
Comparative Assessment of the Antioxidant and Anticancer Activities of Plicosepalus acacia and Plicosepalus curviflorus: Metabolomic Profiling and In Silico Studies

 , ,
, ,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of the Extracts
2.2. Estimation of the Total Phenolic Content
2.3. Evaluation of the In Vitro Antioxidant Activity
2.3.1. DPPH Free Radical Scavenging Activity
2.3.2. Ferric Reducing Antioxidant Power (FRAP) Assay
2.3.3. Total Antioxidant Capacity (TAC) Assay
2.4. Anticancer Activity of P. acacia and P. curviflorus
2.4.1. Cytotoxic Activity
2.4.2. Annexin V/PI Staining and Cell Cycle Analysis
2.4.3. RT-PCR for the Apoptosis-Related Genes
2.5. Metabolomic Profiling by LC/MS/MS
2.6. Statistical Analyses
2.7. Molecular Docking
3. Results and Discussion
3.1. Total Phenolic Content (TPC)
3.2. Evaluation of the In Vitro Antioxidant Activity of P. curviflorus and P. acacia
3.3. Evaluation of the Anticancer Activity of P. acacia and P. curviflorus
3.3.1. Cytotoxicity Using the MTT Assay
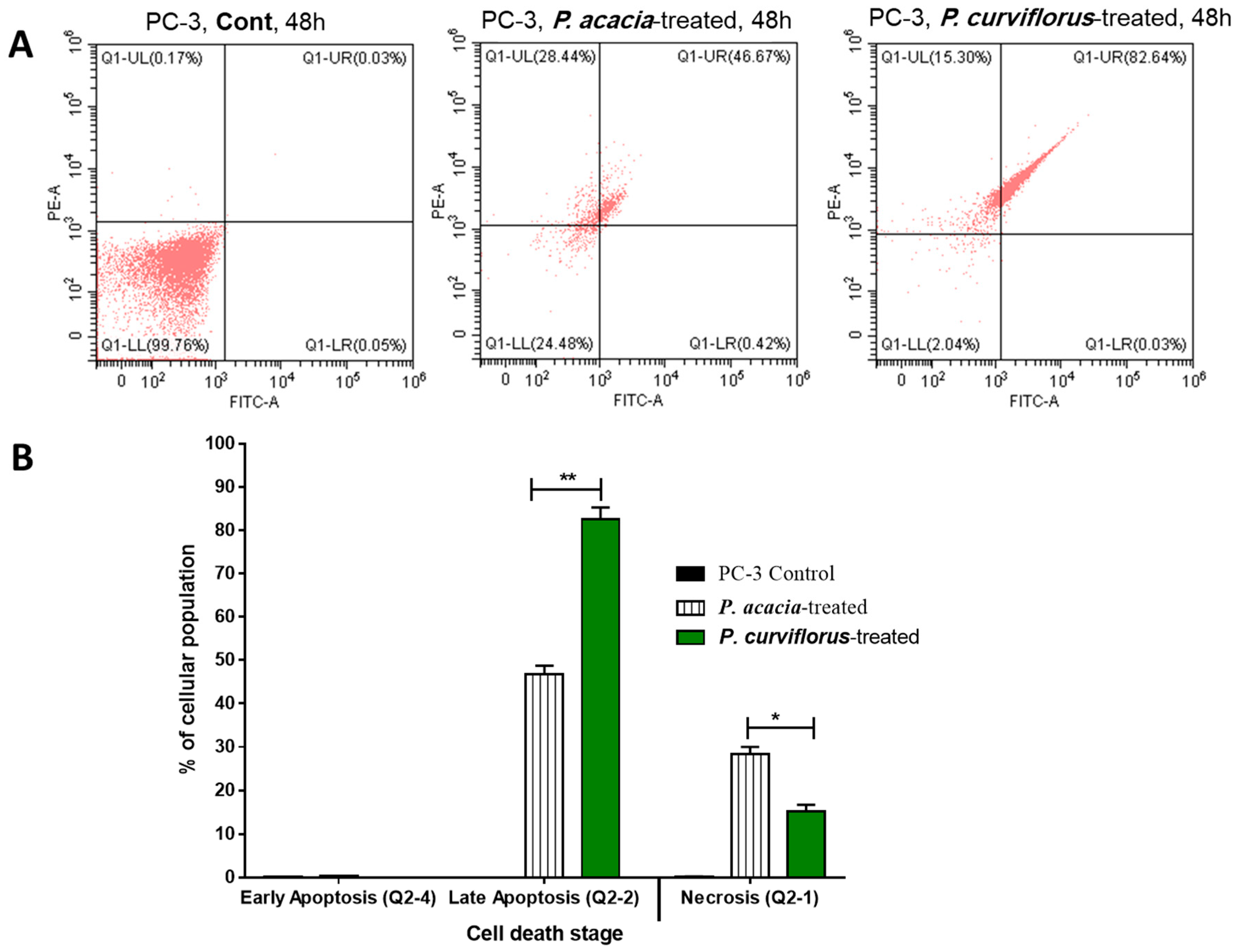
3.3.2. Apoptosis-Inducing Activity
3.3.3. RT-PCR Apoptosis-Related Genes
3.4. Principle Component Analysis (PCA)
3.5. LC-ESI-TOF-MS/MS Analysis of P. curviflorus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rt (min) | Measured m/z | Calculated m/z | Mass Error (ppm) | Adduct | Molecular Formula | MS/MS Spectrum | Deduced Compound | Ref. | |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 1.17 | 169.0137 | 169.0137 | 0 | [M − H]− | C7H6O5 | 169, 125 | Gallic acid | [101,102] |
| 2 | 1.35 | 343.0936 | 343.1029 | −27.1 | [M + H]+ | C15H18O9 | 343, 325, 283 | 1-Caffeoyl-β-D-glucose | [103] |
| 3 | 4.61 | 289.0706 | 289.0712 | −2.07 | [M − H]− | C15H14O6 | 289, 245,205, 179 | Catechin | [104] |
| 4 | 4.87 | 427.1018 | 427.1029 | −2.57 | [M + H]+ | C22H19O9 | 427,275, 150 | Curviflorin | [40] |
| 5 | 4.90 | 183.0309 | 183.0293 | 8.74 | [M − H]− | C8H8O5 | 183, 168, 140, 124 | Methyl gallate | [34,102,105] |
| 6 | 4.95 | 183.0633 | 183.0657 | −13.11 | [M + H]+ | C9H10O4 | 183, 168,140, 123 | Syringaldehyde | [102,106,107] |
| 7 | 5.03 | 153.0572 | 153.0552 | 13.07 | [M + H]+ | C8H8O3 | 153, 135, 125, 93 | Vanillin | [102,106,108] |
| 8 | 6.26 | 609.1453 | 609.1456 | −0.49 | [M − H]− | C27H30O16 | 609,449,301, 300 | Rutin | [34,109] |
| 9 | 6.54 | 463.0896 | 463.0871 | 4.12 | [M − 2H]− | C21H21O12 + | 463, 301,300,227 | Myrtillin | [102,110,111] |
| 10 | 6.71 | 611.1911 | 611.1976 | −10.63 | [M + H]+ | C28H34O15 | 611, 303, 268 | Hesperidin | [34,102,109] |
| 11 | 7.01 | 443.0958 | 443.0978 | −4.51 | [M + H]+ | C22H18O10 | 443,425,151, 123 | 3,3’,4’,5,7-pentahydroxyflavane−5-O-gallate | [40,112] |
| 12 | 7.22 | 447.0930 | 447.0927 | 0.67 | [M − H]− | C21H20O11 | 447, 385, 301, 284 | Quercetrin | [101,109] |
| 13 | 7.28 | 477.1032 | 477.1033 | −0.21 | [M − H]− | C22H22O12 | 477, 314, 285, 271, 243 | Isorhamnetin−3-glucoside | [102,110,111] |
| 14 | 7.56 | 459.0927 | 459.1054 | 27.7 | [M + H]+ | C22H19O11 | 459, 441, 307 | 3,3’,4’,5,5′,7-hexahydroxyflavane−5-O-gallate | [40] |
| 15 | 9.51 | 301.0359 | 301.0348 | 3.65 | [M − H]− | C15H9O7 | 301,284, 255, 151 | Quercetin | [34,109] |
| 16 | 9.94 | 303.0467 | 303.0499 | −10.56 | M + | C15H11O7 + | 303, 284, 274, 257, 247, 229 | Delphinidin | [113] |
| 17 | 9.94 | 315.0544 | 315.0505 | 12.38 | [M − H]− | C16H12O7 | 315, 300, 285, 271, 243, 151 | Isorhamnetin | [41,114] |
| 18 | 9.99 | 285.0390 | 285.0399 | −3.16 | [M − H]− | C15H10O6 | 285, 257, 241,223,197, 151 | Kaempferol | [41] |
| 19 | 20.86 | 599.4351 | 599.4288 | 10.51 | [M + Na]+ | C35H60O6 | 599, 413 | β-Sitosterol−3-O-β-D-glucoside | [115] |
| 20 | 22.03 | 487.3445 | 487.3423 | 4.51 | [M − H]− | C30H48O5 | 487, 425, 279 | Euscaphic acid | [116,117] |
| 21 | 22.73 | 471.3469 | 471.3474 | −1.06 | [M − H]− | C30H48O4 | 471, 453, 409 | Pomolic acid | [116] |
| 22 | 23.03 | 439.3575 | 439.3576 | −0.23 | [M + H+ − H2O]+ | C30H48O2 | 439, 393, 215, 203, 161, 147, 95 | Ursolic acid | [118,119] |
| 23 | 24.54 | 409.387 | 409.3834 | 8.79 | [M + H+ − H2O]+ | C30H50O | 409, 137, 109 | Lupeol | [119,120] |
| 24 | 25.29 | 395.3592 | 395.3678 | −21.75 | [M + H+ − H2O]+ | C29H48O1 | 395, 378, 311, 297, 255, 161, 147 | Stigmasterol | [120,121] |
| 25 | 26.51 | 397.3866 | 397.3834 | 8.05 | [M + H+ − H2O]+ | C29H50O1 | 397, 255, 161, 147 | β-Sitosterol | [120,121,122] |
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jang, E.; Lee, J.H. Promising Anticancer Activities of Alismatis rhizome and its triterpenes via p38 and PI3K/Akt/mTOR Signaling Pathways. Nutrients 2021, 13, 2455. [Google Scholar] [CrossRef] [PubMed]
- Santos-Pereira, C.; Rodrigues, L.R.; Côrte-Real, M. Plasmalemmal V-ATPase as a potential biomarker for lactoferrin-based anticancer therapy. Biomolecules 2022, 12, 119. [Google Scholar] [CrossRef] [PubMed]
- Efenberger-Szmechtyk, M.; Nowak, A.; Nowak, A. Cytotoxic and DNA-damaging effects of Aronia melanocarpa, Cornus mas, and Chaenomeles superba leaf extracts on the human colon adenocarcinoma cell line Caco-2. Antioxidants 2020, 9, 1030. [Google Scholar] [CrossRef] [PubMed]
- Losada-Echeberría, M.; Herranz-López, M.; Micol, V.; Barrajón-Catalán, E. Polyphenols as promising drugs against main breast cancer signatures. Antioxidants 2017, 6, 88. [Google Scholar] [CrossRef] [Green Version]
- Elhady, S.S.; Eltamany, E.E.; Shaaban, A.E.; Bagalagel, A.A.; Muhammad, Y.A.; El-Sayed, N.M.; Ayyad, S.-E.N.; Ahmed, A.A.M.; Elgawish, M.S.; Ahmed, S.A. Jaceidin flavonoid isolated from Chiliadenus montanus attenuates tumor progression in mice via VEGF inhibition: In Vivo and in silico studies. Plants 2020, 9, 1031. [Google Scholar] [CrossRef] [PubMed]
- Torić, J.; Brozovic, A.; Baus Lončar, M.; Brala, C.J.; Marković, A.K.; Benčić, D.; Barbarić, M. Biological activity of phenolic compounds in extra virgin olive oils through their phenolic profile and their combination with anticancer drugs observed in human cervical carcinoma and colon adenocarcinoma cells. Antioxidants 2020, 9, 453. [Google Scholar] [CrossRef]
- World Health Organization. Available online: http://www.who.int/cancer/en/ (accessed on 8 March 2022).
- Huang, Y.J.; Wang, K.L.; Chen, H.Y.; Chiang, Y.F.; Hsia, S.M. Protective effects of epigallocatechin gallate (EGCG) on endometrial, Breast, and Ovarian Cancers. Biomolecules 2020, 10, 1481. [Google Scholar] [CrossRef]
- Koygun, G.; Arslan, E.; Zengin, G.; Orlando, G.; Ferrante, C. Comparison of anticancer activity of Dorycnium pentaphyllum extract on MCF-7 and MCF-12A cell line: Correlation with invasion and adhesion. Biomolecules 2021, 11, 671. [Google Scholar] [CrossRef]
- American Cancer Society. Key Statistics for Ovarian Cancer. Available online: https://www.cancer.org/cancer/ovarian-cancer/about/key-statistics.html (accessed on 18 February 2022).
- American Cancer Society. Trends in Breast Cancer Deaths. Available online: https://www.cancer.org/cancer/breast-cancer/about/how-common-is-breast-cancer.html (accessed on 18 February 2022).
- Lu, L.; Cole, A.; Huang, D.; Wang, Q.; Guo, Z.; Yang, W.; Lu, J. Clinical significance of hepsin and underlying signaling pathways in Prostate Cancer. Biomolecules 2022, 12, 203. [Google Scholar] [CrossRef]
- Varghese, E.; Liskova, A.; Kubatka, P.; Mathews, S.S.; Büsselberg, D. Anti-Angiogenic Effects of Phytochemicals on miRNA Regulating Breast Cancer Progression. Biomolecules 2020, 10, 191. [Google Scholar] [CrossRef] [Green Version]
- Sharifi-Rad, J.; Ozleyen, A.; Boyunegmez, T.T.; Oluwaseun, A.C.; El Omari, N.; Balahbib, A.; Taheri, Y.; Bouyahya, A.; Martorell, M.; Martins, N.; et al. Natural Products and Synthetic Analogs as a Source of Antitumor Drugs. Biomolecules 2019, 9, 679. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Ha, J.; Kim, J.; Cho, Y.; Ahn, J.; Cheon, C.; Kim, S.-H.; Ko, S.-G.; Kim, B. Natural Products for Pancreatic Cancer Treatment: From Traditional Medicine to Modern Drug Discovery. Nutrients 2021, 13, 3801. [Google Scholar] [CrossRef] [PubMed]
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Büsselberg, D. Flavonoids in Cancer and Apoptosis. Cancers 2019, 11, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abotaleb, M.; Liskova, A.; Kubatka, P.; Büsselberg, D. Therapeutic potential of plant phenolic acids in the treatment of cancer. Biomolecules 2020, 10, 221. [Google Scholar] [CrossRef] [Green Version]
- Nowak, J.; Kiss, A.K.; Wambebe, C.; Katuura, E.; Kuźma, Ł. Phytochemical analysis of polyphenols in leaf extract from Vernonia amygdalina Delile plant growing in Uganda. Appl. Sci. 2022, 12, 912. [Google Scholar] [CrossRef]
- Norman, G.; Lewis, L.B.; Davin, S.S. The Nature and Function of Lignins. In Comprehensive Natural Products Chemistry; Barton, D., Nakanishi, K., Meth-Cohn, O., Eds.; Elsevier Science: New York, NY, USA, 1999; pp. 617–745. [Google Scholar]
- Leitão, F.; Moreira, D.L.; De Almeida, M.Z.; Leitãoa, S.G. Secondary metabolites from the mistletoes Ruthanthus marginatus and Struthanthus concinnus (Loranthaceae). Biochem. Syst. Ecol. 2013, 48, 215. [Google Scholar] [CrossRef] [Green Version]
- Moghadamtousi, S.Z.; Hajrezaei, M.; Abdul Kadir, H.; Zandi, K. Loranthus micranthus Linn.: Biological Activities and Phytochemistry. J. Evid.-Based Complement. Altern. Med. 2013, 4, 273712. [Google Scholar]
- Ibrahim, J.A.; Ayodele, A.E. Taxonomic significance of leaf epidermal characters of the family Loranthaceae in Nigeria. World Appl. Sci. J. 2013, 24, 1172–1179. [Google Scholar]
- Ogunmefun, O.T.; Fasola, T.R.; Saba, A.B.; Oridupa, O.A. The toxicity evaluation of Phragmanthera incana (Klotzsch) growing on two plant hosts and its effect on Wistar rats’ haematology and serum biochemistry. Acad. J. Plant Sci. 2013, 6, 92–98. [Google Scholar]
- Loef, M.; Walach, H. Quality of life in cancer patients treated with mistletoe: A systematic review and meta-analysis. BMC Complement. Med. Ther. 2020, 20, 227. [Google Scholar] [CrossRef]
- Orfali, R.; Perveen, S.; Siddiqui, N.A.; Alaam, P.; Alhowiriny, T.A.; Al-Taweel, A.M. Pharmacological Evaluation of Secondary Metabolites and Their Simultaneous Determination in the Arabian Medicinal Plant Plicosepalus curviflorus Using HPTLC Validated Method. J. Anal. Methods Chem. 2019, 3, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badr, J.M.; Shaala, L.A.; Youssef, D.T.A. Loranthin: A new polyhydroxylated flavanocoumarin from Plicosepalus acacia with significant free radical scavenging and antimicrobial activity. Phytochem. Lett. 2013, 6, 113. [Google Scholar] [CrossRef]
- Amina, M.; Al Musayeib, N.M.; Alarfaj, N.A.; El-Tohamy, M.F.; Al-Hamoud, G.A.; Alqenaei, M.K.M. The Fluorescence Detection of Phenolic Compounds in Plicosepalus curviflorus Extract Using Biosynthesized ZnO Nanoparticles and Their Biomedical Potential. Plants 2022, 11, 361. [Google Scholar] [CrossRef] [PubMed]
- Al-Fatimi, M.; Wurster, M.; Schröder, G.; Lindequist, U. Antioxidant, antimicrobial and cytotoxic activities of selected medicinal plants from Yemen. J. Ethnopharmacol. 2007, 111, 657. [Google Scholar] [CrossRef]
- El-Shafei, G.; Al-Hazmi, B.; Mar Ghelani, A.; Al-Moalem, D.; Badr, J.M.; Moneib, N.A. Antimicrobial activity of different extracts of Plicosepalus acacia. Rec. Pharm. Biomed. Sci. 2017, 1, 47. [Google Scholar] [CrossRef] [Green Version]
- Alburyhi, M.M.; Saif, A.A.; Al-Shibani, A.M.; Al-Mahbshi, H.M.; Noman, M.A. Hepatoprotective activity of Plicosepalus acacia extract against carbon tetrachloride-induced hepatic damage in Wistar Albino Rats. Int. J. Pharm. Pharm. Res. 2018, 13, 89. [Google Scholar]
- Kelsey, N.A.; Wilkins, H.M.; Linseman, D.A. Nutraceutical antioxidants as novel neuroprotective agents. Molecules 2010, 15, 7792. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.F.; Brown, P.H.; Lyle, B.J.; Chen, Y.; Black, R.M.; Williams, C.E.; Lin, Y.C.; Cheng, I.H. Roasted coffees high in lipophilic antioxidants and chlorogenic acid lactones are more neuroprotective than green coffees. J. Agric. Food Chem. 2009, 57, 9801. [Google Scholar] [CrossRef]
- Sadi, G.; Gűray, T. Gene expressions of Mn-SOD and GPx-1 in streptozotocin induced diabetes: Effect of antioxidants. Mol. Cell. Biochem. 2009, 327, 127. [Google Scholar] [CrossRef]
- Abdel-Hamed, A.R.; Mehanna, E.T.; Hazem, R.M.; Badr, J.M.; Abo-Elmatty, D.M.; Abdel-Kader, M.S.; Goda, M.S. Plicosepalus acacia Extract and Its Major Constituents, Methyl Gallate and Quercetin, Potentiate Therapeutic Angiogenesis in Diabetic Hind Limb Ischemia: HPTLC Quantification and LC-MS/MS Metabolic Profiling. Antioxidants 2021, 10, 1701. [Google Scholar] [CrossRef]
- Al-Taweel, A.M.; Perveen, S.; Fawzy, G.A.; Alqasoumi, S.I.; El Tahir, K.E. New flavane gallates isolated from the leaves of Plicosepalus curviflorus and their hypoglycemic activity. Fitoterapia 2012, 83, 1610. [Google Scholar] [CrossRef] [PubMed]
- Fawzy, G.A.; Al-Taweel, A.M.; Perveen, S. Anticancer activity of flavane gallates isolated from Plicosepalus curviflorus. Pharmacogn. Mag. 2014, 10, 519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waly, N.M.; Ali, A.E.E.; Jrais, R.N. Botanical and biological studies of six parasitic species of family Loranthaceae growing in Kingdom of Saudi Arabia. Int. J. Environ. Sci. 2014, 4, 196. [Google Scholar]
- Badr, J.M.; Ibrahim, S.R.M.; Abou-Hussein, D.R. Plicosepalin A, a new antioxidant catechin–gallic acid derivative of inositol from the mistletoe Plicosepalus curviflorus. Z. Nat. 2016, 71, 375. [Google Scholar] [CrossRef]
- Al-Musayeib, N.M.; Ibrahim, S.R.M.; Musarat, A.; Al Hamoud, G.A.; Mohamed, G.A. Curviflorside and curviflorin, new naphthalene glycoside and flavanol from Plicosepalus curviflorus. Z. Nat. C. J. Biosci. 2017, 72, 197. [Google Scholar] [CrossRef]
- Al-Taweel, A.M.; Perveen, S.; Alqasoumi, S.I.; Orfali, R.; Aati, H.Y.; Alsultan, E.N.; Alghanem, B.; Shaibah, H. New flavane gallates from the aerial part of an African/Arabian medicinal plant Plicosepalus curviflorus by LC–MS and NMR based molecular characterization. J. King Saud. Univ. Sci. 2021, 33, 101289. [Google Scholar] [CrossRef]
- Eltamany, E.E.; Elhady, S.S.; Ahmed, H.A.; Badr, J.M.; Noor, A.O.; Ahmed, S.A.; Nafie, M.S. Chemical Profiling, Antioxidant, Cytotoxic Activities and Molecular Docking Simulation of Carrichtera annua DC. (Cruciferae). Antioxidants 2020, 9, 1286. [Google Scholar] [CrossRef]
- Yen, G.C.; Duh, P.D. Scavenging effect of methanolic extracts of peanut hulls on free-radical and active-oxygen species. J. Agric. Food Chem. 1994, 42, 629. [Google Scholar] [CrossRef]
- Al Zahrani, N.A.; El-Shishtawy, R.M.; Elaasser, M.M.; Asiri, A.M. Synthesis of Novel Chalcone-Based Phenothiazine Derivatives as Antioxidant and Anticancer Agents. Molecules 2020, 25, 4566. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reaction: Antioxidative activities of products of browning reaction prepared from glucosamine. Jpn. J. Nutr. Diet. 1986, 44, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, I.C.F.R.; Baptista, P.; Vilas-Boas, M.; Barros, L. Free-radical scavenging capacity and reducing power of wild edible mushrooms from northeast Portugal: Individual cap and stipe activity. Food Chem. 2007, 100, 1511. [Google Scholar] [CrossRef]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337. [Google Scholar] [CrossRef] [PubMed]
- Nafie, M.S.; Amer, A.M.; Mohamed, A.K.; Tantawy, E.S. Discovery of novel pyrazolo[3,4-b] pyridine scaffold-based derivatives as potential PIM-1 kinase inhibitors in breast cancer MCF-7 cells. Bioorg. Med. Chem. 2020, 28, 115828. [Google Scholar] [CrossRef] [PubMed]
- Tantawy, E.S.; Amer, A.M.; Mohamed, E.K.; Abd Alla, M.M.; Nafie, M.S. Synthesis, characterization of some pyrazine derivatives as anti-cancer agents: In vitro and in Silico approaches. J. Mol. Struct. 2020, 1210, 128013. [Google Scholar] [CrossRef]
- Nafie, M.S.; Arafa, K.; Sedky, N.K.; Alakhdar, A.A.; Arafa, R.K. Triaryl dicationic DNA minor-groove binders with antioxidant activity display cytotoxicity and induce apoptosis in breast cancer. Chem.-Biol. Interact. 2020, 324, 109087. [Google Scholar] [CrossRef]
- Gad, E.M.; Nafie, M.S.; Eltamany, E.H.; Hammad, M.S.A.G.; Barakat, A.; Boraei, A.T.A. Discovery of New Apoptosis-Inducing Agents for Breast Cancer Based on Ethyl 2-Amino-4,5,6,7-Tetra Hydrobenzo[b]Thiophene-3-Carboxylate: Synthesis, In Vitro, and In Vivo Activity Evaluation. Molecules 2020, 25, 2523. [Google Scholar] [CrossRef]
- Abdelhameed, R.F.A.; Habib, E.S.; Ibrahim, A.K.; Yamada, K.; Abdel-Kader, M.S.; Ahmed, S.A.; Ibrahim, A.K.; Badr, J.M.; Nafie, M.S. Chemical Constituent Profiling of Phyllostachys heterocycla var. Pubescens with Selective Cytotoxic Polar Fraction through EGFR Inhibition in HepG2 cells. Molecules 2021, 26, 940. [Google Scholar] [CrossRef]
- Abdelhameed, R.F.A.; Habib, E.S.; Ibrahim, A.K.; Yamada, K.; Abdel-Kader, M.S.; Ibrahim, A.K.; Ahmed, S.A.; Badr, J.M.; Nafie, M.S. Chemical profiling, cytotoxic activities through apoptosis induction in MCF-7 cells and molecular docking of Phyllostachys heterocycla bark nonpolar extract. J. Biomol. Struct. Dyn. 2021, 2, 1. [Google Scholar] [CrossRef]
- Sarhan, A.A.M.; Boraei, A.T.A.; Barakat, A.; Nafie, M.S. Discovery of hydrazide-based pyridazino[4,5-b]indole scaffold as a new phosphoinositide 3-kinase (PI3K) inhibitor for breast cancer therapy. RSC Adv. 2020, 10, 19534. [Google Scholar] [CrossRef]
- Nafie, M.S.; Tantawy, M.A.; Elmgeed, G.A. Screening of different drug design tools to predict the mode of action of steroidal derivatives as anti-cancer agents. Steroids 2019, 152, 108485. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313. [Google Scholar] [CrossRef] [PubMed]
- Lyu, J.I.; Ryu, J.; Seo, K.-S.; Kang, K.-Y.; Park, S.H.; Ha, T.H.; Ahn, J.-W.; Kang, S.-Y. Comparative Study on Phenolic Compounds and Antioxidant Activities of Hop (Humulus lupulus L.) Strobile Extracts. Plants 2022, 11, 135. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Ray, S. Comparative assessment of total phenolic content and in vitro antioxidant activities of bark and leaf methanolic extracts of Manilkara hexandra (Roxb.) Dubard. J. King Saud. Univ. Sci. 2020, 32, 643. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.N.; Xie, W.H.; Zheng, Y.X.; Cao, J.P.; Cao, P.R.; Chen, Q.J.; Li, X.; Sun, C.D. The Growth of SGC-7901 Tumor Xenografts Was Suppressed by Chinese Bayberry Anthocyanin Extract through Upregulating KLF6 Gene Expression. Nutrients 2016, 8, 599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awad, B.M.; Abd-Alhaseeb, M.M.; Habib, E.S.; Ibrahim, A.K.; Ahmed, S.A. Antitumor activity of methoxylated flavonoids separated from Achillea fragrantissima extract in Ehrlich’s ascites carcinoma model in mice. J. Herbmed. Pharmacol. 2019, 9, 28. [Google Scholar] [CrossRef] [Green Version]
- Ramos, S. Effects of dietary flavonoids on apoptotic pathways related to cancer chemoprevention. J. Nutr. Biochem. 2007, 18, 427. [Google Scholar] [CrossRef] [Green Version]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [Green Version]
- Fan, M.; Chen, G.; Zhang, Y.; Nahar, L.; Sarker, S.D.; Hu, G.; Guo, M. Antioxidant and Anti-Proliferative Properties of Hagenia abyssinica Roots and Their Potentially Active Components. Antioxidants 2020, 9, 143. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.-J.; Hsu, W.-H.; Lee, K.-H.; Chen, K.-C.; Lin, C.-W.; Lee, Y.-L.A.; Ko, T.-P.; Lee, L.-T.; Lee, M.-T.; Chang, M.-S.; et al. Dietary Flavonoids Luteolin and Quercetin Inhibit Migration and Invasion of Squamous Carcinoma through Reduction of Src/Stat3/S100A7 Signaling. Antioxidants 2019, 8, 557. [Google Scholar] [CrossRef] [Green Version]
- Xi, X.; Wang, J.; Qin, Y.; You, Y.; Huang, W.; Zhan, J. The Biphasic Effect of Flavonoids on Oxidative Stress and Cell Proliferation in Breast Cancer Cells. Antioxidants 2022, 11, 622. [Google Scholar] [CrossRef]
- Li, C.; Zhang, L.; Liu, C.; He, X.; Chen, M.; Chen, J. Lipophilic Grape Seed Proanthocyanidin Exerts Anti-Cervical Cancer Effects in HeLa Cells and a HeLa-Derived Xenograft Zebrafish Model. Antioxidants 2022, 11, 422. [Google Scholar] [CrossRef] [PubMed]
- Hafeez, B.B.; Siddiqui, I.A.; Asim, M.; Malik, A.; Afaq, F.; Adhami, V.M.; Saleem, M.; Din, M.; Mukhtar, H. A dietary anthocyanidin delphinidin induces apoptosis of human prostate cancer PC3 cells in vitro and in vivo: Involvement of nuclear factor-kappaB signaling. Cancer Res. 2008, 68, 8564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Choi, H.K.; Kim, Y.K.; Lee, H.J. Delphinidin and Its Glycosides’ War on Cancer: Preclinical Perspectives. Int. J. Mol. Sci. 2021, 22, 11500. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Luo, E.; Liu, X.; Han, B.; Yu, X.; Peng, X. Delphinidin-3-glucoside suppresses breast carcinogenesis by inactivating the Akt/HOTAIR signaling pathway. BMC Cancer 2016, 16, 423. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Harshavardhan, S.; Chirumarry, S.; Poornachandra, Y.; Jang, K.; Kumar, C.; Yoon, Y.-J.; Zhao, B.-X.; Miao, J.-Y.; Shin, D.-S. Design, Synthesis In vitro Anticancer activity and docking studies of (−)-catechin derivatives. Bull. Korean Chem. Soc. 2015, 36, 564. [Google Scholar]
- Tsai, Y.J.; Chen, B.H. Preparation of catechin extracts and nanoemulsions from green tea leaf waste and their inhibition effect on prostate cancer cell PC-3. Int. J. Nanomed. 2016, 11, 1907. [Google Scholar]
- Sun, H.; Yin, M.; Hao, D.; Shen, Y. Anti-Cancer Activity of Catechin against A549 Lung Carcinoma Cells by Induction of Cyclin Kinase Inhibitor p21 and Suppression of Cyclin E1 and P–AKT. Appl. Sci. 2020, 10, 2065. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.; Dong, J. (-)-Epicatechin acts as a potent agonist of the membrane androgen receptor, ZIP9 (SLC39A9), to promote apoptosis of breast and prostate cancer cells. J. Steroid Biochem. Mol. Biol. 2021, 211, 105906. [Google Scholar] [CrossRef]
- Izzo, S.; Naponelli, V.; Bettuzzi, S. Flavonoids as Epigenetic Modulators for Prostate Cancer Prevention. Nutrients 2020, 12, 1010. [Google Scholar] [CrossRef] [Green Version]
- Costea, T.; Nagy, P.; Ganea, C.; Szöllősi, J.; Mocanu, M.M. Molecular Mechanisms and Bioavailability of Polyphenols in Prostate Cancer. Int. J. Mol. Sci. 2019, 20, 1062. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.C.; Yen, C.Y.; Wu, R.S.; Yang, J.S.; Lu, H.F.; Lu, K.W.; Lo, C.; Chen, H.Y.; Tang, N.Y.; Wu, C.C.; et al. The roles of endoplasmic reticulum stress and mitochondrial apoptotic signaling pathway in quercetin-mediated cell death of human prostate cancer PC-3 cells. Environ. Toxicol. 2014, 29, 428. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.J.; Wilson, L.; Jordan, M.A.; Nguyen, V.; Tang, J.; Smiyun, G. Hesperidin suppressed proliferations of both human breast cancer and androgen-dependent prostate cancer cells. Phytother. Res. 2010, 24, S15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, H.; Zhang, J.; Gao, J.; Ge, X.; Lou, G. Hesperidin inhibits HeLa cell proliferation through apoptosis mediated by endoplasmic reticulum stress pathways and cell cycle arrest. BMC Cancer 2015, 15, 682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da, J.; Xu, M.; Wang, Y.; Li, W.; Lu, M.; Wang, Z. Kaempferol Promotes Apoptosis While Inhibiting Cell Proliferation via Androgen-Dependent Pathway and Suppressing Vasculogenic Mimicry and Invasion in Prostate Cancer. Anal. Cell Pathol. 2019, 2019, 1907698. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, G.; Sun, C.; Peng, F.; Yu, L.; Chen, Y.; Tan, Y.; Cao, X.; Tang, Y.; Xie, X.; et al. Chemistry, pharmacokinetics, pharmacological activities, and toxicity of Quercitrin. Phytother. Res. 2022, 36, 1545. [Google Scholar] [CrossRef]
- Zhai, T.; Zhang, X.; Hei, Z.; Jin, L.; Han, C.; Ko, A.T.; Yu, X.; Wang, J. Isorhamnetin Inhibits Human Gallbladder Cancer Cell Proliferation and Metastasis via PI3K/AKT Signaling Pathway Inactivation. Front. Pharmacol. 2021, 12, 628621. [Google Scholar] [CrossRef]
- Cai, F.; Zhang, Y.; Li, J.; Huang, S.; Gao, R. Isorhamnetin inhibited the proliferation and metastasis of androgen-independent prostate cancer cells by targeting the mitochondrion-dependent intrinsic apoptotic and PI3K/Akt/mTOR pathway. Biosci. Rep. 2020, 40, BSR20192826. [Google Scholar] [CrossRef] [Green Version]
- Satari, A.; Amini, S.A.; Raeisi, E.; Lemoigne, Y.; Heidarian, E. Synergetic Impact of Combined 5-Fluorouracil and Rutin on Apoptosis in PC3 Cancer Cells through the Modulation of P53 Gene Expression. Adv. Pharm. Bull. 2019, 9, 462. [Google Scholar] [CrossRef] [Green Version]
- Woyengo, T.; Ramprasath, V.; Jones, P. Anticancer effects of phytosterols. Eur. J. Clin. Nutr. 2009, 63, 813. [Google Scholar] [CrossRef]
- Awad, A.B.; Burr, A.T.; Fink, C.S. Effect of resveratrol and β-sitosterol in combination on reactive oxygen species and prostaglandin release by PC-3 cells. Prostaglandins Leukot. Essent. Fat. Acids 2005, 72, 219. [Google Scholar] [CrossRef]
- von Holtz, R.L.; Fink, C.S.; Awad, A.B. Beta-Sitosterol activates the sphingomyelin cycle and induces apoptosis in LNCaP human prostate cancer cells. Nutr. Cancer 1998, 32, 8. [Google Scholar] [CrossRef]
- Rajavel, T.; Packiyaraj, P.; Suryanarayanan, V.; Singh, S.; Kandasamy, R.; Kasi, P.D. β-Sitosterol targets Trx/Trx1 reductase to induce apoptosis in A549 cells via ROS mediated mitochondrial dysregulation and p53 activation. Sci. Rep. 2018, 8, 2071. [Google Scholar] [CrossRef] [Green Version]
- Dolai, N.; Kumar, A.; Islam, A.; Haldar, P.K. Apoptogenic effects of β-sitosterol glucoside from Castanopsis indica leaves. Nat. Prod. Res. 2016, 30, 482. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, X.; Wang, M.; Lin, Y.; Zhou, S. Stigmasterol simultaneously induces apoptosis and protective autophagy by inhibiting Akt/mTOR pathway in gastric cancer cells. Front. Oncol. 2021, 11, 629008. [Google Scholar] [CrossRef]
- Zhang, Y.; Kong, C.; Zeng, Y.; Wang, L.; Li, Z.; Wang, H.; Xu, C.; Sun, Y. Ursolic acid induces PC-3 cell apoptosis via activation of JNK and inhibition of Akt pathways in vitro. Mol. Carcinog. 2010, 49, 374. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Kong, C.Z.; Wang, L.H.; Li, J.Y.; Liu, X.K.; Xu, B.; Xu, C.L.; Sun, Y.H. Ursolic acid overcomes Bcl-2-mediated resistance to apoptosis in prostate cancer cells involving activation of JNK-induced Bcl-2 phosphorylation and degradation. J. Cell. Biochem. 2010, 109, 764. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Manu, K.A.; Ong, T.H.; Ramachandran, L.; Surana, R.; Bist, P.; Lim, L.H.; Kumar, A.P.; Hui, K.M.; Sethi, G. Inhibition of CXCR4/CXCL12 signaling axis by ursolic acid leads to suppression of metastasis in transgenic adenocarcinoma of mouse prostate model. Int. J. Cancer 2011, 129, 1552. [Google Scholar] [CrossRef]
- Hsu, Y.L.; Kuo, P.L.; Lin, C.C. Proliferative inhibition, cell-cycle dysregulation, and induction of apoptosis by ursolic acid in human non-small cell lung cancer A549 cells. Life Sci. 2004, 75, 2303. [Google Scholar] [CrossRef]
- Prasad, S.; Nigam, N.; Kalra, N.; Shukla, Y. Regulation of signaling pathways involved in Lupeol induced inhibition of proliferation and induction of apoptosis in human prostate cancer cells. Mol. Carcinog. 2008, 47, 916. [Google Scholar]
- Huang, S.-P.; Ho, T.-M.; Yang, C.-W.; Chang, Y.-J.; Chen, J.-F.; Shaw, N.-S.; Horng, J.-C.; Hsu, S.-L.; Liao, M.-Y.; Wu, L.-C.; et al. Chemopreventive Potential of Ethanolic Extracts of Luobuma Leaves (Apocynum venetum L.) in Androgen Insensitive Prostate Cancer. Nutrients 2017, 9, 948. [Google Scholar] [CrossRef] [Green Version]
- Martins, C.A.; Rocha, G.D.G.; Gattass, C.R.; Takiya, C.M. Pomolic acid exhibits anticancer potential against a docetaxel-resistant PC3 prostate cell line. Oncol. Rep. 2019, 42, 328. [Google Scholar] [CrossRef] [PubMed]
- Youn, S.H.; Lee, J.S.; Lee, M.S.; Cha, E.Y.; Thuong, P.T.; Kim, J.R.; Chang, E.S. Anticancer properties of pomolic acid-induced AMP-activated protein kinase activation in MCF7 human breast cancer cells. Biol. Pharm. Bull. 2012, 35, 105. [Google Scholar] [CrossRef] [Green Version]
- Dai, W.; Dong, P.; Liu, J.; Gao, Y.; Hu, Y.; Lin, H.; Song, Y.; Mei, Q. Euscaphic acid inhibits proliferation and promotes apoptosis of nasopharyngeal carcinoma cells by silencing the PI3K/AKT/mTOR signaling pathway. Am. J. Transl. Res. 2019, 11, 2090. [Google Scholar] [PubMed]
- Jang, Y.G.; Ko, E.B.; Choi, K.C. Gallic acid, a phenolic acid, hinders the progression of prostate cancer by inhibition of histone deacetylase 1 and 2 expression. J. Nutr. Biochem. 2020, 84, 108444. [Google Scholar] [CrossRef] [PubMed]
- Civenni, G.; Iodice, M.; Nabavi, S.F.; Habtemariam, S.; Nabavi, S.M.; Catapano, C.V.; Daglia, M. Gallic acid and methyl-3-O-methyl gallate: A comparative study on their effects on prostate cancer stem cells. RSC Adv. 2015, 5, 63800. [Google Scholar] [CrossRef]
- Aborehab, N.M.; Elnagar, M.R.; Waly, N.E. Gallic acid potentiates the apoptotic effect of paclitaxel and carboplatin via overexpression of Bax and P53 on the MCF-7 human breast cancer cell line. J. Biochem. Mol. Toxicol. 2021, 35, e22638. [Google Scholar] [CrossRef]
- Wang, L.; Halquist, M.S.; Sweet, D.H. Simultaneous determination of gallic acid and gentisic acid in organic anion transporter expressing cells by liquid chromatography-tandem mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2013, 937, 91. [Google Scholar] [CrossRef]
- MassBank of North America (MoNA). Available online: https://mona.fiehnlab.ucdavis.edu/ (accessed on 12 May 2022).
- Jaiswal, R.; Matei, M.F.; Glembockyte, V.; Patras, M.A.; Kuhnert, N. Hierarchical key for the LC-MSn identification of all ten regio- and stereoisomers of caffeoylglucose. J. Agric. Food Chem. 2014, 62, 9252. [Google Scholar] [CrossRef]
- Stöggl, W.M.; Huck, C.W.; Bonn, G.K. Structural elucidation of catechin and epicatechin in sorrel leaf extracts using liquid-chromatography coupled to diode array-, fluorescence-, and mass spectrometric detection. J. Sep. Sci. 2004, 27, 524. [Google Scholar] [CrossRef]
- Jiamboonsri, P.; Pithayanukul, P.; Bavovada, R.; Gao, S.; Hu, M. A validated liquid chromatography-tandem mass spectrometry method for the determination of methyl gallate and pentagalloyl glucopyranose: Application to pharmacokinetic studies. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2015, 986–987, 12. [Google Scholar] [CrossRef]
- Tešević, V.; Aljančić, I.; Vajs, V.; Živković, M.B.; Nikićević, N.; Urosevic, I.; Vujisić, L.V. Development and validation of an LC-MS/MS method with multiple reactions monitoring mode for the quantification of vanillin and syringaldehyde in plum brandies. J. Serb. Chem. Soc. 2014, 79, 1537. [Google Scholar] [CrossRef]
- Mohammed, H.A.; Khan, R.A.; Abdel-Hafez, A.A.; Abdel-Aziz, M.; Ahmed, E.; Enany, S.; Mahgoub, S.; Al-Rugaie, O.; Alsharidah, M.; Aly, M.S.A.; et al. Phytochemical Profiling, In vitro and In Silico Anti-Microbial and Anti-Cancer Activity Evaluations and Staph GyraseB and h-TOP-IIβ Receptor-Docking Studies of Major Constituents of Zygophyllum coccineum L. Aqueous-Ethanolic Extract and Its Subsequent Fractions: An Approach to Validate Traditional Phytomedicinal Knowledge. Molecules 2021, 26, 577. [Google Scholar] [PubMed]
- Shen, Y.; Han, C.; Liu, B.; Lin, Z.; Zhou, X.; Wang, C.; Zhu, Z. Determination of vanillin, ethyl vanillin, and coumarin in infant formula by liquid chromatography-quadrupole linear ion trap mass spectrometry. J. Dairy Sci. 2014, 97, 679. [Google Scholar] [CrossRef] [PubMed]
- El Sayed, A.M.; Basam, S.M.; El-Naggar, E.B.A.; Marzouk, H.S.; El-Hawary, S. LC-MS/MS and GC-MS profiling as well as the antimicrobial effect of leaves of selected Yucca species introduced to Egypt. Sci. Rep. 2020, 10, 17778. [Google Scholar] [CrossRef] [PubMed]
- Cantos, E.; Espín, J.C.; Tomás-Barberán, F.A. Varietal differences among the polyphenol profiles of seven table grape cultivars studied by LC−DAD−MS−MS. J. Agric. Food Chem. 2002, 50, 5691. [Google Scholar] [CrossRef] [PubMed]
- Gad El-Hak, H.N.; Mahmoud, H.S.; Ahmed, E.A.; Elnegris, H.M.; Aldayel, T.S.; Abdelrazek, H.M.A.; Soliman, M.T.A.; El-Menyawy, M.A.I. Methanolic Phoenix dactylifera L. Extract Ameliorates Cisplatin-Induced Hepatic Injury in Male Rats. Nutrients 2022, 14, 1025. [Google Scholar] [CrossRef]
- Spáčil, Z.; Nováková, L.; Solich, P. Comparison of positive and negative ion detection of tea catechins using tandem mass spectrometry and ultra-high-performance liquid chromatography. Food Chem. 2010, 123, 535. [Google Scholar] [CrossRef]
- Montoro, P.; Tuberoso, C.I.G.; Perrone, A.; Piacente, S.; Cabras, P.; Pizza, C. Characterisation by liquid chromatography-electrospray tandem mass spectrometry of anthocyanins in extracts of Myrtus communis L. berries used for the preparation of myrtle liqueur. J. Chromatogr. A 2006, 1112, 232. [Google Scholar] [CrossRef]
- Lin, Q.; Li, Y.; Tan, X.M.; Yao, X.C. Simultaneous determination of formononetin, calycosin and isorhamnetin from Astragalus mongholicus in rat plasma by LC-MS/MS and application to pharmacokinetic study. J. Chin. Med. Mater. 2013, 36, 589. [Google Scholar]
- López-Salazar, H.; Camacho-Díaz, B.H.; Ávila-Reyes, S.V.; Pérez-García, M.D.; González-Cortazar, M.; Arenas Ocampo, M.L.; Jiménez-Aparicio, A.R. Identification and quantification of β-sitosterol β-D-glucoside of an ethanolic extract obtained by microwave-assisted extraction from Agave angustifolia Haw. Molecules 2019, 24, 3926. [Google Scholar] [CrossRef] [Green Version]
- Sut, S.; Poloniato, G.; Malagoli, M.; Dall’Acqua, S. Fragmentation of the main triterpene acids of apple by LC-APCI-MSn. J. Mass Spectrom. 2018, 53, 882. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Tao, S.; Zhang, S. Characterization and Quantification of Polyphenols and Triterpenoids in Thinned Young Fruits of Ten Pear Varieties by UPLC-Q TRAP-MS/MS. Molecules 2019, 24, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novotny, L.; Abdel-Hamid, M.E.; Hamza, H.; Masterova, I.; Grancai, D. Development of LC–MS method for determination of ursolic acid: Application to the analysis of ursolic acid in Staphylea holocarpa Hemsl. J. Pharm. Biomed. Anal. 2003, 31, 961. [Google Scholar] [CrossRef]
- Falev, D.I.; Ul’yanovskii, N.V.; Ovchinnikov, D.V.; Faleva, A.V.; Kosyakov, D.S. Screening and semi-quantitative determination of pentacyclic triterpenoids in plants by liquid chromatography-tandem mass spectrometry in precursor ion scan mode. Phytochem. Anal. 2021, 32, 252. [Google Scholar] [CrossRef]
- Mo, S.; Dong, L.; Hurst, W.J.; van Breemen, R.B. Quantitative analysis of phytosterols in edible oils using APCI liquid chromatography–tandem mass spectrometry. Lipids 2013, 48, 949. [Google Scholar] [CrossRef] [Green Version]
- Jiang, K.; Gachumi, G.; Poudel, A.; Shurmer, B.; Bashi, Z.; El-Aneed, A. The Establishment of Tandem Mass Spectrometric Fingerprints of Phytosterols and Tocopherols and the Development of Targeted Profiling Strategies in Vegetable Oils. J. Am. Soc. Mass. Spectrom. 2019, 30, 1700. [Google Scholar] [CrossRef]
- Münger, L.H.; Boulos, S.; Nyström, L. UPLC-MS/MS Based Identification of Dietary Steryl Glucosides by Investigation of Corresponding Free Sterols. Front. Chem. 2018, 6, 342. [Google Scholar] [CrossRef]
- Vinod Prabhu, V.; Elangovan, P.; Niranjali Devaraj, S.; Sakthivel, K.M. Targeting apoptosis by 1,2-diazole through regulation of EGFR, Bcl-2 and CDK-2 mediated signaling pathway in human non-small cell lung carcinoma A549 cells. Gene 2018, 679, 352. [Google Scholar] [CrossRef]
- Li, H.; Li, J.; Su, Y.; Fan, Y.; Guo, X.; Li, L.; Su, X.; Rong, R.; Ying, J.; Mo, X.; et al. A novel 3p22.3 gene CMTM7 represses oncogenic EGFR signaling and inhibits cancer cell growth. Oncogene 2014, 33, 3109. [Google Scholar] [CrossRef] [Green Version]










| Gene | Forward | Reverse |
|---|---|---|
| P53 | 5′-CCCCTCCTGGCCCCTGTCATCTTC-3′ | 5′-GCAGCGCCTCACAACCTCCGTCAT-3′ |
| BAX | 5′-GTTTCATCCAGGATCGAGCAG-3′ | 5′-CATCTTCTTCCAGATGGTGA-3′ |
| CASP-3 | 5′-TGGCCCTGAAATACGAAGTC-3′ | 5′-GGCAGTAGTCGACTCTGAAG-3′ |
| CASP-8 | 5′-AATGTTGGAGGAAAGCAAT-3′ | 5′-CATAGTCGTTGATTATCTTCAGC-3′ |
| CASP-9 | 5′-CGAACTAACAGGCAAGCAGC-3′ | 5′-ACCTCACCAAATCCTCCAGAAC-3′ |
| BCL2 | 5′-CCTGTGGATGACTGAGTACC-3′ | 5′-GAGACAGCCAGGAGAAATCA-3′ |
| β-actin | 5′-GTGACATCCACACCCAGAGG-3′ | 5′-ACAGGATGTCAAAACTGCCC-3′ |
| Sample | DPPH (IC50 in µg/mL) | FRAP (mMol Fe+2/g) | TAC (mg GAE/g) |
|---|---|---|---|
| P. acacia | 60.70 ± 4.28 a | 1.29 ± 0.31 c | 32.67 ± 2.81 c |
| P. curviflorus | 48.28 ± 3.41 b | 1.43 ± 0.54 c | 41.89 ± 3.15 b |
| Ascorbic acid | 10.64 ± 0.82 c | 3.14 ± 0.82 b | 69.23 ± 4.51 a |
| BHT | - | 8.07 ± 0.79 a | - |
| ANOVA (p-value) | <0.001 *** | <0.001 *** | <0.001 *** |
| Sample | IC50 (μg/mL) | |||
|---|---|---|---|---|
| Prostate | Breast | Ovarian | Lung | |
| PC-3 | MDA-MB-231 | A2780 | A549 | |
| P. acacia | 34.12 ± 1.3 a | 86.5 ± 2.01 | NA | 50.6 ± 1.63 |
| P. curviflorus | 25.83 ± 1.2 b | NA | 76.5 ± 1.78 | NA |
| Doxorubicin | 8.23 ± 0.56 c | 7.07 ± 0.64 | 10.63 ± 0.76 | 9.26 ± 0.64 |
| ANOVA (p-value) | <0.001 ***F | <0.001 ***T | <0.001 ***T | <0.001 ***T |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eltamany, E.E.; Goda, M.S.; Nafie, M.S.; Abu-Elsaoud, A.M.; Hareeri, R.H.; Aldurdunji, M.M.; Elhady, S.S.; Badr, J.M.; Eltahawy, N.A. Comparative Assessment of the Antioxidant and Anticancer Activities of Plicosepalus acacia and Plicosepalus curviflorus: Metabolomic Profiling and In Silico Studies. Antioxidants 2022, 11, 1249. https://doi.org/10.3390/antiox11071249
Eltamany EE, Goda MS, Nafie MS, Abu-Elsaoud AM, Hareeri RH, Aldurdunji MM, Elhady SS, Badr JM, Eltahawy NA. Comparative Assessment of the Antioxidant and Anticancer Activities of Plicosepalus acacia and Plicosepalus curviflorus: Metabolomic Profiling and In Silico Studies. Antioxidants. 2022; 11(7):1249. https://doi.org/10.3390/antiox11071249
Chicago/Turabian StyleEltamany, Enas E., Marwa S. Goda, Mohamed S. Nafie, Abdelghafar M. Abu-Elsaoud, Rawan H. Hareeri, Mohammed M. Aldurdunji, Sameh S. Elhady, Jihan M. Badr, and Nermeen A. Eltahawy. 2022. "Comparative Assessment of the Antioxidant and Anticancer Activities of Plicosepalus acacia and Plicosepalus curviflorus: Metabolomic Profiling and In Silico Studies" Antioxidants 11, no. 7: 1249. https://doi.org/10.3390/antiox11071249