
Antioxidant and Neuroprotective Effects of Paeonol against Oxidative Stress and Altered Carrier-Mediated Transport System on NSC-34 Cell Lines

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Radioisotope Compound and Chemical Reagents
2.2. NSC-34 Cell Culture
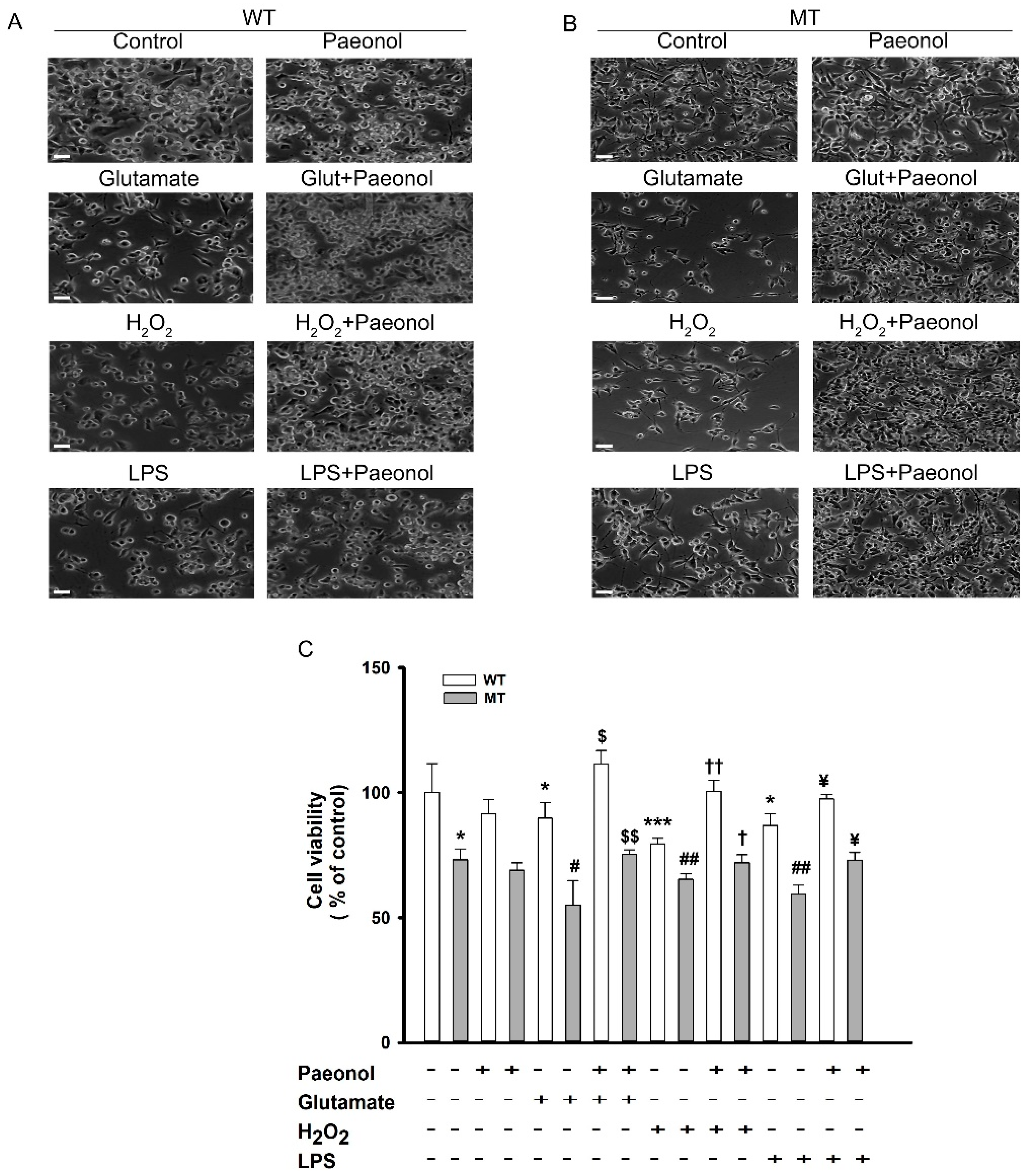
2.3. Cell Viability Assay
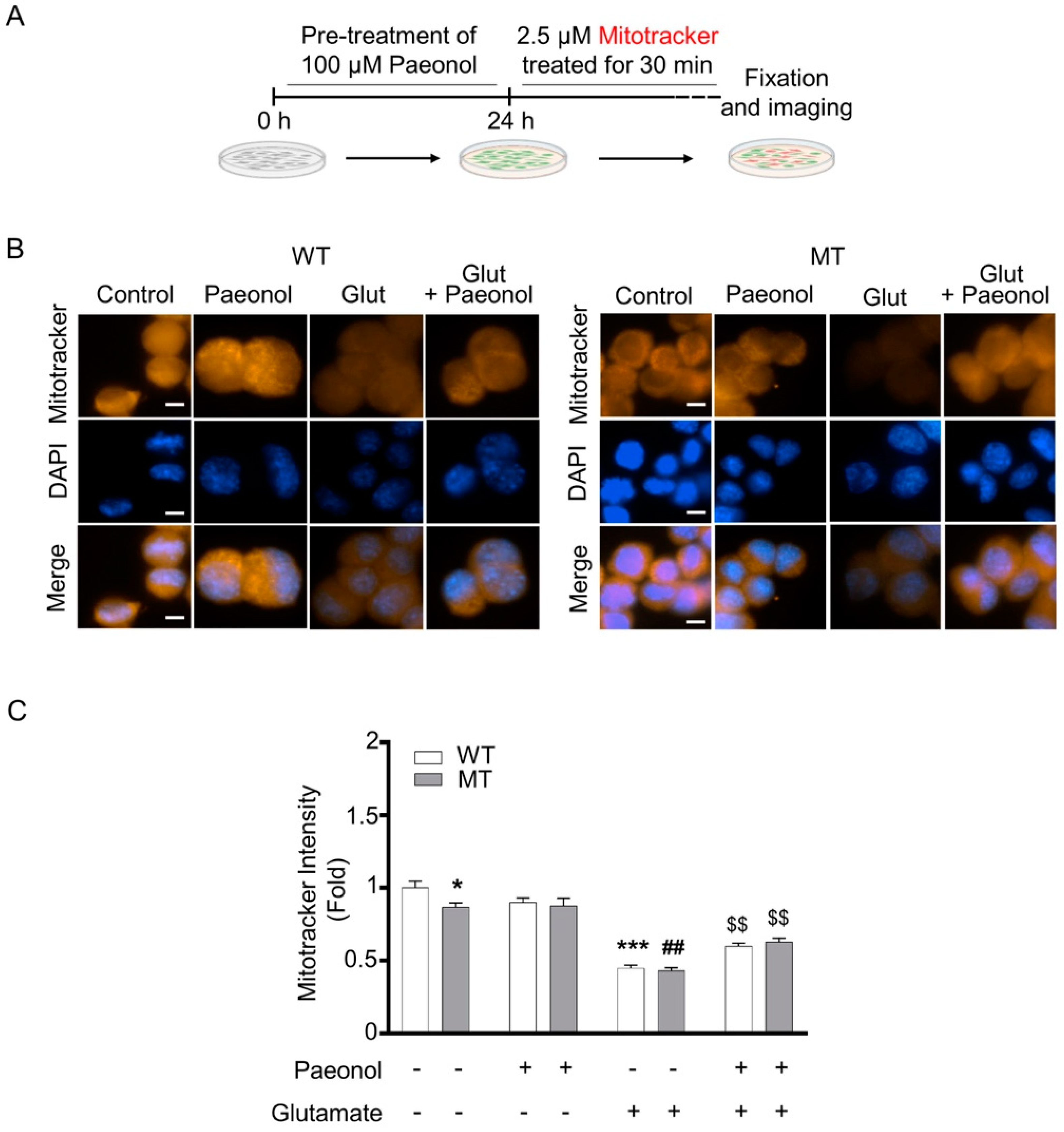
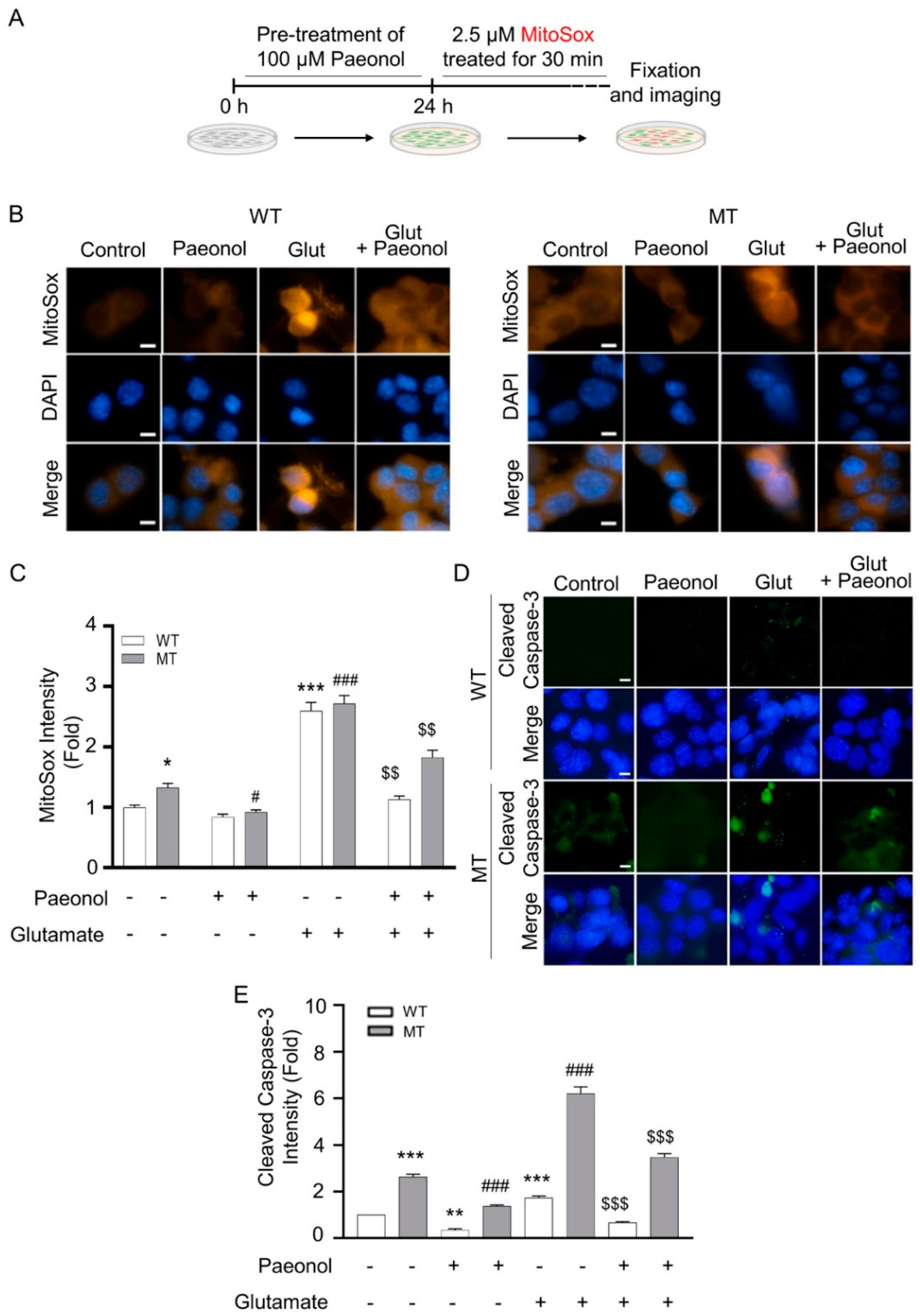
2.4. Measurement of Mitochondrial Membrane Potential (ψ) and Oxidative Stress, and Immunocytochemistry
2.5. In Vitro Uptake Study of [3H]paeonol in Motor Neuron like Cell Line
2.6. Estimation of Kinetic Parameters of [3H]paeonol Uptake in ALS Model Cell Lines
2.7. Small Interfering RNA (siRNA) Transfection Study in NSC-34 Cell Lines
2.8. Quantitative Reverse Transcription-Polymerase Chain Reaction
2.9. ALS Mouse and Immunohistochemistry
2.10. Statistical Analysis
3. Results
3.1. Cytotoxicity Induced by Pro-Inflammatory Cytokines and Neuroprotective Effect of Paeonol in ALS Model Cell Line
3.2. Paeonol Preventing Glutamate-Induced Decrease of Mitochondria Membrane Potential (ψ) and Elevation of Mitochondrial Oxidative Stress in ALS Cell Line Model
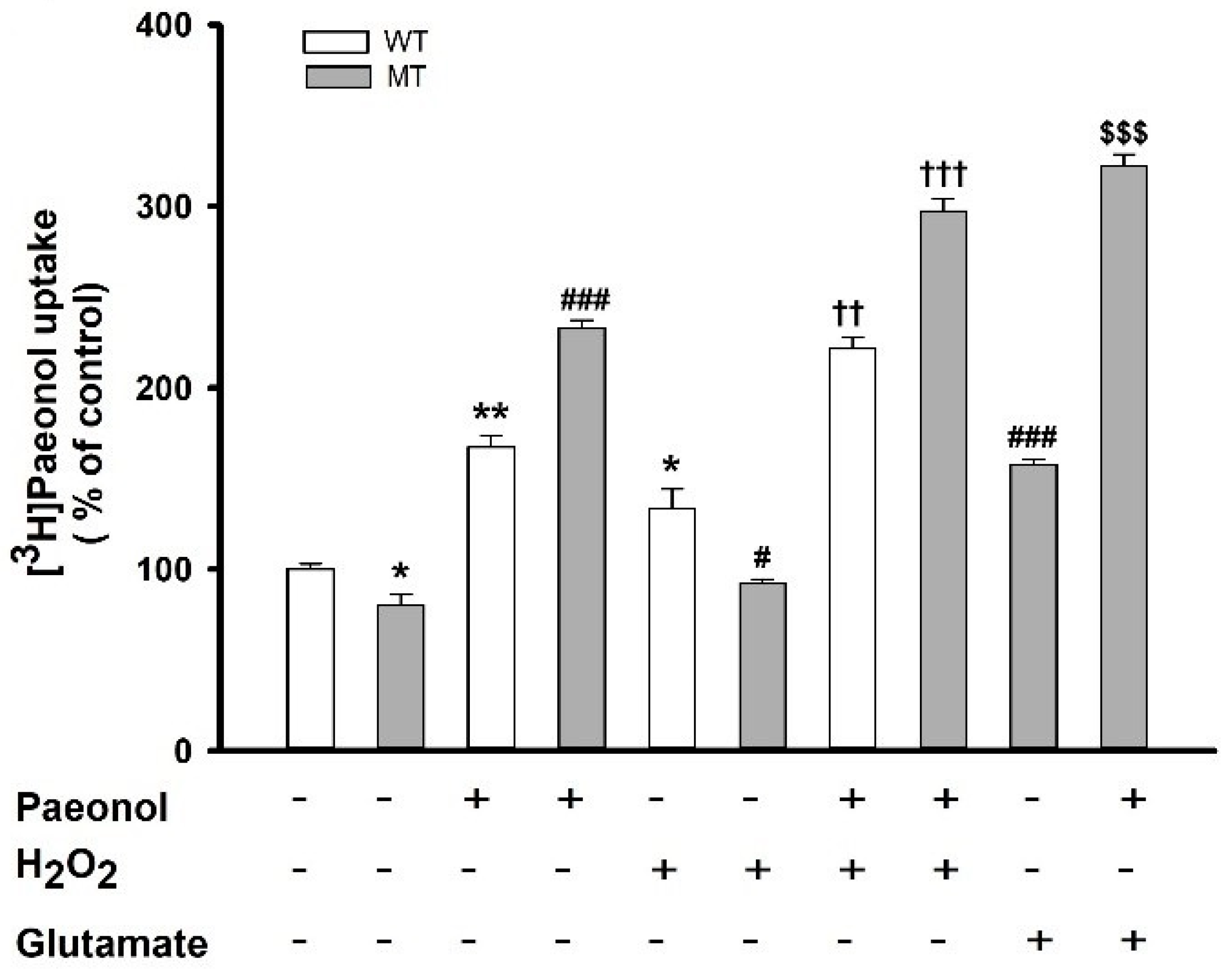
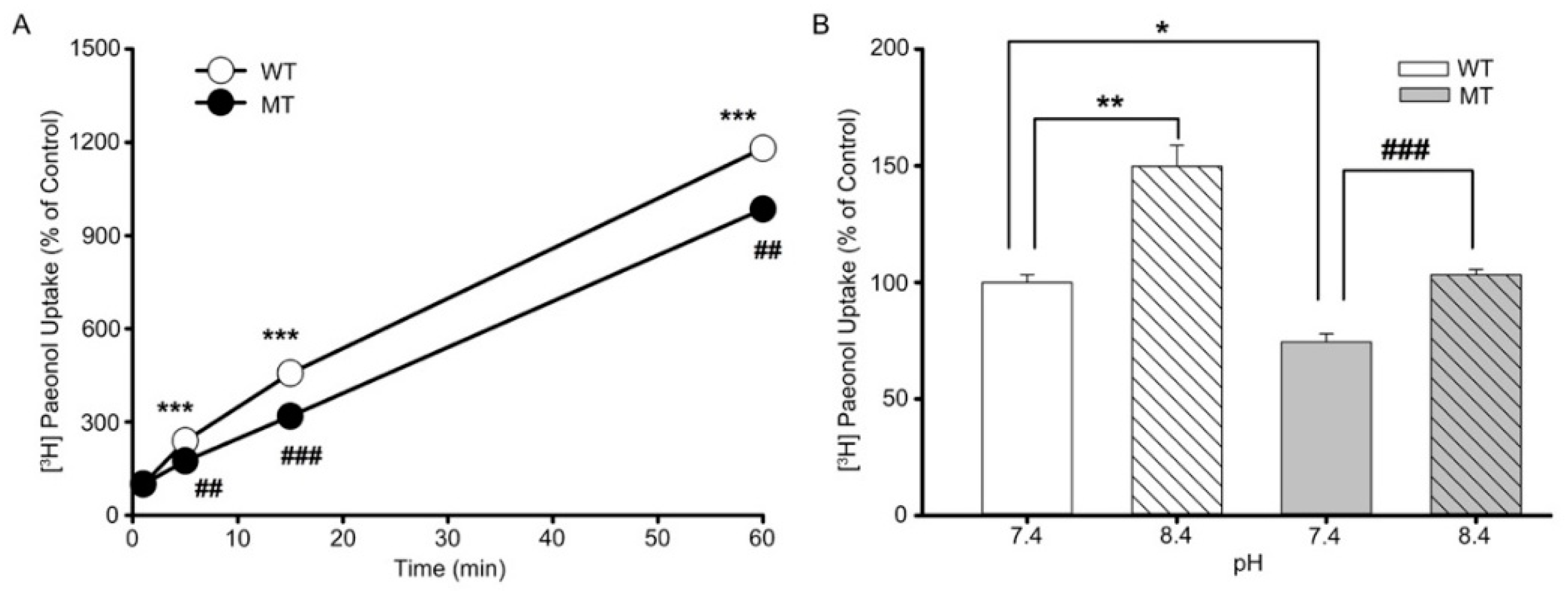
3.3. Characteristics of [3H]paeonol Uptake by ALS Model Cell Lines
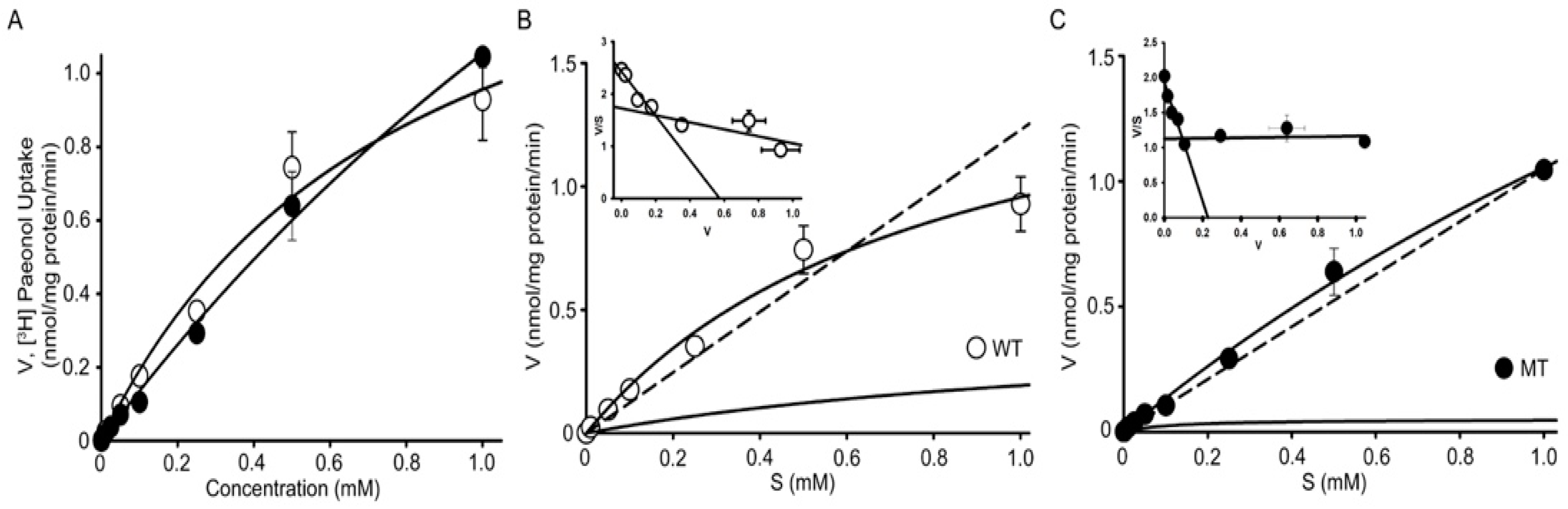
3.4. Analysis of Kinetic Parameters of [3H]paeonol Uptake by NSC-34 Cell Lines
3.5. Effects of Pharmacological Drugs on the Paeonol Transport in ALS Model Cell Lines
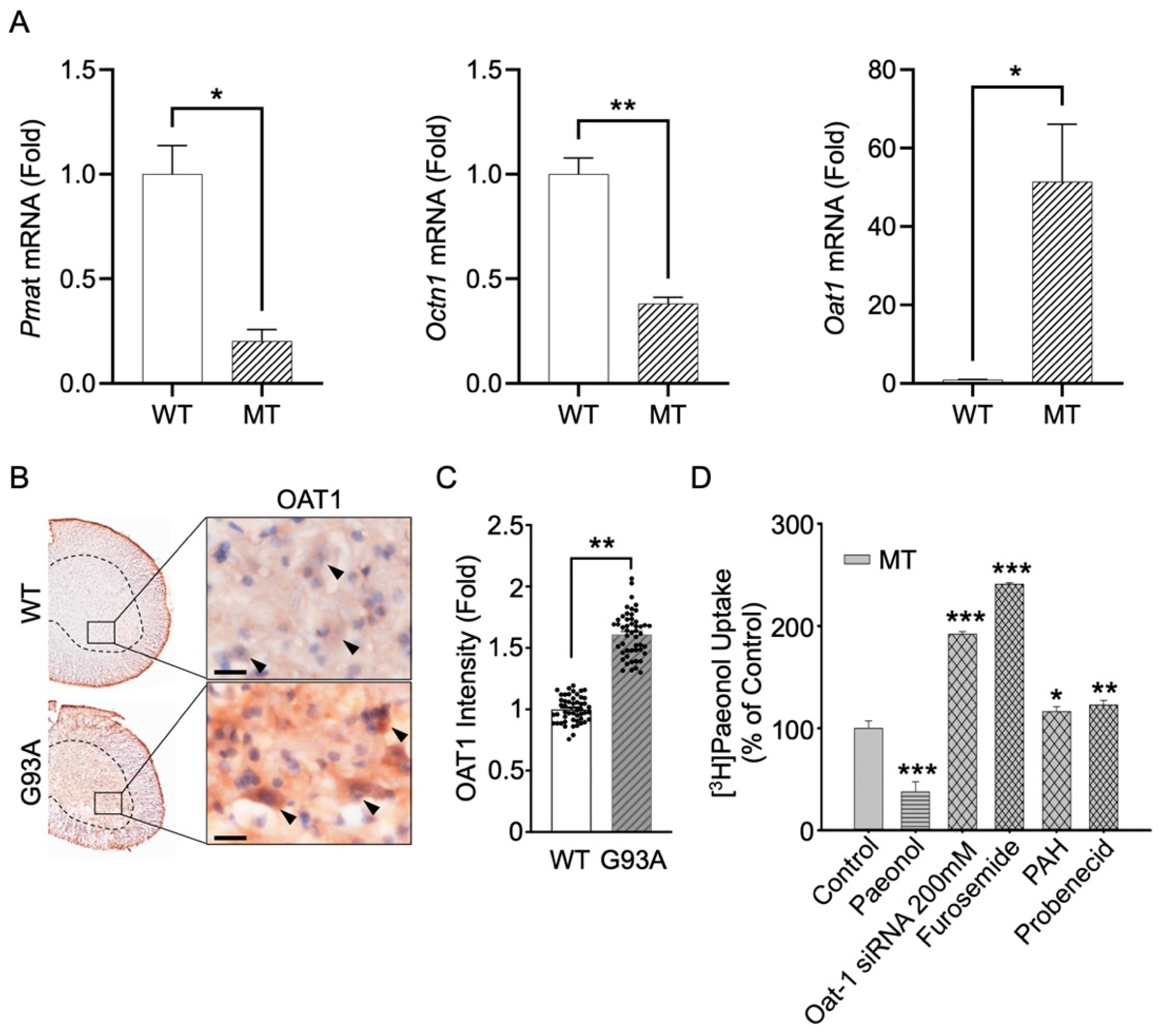
3.6. Knockdown by siRNA Transfection and mRNA Expression of Pmat, Octn1, and Oat1 in Cell and Animal Models of ALS
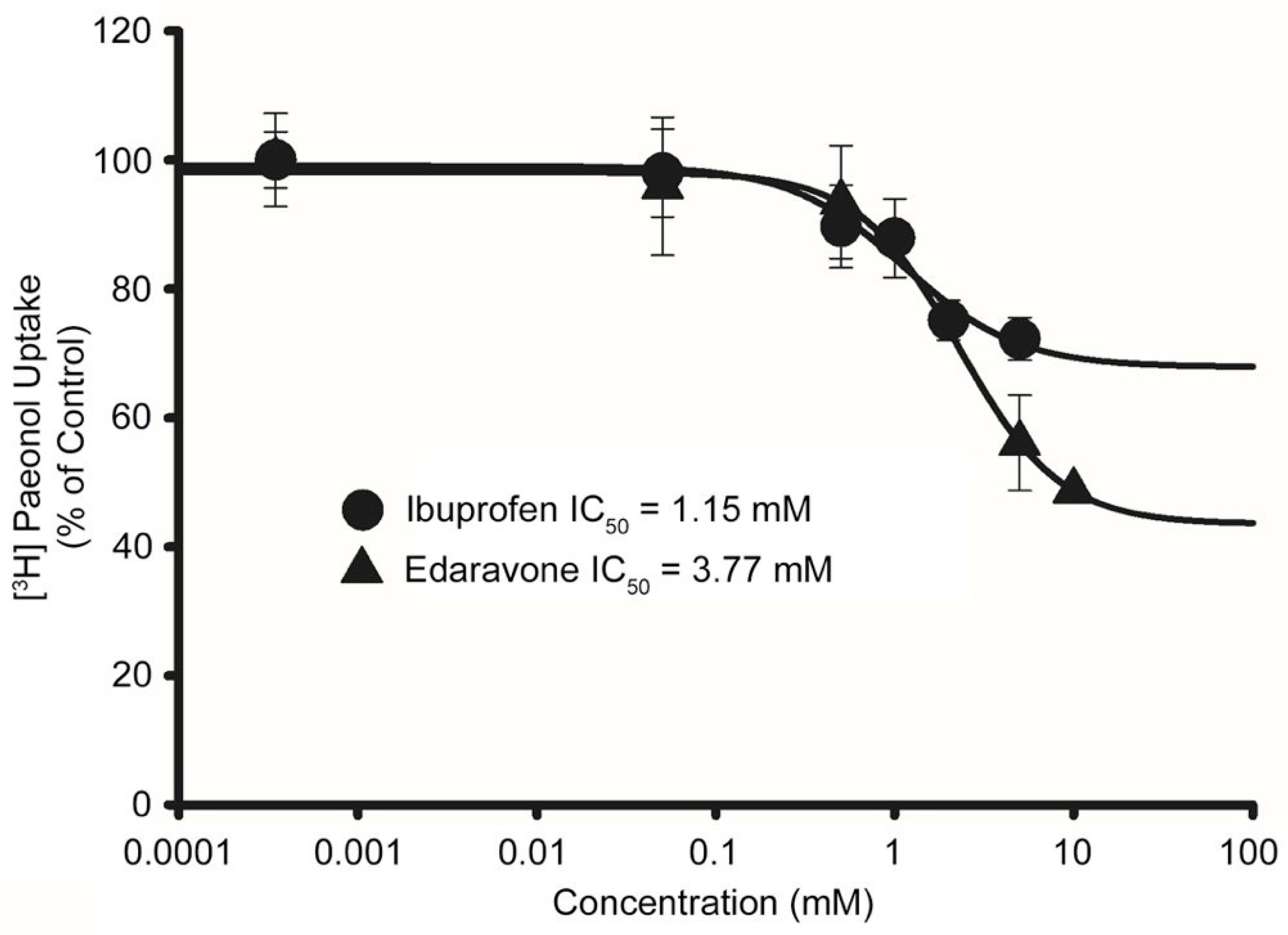
3.7. Determination of Minimal Inhibitory Concentration (IC50) of Drugs in Disease Model Cell Lines
4. Discussion
4.1. Paeonol Restores Motor Neuronal Viability against Multiple Stresses
4.2. Paeonol Modulates Mitochondria Function in ALS Cell Line
4.3. Paeonol Uptake Is Differentially Regulated between WT and MT ALS Cell Line
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, J.; Ryu, H.; Keum, G.; Yoon, Y.J.; Kowall, N.W.; Ryu, H. Therapeutic Targeting of Epigenetic Components in Amyotrophic Lateral Sclerosis (ALS). Curr. Med. Chem. 2014, 21, 3576–3582. [Google Scholar] [CrossRef] [PubMed]
- Tortarolo, M.; Lo Coco, D.; Veglianese, P.; Vallarola, A.; Giordana, M.T.; Marcon, G.; Beghi, E.; Poloni, M.; Strong, M.J.; Iyer, A.M.; et al. Amyotrophic Lateral Sclerosis, a Multisystem Pathology: Insights into the Role of TNF α. Mediators Inflamm. 2017, 2017, 2985051 . [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurney, M.E.; Pu, H.; Chiu, A.Y.; Dal Canto, M.C.; Polchow, C.Y.; Alexander, D.D.; Caliendo, J.; Hentati, A.; Kwon, Y.W.; Deng, H.X.; et al. Motor neuron degeneration in mice that express a human Cu, Zn superoxide dismutase mutation. Science 1994, 264, 1772–1775. [Google Scholar] [CrossRef]
- Lee, J.; Kannagi, M.; Ferrante, R.J.; Kowall, N.W.; Ryu, H. Activation of Ets-2 by oxidative stress induces Bcl-xL expression and accounts for glial survival in amyotrophic lateral sclerosis. FASEB J. 2009, 23, 1739–1749. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Ryu, H.; Kowall, N.W. Differential regulation of neuronal and inducible nitric oxide synthase (NOS) in the spinal cord of mutant SOD1 (G93A) ALS mice. Biochem. Biophys. Res. Commun. 2009, 387, 202–206. [Google Scholar] [CrossRef] [Green Version]
- Ryu, H.; Smith, K.; Camelo, S.I.; Carreras, I.; Lee, J.; Iglesias, A.H.; Dangond, F.; Cormier, K.A.; Cudkowicz, M.E.; Brown, R.H.; et al. Sodium phenylbutyrate prolongs survival and regulates expression of anti-apoptotic genes in transgenic amyotrophic lateral sclerosis mice. J. Neurochem. 2005, 93, 1087–1098. [Google Scholar] [CrossRef]
- Xu, X.; Shen, D.; Gao, Y.; Zhou, Q.; Ni, Y.; Meng, H.; Shi, H.; Le, W.; Chen, S.; Chen, S. A perspective on therapies for amyotrophic lateral sclerosis: Can disease progression be curbed? Transl. Neurodegener. 2021, 10, 1–18. [Google Scholar] [CrossRef]
- Jaiswal, M.K. Riluzole and edaravone: A tale of two amyotrophic lateral sclerosis drugs. Med. Res. Rev. 2019, 39, 733–748. [Google Scholar] [CrossRef]
- Zhang, X.; Zhai, Y.; Yuan, J.; Hu, Y. New insights into Paeoniaceae used as medicinal plants in China. Sci. Rep. 2019, 9, 18469. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Yu, Z.; Li, X.; Zhang, X.; Wang, S.; Yang, S.; Hu, L.; Liu, L. Paeonol for the treatment of atherosclerotic cardiovascular disease: A pharmacological and cechanistic overview. Front. Cardiovasc. Med. 2021, 8, 690116. [Google Scholar] [CrossRef]
- Zhang, M.; Li, Y.; Zhu, T. The theoretical investigation on properties of paeonol and its isomers. Mol. Phys. 2021, 119, e1925363. [Google Scholar] [CrossRef]
- Zhou, H.M.; Sun, Q.X.; Cheng, Y. Paeonol enhances the sensitivity of human ovarian cancer cells to radiotherapy-induced apoptosis due to downregulation of the phosphatidylinositol-3-kinase/Akt/phosphatase and tensin homolog pathway and inhibition of vascular endothelial growth factor. Exp. Ther. Med. 2017, 14, 3213–3220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gyawali, A.; Kim, M.H.; Kang, Y.S. A novel organic cation transporter involved in paeonol transport across the inner blood-retinal barrier and changes in uptake in high glucose conditions. Exp. Eye Res. 2021, 202, 108387. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.M. Neuroprotective therapeutics for Alzheimers disease: Progress and prospects. Trends Pharmacol. Sci. 2011, 32, 141–147. [Google Scholar] [CrossRef]
- Zhang, L.; Li, D.; Liu, L. Paeonol: Pharmacological effects and mechanisms of action. Int. Immunopharmacol. 2019, 72, 413–421. [Google Scholar] [CrossRef]
- Nam, K.N.; Woo, B.C.; Moon, S.K.; Park, S.U.; Park, J.Y.; Hwang, J.W.; Bae, H.S.; Ko, C.N.; Lee, E.H. Paeonol attenuates inflammation-mediated neurotoxicity and microglial activation. Neural Regen. Res. 2013, 8, 1637–1643. [Google Scholar] [CrossRef]
- Shi, X.; Chen, Y.H.; Liu, H.; Qu, H.D. Therapeutic effects of paeonol on methyl-4-phenyl-1,2,3,6-tetrahydropyridine/probenecid-induced Parkinson’s disease in mice. Mol. Med. Rep. 2016, 14, 2397–2404. [Google Scholar] [CrossRef] [Green Version]
- Gyawali, A.; Latif, S.; Choi, S.-H.; Hyeon, S.J.; Ryu, H.; Kang, Y.-S. Monocarboxylate transporter functions and neuroprotective effects of valproic acid in experimental models of amyotrophic lateral sclerosis. J. Biomed. Sci. 2022, 29, 2. [Google Scholar] [CrossRef]
- Gyawali, A.; Gautam, S.; Hyeon, S.J.; Ryu, H.; Kang, Y.S. L-Citrulline level and transporter activity are altered in experimental models of amyotrophic lateral sclerosis. Mol. Neurobiol. 2021, 58, 647–657. [Google Scholar] [CrossRef]
- Latif, S.; Kang, Y. Change in cationic amino acid transport system and effect of lysine pretreatment on inflammatory state in amyotrophic lateral sclerosis cell model. Biomol. Ther. 2021, 29, 498–505. [Google Scholar] [CrossRef]
- Gyawali, A.; Kang, Y.S. Pretreatment effect of inflammatory stimuli and characteristics of tryptophan transport on brain capillary endothelial (Tr-BBB) and motor neuron like (NSC-34) cell lines. Biomedicines 2021, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Gyawali, A.; Kang, Y.S. Transport alteration of 4-phenyl butyric acid mediated by a sodium- and proton-coupled monocarboxylic acid transporter system in ALS model cell lines (NSC-34) under inflammatory states. J. Pharm. Sci. 2021, 110, 1374–1384. [Google Scholar] [CrossRef] [PubMed]
- Gyawali, A.; Hyeon, S.J.; Ryu, H.; Kang, Y.S. The alteration of l-carnitine transport and pretreatment effect under glutamate cytotoxicity on motor neuron-like NSC-34 lines. Pharmaceutics 2021, 13, 551. [Google Scholar] [CrossRef] [PubMed]
- Tovar-Y-Romo, L.B.; Santa-Cruz, L.D.; Tapia, R. Experimental models for the study of neurodegeneration in amyotrophic lateral sclerosis. Mol. Neurodegener. 2009, 4, 31. [Google Scholar] [CrossRef] [Green Version]
- Himaya, S.W.A.; Ryu, B.M.; Qian, Z.J.; Kim, S.K. Paeonol from Hippocampus kuda Bleeler suppressed the neuro-inflammatory responses in vitro via NF-κB and MAPK signaling pathways. Toxicol. Vitr. 2012, 26, 878–887. [Google Scholar] [CrossRef]
- Hosaka, T.; Tsuji, H.; Tamaoka, A. Biomolecular modifications linked to oxidative stress in amyotrophic lateral sclerosis: Determining promising biomarkers related to oxidative stress. Processes 2021, 9, 1667. [Google Scholar] [CrossRef]
- Sasaki, S.; Horie, Y.; Iwata, M. Mitochondrial alterations in dorsal root ganglion cells in sporadic amyotrophic lateral sclerosis. Acta Neuropathol. 2007, 114, 633–639. [Google Scholar] [CrossRef]
- Dunlop, R.A.; Rodgers, K.J.; Dean, R.T. Recent developments in the intracellular degradation of oxidized proteins. Free Radic. Biol. Med. 2002, 33, 894–906. [Google Scholar] [CrossRef]
- Adki, K.M.; Kulkarni, Y.A. Chemistry, pharmacokinetics, pharmacology and recent novel drug delivery systems of paeonol. Life Sci. 2020, 250, 117544. [Google Scholar] [CrossRef]
- Gyawali, A.; Krol, S.; Kang, Y.S. Involvement of a novel organic cation transporter in paeonol transport across the blood-brain barrier. Biomol. Ther. 2019, 27, 290–301. [Google Scholar] [CrossRef]
- Burckhardt, G. Drug transport by Organic Anion Transporters (OATs). Pharmacol. Ther. 2012, 136, 106–130. [Google Scholar] [CrossRef]
- Sweet, D.H.; Miller, D.S.; Pritchard, J.B.; Fujiwara, Y.; Beier, D.R.; Nigam, S.K. Impaired organic anion transport in kidney and choroid plexus of organic anion transporter 3 (Oat3 (Slc22a8)) knockout mice. J. Biol. Chem. 2002, 277, 26934–26943. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Ryu, H.; Kowall, N.W. Motor neuronal protection by l-arginine prolongs survival of mutant SOD1 (G93A) ALS mice. Biochem. Biophys. Res. Commun. 2009, 384, 524–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cashman, N.R.; Durham, H.D.; Blusztajn, J.K.; Oda, K.; Tabira, T.; Shaw, I.T.; Dahrouge, S.; Antel, J.P. Neuroblastoma × spinal cord (NSC) hybrid cell lines resemble developing motor neurons. Dev. Dyn. 1992, 194, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Hyeon, S.J.; Park, J.; Yoo, J.; Kim, S.H.; Hwang, Y.J.; Kim, S.C.; Liu, T.; Shim, H.S.; Kim, Y.; Cho, Y.; et al. Dysfunction of X-linked inhibitor of apoptosis protein (XIAP) triggers neuropathological processes via altered p53 activity in Huntington’s disease. Prog. Neurobiol. 2021, 204, 102110. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.Y.; Kim, Y.; Ryu, H.; Kang, Y.S. The alteration of serine transporter activity in a cell line model of amyotrophic lateral sclerosis (ALS). Biochem. Biophys. Res. Commun. 2017, 483, 135–141. [Google Scholar] [CrossRef]
- Lee, N.Y.; Choi, H.M.; Kang, Y.S. Choline Transport via Choline Transporter-like Protein 1 in Conditionally Immortalized Rat Syncytiotrophoblast Cell Lines TR-TBT. Placenta 2009, 30, 368–374. [Google Scholar] [CrossRef]
- Van Den Bosch, L.; Van Damme, P.; Bogaert, E.; Robberecht, W. The role of excitotoxicity in the pathogenesis of amyotrophic lateral sclerosis. Biochim. Biophys. Acta-Mol. Basis Dis. 2006, 1762, 1068–1082. [Google Scholar] [CrossRef] [Green Version]
- Pandya, R.S.; Zhu, H.; Li, W.; Bowser, R.; Friedlander, R.M.; Wang, X. Therapeutic neuroprotective agents for amyotrophic lateral sclerosis. Cell. Mol. Life Sci. 2013, 70, 4729–4745. [Google Scholar] [CrossRef]
- Liu, J.; Feng, L.; Ma, D.; Zhang, M.; Gu, J.; Wang, S.; Fu, Q.; Song, Y.; Lan, Z.; Qu, R.; et al. Neuroprotective effect of paeonol on cognition deficits of diabetic encephalopathy in streptozotocin-induced diabetic rat. Neurosci. Lett. 2013, 549, 63–68. [Google Scholar] [CrossRef]
- Lee, N.Y.; Kang, Y.S. Taurine protects glutamate neurotoxicity in motor neuron cells. Adv. Exp. Med. Biol. 2017, 975, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhu, G.; Yang, S.; Wang, X.; Cheng, H.; Wang, F.; Li, X.; Li, Q. Paeonol prevents excitotoxicity in rat pheochromocytoma PC12 cells via downregulation of ERK activation and inhibition of apoptosis. Planta Med. 2011, 77, 1695–1701. [Google Scholar] [CrossRef]
- Tseng, Y.T.; Hsu, Y.Y.; Shih, Y.T.; Lo, Y.C. Paeonol attenuates microglia-mediated inflammation and oxidative stress-induced neurotoxicity in rat primary microglia and cortical neurons. Shock 2012, 37, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Culmsee, C.; Zai, F.Y. Apoptotic and antiapoptotic mechanisms in stroke. Cell Tissue Res. 2000, 301, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Zhang, Q. Paeonol protects melanocytes against hydrogen peroxide-induced oxidative stress through activation of Nrf2 signaling pathway. Drug Dev. Res. 2021, 82, 861–869. [Google Scholar] [CrossRef]
- Barber, S.C.; Shaw, P.J. Oxidative stress in ALS: Key role in motor neuron injury and therapeutic target. Free Radic. Biol. Med. 2010, 48, 629–641. [Google Scholar] [CrossRef]
- Muyderman, H.; Chen, T. Mitochondrial dysfunction in amyotrophic lateral sclerosis-A valid pharmacological target? Br. J. Pharmacol. 2014, 171, 2191–2205. [Google Scholar] [CrossRef]
- Lin, C.; Lin, H.Y.; Chen, J.H.; Tseng, W.P.; Ko, P.Y.; Liu, Y.S.; Yeh, W.L.; Lu, D.Y. Effects of paeonol on anti-neuroinflammatory responses in microglial cells. Int. J. Mol. Sci. 2015, 16, 8844–8860. [Google Scholar] [CrossRef] [Green Version]
- Carriedo, S.G.; Sensi, S.L.; Yin, H.Z.; Weiss, J.H. AMPA exposures induce mitochondrial Ca2+ overload and ROS generation in spinal motor neurons in vitro. J. Neurosci. 2000, 20, 240–250. [Google Scholar] [CrossRef] [Green Version]
- Salomon, J.J.; Gausterer, J.C.; Selo, M.A.; Hosoya, K.I.; Huwer, H.; Schneider-Daum, N.; Lehr, C.M.; Ehrhardt, C. OCTN2-mediated acetyl-l-carnitine transport in human pulmonary epithelial cells in vitro. Pharmaceutics 2019, 11, 396. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Chen, S.; Lu, K.; Wang, F.; Deng, J.; Xu, Z.; Wang, X.; Zhou, Q.; Le, W.; Zhao, Y. Verapamil ameliorates motor neuron degeneration and improves lifespan in the SOD1G93A mouse model of als by enhancing autophagic flux. Aging Dis. 2019, 10, 1159–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.Y.; Chen, J.F.; Chen, Y.; Zhuang, Y.H.; Sun, M.; Behnisch, T. The neurotoxin 1-methyl-4-phenylpyridinium (MPP+) alters hippocampal excitatory synaptic transmission by modulation of the GABAergic system. Front. Cell. Neurosci. 2015, 9, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuuchi, H.; Katsura, T.; Ashida, K.; Hashimoto, Y.; Inui, K.I. Diphenhydramine transport by pH-dependent tertiary amine transport system in Caco-2 cells. Am. J. Physiol.-Gastrointest. Liver Physiol. 2000, 278, 563–569. [Google Scholar] [CrossRef]
- Boxberger, K.H.; Hagenbuch, B.; Lampe, J.N. Common drugs inhibit human organic cation transporter 1 (oct1)-mediated neurotransmitter uptake. Drug Metab. Dispos. 2014, 42, 990–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- André, P.; Debray, M.; Scherrmann, J.M.; Cisternino, S. Clonidine transport at the mouse blood-brain barrier by a new H antiporter that interacts with addictive drugs. J. Cereb. Blood Flow Metab. 2009, 29, 1293–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-H.; Jung, S.-J.; Um, S.-Y.; Na, M.-A.; Choi, M.-J.; Chung, M.-W.; Oh, H.-Y. Effect of Cimetidine on the Transport of Quinolone Antibiotics in Caco-2 Cell monolayers. Biomol. Ther. 2007, 15, 102–107. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, N.; Takahashi, T.; Iwase, Y.; Kusuhara, H.; Niwa, T.; Sugiyama, Y. Human organic anion transporters 1 (hOAT1/SLC22A6) and 3 (hOAT3/SLC22A8) transport edaravone (MCI-186; 3-methyl-1-phenyl-2-pyrazolin-5-one) and its sulfate conjugate. Drug Metab. Dispos. 2007, 35, 1429–1434. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; George, R.L.; Huang, W.; Wang, H.; Conway, S.J.; Leibach, F.H.; Ganapathy, V. Structural and functional characteristics and tissue distribution pattern of rat OCTN1, an organic cation transporter, cloned from placenta. Biochim. Biophys. Acta-Biomembr. 2000, 1466, 315–327. [Google Scholar] [CrossRef] [Green Version]
- Yonezawa, A.; Inui, K.I. Importance of the multidrug and toxin extrusion MATE/SLC47A family to pharmacokinetics, pharmacodynamics/toxicodynamics and pharmacogenomics. Br. J. Pharmacol. 2011, 164, 1817–1825. [Google Scholar] [CrossRef] [Green Version]
- Wang, J. The plasma membrane monoamine transporter (PMAT): Structure, function, and role in organic cation disposition. Clin. Pharmacol. Ther. 2016, 100, 489–499. [Google Scholar] [CrossRef] [Green Version]
- Ashrafian, H.; Bogle, R.G. The organic anion transporter family: From physiology to ontogeny and the clinic. J. Intensive Care Soc. 2004, 5, 108–111. [Google Scholar] [CrossRef]
- Lee, W.; Ha, J.M.; Sugiyama, Y. Post-translational regulation of the major drug transporters in the families of organic anion transporters and organic anion-transporting polypeptides. J. Biol. Chem. 2020, 295, 17349–17364. [Google Scholar] [CrossRef] [PubMed]
- Evers, R.; Piquette-Miller, M.; Polli, J.W.; Russel, F.G.M.; Sprowl, J.A.; Tohyama, K.; Ware, J.A.; de Wildt, S.N.; Xie, W.; Brouwer, K.L.R. Disease-associated changes in drug transporters may impact the pharmacokinetics and/or toxicity of drugs: A white paper from the international transporter consortium. Clin. Pharmacol. Ther. 2018, 104, 900–915. [Google Scholar] [CrossRef] [PubMed]
- Wolff, N.A.; Thies, K.; Kuhnke, N.; Reid, G.; Friedrich, B.; Lang, F.; Burckhardt, G. Protein kinase C activation downregulates human organic anion transporter 1-mediated transport through carrier internalization. J. Am. Soc. Nephrol. 2003, 14, 1959–1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauvant, C.; Holzinger, H.; Gekle, M. Short-term regulation of basolateral organic anion uptake in proximal tubular opossum kidney cells: Prostaglandin E2 acts via receptor-mediated activation of protein kinase A. J. Am. Soc. Nephrol. 2003, 14, 3017–3026. [Google Scholar] [CrossRef] [Green Version]
- Nozaki, Y.; Kusuhara, H.; Kondo, T.; Hasegawa, M.; Shiroyanagi, Y.; Nakazawa, H.; Okano, T.; Sugiyama, Y. Characterization of the uptake of organic anion transporter (OAT) 1 and OAT3 substrates by human kidney slices. J. Pharmacol. Exp. Ther. 2007, 321, 362–369. [Google Scholar] [CrossRef]
- Hagos, F.T.; Daood, M.J.; Ocque, J.A.; Nolin, T.D.; Bayir, H.; Poloyac, S.M.; Kochanek, P.M.; Clark, R.S.B.; Empey, P.E. Probenecid, an organic anion transporter 1 and 3 inhibitor, increases plasma and brain exposure of N-acetylcysteine. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Severin, M.J.; Hazelhoff, M.H.; Bulacio, R.P.; Mamprin, M.E.; Brandoni, A.; Torres, A.M. Impact of the induced organic anion transporter 1 (Oat1) renal expression by furosemide on the pharmacokinetics of organic anions. Nephrology 2017, 22, 642–648. [Google Scholar] [CrossRef]
- Ohtsuki, S.; Asaba, H.; Takanaga, H.; Deguchi, T.; Hosoya, K.I.; Otagiri, M.; Terasaki, T. Role of blood-brain barrier organic anion transporter 3 (OAT3) in the efflux of indoxyl sulfate, a uremic toxin: Its involvement in neurotransmitter metabolite clearance from the brain. J. Neurochem. 2002, 83, 57–66. [Google Scholar] [CrossRef]
- Sugiyama, D.; Kusuhara, H.; Shitara, Y.; Abe, T.; Meier, P.J.; Sekine, T.; Endou, H.; Suzuki, H.; Sugiyama, Y. Characterization of the efflux transport of 17β-estradiol-D-17β-glucuronide from the brain across the blood-brain barrier. J. Pharmacol. Exp. Ther. 2001, 298, 316–322. [Google Scholar]
- Leidgens, V.; Seliger, C.; Jachnik, B.; Welz, T.; Leukel, P.; Vollmann-Zwerenz, A.; Bogdahn, U.; Kreutz, M.; Grauer, O.M.; Hau, P. Ibuprofen and diclofenac restrict migration and proliferation of human glioma cells by distinct molecular mechanisms. PLoS ONE 2015, 10, e0140613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Tahara, M.; Todo, S. The novel antioxidant edaravone: From bench to bedside. Cardiovasc. Ther. 2008, 26, 101–114. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | WT | MT |
|---|---|---|
| Km1 (mM) | 0.011 ± 0.009 | 0.098 ± 0.006 |
| Km2 (mM) | 0.821 ± 0.344 | 5.35 ± 1.52 * |
| Vmax1 (nmol/mg protein/min) | 0.183 ± 0.019 | 0.161 ± 0.009 |
| Vmax2 (nmol/mg protein/min) | 1.82 ± 0.56 | 7.19 ± 1.50 ** |
| Pharmacological Compounds | Conc. (mM) | [3H]Paeonol Uptake (% of Control) | |
|---|---|---|---|
| WT (hSOD1WT) | MT (hSODG93A) | ||
| Control | 100 ± 3 | 100 ± 3 | |
| +Paeonol | 1 | 36.8 ± 0.8 *** | 35.4 ± 1.3 *** |
| +Edaravone | 1 | 62.1 ± 7.0 *** | 67.9 ± 5.4 *** |
| +Ibuprofen | 1 | -- | 78 ± 7 * |
| +L-Carnitine | 1 | 82.7 ± 3.6 ** | 73.9 ± 2.3 *** |
| +MPP+ | 1 | 63.3 ± 6.3 *** | 73.8 ± 2.7 ** |
| +Quinidine | 1 | 63.9 ± 8.4 * | 73.4 ± 4.8 * |
| +Verapamil | 1 | 75.2 ± 4.5 * | 73.1 ± 7.5 ** |
| +Diphenhydramine | 1 | 77.5 ± 6.0 ** | 75.3 ± 5.3 ** |
| +Clonidine | 1 | 72.6 ± 6.0 * | 83.7 ± 2.6 ** |
| +Tramadol | 1 | 82.9 ± 4.5 * | 80.5 ± 6.7 *** |
| +Cimetidine | 1 | 86.6 ± 2.3 * | 87.3 ± 4.2 * |
| +ALC | 1 | 73.4 ± 10.5 * | 82.0 ± 6.4 ** |
| +TEA | 1 | 90.7 ± 1.8 | 146 ± 6.6 ** |
| +PAH | 1 | 102 ± 10 | 116 ± 5 * |
| siRNA | [3H]Paeonol Uptake (% of Control) | |
|---|---|---|
| WT | MT | |
| Control | 100 ± 4 | 100 ± 4 |
| Octn1 | 105 ± 2 | 85.1± 10.4 ***,### |
| Octn2 | 104 ± 6 | 102 ± 3 |
| MATE | 94.9 ± 5.0 | 93.4 ± 4.8 |
| Pmat | 80.4 ± 8.5 * | 77.5 ± 10.1 * |
| Oct2 | 102 ± 4 | 107 ± 5 |
| SMCT1 | 100 ± 5 | 101 ± 1 |
| SMCT2 | 104 ± 9 | 103 ± 6 |
| Oat1 | 101 ± 2 | 193 ± 3 ***,### |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latif, S.; Choi, S.-H.; Gyawali, A.; Hyeon, S.J.; Kang, Y.-S.; Ryu, H. Antioxidant and Neuroprotective Effects of Paeonol against Oxidative Stress and Altered Carrier-Mediated Transport System on NSC-34 Cell Lines. Antioxidants 2022, 11, 1392. https://doi.org/10.3390/antiox11071392
Latif S, Choi S-H, Gyawali A, Hyeon SJ, Kang Y-S, Ryu H. Antioxidant and Neuroprotective Effects of Paeonol against Oxidative Stress and Altered Carrier-Mediated Transport System on NSC-34 Cell Lines. Antioxidants. 2022; 11(7):1392. https://doi.org/10.3390/antiox11071392
Chicago/Turabian StyleLatif, Sana, Seung-Hye Choi, Asmita Gyawali, Seung Jae Hyeon, Young-Sook Kang, and Hoon Ryu. 2022. "Antioxidant and Neuroprotective Effects of Paeonol against Oxidative Stress and Altered Carrier-Mediated Transport System on NSC-34 Cell Lines" Antioxidants 11, no. 7: 1392. https://doi.org/10.3390/antiox11071392