013. [Google Scholar] [CrossRef] [PubMed]
- Fullard, M.E.; Xie, S.X.; Marek, K.; Stern, M.; Jennings, D.; Siderowf, A.; Willis, A.W.; Chen-Plotkin, A.S. Vitamin D in the Parkinson associated risk syndrome (PARS) study. Mov. Disord. 2017, 32, 1636–1640. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, H.; Ishihara, T.; Yokota, O.; Terada, S.; Trojanowski, J.Q.; Lee, V.M.-Y.; Kuroda, S. Effects of α-tocopherol on an animal model of tauopathies. Free Radic. Biol. Med. 2004, 37, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Masaki, K.H.; Losonczy, K.G.; Izmirlian, G.; Foley, D.J.; Ross, G.W.; Petrovitch, H.; Havlik, R.; White, L.R. Association of vitamin E and C supplement use with cognitive function and dementia in elderly men. Neurology 2000, 54, 1265–1272. [Google Scholar] [CrossRef]
- Maison, S.F.; Liu, X.-P.; Eatock, R.A.; Sibley, D.R.; Grandy, D.K.; Liberman, M.C. Dopaminergic Signaling in the Cochlea: Receptor Expression Patterns and Deletion Phenotypes. J. Neurosci. 2012, 32, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; West, M.B.; Cai, Q.; Cheng, W.; Ewert, D.L.; Li, W.; Floyd, R.A.; Kopke, R.D. Antioxidants reduce neurodegeneration and accumulation of pathologic Tau proteins in the auditory system after blast exposure. Free Radic. Biol. Med. 2017, 108, 627–643. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Yan, L.; Lewith, G.; Liu, J. Chinese herbal medicine for idiopathic sudden sensorineural hearing loss: A systematic review of randomised clinical trials. Clin. Otolaryngol. 2013, 38, 455–473. [Google Scholar] [CrossRef] [PubMed]
- Kaya, H.; Koç, A.K.; Sayın, İ.; Güneş, S.; Altıntaş, A.; Yeğin, Y.; Kayhan, F.T. Vitamins A, C, and E and selenium in the treatment of idiopathic sudden sensorineural hearing loss. Eur. Arch. Oto-Rhino-Laryngol. 2015, 272, 1119–1125. [Google Scholar] [CrossRef]
- Curhan, S.G.; Stankovic, K.M.; Eavey, R.D.; Wang, M.; Stampfer, M.J.; Curhan, G.C. Carotenoids, vitamin A, vitamin C, vitamin E, and folate and risk of self-reported hearing loss in women. Am. J. Clin. Nutr. 2015, 102, 1167–1175. [Google Scholar] [CrossRef] [Green Version]
- Shargorodsky, J.; Curhan, S.G.; Eavey, R.; Curhan, G.C. A prospective study of vitamin intake and the risk of hearing loss in men. Otolaryngol. Neck Surg. 2010, 142, 231–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopinath, B.; Flood, V.M.; McMahon, C.M.; Burlutsky, G.; Spankovich, C.; Hood, L.J.; Mitchell, P. Dietary antioxidant intake is associated with the prevalence but not incidence of age-related hearing loss. J. Nutr. Health Aging 2011, 15, 896–900. [Google Scholar] [CrossRef] [PubMed]
- Aruoma, O.I.; Hayashi, Y.; Marotta, F.; Mantello, P.; Rachmilewitz, E.; Montagnier, L. Applications and bioefficacy of the functional food supplement fermented papaya preparation. Toxicology 2010, 278, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Fardet, A.; Rock, E. In vitro and in vivo antioxidant potential of milks, yoghurts, fermented milks and cheeses: A narrative review of evidence. Nutr. Res. Rev. 2018, 31, 52–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da-Costa-Rocha, I.; Bonnlaender, B.; Sievers, H.; Pischel, I.; Heinrich, M. Hibiscus sabdariffa L.–A phytochemical and pharmacological review. Food Chem. 2014, 165, 424–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieto, G.; Ros, G.; Castillo, J. Antioxidant and antimicrobial properties of rosemary (Rosmarinus officinalis L.): A review. Medicines 2018, 5, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera, C.; Artacho, R.; Giménez, R. Beneficial effects of green tea—A review. J. Am. Coll. Nutr. 2006, 25, 79–99. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Abu-Ghannam, N. Bioactive potential and possible health effects of edible brown seaweeds. Trends Food Sci. Technol. 2011, 22, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Dimidi, E.; Cox, S.R.; Rossi, M.; Whelan, K. Fermented Foods: Definitions and Characteristics, Impact on the Gut Microbiota and Effects on Gastrointestinal Health and Disease. Nutrients 2019, 11, 1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Mori, A.; Chen, Q.; Zhao, B. Fermented papaya preparation attenuates β-amyloid precursor protein: β-amyloid–mediated copper neurotoxicity in β-amyloid precursor protein and β-amyloid precursor protein Swedish mutation overexpressing SH-SY5Y cells. Neuroscience 2006, 143, 63–72. [Google Scholar] [CrossRef]
- Logozzi, M.; Di Raimo, R.; Mizzoni, D.; Andreotti, M.; Spada, M.; Macchia, D.; Fais, S. Beneficial Effects of Fermented Papaya Preparation (FPP®) Supplementation on Redox Balance and Aging in a Mouse Model. Antioxidants 2020, 9, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fardet, A.; Boirie, Y. Associations between food and beverage groups and major diet-related chronic diseases: An exhaustive review of pooled/meta-analyses and systematic reviews. Nutr. Rev. 2014, 72, 741–762. [Google Scholar] [CrossRef]
- Ismail, A.; Ikram, E.H.K.; Nazri, H.S.M. Roselle (Hibiscus sabdariffa L.) seeds-nutritional composition, protein quality and health benefits. Food 2008, 2, 1–16. [Google Scholar]
- Olaleye, M.T.; Rocha, B.T.J. Acetaminophen-induced liver damage in mice: Effects of some medicinal plants on the oxidative defense system. Exp. Toxicol. Pathol. 2008, 59, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Mohd-Esa, N.; Hern, F.S.; Ismail, A.; Yee, C.L. Antioxidant activity in different parts of roselle (Hibiscus sabdariffa L.) extracts and potential exploitation of the seeds. Food Chem. 2010, 122, 1055–1060. [Google Scholar] [CrossRef]
- Frank, T.; Netzel, G.; Kammerer, D.R.; Carle, R.; Kler, A.; Kriesl, E.; Bitsch, I.; Bitsch, R.; Netzel, M. Consumption of Hibiscus sabdariffa L. aqueous extract and its impact on systemic antioxidant potential in healthy subjects. J. Sci. Food Agric. 2012, 92, 2207–2218. [Google Scholar] [CrossRef]
- Wu, J.W.; Lee, M.-H.; Ho, C.-T.; Chang, S.S. Elucidation of the chemical structures of natural antioxidants isolated from rosemary. J. Am. Oil Chem. Soc. 1982, 59, 339–345. [Google Scholar] [CrossRef]
- Habtemariam, S. The Therapeutic Potential of Rosemary (Rosmarinus officinalis) Diterpenes for Alzheimer’s Disease. Evid. Based. Complement. Alternat. Med. 2016, 2016, 2680409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, N.S.L.; Menzies, R.; Hodgson, F.; Wedgewood, P.; Howes, M.-J.R.; Brooker, H.J.; Wesnes, K.A.; Perry, E.K. A randomised double-blind placebo-controlled pilot trial of a combined extract of sage, rosemary and melissa, traditional herbal medicines, on the enhancement of memory in normal healthy subjects, including influence of age. Phytomedicine 2018, 39, 42–48. [Google Scholar] [CrossRef]
- Li, Y.H.; Jiang, P.S.; Su, Y.W.; Liu, Y.M. Role of green tea polyphenols in noise-induced hearing loss. Chinese J. Ind. Hyg. Occup. Dis. 2017, 35, 561–565. [Google Scholar]
- Gu, L.-T.; Yang, J.; Su, S.-Z.; Liu, W.-W.; Shi, Z.-G.; Wang, Q.-R. Green tea polyphenols protects cochlear hair cells from ototoxicity by inhibiting Notch signalling. Neurochem. Res. 2015, 40, 1211–1219. [Google Scholar] [CrossRef]
- Heo, S.-J.; Park, E.-J.; Lee, K.-W.; Jeon, Y.-J. Antioxidant activities of enzymatic extracts from brown seaweeds. Bioresour. Technol. 2005, 96, 1613–1623. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.Y.; Byon, S.-H.; Shin, H.-C.; Han, S.E.; Kim, J.Y.; Byun, J.Y.; Lee, J.D.; Park, M.K. Protective effects of the seaweed phlorotannin polyphenolic compound dieckol on gentamicin-induced damage in auditory hair cells. Int. J. Pediatr. Otorhinolaryngol. 2016, 83, 31–36. [Google Scholar] [CrossRef]
- Zhang, L.; Du, Z.; He, L.; Liang, W.; Liu, K.; Gong, S. ROS-Induced Oxidative Damage and Mitochondrial Dysfunction Mediated by Inhibition of SIRT3 in Cultured Cochlear Cells. Neural Plast. 2022, 2022, 5567174. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Kamogashira, T.; Fujimoto, C.; Iwasaki, S.; Yamasoba, T. Pyrroloquinoline quinone (PQQ) protects mitochondrial function of HEI-OC1 cells under premature senescence. npj Aging 2022, 8, 3. [Google Scholar] [CrossRef]
- Ki, D.-W.; Kim, D.-W.; Song, J.-G.; Woo, E.-E.; Choi, D.-C.; Lee, I.-K.; Yun, B.-S. New antioxidants from the culture broth of Coprinopsis echinospora. J. Antibiot. 2022, 75, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Lutchmanen Kolanthan, V.; Brown, A.; Soobramaney, V.; Philibert, E.G.; Francois Newton, V.; Hosenally, M.; Sokeechand, B.N.; Petkar, G.; Moga, A.; Andres, P.; et al. Clinical Evaluation of Indian Sandalwood Oil and Its Protective Effect on the Skin against the Detrimental Effect of Exposome. Cosmetics 2022, 9, 35. [Google Scholar] [CrossRef]
- Tadokoro, K.; Morihara, R.; Ohta, Y.; Hishikawa, N.; Kawano, S.; Sasaki, R.; Matsumoto, N.; Nomura, E.; Nakano, Y.; Takahashi, Y.; et al. Clinical Benefits of Antioxidative Supplement Twendee X for Mild Cognitive Impairment: A Multicenter, Randomized, Double-Blind, and Placebo-Controlled Prospective Interventional Study. J. Alzheimer’s Dis. 2019, 71, 1063–1069. [Google Scholar] [CrossRef] [PubMed]
- Miyata, H.; Toyoda, Y.; Takada, T.; Hiragi, T.; Kubota, Y.; Shigesawa, R.; Koyama, R.; Ikegaya, Y.; Suzuki, H. Identification of an exporter that regulates vitamin C supply from blood to the brain. iScience 2022, 25, 103642. [Google Scholar] [CrossRef] [PubMed]
- Francois-Newton, V.; Brown, A.; Andres, P.; Mandary, M.B.; Weyers, C.; Latouche-Veerapen, M.; Hettiarachchi, D. Antioxidant and anti-aging potential of Indian sandalwood oil against environmental stressors in vitro and ex vivo. Cosmetics 2021, 8, 53. [Google Scholar] [CrossRef]
- Liu, X.; Yamashita, T.; Shang, J.; Shi, X.; Morihara, R.; Huang, Y.; Sato, K.; Takemoto, M.; Hishikawa, N.; Ohta, Y.; et al. Twendee X Ameliorates Phosphorylated Tau, α-Synuclein and Neurovascular Dysfunction in Alzheimer’s Disease Transgenic Mice With Chronic Cerebral Hypoperfusion. J. Stroke Cerebrovasc. Dis. 2019, 28, 104310. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Yamashita, T.; Yu, H.; Bian, Z.; Hu, X.; Feng, T.; Tadokoro, K.; Morihara, R.; Abe, K. Neuroprotective and Therapeutic Effects of Tocovid and Twendee-X on Aβ Oligomer-Induced Damage in the SH-SY5Y Cell Line. Neurodegener. Dis. 2021, 21, 117–125. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Wu, J.-L.; Shih, T.-S.; Tsai, P.-J.; Sun, Y.-M.; Ma, M.-C.; Guo, Y.L. N-Acetyl-cysteine against noise-induced temporary threshold shift in male workers. Hear. Res. 2010, 269, 42–47. [Google Scholar] [CrossRef]
- Brigelius-Flohé, R.; Traber, M.G. Vitamin E: Function and metabolism. FASEB J. 1999, 13, 1145–1155. [Google Scholar] [CrossRef]
- Oo, S.M.; Oo, H.K.; Takayama, H.; Ishii, K.; Takeshita, Y.; Goto, H.; Nakano, Y.; Kohno, S.; Takahashi, C.; Nakamura, H.; et al. Selenoprotein P-mediated reductive stress impairs cold-induced thermogenesis in brown fat. Cell Rep. 2022, 38, 110566. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Antioxidants | Potential Role in the Prevention/Therapy of Hearing Loss (References) | |
|---|---|---|
| Medicine | Edaravone | Preserved ABR waves against ototoxic drugs in an animal experiment [158]. |
| NAC | Protected hair cell strucure against cisplatin-induced ototoxicity in animal experiments [168,183], and ABR threshold was preserved against drug- and noise-induced hearing loss in human studies [159,184,229]. | |
| Ebselen | Preserved hair cells and ABR threshold against drug- and noise-induced hearing loss in animal experiments [160,164,165,180,181]. | |
| Q-ter | Has a potential to preserve hearing level against noise-induced hearing loss in a clnical study [171] and ameliorated ABR threshold after ototoxic drug and noise exposure [161,174]. | |
| Methionine | The hair cell structure and ABR threshold were preserved against drug- and noise-induced hearing loss in animal experiments [162,167,175]. | |
| L-carnitine | Prevented changes in hearing threshold and cochlear damage in newborn guinea pigs exposed to gentamicin in utero [163] | |
| CoQ10 | Prevented ototoxic apoptosis of the hair cells in a culture experiment [166], and PTA threshold was improved in a human study [170]. | |
| Vitamin C | ABR threshold was preserved against ototoxic drugs in an animal experiment [179]. | |
| GSH | Improved ABR threshold shift after noise exposure, and sensory epithelium was preserved in animal experiments [172,173]. | |
| Resveratrol | Prevented ABR threshold shift after noise exposure in an animal experiment [176]. | |
| Anenexin | Protected stereocillia, hair cells, and cochlear nerve ater noise exposure in an animal experiment [182]. | |
| Natural foods | Green tea | Hair cell loss against noise exposure was decreased (216), and mechanotransduction currents from hair cells against ototoxic drugs were maintained [217]. |
| Brown seaweeds | Enabled to prevent cell damage due to ototoxic drugs in culture experiments [218,219] | |
| New antioxidants | SIRT3-inhibitor | Aggravates cochlear damage due to loss of ribbon synapses and hair cells, increase in apoptosis of hair cells, and ROS production [220]. |
| Pyrroloquinoline quinone | Pyrroloquinoline quinone-treated cells showed decreased mitochondrial potential, promoted mitochondrial fusion, and accelerated mitochondrial movement [221]. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kishimoto-Urata, M.; Urata, S.; Fujimoto, C.; Yamasoba, T. Role of Oxidative Stress and Antioxidants in Acquired Inner Ear Disorders. Antioxidants 2022, 11, 1469. https://doi.org/10.3390/antiox11081469
Kishimoto-Urata M, Urata S, Fujimoto C, Yamasoba T. Role of Oxidative Stress and Antioxidants in Acquired Inner Ear Disorders. Antioxidants. 2022; 11(8):1469. https://doi.org/10.3390/antiox11081469
Chicago/Turabian StyleKishimoto-Urata, Megumi, Shinji Urata, Chisato Fujimoto, and Tatsuya Yamasoba. 2022. "Role of Oxidative Stress and Antioxidants in Acquired Inner Ear Disorders" Antioxidants 11, no. 8: 1469. https://doi.org/10.3390/antiox11081469