
Therapeutic Intervention in Cancer by Isoliquiritigenin from Licorice: A Natural Antioxidant and Redox Regulator


Abstract
:1. Introduction
2. Licorice (Glycyrrhiza Radix)
3. Isoliquiritigenin (ISL) and Its Biological Properties
3.1. Anti-Inflammatory Effects of ISL
3.2. Anti-Microbial Activity of ISL
3.3. Anti-Diabetic Effects of ISL
3.4. Immunomodulatory Property of ISL
3.5. Anti-Angiogenic Property of ISL
3.6. Potential Role of ISL in Cancer Development
3.7. ISL Modulates Antioxidant Enzymes and Phase II Enzymes in Neuroprotection and Chemoprevention
4. ISL and Pancreatic Cancer
4.1. ISL Promotes Apoptosis in Cancer Cells by Homeostatic Regulation of ROS
4.2. Antioxidant Role of Autophagy
4.3. ISL and Autophagy in PDAC Regulation
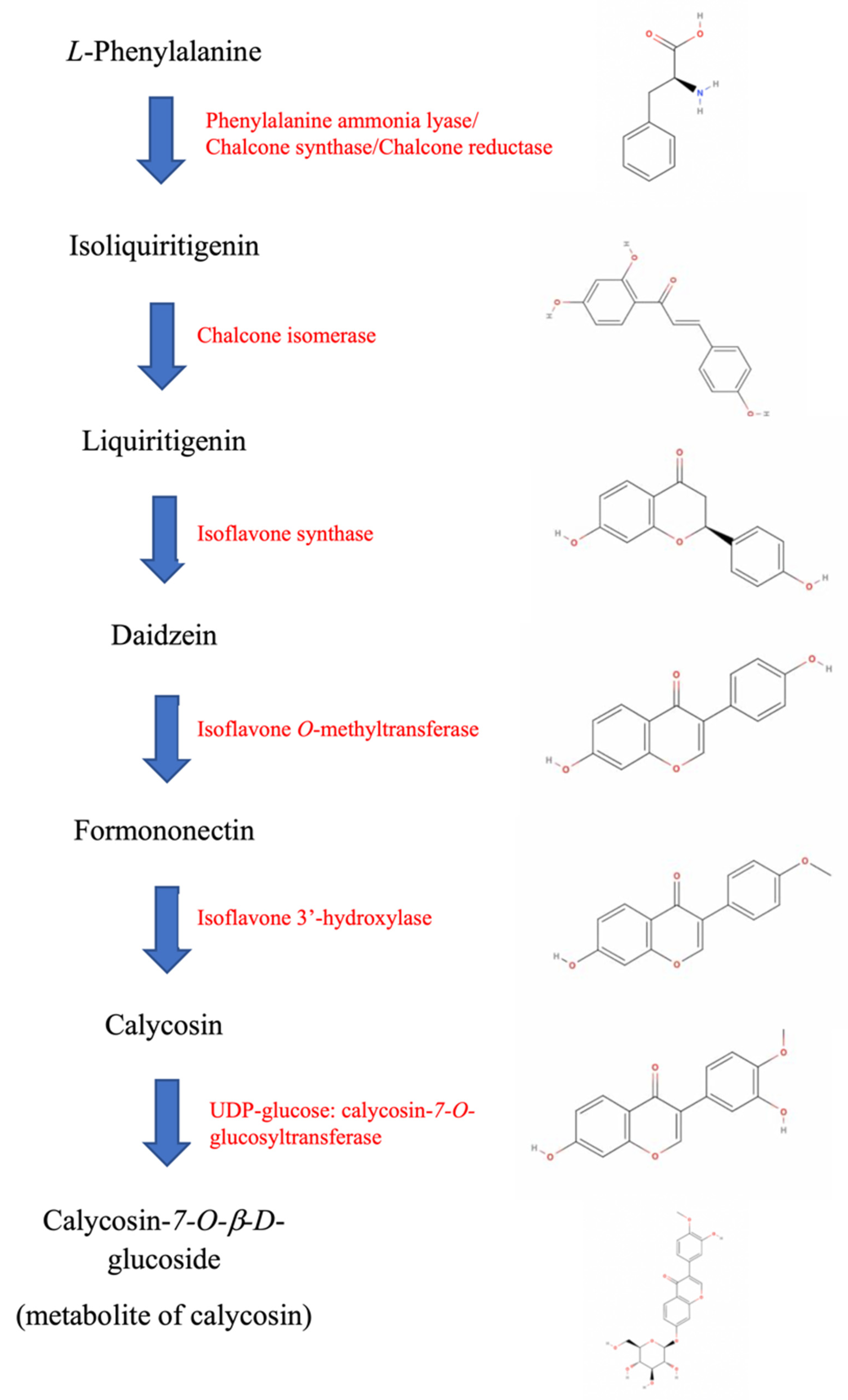
5. Insight from the ISL-Calysosin Isoflavonoid Biosynthetic Pathway
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McGuigan, A.; Kelly, P.; Turkington, R.C.; Jones, C.; Coleman, H.G.; McCain, R.S. Pancreatic cancer: A review of clinical diagnosis, epidemiology, treatment and outcomes. World J. Gastroenterol. 2018, 24, 4846–4861. [Google Scholar] [CrossRef] [PubMed]
- Ansari, D.; Tingstedt, B.; Andersson, B.; Holmquist, F.; Sturesson, C.; Williamsson, C.; Sasor, A.; Borg, D.; Bauden, M.; Andersson, R. Pancreatic cancer: Yesterday, today and tomorrow. Future Oncol. 2016, 12, 1929–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbassi, R.; Schmid, R.M. Evolving Treatment Paradigms for Pancreatic Cancer. Visc. Med. 2019, 35, 362–372. [Google Scholar] [CrossRef]
- Jia, D.; Park, J.H.; Jung, K.H.; Levine, H.; Kaipparettu, B.A. Elucidating the Metabolic plasticity of cancer: Mitochondrial reprogramming and hybrid metabolic states. Cells 2018, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Jiang, Y.; Meisenhelder, J.; Yang, W.; Hawke, D.H.; Zheng, Y.; Xia, Y.; Aldape, K.; He, J.; Hunter, T.; et al. Mitochondria-translocated pgk1 functions as a protein kinase to coordinate glycolysis and the TCA cycle in tumorigenesis. Mol. Cell 2016, 61, 705–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoncu, R.; Efeyan, A.; Sabatini, D.M. mTOR: From growth signal integration to cancer, diabetes and ageing. Nat. Rev. Mol. Cell Biol. 2011, 12, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Banim, P.J.R.; Luben, R.; McTaggart, A.; Welch, A.; Wareham, N.; Khaw, K.-T.; Hart, A.R. Dietary antioxidants and the aetiology of pancreatic cancer: A cohort study using data from food diaries and biomarkers. Gut 2012, 62, 1489–1496. [Google Scholar] [CrossRef]
- Han, X.; Li, J.; Brasky, T.M.; Xun, P.; Stevens, J.; White, E.; Gammon, M.D.; He, K. Antioxidant intake and pancreatic cancer risk: The VITamines And Lifestyle (VITAL) study. Cancer 2013, 119, 1314–1320. [Google Scholar] [CrossRef] [Green Version]
- Lucas, A.L.; Bosetti, C.; Boffetta, P.; Negri, E.; Tavani, A.; Serafini, M.; Polesel, J.; Serraino, D.; La Vecchia, C.; Rossi, M. Dietary total antioxidant capacity and pancreatic cancer risk: An Italian case-control study. Br. J. Cancer 2016, 115, 102–107. [Google Scholar] [CrossRef] [Green Version]
- DeNicola, G.M.; Karreth, F.A.; Humpton, T.J.; Gopinathan, A.; Wei, C.; Frese, K.; Mangal, D.; Yu, K.H.; Yeo, C.J.; Calhoun, E.S.; et al. Oncogene-induced Nrf2 transcription promotes ROS detoxification and tumorigenesis. Nature 2011, 475, 106–109. [Google Scholar] [CrossRef]
- Moloney, J.N.; Cotter, T.G. ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C. Flavonoid antioxidants. Curr. Med. Chem. 2001, 8, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Poljsak, B.; Šuput, D.; Milisav, I. Achieving the balance between ROS and antioxidants: When to use the synthetic antioxidants. Oxid. Med. Cell. Longev. 2013, 2013, 956792. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhu, P.; Zhang, X.; Ma, Y.; Li, W.; Chen, J.; Guo, H.; Bucala, R.; Zhuang, J.; Li, J. Natural antioxidant-isoliquiritigenin ameliorates contractile dysfunction of hypoxic cardiocytes via AMPK signaling pathway. Mediat. Inflam. 2013, 2013, 390890. [Google Scholar] [CrossRef]
- Chen, T.; Deng, S.; Lin, R. The inhibitory effect of isoliquiritigenin on the proliferation of human arterial smooth muscle cell. BMC Pharmacol. Toxicol. 2017, 18, 57. [Google Scholar] [CrossRef]
- Yao, D.; Shi, B.; Wang, S.; Bao, L.; Tan, M.; Shen, H.; Zhang, Z.; Pan, X.; Yang, Y.; Wu, Y.; et al. Isoliquiritigenin ameliorates ischemia-induced myocardial injury via modulating the Nrf2/HO-1 pathway in mice. Drug Design Develop. Ther. 2022, 16, 1273–1287. [Google Scholar] [CrossRef]
- Cao, L.; Li, H.; Yan, M.; Li, Z.; Gong, H.; Jiang, P.; Deng, Y.; Fang, P.; Zhang, B. The protective effects of isoliquiritigenin and glycyrrhetinic acid against triptolide-induced oxidative stress in HepG2 cells involve Nrf2 activation. Evid. Comp. Altern. Med. 2016, 2016, 8912184. [Google Scholar] [CrossRef]
- Liu, X.; Zhu, Q.; Zhang, M.; Yin, T.; Xu, R.; Xiao, W.; Wu, J.; Deng, B.; Gao, X.; Gong, W.; et al. Isoliquiritigenin ameliorates acute pancreatitis in mice via inhibition of oxidative stress and modulation of the Nrf2/HO-1 pathway. Oxid. Med. Cell. Longev. 2018, 2018, 7161592. [Google Scholar] [CrossRef]
- Chopra, R.N.; Chopra, I.C.; Handa, K.L.; Kapur, L.D. Indigenous Drugs of India; Dhur & Sons Private Ltd.: Kolkata, India, 1958. [Google Scholar]
- Karahan, F.; Avşar, C.; Ozyigit, I.I.; Berber, I. Antimicrobial and antioxidant activities of medicinal plant Glycyrrhiza glabra var. glandulifera from different habitats. Biotech. Biotech. Equip. 2016, 30, 797–804. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Shang, E.; Zhao, J.; Fan, X.; Duan, J.; Qian, D.; Tao, W.; Tang, Y. Data mining and frequency analysis for licorice as a “Two-Face” herb in Chinese formulae based on Chinese Formulae Database. Phytomedicine 2014, 21, 1281–1286. [Google Scholar] [CrossRef]
- Mukhopadhyay, M.; Panja, P. A novel process for extraction of natural sweetener from licorice (Glycyrrhiza glabra) roots. Sep. Purif. Technol. 2008, 63, 539–545. [Google Scholar] [CrossRef]
- Hayashi, H.; Sudo, H. Economic importance of licorice. Plant Biotechnol. 2009, 26, 101–104. [Google Scholar] [CrossRef] [Green Version]
- China Food and Drug Administration. Chinese Pharmacopoeia, 10th ed.; China Food and Drug Administration: Beijing, China, 2005.
- Pastorino, G.; Cornara, L.; Soares, S.; Rodrigues, F.; Oliveira, M.B.P.P. Liquorice (Glycyrrhiza glabra): A phytochemical and pharmacological review. Phytother. Res. 2018, 32, 2323–2339. [Google Scholar] [CrossRef] [PubMed]
- Rizzato, G.; Scalabrin, E.; Radaelli, M.; Capodaglio, G.; Piccolo, O. A new exploration of licorice metabolome. Food Chem. 2017, 221, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Hussein, M.A.; Pierce, S.; Martens, C.; Shahagadkar, P.; Munirathinam, G. Oncopreventive and oncotherapeutic potential of licorice triterpenoid compound glycyrrhizin and its derivatives: Molecular insights. Pharmacol. Res. 2022, 178, 106138. [Google Scholar] [CrossRef]
- Farese, R.V.; Biglieri, E.G.; Shackleton, C.H.; Irony, I.; Gomez-Fontes, R. Licorice-Induced Hypermineralocorticoidism. N. Engl. J. Med. 1991, 325, 1223–1227. [Google Scholar] [CrossRef]
- Han, Y.J.; Kang, B.; Yang, E.-J.; Choi, M.-K.; Song, I.-S. Simultaneous Determination and Pharmacokinetic Characterization of Glycyrrhizin, Isoliquiritigenin, Liquiritigenin, and Liquiritin in Rat Plasma Following Oral Administration of Glycyrrhizae Radix Extract. Molecules 2019, 24, 1816. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.Q.; Yang, R.; Yuan, B.C.; Liu, Y.; Liu, C.S. The antiviral and antimicrobial activities of licorice, a widely-used Chinese herb. Acta Pharm. Sin. B. 2015, 5, 310–315. [Google Scholar] [CrossRef] [Green Version]
- Asl, M.N.; Hosseinzadeh, H. Review of pharmacological effects of Glycyrrhiza Sp. and its bioactive compounds. Phytother. Res. 2008, 22, 709–724. [Google Scholar] [CrossRef]
- Kwon, H.M.; Choi, Y.J.; Choi, J.S.; Kang, S.W.; Bae, J.Y.; Kang, I.J.; Jun, J.G.; Lee, S.S.; Lim, S.S.; Kang, Y.H. Blockade of cytokine- induced endothelial cell adhesion molecule expression by licorice isoliquiritigenin through NF-kappa B signal disruption. Exp. Biol. Med. 2007, 232, 235–245. [Google Scholar]
- Hattori, T.; Sadakane, C.; Koseki, J.; Kase, Y.; Takeda, S. Saireito probably prevents mesangial cell proliferation in HIGA mice via PDGF-BB tyrosine kinase inhibition. Clin. Exp. Nephrol. 2007, 11, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhang, X.; Chen, X.; Li, Y.; Ke, Z.; Tang, T.; Chai, H.; Guo, A.M.; Chen, H.; Yang, J. Isoliquiritigenin, a flavonoid from licorice, blocks M2 macrophage polarization in colitis-associated tumorigenesis through downregulating PGE2 and IL-6. Toxicol. Appl. Pharmacol. 2014, 279, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sun, Y.N.; Yan, X.T.; Yang, S.Y.; Kim, S.; Lee, Y.M.; Koh, Y.-S.; Kim, Y.H. Flavonoids from Astragalus membranaceus and their inhibitory effects on LPS-stimulated pro-inflammatory cytokine production in bone marrow-derived dendritic cells. Arch. Pharmacal Res. 2013, 37, 186–192. [Google Scholar] [CrossRef]
- Yang, N.; Patil, S.; Zhuge, J.; Wen, M.C.; Bolleddula, J.; Doddaga, S.; Goldfarb, J.; Sampson, H.A.; Li, X.M. Glycyrrhiza uralensis flavonoids present in anti-asthma formula, ASHMITM, inhibit memory Th2 responses in vitro and in vivo. Phytother. Res. 2013, 27, 1381–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.K.; Papaemmanouil, A.; Bhowruth, V.; Bhatt, A.; Dover, L.G.; Besra, G.S. Flavonoid inhibitors as novel antimycobacterial agents targeting Rv0636, a putative dehydratase enzyme involved in Mycobacterium tuberculosis fatty acid synthase II. Microbiology 2007, 153, 3314–3322. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Mei, W.; Gong, M.; Zuo, W.; Bai, H.; Dai, H. Antibacterial Activity of the Flavonoids from Dalbergia odorifera on Ralstonia solanacearum. Molecules 2011, 16, 9775–9782. [Google Scholar] [CrossRef] [Green Version]
- Feldman, M.; Santos, J.; Grenier, D. Comparative Evaluation of Two Structurally Related Flavonoids, Isoliquiritigenin and Liquiritigenin, for Their Oral Infection Therapeutic Potential. J. Nat. Prod. 2011, 74, 1862–1867. [Google Scholar] [CrossRef]
- Oldoni, T.L.C.; Cabral, I.S.R.; Regitano d’Arce, M.A.B.; Rosalen, P.L.; Ikegaki, M.; Nascimento, A.M.; Alencar, S.M. Isolation and analysis of bioactive isoflavonoids and chalone from a new type of Brazilian propolis. Sep. Purif. Technol. 2011, 77, 208–213. [Google Scholar] [CrossRef]
- Boyapelly, K.; Bonin, M.-A.; Traboulsi, H.; Cloutier, A.; Phaneuf, S.C.; Fortin, D.; Cantin, A.M.; Richter, M.V.; Marsault, E. Synthesis and Characterization of a Phosphate Prodrug of Isoliquiritigenin. J. Nat. Prod. 2017, 80, 879–886. [Google Scholar] [CrossRef]
- Adianti, M.; Aoki, C.; Komoto, M.; Deng, L.; Shoji, I.; Wahyuni, T.S.; Lusida, M.I.; Fuchino, H.; Kawahara, N.; Hotta, H. Anti-hepatitis C virus compounds obtained from Glycyrrhiza uralensis and other Glycyrrhiza species. Microbiol. Immunol. 2014, 58, 180–187. [Google Scholar] [CrossRef]
- Lee, Y.S.; Kim, S.H.; Jung, S.H.; Kim, J.K.; Pan, C.-H.; Lim, S.S. Aldose Reductase Inhibitory Compounds from Glycyrrhiza uralensis. Biol. Pharm. Bull. 2010, 33, 917–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.W.; Kang, Y.H.; Kim, J.L.; Kwun, I.S.; Li, J.; Sung, H.Y. Isoliquiritigenin entails blockade of TGF-beta1-SMAD signaling for retarding high glucose-induced mesangial matrix accumulation. J. Agric. Food Chem. 2010, 58, 3205–3212. [Google Scholar]
- Kanazawa, M.; Satomi, Y.; Mizutani, Y.; Ukimura, O.; Kawauchi, A.; Sakai, T.; Baba, M.; Okuyama, T.; Nishino, H.; Miki, T. Isoliquiritigenin Inhibits the Growth of Prostate Cancer. Eur. Urol. 2003, 43, 580–586. [Google Scholar] [CrossRef]
- Cong, D.; Jiang, Y.; Liu, Y.; Yu, J.; Zhang, H.; Zhao, C. Screening and determination for potential inhibitory α-glucosidase constituents from Dalberigia odorifera T. Chen using ultrafiltration-LC/ESI-Ms(n). Biomed. Chromatogr. 2013, 27, 1621–1629. [Google Scholar]
- Park, S.-J.; Song, H.-Y.; Youn, H.-S. Suppression of the TRIF-dependent signaling pathway of toll-like receptors by isoliquiritigenin in RAW264.7 macrophages. Mol. Cells 2009, 28, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-J.; Youn, H.-S. Suppression of homodimerization of toll-like receptor 4 by isoliquiritigenin. Phytochemistry 2010, 71, 1736–1740. [Google Scholar] [CrossRef]
- Park, S.J.; Youn, H.S. Isoliquiritigenin suppresses the toll-interleukin-1 receptor domain-containing adapter inducing interferon-beta (TRIF)-dependent signaling pathway of toll- like receptors by targeting TBK1. J. Agric. Food Chem. 2010, 58, 4701–4705. [Google Scholar] [CrossRef]
- Guo, A.; He, D.; Xu, H.-B.; Geng, C.-A.; Zhao, J. Promotion of regulatory T cell induction by immunomodulatory herbal medicine licorice and its two constituents. Sci. Rep. 2015, 5, 14046. [Google Scholar] [CrossRef] [Green Version]
- Jhanji, V.; Liu, H.; Law, K.; Lee, V.Y.; Huang, S.F.; Pang, C.P.; Yam, G.H. Isoliquiritigenin from licorice root suppressed neovascularisation in experimental ocular an- giogenesis models. Br. J. Ophthalmol. 2011, 95, 1309–1315. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, N.; Han, S.; Wang, D.; Mo, S.; Yu, L.; Huang, H.; Tsui, K.; Shen, J.; Chen, J. Dietary Compound Isoliquiritigenin Inhibits Breast Cancer Neoangiogenesis via VEGF/VEGFR-2 Signaling Pathway. PLoS ONE 2013, 8, e68566. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.W.; Choi, J.S.; Choi, Y.J.; Bae, J.Y.; Li, J.; Kim, D.S.; Kim, J.L.; Shin, S.Y.; Lee, Y.J.; Kwun, I.S.; et al. Licorice isoliquiritigenin dampens angiogenic activity via inhibition of MAPK-responsive signaling pathways leading to induction of matrix metalloproteinases. J. Nutr. Biochem. 2010, 21, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Zhang, B.; Chen, N.; Chen, X.-Y.; Liu, L.-L.; Zheng, Q.-S.; Wang, Z.-P. Isoliquiritigenin treatment induces apoptosis by increasing intracellular ROS levels in HeLa cells. J. Asian Nat. Prod. Res. 2012, 14, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yeung, E.D.; Wang, J.; Panzhinskiy, E.E.; Tong, C.; Li, W.; Li, J. Isoliquiritigenin, a natural anti-oxidant, selectively inhibits the proliferation of prostate cancer cells. Clin. Exp. Pharmacol. Physiol. 2010, 37, 841–847. [Google Scholar] [PubMed]
- Lee, Y.M.; Lim, D.Y.; Choi, H.J.; Jung, J.I.; Chung, W.-Y.; Park, J.H.Y. Induction of Cell Cycle Arrest in Prostate Cancer Cells by the Dietary Compound Isoliquiritigenin. J. Med. Food 2009, 12, 8–14. [Google Scholar] [CrossRef]
- Cho, H.J.; Chung, W.Y.; Kwon, G.T.; Moon, A.; Park, J.H.; Park, K.K. Isoliquiritigenin inhibits migration and invasion of prostate cancer cells: Possible mediation by decreased JNK/AP-1 signaling. J. Nutr. Biochem. 2009, 20, 663–676. [Google Scholar]
- Li, Y.; Zhao, H.; Wang, Y.; Zheng, H.; Yu, W.; Chai, H.; Zhang, J.; Falck, J.R.; Guo, A.M.; Yue, J.; et al. Isoliquiritigenin induces growth inhibition and apoptosis through downregulating arachidonic acid metabolic network and the deactivation of PI3K/Akt in human breast cancer. Toxicol. Appl. Pharmacol. 2013, 272, 37–48. [Google Scholar] [CrossRef]
- Chen, G.; Hu, X.; Zhang, W.; Xu, N.; Wang, F.-Q.; Jia, J.; Zhang, W.-F.; Sun, Z.-J.; Zhao, Y.-F. Mammalian target of rapamycin regulates isoliquiritigenin-induced autophagic and apoptotic cell death in adenoid cystic carcinoma cells. Apoptosis 2011, 17, 90–101. [Google Scholar] [CrossRef]
- Sun, Z.-J.; Chen, G.; Zhang, W.; Hu, X.; Huang, C.-F.; Wang, Y.-F.; Jia, J.; Zhao, Y.-F. Mammalian Target of Rapamycin Pathway Promotes Tumor-Induced Angiogenesis in Adenoid Cystic Carcinoma: Its Suppression by Isoliquiritigenin through Dual Activation of c-Jun NH2-Terminal Kinase and Inhibition of Extracellular Signal-Regulated Kinase. J. Pharmacol. Exp. Ther. 2010, 334, 500–512. [Google Scholar] [CrossRef]
- Auyeung, K.K.-W.; Ko, J.K.-S. Novel herbal flavonoids promote apoptosis but differentially induce cell cycle arrest in human colon cancer cell. Investig. New Drugs 2010, 28, 1–13. [Google Scholar] [CrossRef]
- Wang, K.L.; Yu, Y.C.; Hsia, S.M. Perspectives on the role of isoliquiritigenin in cancer. Cancers 2021, 13, 115. [Google Scholar] [CrossRef]
- Shi, D.; Yang, J.; Jiang, Y.; Wen, L.; Wang, Z.; Yang, B. The antioxidant activity and neuroprotective mechanism of isoliquiritigenin. Free Rad. Biol. Med. 2020, 152, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Song, L.L.; Kosmeder, J.W., 2nd; Lee, S.K.; Gerhäuser, C.; Lantvit, D.; Moon, R.C.; Moriarty, R.M.; Pezzuto, J.M. Cancer chemopreventive activity mediated by 4′-bromoflavone, a potent inducer of phase II detoxification enzymes. Cancer Res. 1999, 59, 578–585. [Google Scholar] [PubMed]
- Cuendet, M.; Oteham, C.P.; Moon, R.C.; Pezzuto, J.M. Quinone Reductase Induction as a Biomarker for Cancer Chemoprevention. J. Nat. Prod. 2006, 69, 460–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinnen, R.D.; Mao, Y.; Fine, R.L. The Use of Fluorescent Probes in the Study of Reactive Oxygen Species in Pancreatic Cancer Cells. In Pancreatic Cancer; Humana Press: Totowa, NJ, USA, 2012; pp. 321–329. [Google Scholar]
- Giuliani, C.M.; Dass, C.R. Autophagy and cancer: Taking the toxic out of cytotoxics. J. Pharm. Pharmacol. 2013, 65, 777–789. [Google Scholar] [CrossRef]
- Poothong, J.; Jang, I.; Kaufman, R.J. Defects in protein folding and/or quality control cause protein aggregation in the ER. Prog. Mol. Subcell. Biol. 2021, 59, 115–143. [Google Scholar]
- Wang, Y.; Zhang, Z.; Auyeung, K.K.-W.; Cho, C.-H.; Yung, K.K.-L.; Ko, J.K.-S. Cryptotanshinone-Induced p53-Dependent Sensitization of Colon Cancer Cells to Apoptotic Drive by Regulation of Calpain and Calcium Homeostasis. Am. J. Chin. Med. 2020, 48, 1179–1202. [Google Scholar] [CrossRef]
- Marchi, S.; Giorgi, C.; Suski, J.M.; Agnoletto, C.; Bononi, A.; Bonora, M.; De Marchi, E.; Missiroli, S.; Patergnani, S.; Poletti, F.; et al. Mitochondria-Ros Crosstalk in the Control of Cell Death and Aging. J. Signal Transduct. 2011, 2012, 329635. [Google Scholar] [CrossRef] [Green Version]
- Donadelli, M.; Dando, I.; Zaniboni, T.; Costanzo, C.; Dalla Pozza, E.; Scupoli, M.T.; Scarpa, A.; Zappavigna, S.; Marra, M.; Abbruzzese, A.; et al. Gemcitabine/cannabinoid combination triggers autophagy in pancreatic cancer cells through a ROS-mediated mechanism. Cell Death Dis. 2011, 2, e152. [Google Scholar] [CrossRef]
- Yuan, X.; Yu, B.; Wang, Y.; Jiang, J.; Liu, L.; Zhao, H.; Qi, W.; Zheng, Q. Involvement of endoplasmic reticulum stress in isoliquiritigenin-induced SKOV-3 cell apoptosis. Recent Pat. Anti-Cancer Drug Dis. 2013, 8, 191–199. [Google Scholar] [CrossRef]
- Zhang, L.; Li, J.; Zong, L.; Chen, X.; Chen, K.; Jiang, Z.; Nan, L.; Li, X.; Li, W.; Shan, T.; et al. Reactive Oxygen Species and Targeted Therapy for Pancreatic Cancer. Oxidative Med. Cell. Longev. 2016, 2016, 1616781. [Google Scholar] [CrossRef] [Green Version]
- Ju, H.-Q.; Gocho, T.; Aguilar, M.; Wu, M.; Zhuang, Z.-N.; Fu, J.; Yanaga, K.; Huang, P.; Chiao, P.J. Mechanisms of Overcoming Intrinsic Resistance to Gemcitabine in Pancreatic Ductal Adenocarcinoma through the Redox Modulation. Mol. Cancer Ther. 2015, 14, 788–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, J.; Namba, T.; Takabatake, Y.; Kimura, T.; Takahashi, A.; Yamamoto, T.; Minami, S.; Sakai, S.; Fujimura, R.; Kaimori, J.-Y.; et al. Antioxidant role of autophagy in maintaining the integrity of glomerular capillaries. Autophagy 2018, 14, 53–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filomeni, G.; De Zio, D.; Cecconi, F. Oxidative stress and autophagy: The clash between damage and metabolic needs. Cell Death Differ. 2015, 22, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Ding, Z.; Du, K.; Ye, X.; Cheng, S. Reactive Oxygen Species as a Link between Antioxidant Pathways and Autophagy. Oxidative Med. Cell. Longev. 2021, 2021, 5583215. [Google Scholar] [CrossRef] [PubMed]
- Giordano, S.; Darley-Usmar, V.; Zhang, J. Autophagy as an essential cellular antioxidant pathway in neurodegenerative disease. Redox. Biol. 2014, 2, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Underwood, B.R.; Imarisio, S.; Fleming, A.; Rose, C.; Krishna, G.; Heard, P.; Quick, M.; Korolchuk, V.I.; Renna, M.; Sarkar, S.; et al. Antioxidants can inhibit basal autophagy and enhance neurodegeneration in models of polyglutamine disease. Human Mol. Genet. 2010, 19, 3413–3429. [Google Scholar] [CrossRef]
- El Hout, M.; Cosialls, E.; Mehrpour, M.; Hamaï, A. Crosstalk between autophagy and metabolic regulation of cancer stem cells. Mol. Cancer. 2020, 19, 27. [Google Scholar] [CrossRef]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef]
- Cantó, C.; Auwerx, J. AMP-activated protein kinase and its downstream transcriptional pathways. Cell. Mol. Life Sci. 2010, 67, 3407–3423. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Qian, W.; Li, J.; Jiang, Z.; Cheng, L.; Yan, B.; Cao, J.; Sun, L.; Zhou, C.; Lei, M.; et al. Loss of AMPK activation promotes the invasion and metastasis of pancreatic cancer through an HSF1-dependent pathway. Mol. Oncol. 2017, 11, 1475–1492. [Google Scholar] [CrossRef]
- Jiang, X.; Tan, H.-Y.; Teng, S.; Chan, Y.-T.; Wang, D.; Wang, N. The Role of AMP-Activated Protein Kinase as a Potential Target of Treatment of Hepatocellular Carcinoma. Cancers 2019, 11, 647. [Google Scholar] [CrossRef] [Green Version]
- Faubert, B.; Vincent, E.E.; Poffenberger, M.C.; Jones, R.G. The AMP-activated protein kinase (AMPK) and cancer: Many faces of a metabolic regulator. Cancer Lett. 2015, 356, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wang, X.; Contino, G.; Liesa, M.; Sahin, E.; Ying, H.; Bause, A.; Li, Y.; Stommel, J.M.; Dell’Antonio, G.; et al. Pancreatic cancers require autophagy for tumor growth. Genes Dev. 2011, 25, 717–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartlett, B.J.; Isakson, P.; Lewerenz, J.; Sanchez, H.; Kotzebue, R.W.; Cumming, R.C.; Harris, G.L.; Nezis, I.P.; Schubert, D.R.; Simonsen, A.; et al. p62, Ref(2)P and ubiquitinated proteins are conserved markers of neuronal aging, aggregate formation and progressive autophagic defects. Autophagy 2011, 7, 572–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanida, I.; Minematsu-Ikeguchi, N.; Ueno, T.; Kominami, E. Lysosomal Turnover, but Not a Cellular Level, of Endogenous LC3 is a Marker for Autophagy. Autophagy 2005, 1, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Rubinsztein, D.C.; Cuervo, A.M.; Ravikumar, B.; Sarkar, S.; Korolchuk, V.I.; Kaushik, S.; Klionsky, D.J. In search of an autophagomometer. Autophagy 2009, 5, 585–589. [Google Scholar] [CrossRef]
- Ma, X.; Tu, P.; Chen, Y.; Zhang, T.; Wei, Y.; Ito, Y. Preparative isolation and purification of two isoflavones from Astragalus membranaceus Bge. var. mongholicus (Bge.) Hsiao by high-speed counter-current chromatography. J. Chromatogr. A 2003, 992, 193–197. [Google Scholar] [CrossRef]
- Pan, H.; Fang, C.; Zhou, T.; Wang, Q.; Chen, J. Accumulation of calycosin and its 7-O-β-D-glucoside and related gene expression in seedlings of Astragalus membranaceus Bge. var. mongholicus (Bge.) Hsiao induced by low temperature stress. Plant Cell Rep. 2007, 26, 1111–1120. [Google Scholar]
- Li, T.W.; Huang, H.; Cui, Y.; Wang, S.Y. Huangqi bu chong dui sai ting yun dong yuan mian yi ji neng ying xiang de yan jiu [Study on the effect of astragalus supplementation on the immune function of rowers]. Nong Cun Jing Ji Yu Ke Ji 2016, 12, 160–161. [Google Scholar]
- Ren, C.X. Huangqi dui dong xun da qiang du xun lian qi zhong zhang pao yun dong yuan ti ye mian yi de ying xiang [Effects of Astragalus on the humoral immunity of mid long-distance runners during high-intensity training in winter]. Shenzhou 2012, 26, 39–40. [Google Scholar]
- Yan, M.; Liu, W.; Fu, Y.J.; Zu, Y.G.; Chen, C.Y.; Luo, M. Optimisation of the microwave-assisted extraction process for four main astragalosides in Radix Astragali. Food Chem. 2010, 119, 1663–1670. [Google Scholar] [CrossRef]
- Auyeung, K.K.W.; Han, Q.; Ko, J.K.S. Astragalus membranaceus: A review of its protection against inflammation and gastrointestinal cancers. Am. J. Chin. Med. 2016, 44, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Auyeung, K.K.; Sze, S.C.; Zhang, S.; Yung, K.K.; Ko, J.K. The dual roles of calycosin in growth inhibition and metastatic progression during pancreatic cancer development: A “TGF-β paradox”. Phytomedicine 2020, 68, 153177. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhang, Z.; Yung, K.K.; Ko, J.K. MUC1 is responsible for the pro-metastatic potential of calycosin in pancreatic ductal adenocarcinoma. Am. J. Cancer Res. 2022. accepted. [Google Scholar]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Zeng, S.; Pöttler, M.; Lan, B.; Crützmann, R.; Pilarsky, C.; Yang, H. Chemoresistance in pancreatic cancer. Int. J. Mol. Sci. 2019, 20, 4504. [Google Scholar] [CrossRef] [Green Version]
- Neoptolemos, J.P.; Kleeff, J.; Michl, P.; Costello, E.; Greenhalf, W.; Palmer, D.H. Therapeutic developments in pancreatic cancer: Current and future perspectives. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 333–348. [Google Scholar] [CrossRef]
- Murakami, T.; Hiroshima, Y.; Matsuyama, R.; Homma, Y.; Hoffman, R.M.; Endo, I. Role of the tumor microenvironment in pancreatic cancer. Ann. Gastroenterol. Surg. 2019, 3, 130–137. [Google Scholar] [CrossRef]
- Amaravadi, R.K.; Kimmelman, A.C.; Debnath, J. Targeting Autophagy in Cancer: Recent Advances and Future Directions. Cancer Discov. 2019, 9, 1167–1181. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, D.; Bläuer, M.; Hirota, M.; Ikonen, N.H.; Sand, J.; Laukkarinen, J. Autophagy is needed for the growth of pancreatic adenocarcinoma and has a cytoprotective effect against anticancer drugs. Eur. J. Cancer 2014, 50, 1382–1390. [Google Scholar] [CrossRef]
- Townsend, K.N.; Hughson, L.R.K.; Schlie, K.; Poon, V.; Westerback, A.; Lum, J.J. Autophagy inhibition in cancer therapy: Metabolic considerations for antitumor immunity. Immunol. Rev. 2012, 249, 176–194. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Chen, R.; Wang, Z.; Huang, Z.; Kong, N.; Zhang, M.; Han, W.; Lou, F.; Yang, J.; Zhang, Q.; et al. Autophagy and chemotherapy resistance: A promising therapeutic target for cancer treatment. Cell Death Dis. 2013, 4, e838. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-T.; Tan, H.-L.; Shui, G.; Bauvy, C.; Huang, Q.; Wenk, M.R.; Ong, C.N.; Codogno, P.; Shen, H.-M. Dual Role of 3-Methyladenine in Modulation of Autophagy via Different Temporal Patterns of Inhibition on Class I and III Phosphoinositide 3-Kinase. J. Biol. Chem. 2010, 285, 10850–10861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Commisso, C.; Davidson, S.M.; Soydaner-Azeloglu, R.G.; Parker, S.J.; Kamphorst, J.J.; Hackett, S.; Grabocka, E.; Nofal, M.; Drebin, J.A.; Thompson, C.B.; et al. Macropinocytosis of protein is an amino acid supply route in Ras-transformed cells. Nature 2013, 497, 633–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Pharmaceutical Name: | Glycyrrhiza Radix |
|---|---|
| English name: | Licorice root |
| Common Names: | Licorice root, licorice, liquorice, sweet root, Gan Cao (Chinese licorice) |
| Family: | Fabaceae |
| Common species in Latin: | Glycyrrhiza uralensis FISHCH. (Gan Cao, Chinese licorice) Glycyrrhiza inflate BAT. (Zhang Guo Gan Cao) Glycyrrhiza glabra L. (Guang Guo Gan Cao) Glycyrrhiza glabra (European licorice) Glycyrrhiza lepidota (American licorice) |
| Name of Isoflavonoid | Molecular Structures | |
|---|---|---|
| Glycyrrhizin |  |  |
| 18α-Glycyrrhetinic acid |  |  |
| 18β-Glycyrrhetinic acid |  |  |
| Licochalcone A |  |  |
| Licochalcone E |  |  |
| Glabridin |  |  |
| Liquiritigenin |  |  |
| Isoliquiritigenin |  |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Yung, K.K.-L.; Ko, J.K.-S. Therapeutic Intervention in Cancer by Isoliquiritigenin from Licorice: A Natural Antioxidant and Redox Regulator. Antioxidants 2022, 11, 1349. https://doi.org/10.3390/antiox11071349
Zhang Z, Yung KK-L, Ko JK-S. Therapeutic Intervention in Cancer by Isoliquiritigenin from Licorice: A Natural Antioxidant and Redox Regulator. Antioxidants. 2022; 11(7):1349. https://doi.org/10.3390/antiox11071349
Chicago/Turabian StyleZhang, Zhu, Ken Kin-Lam Yung, and Joshua Ka-Shun Ko. 2022. "Therapeutic Intervention in Cancer by Isoliquiritigenin from Licorice: A Natural Antioxidant and Redox Regulator" Antioxidants 11, no. 7: 1349. https://doi.org/10.3390/antiox11071349