
A Dietary Antioxidant Formulation Ameliorates DNA Damage Caused by γ-Irradiation in Normal Human Bronchial Epithelial Cells In Vitro

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals, Kits, Antibodies, and AOX2 Formulation
2.2. Cell Culture
2.3. Irradiation by Gammacell Irradiator
2.4. Cell Viability by MTS Assay
2.5. Clonogenic Cell Survival Assay
2.6. Measurement of Intracellular ROS
2.7. DNA Fragmentation Analysis
2.8. γ-H2AX Immunofluorescence Assay
2.9. Comet Assay
2.10. Western Blotting
2.11. Statistical Analysis
3. Results
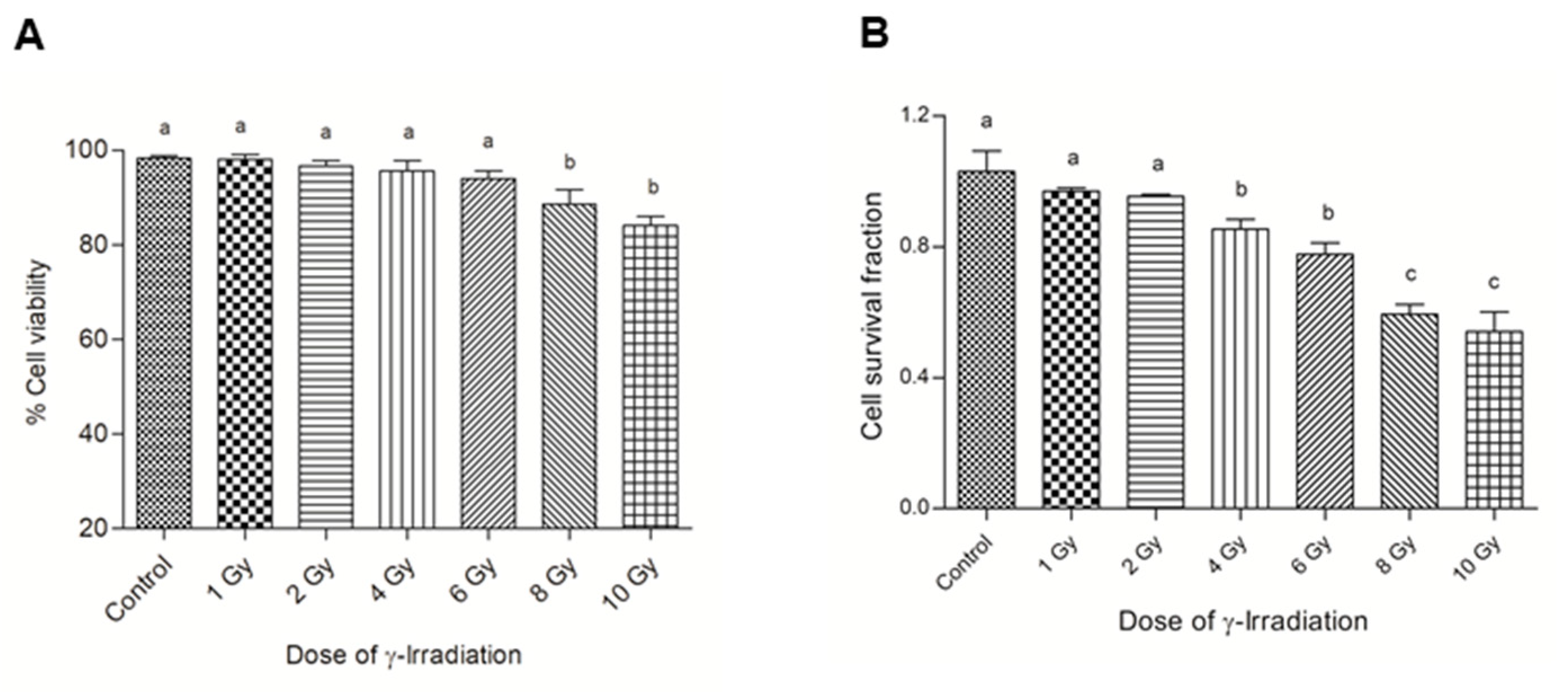
3.1. Assessing the Sensitivity of Normal Bronchial Epithelial Cells to γ-Irradiation
3.2. AOX2 Is Not Cytotoxic at Lower Concentrations
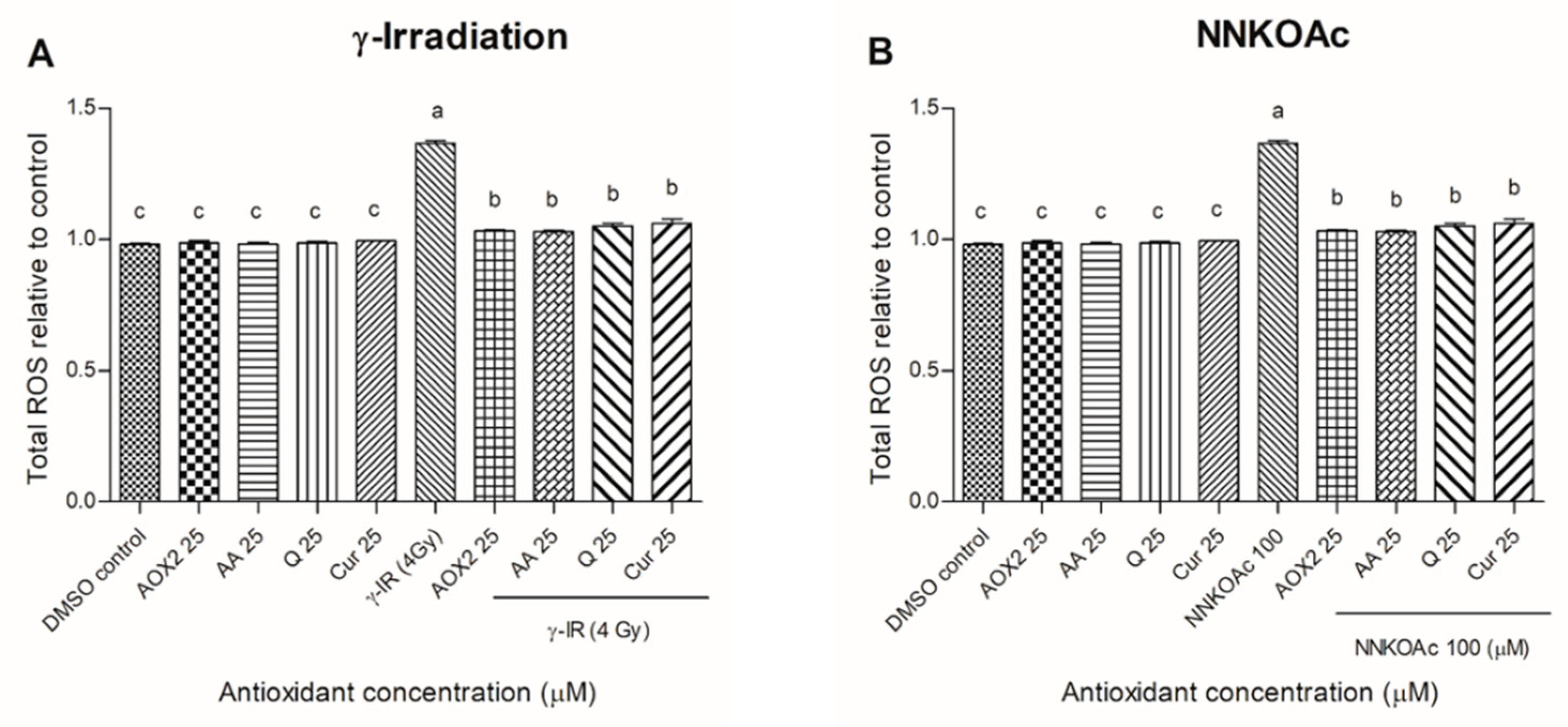
3.3. AOX2 Reduced the γ-Irradiation- and Carcinogen-Induced Intracellular ROS
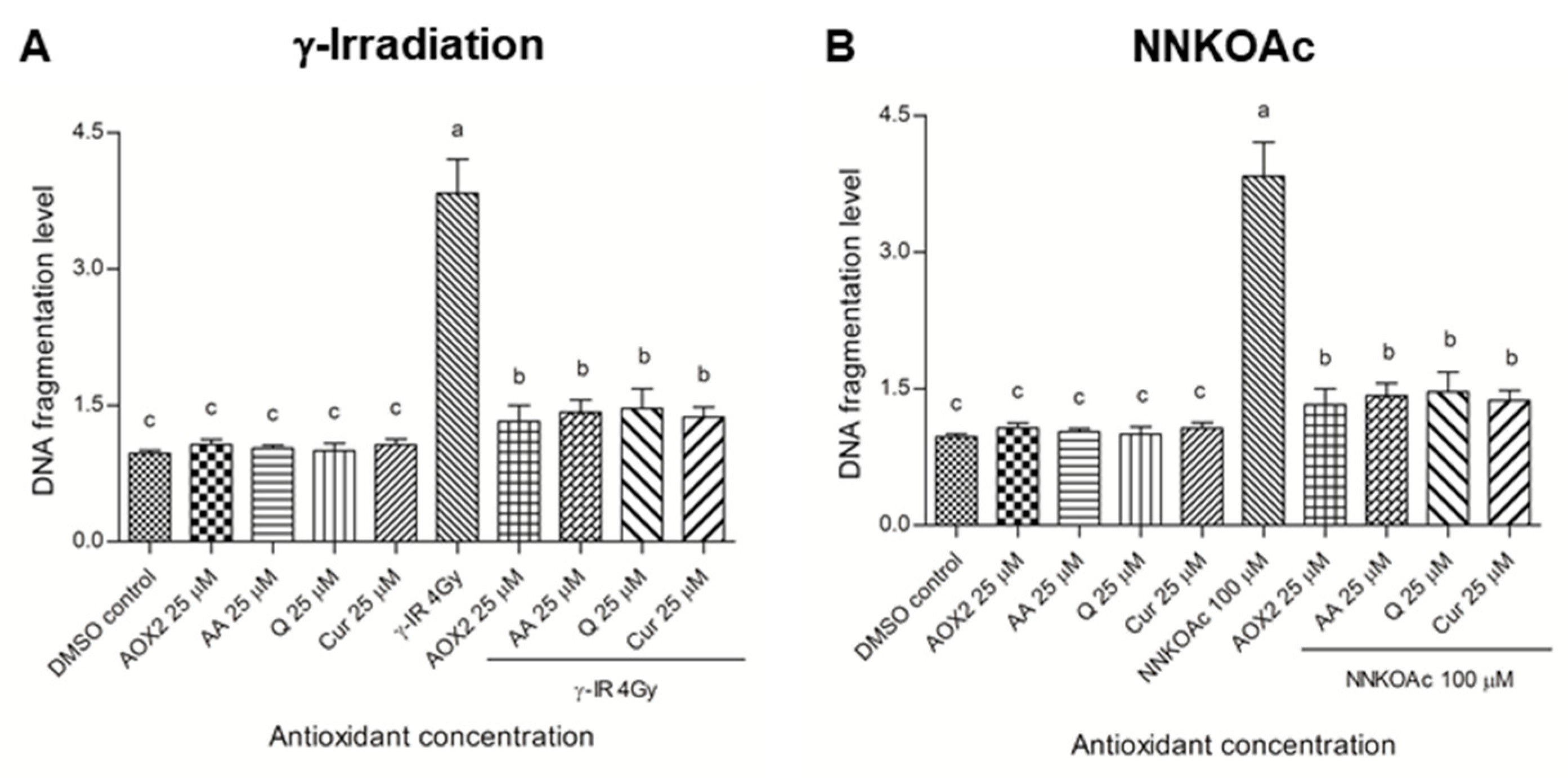
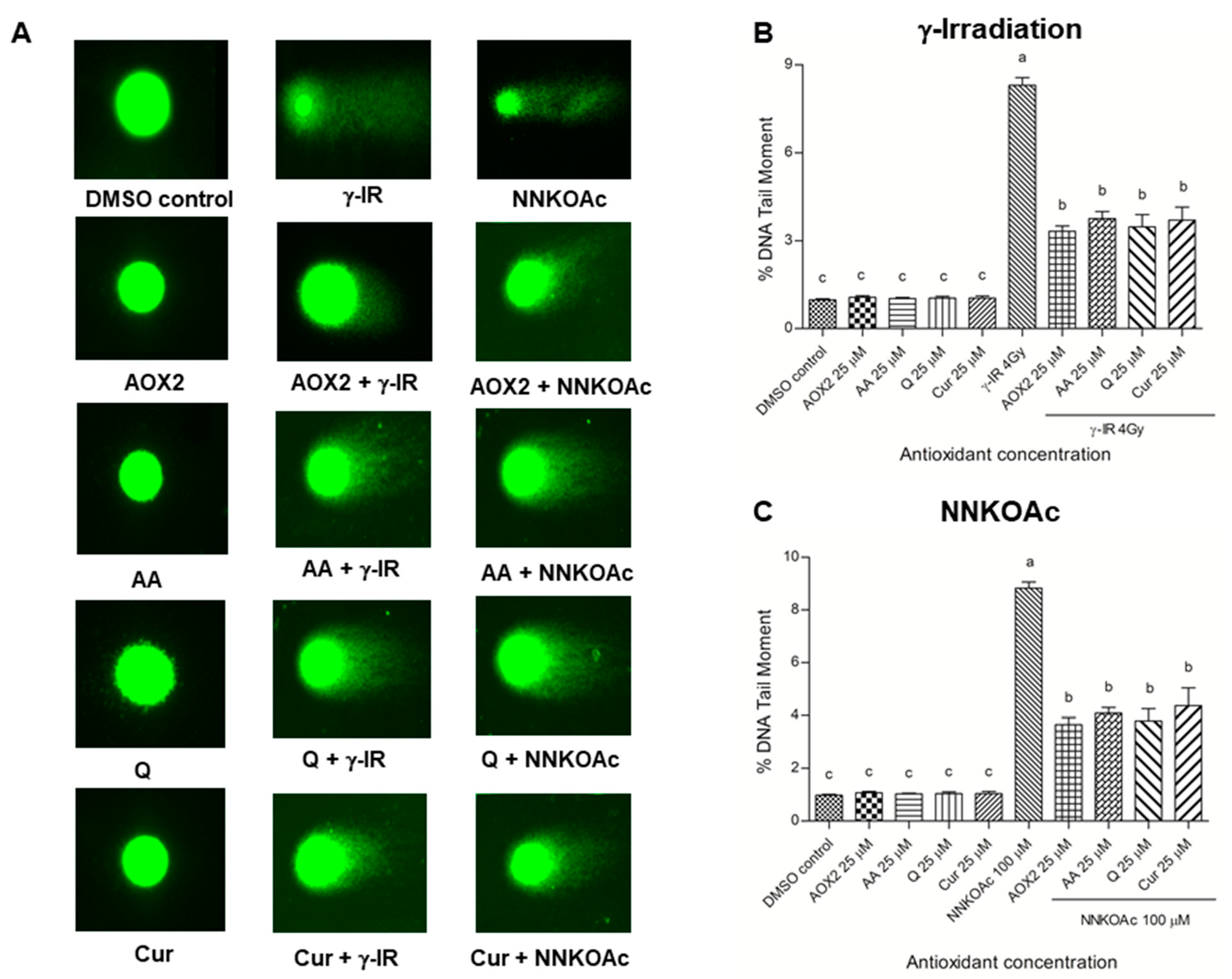
3.4. AOX2 Ameliorates DNA Damage Caused by Carcinogenic Factors
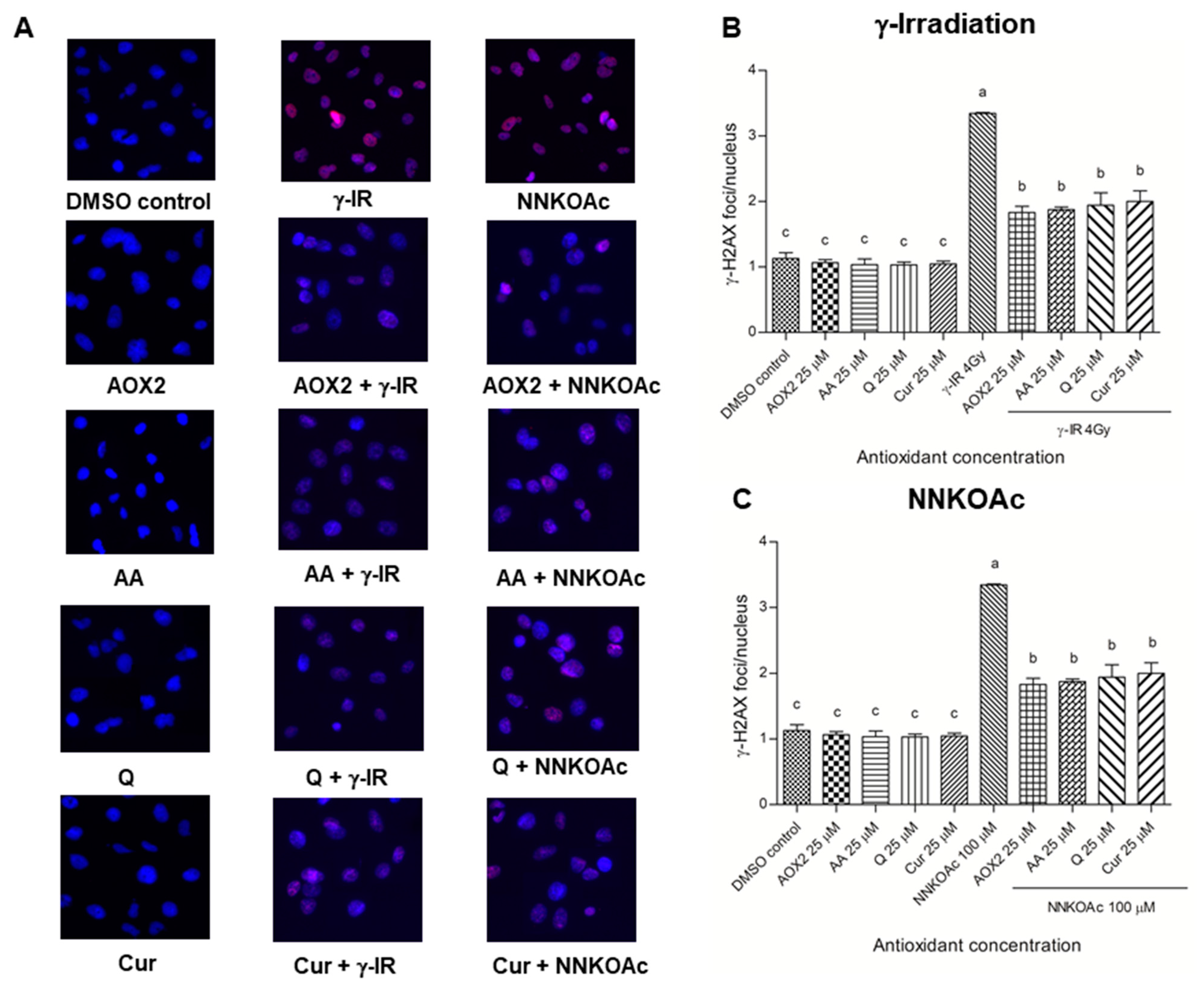
3.5. Dietary Antioxidants Affect DDR Cell Signaling
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Malvezzi, M.; Bertuccio, P.; Levi, F.; La Vecchia, C.; Negri, E. European cancer mortality predictions for the year 2012. Ann. Oncol. 2012, 23, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Wohlbold, L.; Fisher, R.P. Behind the wheel and under the hood: Functions of cyclin-dependent kinases in response to DNA damage. DNA Repair 2009, 8, 1018–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duma, N.; Santana-Davila, R.; Molina, J.R. Non-small cell lung cancer: Epidemiology, screening, diagnosis, and treatment. Mayo Clin. Proc. 2019, 94, 1632–1640. [Google Scholar] [CrossRef] [PubMed]
- Ehrhart, E.; Segarini, P.; Tsang, M.; Carroll, A.G.; Barcellos-Hoff, M.H. Latent transforming growth factor beta1 activation in situ: Quantitative and functional evidence after low-dose gamma-irradiation. FASEB J. 1997, 11, 991–1002. [Google Scholar] [CrossRef]
- Davis, J.D.; Lin, S.-Y. DNA damage and breast cancer. World J. Clin. Oncol. 2011, 2, 329–338. [Google Scholar] [CrossRef]
- Marmary, Y.; Adar, R.; Gaska, S.; Wygoda, A.; Maly, A.; Cohen, J.; Eliashar, R.; Mizrachi, L.; Orfaig-Geva, C.; Baum, B.J.; et al. Radiation-induced loss of salivary gland function isdriven by cellular senescence and prevented by IL6 modulation. Cancer Res. 2016, 76, 1170–1180. [Google Scholar] [CrossRef] [Green Version]
- Kitabatake, K.; Yoshida, E.; Kaji, T.; Tsukimoto, M. Involvement of adenosine A2B receptor in radiation-induced translocation of epidermal growth factor receptor and DNA damage response leading to radioresistance in human lung cancer cells. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129457. [Google Scholar] [CrossRef]
- Hall, S.; Rudrawar, S.; Zunk, M.; Bernaitis, N.; Arora, D.; McDermott, C.M.; Anoopkumar-Dukie. Protection against radiotherapy-induced toxicity. Antioxidants 2016, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Lou, Y.; Guo, Z.; Zhu, Y.; Kong, M.; Zhang, R.; Lu, L.; Wu, F.; Liu, Z.; Wu, J. Houttuynia cordata Thunb. and its bioactive compound 2-undecanone significantly suppress benzo(a)pyrene-induced lung tumorigenesis by activating the Nrf2-HO-1/NQO-1 signaling pathway. J. Exp. Clin. Cancer Res. 2019, 38, 242. [Google Scholar] [CrossRef]
- Ibuki, Y.; Shikata, M.; Toyooka, T. γ-H2AX is a sensitive marker of DNA damage induced by metabolically activated 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone. Toxicol. In Vitro 2015, 29, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Amararathna, M.; Hoskin, D.W.; Rupasinghe, H.P.V. Anthocyanin-rich haskap (Lonicera caerulea L.) berry extracts reduce nitrosamine-induced DNA damage in human normal lung epithelial cells in vitro. Food Chem. Toxicol. 2020, 141, 111404. [Google Scholar] [CrossRef] [PubMed]
- George, V.C.; Rupasinghe, H.P.V. Apple flavonoids suppress carcinogen-induced DNA damage in normal human bronchial epithelial cells. Oxid. Med. Cell. Longev. 2017, 2017, 1767198. [Google Scholar] [CrossRef]
- Nakamura, A.J.; Rao, V.A.; Pommier, Y.; Bonner, W.M. The complexity of phosphorylated H2AX foci formation and DNA repair assembly at DNA double-strand breaks. Cell Cycle 2010, 9, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.; Tho, L.M.; Xu, N.; Gillespie, D.A. The ATM-Chk2 and ATR-Chk1 pathways in DNA damage signaling and cancer. Adv. Cancer Res. 2010, 108, 73–112. [Google Scholar]
- Berquist, B.R.; Wilson, D.M. Pathways for repairing and tolerating the spectrum of oxidative DNA lesions. Cancer Lett. 2012, 327, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Rupasinghe, H.P.V. Flavonoids and their disease prevention and treatment potential: Recent advances and future perspectives. Molecules 2020, 25, 4746. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, S.S.; Rahman, S.; Rupasinghe, H.P.V.; Vazhappilly, C.G. Dietary Flavonoids in p53—Mediated Immune Dysfunctions Linking to Cancer Prevention. Biomedicines 2020, 8, 286. [Google Scholar] [CrossRef]
- Suraweera, T.L.; Rupasinghe, H.P.V.; Dellaire, G.; Xu, Z. Regulation of Nrf2/ARE pathway by dietary flavonoids: A friend or foe for cancer management? Antioxidants 2020, 9, 973. [Google Scholar] [CrossRef]
- Merlin, J.P.J.; Rupasinghe, H.P.V.; Dellaire, G.; Murphy, K. Role of dietary antioxidants in p53-mediated cancer chemoprevention and tumor suppression. Oxid. Med. Cell. Longev. 2021, 2021, 9924328. [Google Scholar] [CrossRef]
- Merlin, J.P.J.; Dellaire, G.; Murphy, K.; Rupasinghe, H.P.V. Vitamin-containing antioxidant formulation reduces carcinogen-induced DNA damage through ATR/Chk1 signaling in bronchial epithelial cells in vitro. Biomedicines 2021, 9, 1665. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Cromie, M.M.; Cai, Q.; Lv, T.; Singh, K.; Gao, W. Curcumin and vitamin E protect against adverse effects of benzo[a]pyrene in lung epithelial cells. PLoS ONE 2014, 9, e92992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koklesova, L.; Liskova, A.; Samec, M.; Qaradakhi, T.; Zulli, A.; Smejkal, K.; Kajo, K.; Jakubikova, J.; Behzadi, P.; Pec, M.; et al. Genoprotective activities of plant natural substances in cancer and chemopreventive strategies in the context of 3P medicine. EPMA J. 2020, 11, 261–287. [Google Scholar] [CrossRef] [PubMed]
- Alcaraz, M.; Olivares, A.; Achel, D.G.; Garcia-Gamuz, J.A.; Castillo, J.; Alcaraz-Saura, M. Genoprotective effect of some flavonoids against genotoxic damage induced by X-rays in vivo: Relationship between structure and activity. Antioxidants 2022, 11, 94. [Google Scholar] [CrossRef]
- Kubatka, P.; Mazurakova, A.; Samec, M.; Koklesova, L.; Zhai, K.; Al-Ishaq, R.; Kajo, K.; Biringer, K.; Vybohova, D.; Brockmueller, A.; et al. Flavonoids against non-physiologic inflammation attributed to cancer initiation, development, and progression-3PM pathways. EPMA J. 2021, 12, 559–587. [Google Scholar] [CrossRef]
- Kitabatake, K.; Kaji, T.; Tsukimoto, M. ATP and ADP enhance DNA damage repair in γ-irradiated BEAS-2B human bronchial epithelial cells through activation of P2X7 and P2Y12 receptors. Toxicol. Appl. Pharmacol. 2020, 407, 115240. [Google Scholar] [CrossRef]
- Wang, H.; Joseph, J.A. Quantifying cellular oxidative stress by dichlorofluorescein assay using microplate reader. Free Radic. Biol. Med. 1999, 27, 612–616. [Google Scholar] [CrossRef]
- Ivashkevich, A.; Redon, C.E.; Nakamura, A.J.; Martin, R.F.; Martin, O.A. Use of the gamma-H2AX assay to monitor DNA damage and repair in cancer research. Cancer Lett. 2012, 327, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Vadnais, C.; Chen, R.; Fraszczak, J.; Yu, Z.; Boulais, J.; Pinder, J.; Frank, D.; Khandanpour, C.; Hebert, J.; Dellaire, G.; et al. GFI1 facilitates efficient DNA repair by regulating PRMT1 dependent methylation of MRE11 and 53BP1. Nat. Commun. 2018, 9, 1418. [Google Scholar] [CrossRef]
- Voller, A.; Bartlett, A.; Bidwell, D.E. Enzyme immunoassays with special reference to ELISA techniques. J. Clin. Pathol. 1978, 31, 507–520. [Google Scholar] [CrossRef] [Green Version]
- Rogakou, E.P.; Pilch, D.R.; Orr, A.H.; Ivanova, V.S.; Bonner, W.M. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J. Biol. Chem. 1998, 273, 5858–5868. [Google Scholar] [CrossRef] [Green Version]
- Mozaffarieh, M.; Schoetzau, A.; Sauter, M.; Grieshaber, M.; Orgul, S.; Golubnitschaja, O.; Flammer, J. Comet assay analysis of single–stranded DNA breaks in circulating leukocytes of glaucoma patients. Mol. Vis. 2008, 14, 1584–1588. [Google Scholar] [PubMed]
- Kodama, Y.; Noda, A.; Booth, C.; Breault, D.; Suda, T.; Hendry, J.; Shinohara, T.; Rube, C.; Nishimura, E.K.; Mitani, H.; et al. International Workshop: Radiation Effects on Mutation in Somatic and Germline Stem Cells. Int. J. Radiat. Biol. 2012, 88, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Lahtz, C.; Bates, S.E.; Jiang, Y.; Li, A.X.; Wu, X.; Hahn, M.A.; Pfeifer, G.P. Gamma irradiation does not induce detectable changes in DNA methylation directly following exposure of human cells. PLoS ONE 2012, 7, e44858. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sim, M.-K.; Loke, W.K.; Chinnathambi, A.; Alharbi, S.A.; Tang, F.R.; Sethi, G. Potential protective effects of ursolic acid against gamma irradiation-induced damage are mediated through the modulation of diverse inflammatory mediators. Front. Pharmacol. 2017, 8, 352. [Google Scholar] [CrossRef] [Green Version]
- Idzko, M.; Ferrari, D.; Eltzschig, H.K. Nucleotide signalling during inflammation. Nature 2014, 509, 310–317. [Google Scholar] [CrossRef] [Green Version]
- Borrego-Soto, G.; Ortiz-Lopez, R.; Rojas-Martinez, A. Ionizing radiation-induced DNA injury and damage detection in patients with breast cancer. Genet. Mol. Biol. 2015, 38, 420–432. [Google Scholar] [CrossRef]
- Hecht, S.S.; Stepanov, I.; Carmella, S.G. Exposure and metabolic activation biomarkers of carcinogenic tobacco-specific nitrosamines. Acc. Chem. Res. 2016, 49, 106–114. [Google Scholar] [CrossRef] [Green Version]
- Hecht, S.S.; Trushin, N.; Reid-Quinn, C.A.; Burak, E.S.; Jones, A.B.; Southers, J.L.; Gombar, C.T.; Carmella, S.G.; Anderson, L.M.; Rice, J.M. Metabolism of the tobacco-specific nitrosamine 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone in the patas monkey: Pharmacokinetics and characterization of glucuronide metabolites. Carcinogenesis 1993, 14, 229–236. [Google Scholar] [CrossRef]
- Ma, B.; Villalta, P.W.; Zarth, A.T.; Kotandeniya, D.; Upadhyaya, P.; Stepanov, I.; Hecht, S.S. Comprehensive high-resolution mass spectrometric analysis of DNA phosphate adducts formed by the tobacco-specific lung carcinogen 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone. Chem. Res. Toxicol. 2015, 28, 2151–2159. [Google Scholar] [CrossRef]
- Cloutier, J.F.; Drouin, R.; Weinfeld, M.; O’Connor, T.R.; Castonguay, A. Characterization and mapping of DNA damage induced by reactive metabolites of 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK) at nucleotide resolution in human genomic DNA. J. Mol. Biol. 2001, 313, 539–557. [Google Scholar] [CrossRef] [PubMed]
- Szeinfeld, D.; De Villiers, N. Radioprotective properties of ATP and modification of acid phosphatase response after a lethal dose of whole body p (66MeV)/Be neutron radiation to BALB/c mice. Cancer Biochem. Biophys. 1992, 13, 123–132. [Google Scholar] [PubMed]
- Nikolov, I.; Rogozkin, V.D.; Pantev, T.; Chertkov, K.S.; Dikovenko, E.A.; Davidova, S.A. Protection of monkeys against prolonged gamma-irradiation. Strahlenther. Onkol. 1986, 162, 200–204. [Google Scholar]
- Agteresch, H.J.; Dagnelie, P.C.; van den Berg, J.W.; Wilson, J.H. Adenosine triphosphate: Established and potential clinical applications. Drugs 1999, 58, 211–232. [Google Scholar] [CrossRef] [PubMed]
- Molkentine, J.M.; Fujimoto, T.N.; Horvath, T.D.; Grossberg, A.J.; Garcia, C.J.G.; Deorukhkar, A.; de la Cruz Bonilla, M.; Lin, D.; Samuel, E.L.G.; Chan, W.K.; et al. Enteral activation of WR-2721 mediates radioprotection and improved survival from lethal fractionated radiation. Sci. Rep. 2019, 9, 1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, J.; Yang, S.; Seng, S. Mechanisms of cancer induction by tobacco-specific NNK and NNN. Cancers 2014, 6, 1138–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, V.C.; Kumar, D.R.; Suresh, P.K.; Kumar, R.A. Antioxidant, DNA protective efficacy and HPLC analysis of Annona muricata (soursop) extracts. J. Food Sci. Technol. 2015, 52, 2328–2335. [Google Scholar] [CrossRef]
- Ramdzan, Z.M.; Ginjala, V.; Pinder, J.B.; Chung, D.; Donovan, C.M.; Kaur, S.; Leduy, L.; Dellaire, G.; Ganesan, S.; Nepveu, A. The DNA repair function of CUX1 contributes to radioresistance. Oncotarget 2017, 8, 19021–19038. [Google Scholar] [CrossRef] [Green Version]
- Clapier, C.R.; Cairns, B.R. The biology of chromatin remodeling complexes. Annu. Rev. Biochem. 2009, 78, 273–304. [Google Scholar] [CrossRef]
- Olive, P.L.; Banath, J.P. Kinetics of H2AX phosphorylation after exposure to cisplatin. Cytom. B. Clin. Cytom. 2009, 76, 79–90. [Google Scholar] [CrossRef]
- Liu, Q.; Guntuku, S.; Cui, X.-S.; Matsuoka, S.; Cortez, D.; Tamai, K.; Luo, G.; Carattini-Rivera, S.; DeMayo, F.; Bradley, A.; et al. Chk1 is an essential kinase that is regulated by Atr and required for the G2/M DNA damage checkpoint. Genes Dev. 2000, 14, 1448–1459. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Albino, A.P.; Jorgensen, E.; Traganos, F.; Darzynkiewicz, Z. DNA damage response induced by tobacco smoke in normal human bronchial epithelial and A549 pulmonary adenocarcinoma cells assessed by laser scanning cytometry. Cytom. A 2009, 75, 840–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Ahn, K.S.; Alharbi, S.A.; Shair, O.H.M.; Arfuso, F.; Sethi, G.; Chinnathambi, A.; Tang, F.R. Celastrol alleviates gamma irradiation-induced damage by modulating diverse inflammatory mediators. Int. J. Mol. Sci. 2020, 21, 1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teafatiller, T.; Agrawal, S.; Robles, G.D.; Rahmatpanah, F.; Subramanian, V.S.; Agrawal, A. Vitamin C enhances antiviral functions of lung epithelial cells. Biomolecules 2021, 11, 1148. [Google Scholar] [CrossRef] [PubMed]
- Corti, A.; Franzini, M.; Casini, A.F.; Paolicchi, A.; Pompella, A. Vitamin C supply to bronchial epithelial cells linked to glutathione availability in elf—A role for secreted γ-glutamyltransferase? J. Cyst. Fibros. 2008, 7, 174–178. [Google Scholar] [CrossRef]
- Pratheeshkumar, P.; Son, Y.-O.; Divya, S.P.; Wang, L.; Turcios, L.; Roy, R.V.; Hitron, J.A.; Kim, D.; Dai, J.; Asha, P.; et al. Quercetin inhibits Cr(VI)-induced malignant cell transformation by targeting miR-21-PDCD4 signaling pathway. Oncotarget 2017, 8, 52118–52131. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Maka, J.C.W.; Man, R.Y.K.; Leung, S.W.S. Flavonoids reduces lipopolysaccharide-induced release of inflammatory mediators in human bronchial epithelial cells: Structure-activity relationship. Eur. J. Pharmacol. 2019, 865, 172731. [Google Scholar] [CrossRef]
- Yang, S.; Huang, X.-L.; Chen, J.; Mao, L.-N.; Liu, X.; Yuan, W.-S.; Wu, X.-J.; Luo, G.-W. Curcumin protects BEAS-2B cells from PM2.5-induced oxidative stress and inflammation by activating NRF2/antioxidant response element pathways. Int. J. Mol. Med. 2021, 47, 45. [Google Scholar] [CrossRef]
- Reis, R.; Orak, D.; Yilmaz, D.; Cimen, H.; Sipahi, H. Modulation of cigarette smoke extract-induced human bronchial epithelial damage by eucalyptol and curcumin. Hum. Exp. Toxicol. 2021, 40, 1445–1462. [Google Scholar] [CrossRef]
- Boersma, M.G.; van der Woude, H.; Bogaards, J.; Boeren, S.; Vervoort, J.; Cnubben, N.H.P.; van Iersel, M.L.P.S.; van Bladeren, P.J.; Rietjens, I.M.C.M. Regioselectivity of Phase II metabolism of luteolin and quercetin by UDP-glucuronosyl transferases. Chem. Res. Toxicol. 2002, 15, 662–670. [Google Scholar] [CrossRef]
- Lykkesfeldt, J.; Tveden-Nyborg, P. The pharmacokinetics of vitamin C. Nutrients 2019, 11, 2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducker, G.S.; Rabinowitz, J.D. One-carbon metabolism in health and disease. Cell Metab. 2017, 25, 27–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andres, E.; Dali-Youcef, N. Cobalamin (Vitamin B12) metabolism and disorders. In Encyclopedia of Food and Health; Elsevier: Amsterdam, The Netherlands, 2016; pp. 160–165. [Google Scholar]
- Solomonson, A.; DeBerardinis, R.J. Lipoic acid metabolism and mitochondrial redox regulation. J. Biol. Chem. 2018, 293, 7522–7530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartolini, D.; Marinelli, R.; Giusepponi, D.; Galarini, R.; Barola, C.; Stabile, A.M.; Sebastiani, B.; Paoletti, F.; Betti, M.; Rende, M.; et al. Alpha-tocopherol metabolites (The vitamin E metabolome) and their interindividual variability during supplementation. Antioxidants 2021, 10, 173. [Google Scholar] [CrossRef]
- Turunen, M.; Olsson, J.; Dallner, G. Metabolism and function of coenzyme Q. BBA 2004, 1660, 171–199. [Google Scholar] [CrossRef] [Green Version]
- Murillo, A.G.; Hu, S.; Fernandez, M.L. Zeaxanthin: Metabolism, properties, and antioxidant protection of eyes, heart, liver, and skin. Antioxidants 2019, 8, 390. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Kim, B.; Lee, J.-Y. Astaxanthin structure, metabolism, and health benefits. J. Hum. Nutr. Food. Sci. 2013, 1, 1003. [Google Scholar]
- Zhou, L.; Jiao, L.; Ju, J.; Ma, X. Effect of sodium selenite on the metabolite profile of Epichloe sp. Mycelia from Festuca sinensis in solid culture. Biol. Trace Elem. Res. 2022, 1, 1–15. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Merlin, J.P.J.; Mathavarajah, S.; Dellaire, G.; Murphy, K.P.J.; Rupasinghe, H.P.V. A Dietary Antioxidant Formulation Ameliorates DNA Damage Caused by γ-Irradiation in Normal Human Bronchial Epithelial Cells In Vitro. Antioxidants 2022, 11, 1407. https://doi.org/10.3390/antiox11071407
Merlin JPJ, Mathavarajah S, Dellaire G, Murphy KPJ, Rupasinghe HPV. A Dietary Antioxidant Formulation Ameliorates DNA Damage Caused by γ-Irradiation in Normal Human Bronchial Epithelial Cells In Vitro. Antioxidants. 2022; 11(7):1407. https://doi.org/10.3390/antiox11071407
Chicago/Turabian StyleMerlin, J. P. Jose, Sabateeshan Mathavarajah, Graham Dellaire, Kieran P. J. Murphy, and H. P. Vasantha Rupasinghe. 2022. "A Dietary Antioxidant Formulation Ameliorates DNA Damage Caused by γ-Irradiation in Normal Human Bronchial Epithelial Cells In Vitro" Antioxidants 11, no. 7: 1407. https://doi.org/10.3390/antiox11071407