
High Exogenous Antioxidant, Restorative Treatment (Heart) for Prevention of the Six Stages of Heart Failure: The Heart Diet

 ,
,  ,
,  ,
,
Abstract
:1. In Sanskrit
2. Translation in English
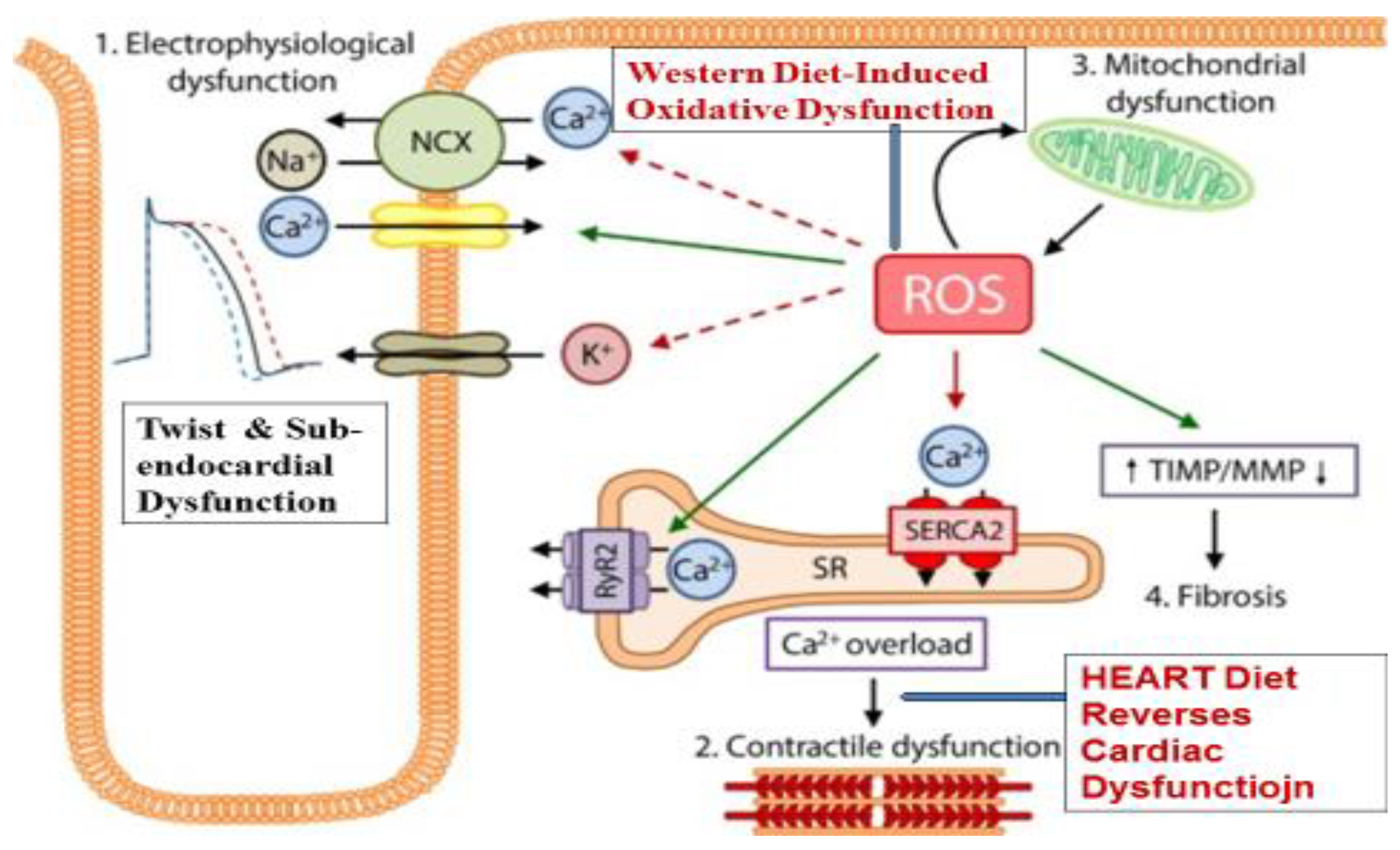
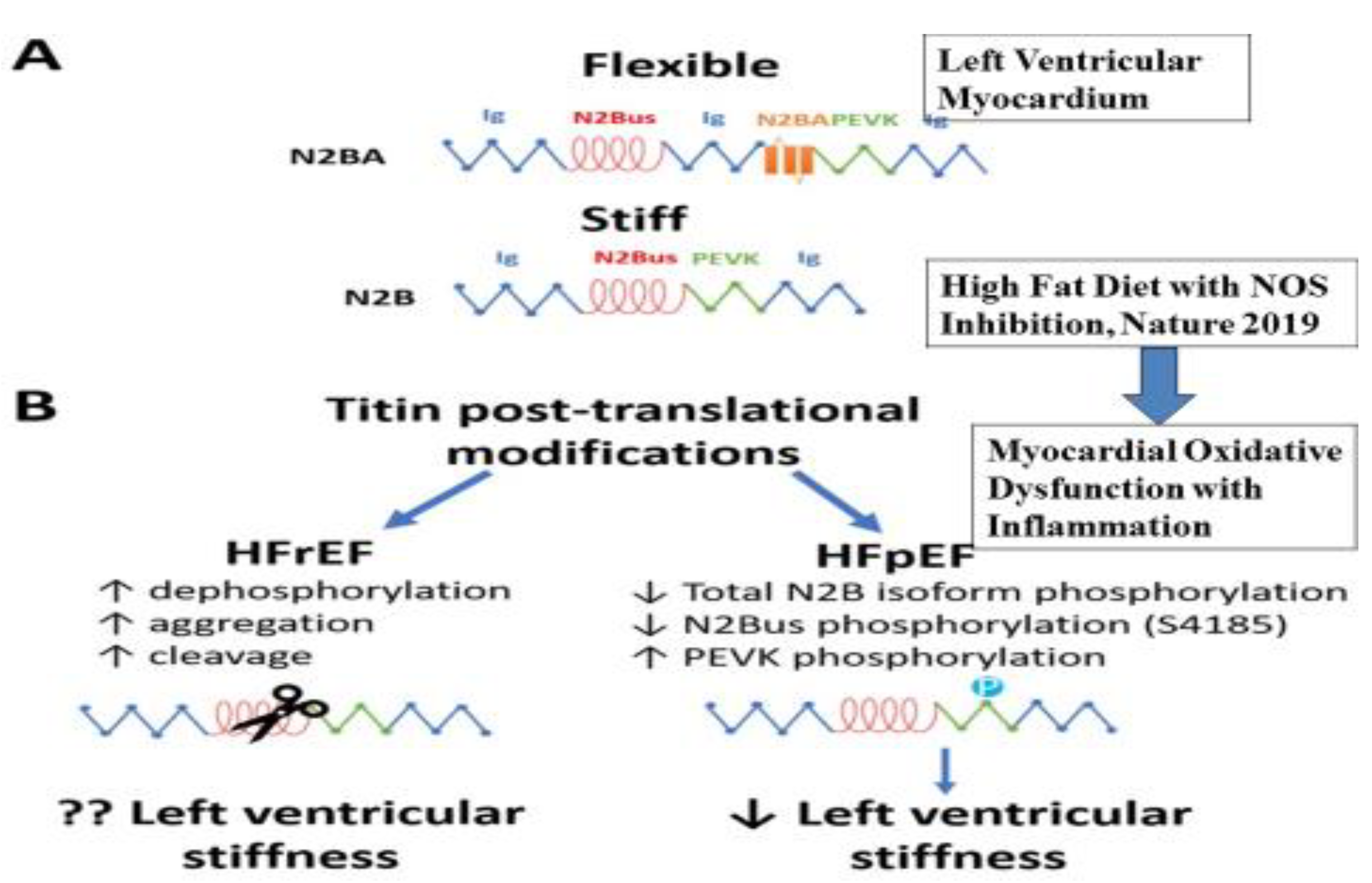
3. Oxidative Dysfunction in Heart Failure
4. Left Ventricular Twist as Function of the Heart
5. Oxidative Dysfunction and Inflammation as Targets for Therapeutic Antioxidants
6. Effects of HEART Diet in Heart Failure
7. Dietary Fat and Risk of Heart Failure
8. Mechanisms of Diet and Obesity in Heart Failure
9. Epidemiological Studies on Diet and Risk of Heart Failure
10. Protective Dietary Patterns in the Prevention of Heart Failure
11. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van der Pol, A.; van Gilst, W.H.; Voors, A.A.; van der Meer, P. Treating oxidative stress in heart failure: Past, present and future. Eur. J. Heart Fail. 2019, 21, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.B.; Torshin, V.I.; Chibisov, S.; Goyal, R.K.; Watanabe, S.; Nakagami, H.; Mogi, M.; Nayak, B.N. Can protective factors inculcate molecular adaptations of cardiomyocyte in the prevention of chronic heart failure? World Heart J. 2019, 11, 149–157. [Google Scholar]
- Singh, R.B.; Sozzi, F.B.; Fedacko, J.; Hristova, K.; Fatima, G.; Pella, D.; Cornelissen, G.; Isaza, A.; Pella, D.; Singh, J.; et al. Pre-heart failure at 2D- and 3D-speckle tracking echocardiography: A comprehensive review. Echocardiography 2022, 39, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Najjar, R.S.; Feresin, R.G. Protective role of polyphenols in heart failure: Molecular targets and cellular mechanisms underlying their therapeutic potential. Int. J. Mol. Sci. 2021, 22, 1668. [Google Scholar] [CrossRef]
- Singh, R.B.; Komatsu, T.; Lee, M.C.; Watanabe, S.; Nwozo, S.O.; Kiyoi, T.; Mogi, M.; Gaur, S.S.; Gautam, R. Effects of behavioral risk factors with reference to smoking on pathophysiology of cardiomyocyte dysfunction. World Heart J. 2020, 12, 9–14. [Google Scholar]
- Singh, R.B.; Fedacko, J.; Goyal, R.; Rai, R.H.; Nandave, M.; Tonk, R.K.; Gaur, S.S.; Gautam, R.; Chibisov, S. Pathophysiology and significance of troponin t, in heart failure, with reference to behavioral risk factors. World Heart J. 2020, 12, 15–22. [Google Scholar]
- Takimoto, E.; Kass, D.A. Role of oxidative stress in cardiac hypertrophy and remodeling. Hypertension 2007, 49, 241–248. [Google Scholar] [CrossRef]
- Singh, R.B.; Elkilany, G.; Fedacko, J.; Hristova, K.; Palmiero, P.; Singh, J.; Manal, M.A.; Badran, H.M. Evolution of the natural history of myocardial twist and diastolic dysfunction as cardiac dysfunction. In Chronic Heart Failure, Pathophysiology and Management; Singh, R.B., Fedacko, J., Elkilany, G., Hristova, K., Eds.; Elsevier: Cambridge, MA, USA, 2023; in press. [Google Scholar]
- Mirończuk-Chodakowska, I.; Witkowska, A.M.; Zujko, M.E. Endogenous non-enzymatic antioxidants in the human body. Adv. Med. Sci. 2018, 63, 68–78. [Google Scholar] [CrossRef]
- Laura, D.C.; Alla, B.; Ahmed, E.S. Nutrigenetics and modulation of oxidative stress. Ann. Nutr. Metab. 2012, 60 (Suppl. S3), 27–36. [Google Scholar] [CrossRef] [Green Version]
- González, R.; Ballester, I.; López-Posadas, R.; Suárez, M.D.; Zarzuelo, A.; Martínez-Augustin, O.; Sánchez de Medina, F. Effects of flavonoids and other polyphenols on inflammation. Crit. Rev. Food Sci. Nutr. 2011, 51, 331–362. [Google Scholar] [CrossRef]
- Singh, R.B.; Fedacko, J.; Elkilany, G.; Hristova, K.; Palmiero, P.; Pella, D.; Cornelissen, G.; Isaza, A.; Pella, D. 2020 Guidelines on Pre-Heart Failure in the Light of 2D and 3D Speckle Tracking Echocardiography. A Scientific Statement of the International College of Cardiology. World Heart J. 2020, 12, 50–70. [Google Scholar]
- Mann, D.L.; Bristow, M.R. Mechanisms and models in heart failure: The biomechanical model and beyond. Circulation 2005, 111, 2837–2849. [Google Scholar] [CrossRef] [Green Version]
- Halabi, A.; Yang, H.; Wright, L.; Potter, E.; Huynh, Q.; Negishi, K.; Marwick, T.H. Evolution of myocardial dysfunction in asymptomatic patients at risk of heart failure. JACC Cardiovasc. Imaging 2021, 14, 350–361. [Google Scholar] [CrossRef]
- Lower, R. Tractatus de Corde; Oxford University Press: London, UK, 1669. [Google Scholar]
- Sengupta, P.P.; Tajik, A.J.; Chandrasekaran, K.; Khandheria, B.K. Twist mechanics of the left ventricle: Principles and application. JACC Cardiovasc. Imaging 2008, 1, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Nakatani, S. Left ventricular rotation and twist: Why should we learn? J. Cardiovasc. Ultrasound 2011, 19, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.K.; Kiotsekoglou, A.; Nanda, N.C. Echocardiography 2020: Toward deciphering the “Rosetta stone” of left ventricular diastolic function. Echocardiography 2020, 37, 1886–1889. [Google Scholar] [CrossRef]
- Khouri, M.G.; Peshock, R.M.; Ayers, C.R.; de Lemos, J.A.; Drazner, M.H. A 4-tiered classification of left ventricular hypertrophy based on left ventricular geometry: The Dallas heart study. Circ. Cardiovasc. Imaging 2010, 3, 164–171. [Google Scholar] [CrossRef]
- Lacalzada, J.; de la Rosa, A.; Izquierdo, M.M.; Jiménez, J.J.; Iribarren, J.L.; García-González, M.J.; López, B.M.; Duque, M.A.; Barragán, A.; Hernández, C.; et al. Left ventricular global longitudinal systolic strain predicts adverse remodeling and subsequent cardiac events in patients with acute myocardial infarction treated with primary percutaneous coronary intervention. Int. J. Cardiovasc. Imaging 2015, 31, 575–584. [Google Scholar] [CrossRef]
- Hristova, K.; Singh, R.B.; Fedacko, J.; Toda, E.; Kumar, A.; Saxena, M.; Baby, A.; Takahashi, T.; De Meester, F.; Wilson, D.W. Causes and risk factors of congestive heart failure in India. World Heart J. 2013, 5, 13–20. [Google Scholar]
- Fedacko, J.; Singh, R.B.; Gupta, A.; Hristova, K.; Toda, E.; Kumar, A.; Saxena, M.; Baby, A.; Singh, R.; Takahashi, T.; et al. Inflammatory mediators in chronic heart failure in North India. Acta Cardiol. 2014, 69, 391–984. [Google Scholar] [CrossRef]
- Simmonds, S.J.; Cuijpers, I.; Heymans, S.; Jones, E.A.V. Cellular and Molecular Differences between HFpEF and HFrEF: A Step Ahead in an Improved Pathological Understanding. Cells 2020, 9, 242. [Google Scholar] [CrossRef] [Green Version]
- Elkilany, G.; Singh, R.B.; Hristova, K.; Milovanovic, B.; Chaves, H.; Wilson, D.W.; Saboo, B.; Mahashwari, A. Beyond drug therapy, nutritional perspectives in the management of chronic heart failure. World Heart J. 2015, 7, 83–88. [Google Scholar]
- Singh, R.B.; Fedacko Mojto, V.; Pella, D. Coenzyme Q10 modulates remodeling possibly by decreasing angiotensin-converting enzyme in patients with acute coronary syndrome. Antioxidants 2018, 7, 99. [Google Scholar] [CrossRef] [Green Version]
- Fedacko, J.; Singh, R.B.; Niaz, M.A.; Bharadwaj, K.; Verma, N.; Gupta, A.K.; Singh, R.B. Association of coronary protective factors among patients with acute coronary syndromes. J. Cardiol. Ther. 2016, 4, 671–677. [Google Scholar] [CrossRef]
- Singh, R.B.; Cornelissen, G.; Takahashi, T.; Shastun, S.; Hristova, K.; Chibisov, S.; Keim, M.; Abramova, M.; Otsuka, K.; Saboo, B.; et al. Brain-heart interactions and circadian rhythms in chronic heart failure. World Heart J. 2015, 7, 129–142. [Google Scholar]
- Singh, R.B.; Hristova, K.; Fedacko, J.; El-Kilany, G.; Cornelissen, G. Chronic heart failure: A disease of the brain. Heart Fail. Rev. 2018, 24, 301–307. [Google Scholar] [CrossRef]
- Singh, R.B.; Wilczynska Fedacko, J.; Takahashi, T.; Niaz, M.A.; Jain SFatima, G.; Manal, M.A.; Abla, M.A.S. Association of Indo-Mediterranean neuroprotective dietary (MIND) pattern with memory impairment and dementia, in an urban population of north India. Int. J. Clin. Nutr. 2021, 21, 11–20. [Google Scholar]
- Wilczynska, A.; Fedacko, J.; Hristova, K.; Alkilany, G.; Fatima, G.; Tyagi, G.; Mojto, V.; Suchday, S. Association of dietary pattern and depression with risk of cardiovascular diseases. Int. J. Clin. Nutr. 2017, 17, 1–10. [Google Scholar]
- Singh, R.B.; Gvozdjakova, A.; Singh, J.; Shastun, S.; Dhalla, N.S.; Pella, D.; Fedacko, J.; Cornelissen, G. Omega-3 PUFA, Omega-6 PUFA and mitochondrial dysfunction in relation to remodelling. In Recent Advances in Mitochondrial Medicine and Coenzyme Q10; Gvozdjakova, A., Cornelissen, G., Singh, R.B., Eds.; Nova Science Publishers: Hauppauge, NY, USA, 2018; Chapter 23; pp. 353–368. [Google Scholar]
- Nettleton, J.A.; Steffen, L.M.; Loehr, L.R.; Rosamond, W.D.; Folsom, A.R. Incident heart failure is associated with lower whole-grain intake and greater high-fat dairy and egg intake in the Atherosclerosis Risk in Communities (ARIC) study. J. Am. Diet. Assoc. 2008, 108, 1881–1887. [Google Scholar] [CrossRef] [Green Version]
- Ashaye, A.; Gaziano, J.; Djoussé, L. Red meat consumption and risk of heart failure in male physicians. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 941–946. [Google Scholar] [CrossRef] [Green Version]
- Kaluza, J.; Åkesson, A.; Wolk, A. Long-term processed and unprocessed red meat consumption and risk of heart failure: A prospective cohort study of women. Int. J. Cardiol. 2015, 193, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Kaluza, J.; Akesson, A.; Wolk, A. Processed and unprocessed red meat consumption and risk of heart failure: Prospective study of men. Circ. Heart Fail. 2014, 7, 552–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levitan, E.B.; Mittleman, M.A.; Wolk, A. Dietary glycemic index, dietary glycemic load, and incidence of heart failure events: A prospective study of middle-aged and elderly women. J. Am. Coll. Nutr. 2010, 29, 65–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, S.F.; Lin, R.C.; Laybutt, D.R.; Barres, R.; Owens, J.A.; Morris, M.J. Chronic high-fat diet in fathers programs β-cell dysfunction in female rat offspring. Nature 2010, 467, 963–966. [Google Scholar] [CrossRef]
- Schiattarella, G.G.; Altamirano, F.; Tong, D.; French, K.M.; Villalobos, E.; Kim, S.Y.; Luo, X.; Jiang, N.; May, H.I.; Wang, Z.V.; et al. Nitrosative stress drives heart failure with preserved ejection fraction. Nature 2019, 568, 351–356. [Google Scholar] [CrossRef]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; DuGar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.-M. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Heaney, L.M.; Bhandari, S.S.; Jones, D.J.L.; Ng, L.L. Trimethylamine N-oxide and prognosis in acute heart failure. Heart 2016, 102, 841–848. [Google Scholar] [CrossRef] [Green Version]
- Dambrova, M.; Latkovskis, G.; Kuka, J.; Strele, I.; Konrade, I.; Grinberga, S.; Hartmane, D.; Pugovics, O.; Erglis, A.; Liepinsh, E. Diabetes is associated with higher trimethylamine N-oxide plasma levels. Exp. Clin. Endocrinol. Diabetes 2016, 124, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Papandreou, C.; Moré, M.; Bellamine, A. Trimethylamine N-Oxide in relation to cardiometabolic health—Cause or effect? Nutrients 2020, 12, 1330. [Google Scholar] [CrossRef]
- O’Shea, K.M.; Khairallah, R.J.; Sparagna, G.C.; Xu, W.; Hecker, P.A.; Robillard-Frayne, I.; Des Rosiers, C.; Kristian, T.; Murphy, R.C.; Fiskum, G.; et al. Dietary omega-3 fatty acids alter cardiac mitochondrial phospholipid composition and delay Ca2+-induced permeability transition. J. Mol. Cell. Cardiol. 2009, 47, 819–827. [Google Scholar] [CrossRef] [Green Version]
- Miatello, R.; Vazquez, M.; Renna, N.; Cruzado, M.; Zumino, A.P.; Risler, N. Chronic administration of resveratrol prevents biochemical cardiovascular changes in fructose-fed rats. Am. J. Hypertens. 2005, 18, 864–870. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Lemon, S.C.; Olendzki, B.; Magner, R.; Li, W.; Culver, A.L.; Ockene, I.; Goldberg, R.J. The dietary quality of persons with heart failure in NHANES 1999–2006. J. Gen. Intern. Med. 2010, 25, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Rahman, I.; Wolk, A.; Latsson, S.C. The relationship between sweetened beverages consumption and risk of heart failure in men. Heart 2015, 101, 1961–1965. [Google Scholar] [CrossRef] [Green Version]
- Stanley, W.C.; Dabkowski, E.R.; Ribeiro, R.F., Jr.; O’Connell, K.A. Dietary fat and heart failure: Moving from lipotoxicity to lipoprotection. Circ. Res. 2012, 110, 764–776. [Google Scholar] [CrossRef] [Green Version]
- Tikellis, C.; Thomas, M.C.; Harcourt, B.E.; Coughlan, M.T.; Pete, J.; Bialkowski, K.; Tan, A.; Bierhaus, A.; Cooper, M.E.; Forbes, J.M. Cardiac inflammation associated with a Western diet is mediated via activation of RAGE by AGEs. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E323–E330. [Google Scholar] [CrossRef] [Green Version]
- Butler, T.J.; Ashford, D.; Seymour, A.M. Western diet increases cardiac ceramide content in healthy and hypertrophied hearts. Nutr. Metab. Cardiovas. Dis. 2017, 27, 991–998. [Google Scholar] [CrossRef]
- Lopaschuk, G.D.; Folmes, C.D.; Stanley, W.C. Cardiac energy metabolism in obesity. Circ. Res. 2007, 101, 335–347. [Google Scholar] [CrossRef] [Green Version]
- Levitan, E.B.; Wolk, A.; Mittleman, M.A. Relation of consistency with the dietary approaches to stop hypertension diet and incidence of heart failure in men aged 45 to 79 years. Am. J. Cardiol. 2009, 104, 1416–1420. [Google Scholar] [CrossRef] [Green Version]
- Levitan, E.B.; Wolk, A.; Mittleman, M.A. Consistency with the DASH diet and incidence of heart failure. Arch. Intern. Med. 2009, 169, 851–857. [Google Scholar] [CrossRef]
- Hall, J.E.; da Silva, A.A.; do Carmo, J.M.; Dubinion, J.; Hamza, S.; Munusamy, S.; Smith, G.; Stec, D.E. Obesity-induced hypertension: Role of sympathetic nervous system, leptin, and melanocortins. J. Biol. Chem. 2010, 285, 17271–17276. [Google Scholar] [CrossRef] [Green Version]
- Okere, I.C.; Chandler, M.P.; McElfresh, T.A.; Rennison, J.H.; Sharov, V.; Sabbah, H.N.; Tserng, K.Y.; Hoit, B.D.; Ernsberger, P.; Young, M.E.; et al. Differential effects of saturated and unsaturated fatty acid diets on cardiomyocyte apoptosis, adipose distribution, and serum leptin. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H38–H44. [Google Scholar] [CrossRef]
- Fitzgerald, S.M.; Henegar, J.R.; Brands, M.W.; Henegar, L.K.; Hall, J.E. Cardiovascular and renal responses to a high-fat diet in Osborne-Mendel rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 281, R547–R552. [Google Scholar] [CrossRef]
- Pladevall, M.; Williams, K.; Guyer, H.; Sadurni, J.; Falces, C.; Ribes, A.; Pare, C.; Brotons, C.; Gabriel, R.; Serrano-Rios, M.; et al. The association between leptin and left ventricular hypertrophy: A population-based cross-sectional study. J. Hypertens. 2003, 21, 1467–1473. [Google Scholar] [CrossRef]
- Schaffer, J.E. Lipotoxicity: When tissues overeat. Curr. Opin. Lipidol. 2003, 14, 281–287. [Google Scholar] [CrossRef]
- Lennie, T.A.; Andreae, C.; Rayens, M.K.; Song, E.K.; Dunbar, S.B.; Pressler, S.J.; Heo, S.; Kim, J.; Moser, D.K. Micronutrient Deficiency Independently Predicts Time to Event in Patients with Heart Failure. J. Am. Heart Assoc. 2018, 7, e007251. [Google Scholar] [CrossRef]
- Song, E.K.; Kang, S.M. Micronutrient deficiency independently predicts adverse health outcomes in patients with heart failure. J. Cardiovasc. Nurs. 2017, 32, 47–53. [Google Scholar] [CrossRef]
- Arcand, J.; Floras, V.; Ahmed, M.; Al-Hesayen, A.; Ivanov, J.; Allard, J.P.; Newton, G.E. Nutritional inadequacies in patients with stable heart failure. J. Am. Diet. Assoc. 2009, 109, 1909–1913. [Google Scholar] [CrossRef]
- Colin-Ramirez, E.; Castillo-Martinez, L.; Orea-Tejeda, A.; Zheng, Y.; Westerhout, C.M.; Ezekowitz, J.A. Dietary fatty acids intake and mortality in patients with heart failure. Nutrition 2014, 30, 1366–1371. [Google Scholar] [CrossRef]
- Abshire, M.; Xu, J.; Baptiste, D.; Almansa, J.R.; Xu, J.; Cummings, A.; Andrews, M.J.; Dennison Himmelfarb, C. Nutritional interventions in heart failure: A systematic review of the literature. J. Card. Fail. 2015, 21, 989–999. [Google Scholar] [CrossRef] [Green Version]
- Djoussé, L.; Petrone, A.B.; Gaziano, J.M. Consumption of fried foods and risk of heart failure in the physicians’ health study. J. Am. Heart Assoc. 2015, 4, e001740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lara, K.M.; Levitan, E.B.; Gutierrez, O.M.; Shikany, J.M.; Safford, M.M.; Judd, S.E.; Rosenson, R.S. Dietary patterns and incident heart failure in US adults without known coronary disease. J. Am. Coll. Cardiol. 2019, 73, 2036–2045. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, H.E.K.; Voutilainen, S.; Kosiken, T.L.; Mursu, T.P.; Toumainen, T.P.; Virtanen, J.K. Intake of different dietary proteins and risk of heart failure in men: The Kuopio Ischaemic Heart Disease Risk Factor Study. Circ. Heart Fail. 2018, 11, e004531. [Google Scholar] [CrossRef] [PubMed]
- Godos, J.; Micek, A.; Brzostek, T.; Toledo, E.; Iacoviello, L.; Astrup, A.; Franco, O.H.; Galvano, F.; Martinez-Gonzalez, M.A.; Grosso, G. Egg consumption and cardiovascular risk: A dose–response meta-analysis of prospective cohort studies. Eur. J. Nutr. 2020, 60, 1833–1862. [Google Scholar] [CrossRef]
- Nwozo, S.O.; Orojobi, F.; Adaramoye, O.A. Hypolipidemic and antioxidant potentials of Xylopia aethiopica seed extract in hypercholesterolaemic rats. J. Med. Foods 2011, 14, 114–119. [Google Scholar] [CrossRef]
- Nwozo, S.O.; Lewis, Y.T.; Oyinloye, B.E. The effects of Piper guineese versus Sesamum indicum aqueous extracts on lipid metabolism and antioxidants in hypercholesterolemic rats. Iran. J. Med. Sci. (IJMS) 2017, 42, 449–456. [Google Scholar]
- Singh, R.B.; Rastogi, S.S.; Verma, R.; Laxmi, B.; Singh Reema Ghosh, S.; Niaz, M.A. Randomized, controlled trial of cardioprotective diet in patients with recent acute myocardial infarction: Results of one year follow up. BMJ 2002, 304, 1115–1119. [Google Scholar]
- Singh, R.B.; Dubnov, G.; Niaz, M.A.; Ghosh, S.; Singh, R.; Rastogi, S.S.; Manor, O.; Pella, D.; Berry, E.M. Effect of an Indo-Mediterranean diet on progression of coronary disease in high risk patients: A randomized single blind trial. Lancet 2002, 360, 1455–1461. [Google Scholar] [CrossRef]
- Papandreou, C.; Hernández-Alonso, P.; Bulló, M.; Ruiz-Canela, M.; Li, J.; Guasch-Ferré, M.; Toledo, E.; Clish, C.; Corella, D.; Estruch, R.; et al. High plasma glutamate and a low glutamine-to-glutamate ratio are associated with increased risk of heart failure but not atrial fibrillation in the Prevención con Dieta Mediterránea (PREDIMED) Study. J. Nutr. 2020, 150, 2882–2889. [Google Scholar] [CrossRef]
- Wirth, J.; di Giuseppe, R.; Boeing, H.; Weikert, C. A Mediterranean-style diet, its components and the risk of heart failure: A prospective population-based study in a non-Mediterranean country. Eur. J. Clin. Nutr. 2016, 70, 1015–1021. [Google Scholar] [CrossRef]
- Djoussé, L.; Akinkuolie, A.O.; Wu, J.H.; Ding, E.L.; Gaziano, J.M. Fish consumption, omega-3 fatty acids and risk of heart failure: A meta-analysis. Clin. Nutr. 2012, 31, 846–853. [Google Scholar] [CrossRef] [Green Version]
- Djoussé, L.; Gaziano, J.M. Breakfast cereals and risk of heart failure in the Physicians’ Health Study I. Arch. Intern. Med. 2007, 167, 2080–2085. [Google Scholar] [CrossRef] [Green Version]
- Mozaffarian, D.; Lemaitre, R.N.; King, I.B.; Song, X.; Spiegelman, D.; Sacks, F.M.; Rimm, E.B.; Siscovick, D.S. Circulating long-chain omega-3 fatty acids and incidence of congestive heart failure in older adults: The Cardiovascular Health Study. Ann. Intern. Med. 2011, 155, 160–170. [Google Scholar] [CrossRef]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Nadal-Ginard, B.; Kajstura, J.; Leri, A.; Anversa, P. Myocyte death, growth, and regeneration in cardiac hypertrophy and failure. Circ. Res. 2003, 92, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Abel, E.D.; Doenst, T. Mitochondrial adaptations to physiological vs. pathological cardiac hypertrophy. Cardiovasc. Res. 2011, 90, 234–242. [Google Scholar] [CrossRef]
- Paulus, W.J. Unfolding discoveries in heart failure. N. Engl. J. Med. 2020, 382, 679–682. [Google Scholar] [CrossRef]
- Amgalan, D.; Kitsis, R.N. A mouse model for the most common form of heart failure. Nature 2019, 568, 324–325. [Google Scholar] [CrossRef] [Green Version]
- Bogiatzi, C.; Gloor, G.; Allen-Vercoe, E.; Reid, G.; Wong, R.G.; Urquhart, B.L.; Dinculescu, V.; Ruetz, K.N.; Velenosi, T.J.; Pignanelli, M.; et al. Metabolic products of the intestinal microbiome and extremes of atherosclerosis. Atherosclerosis 2018, 273, 91–97. [Google Scholar] [CrossRef]
- Spence, J.D.; Srichaikul, K.K.; Jenkins, D.J.A. Cardiovascular Harm from Egg Yolk and Meat: More Than Just Cholesterol and Saturated Fat. J. Am. Heart Assoc. 2021, 10, e017066. [Google Scholar] [CrossRef]
- Magomedova, A.G. Characteristics of nutrition and health of pupils in the regions of Russia. Munic. Educ. Innov. Exp. 2021, 3, 56–64. [Google Scholar] [CrossRef]
- Wang, W.; Kang, P.M. Oxidative stress and antioxidant treatments in cardiovascular diseases. Antioxidants 2020, 9, 1292. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stages (HT on HF) | Manifestations | 2D Echocardio Graphic | 2D Speckle Tracking Echo | 3D Speckle Tracking Echo |
|---|---|---|---|---|
| Stage A | Mild to moderate Oxidative dysfunction, neuro-humoral dysfunction begins. | Increasing filling pressure with abnormal relaxation | Dysfunctional untwist rate LA strain reduced. | Dysfunctional Untwist Rate, LA strain reduced. |
| Stage B | Moderate oxidative dysfunction, hyper-rotation. Sub-endocardial dysfunction. | Dysfunction of systole | Dysfunctional untwist rate and Increased diastolic pressure. LA strain decreased | Dysfunctional untwist rate and Increased diastolic pressure. LA strain decreased |
| Stage C, PHF | Asymptomatic Physio-Pathological remodeling+ | EF% normal > 53% | Normal GLS −20–−23% ≥−27.0% area strain | Normal GLS −17–21% Normal AS −31–−36 % |
| Stage D, PHF and HFpEF | Pathological remodeling disease without symptoms of HF but elevated Natriuretic peptide, dyspnea on exertion | EF% ≥ 50% Systolic LV dysfunction. | EF% 40–49% Impaired GLS–16–20% Impaired GCS, GRS Impaired early diastolic SR, right ventricular LS, and global RV longitudinal SR | Impaired GLS –16–20% Impaired AS −27–31% |
| Stage E, HFmrEF | Structural heart disease with symptoms of HF | EF% 40–49% Grade 1, diastolic dysfunction | Reduced GLS−12–16% Reduced GCS, GRS, Treat with ACE,ARB.ARNI | GLS ≤ −16% AS ≤ −27 % |
| Stage F, HFrEF | Refractory class III HF | EF% < 40% | All above GLS < −12% Treated with ARNI | GLS < −13% AS < −27% |
| Indogenous Antioxidants | Exogenous Antioxidants from HEART Diet |
|---|---|
| Enzymes | Vitamins |
| Superoxide dismutase (SOD) | Vitamin C, ascorbic acid, ascorbate |
| Glutathion peroxidase (GPS) | Vitaminss, E, tocopherol, tocotrienol |
| Glutathion reductase | Vitamin A, vitamin D |
| Glutathion-S-transferase | Polyphenolics and favonoids |
| Paraoxanase | Quercitin, resveratrol |
| Thioredoxin reductase | Catechins; Flavonols, Flavanols |
| Heme- oxygenase | Curcumin |
| Aldehyde dehydrogenase | Anthrocyanins |
| 8-Oxyguanine glycoselase | Phenolic acid |
| Catalase (Iron dependent) | Isoflavons/Genestein |
| Non-enzyme antioxidant | Carotinoids |
| Bilirubin | Alpha-carotine, beta-carotine |
| Coenzyme Q10 | Zeaxanthin |
| L-carnitine | Lutein |
| Alpha-lipoic acid | Lycopine |
| Melatonin | Beta-cryptixanthin |
| Uric acid, cholesterol | Minerals |
| Metal binding proteins | Magnesium |
| Metallothioneine | Selinium, cromium |
| Lactoferrin | Zinc, copper |
| Transferrin | Fiber in the diet; oligosaccharides, polysaccharides |
| Ferritin | Fatty acids; Omega-3 and Monounsaturated |
| Ceruloplasmin (Cu dependent) | Amino acids; L-theanine, arginine, L-tryptophan |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, R.B.; Fedacko, J.; Pella, D.; Fatima, G.; Elkilany, G.; Moshiri, M.; Hristova, K.; Jakabcin, P.; Vaňova, N. High Exogenous Antioxidant, Restorative Treatment (Heart) for Prevention of the Six Stages of Heart Failure: The Heart Diet. Antioxidants 2022, 11, 1464. https://doi.org/10.3390/antiox11081464
Singh RB, Fedacko J, Pella D, Fatima G, Elkilany G, Moshiri M, Hristova K, Jakabcin P, Vaňova N. High Exogenous Antioxidant, Restorative Treatment (Heart) for Prevention of the Six Stages of Heart Failure: The Heart Diet. Antioxidants. 2022; 11(8):1464. https://doi.org/10.3390/antiox11081464
Chicago/Turabian StyleSingh, Ram B., Jan Fedacko, Dominik Pella, Ghizal Fatima, Galal Elkilany, Mahmood Moshiri, Krasimira Hristova, Patrik Jakabcin, and Natalia Vaňova. 2022. "High Exogenous Antioxidant, Restorative Treatment (Heart) for Prevention of the Six Stages of Heart Failure: The Heart Diet" Antioxidants 11, no. 8: 1464. https://doi.org/10.3390/antiox11081464