
Protective Effects of Diets Rich in Polyphenols in Cigarette Smoke (CS)-Induced Oxidative Damages and Associated Health Implications

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Cigarette Smoking, Oxidative Damages and Health Implications
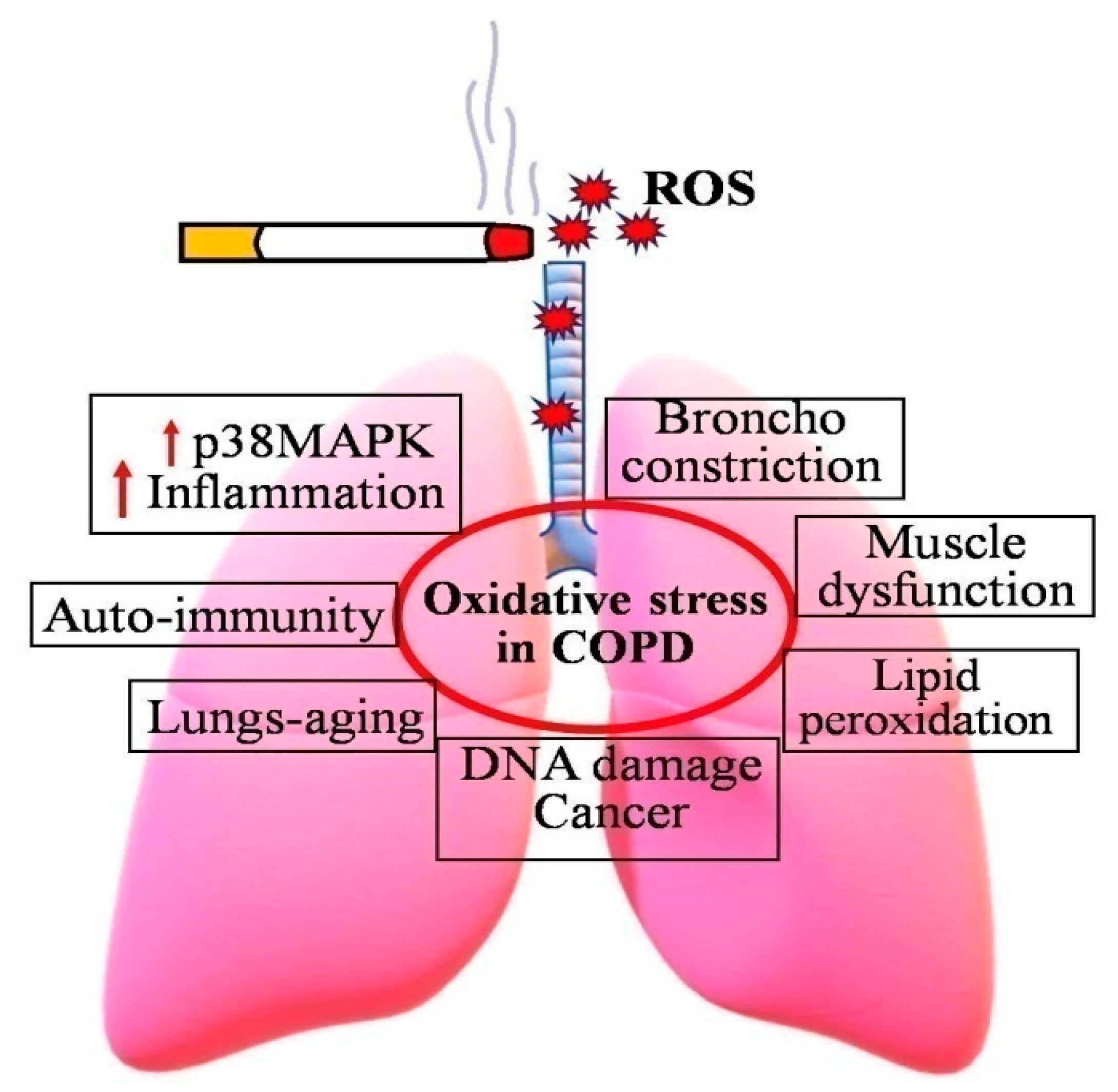
2.1. Cigarette Smoking and Chronic Obstructive Pulmonary Disease (COPD)
2.1.1. COPD and Inflammation
2.1.2. COPD and Autoimmunity
2.1.3. COPD and DNA Damage
2.2. Cigarette Smoking and Lipoprotein Oxidation
2.3. Cigarette Smoking and Abnormal Nitric Oxide (NO) Metabolism
2.4. Cigarette Smoking and Thrombogenesis
2.5. Cigarette Smoking and Endothelial Dysfunction
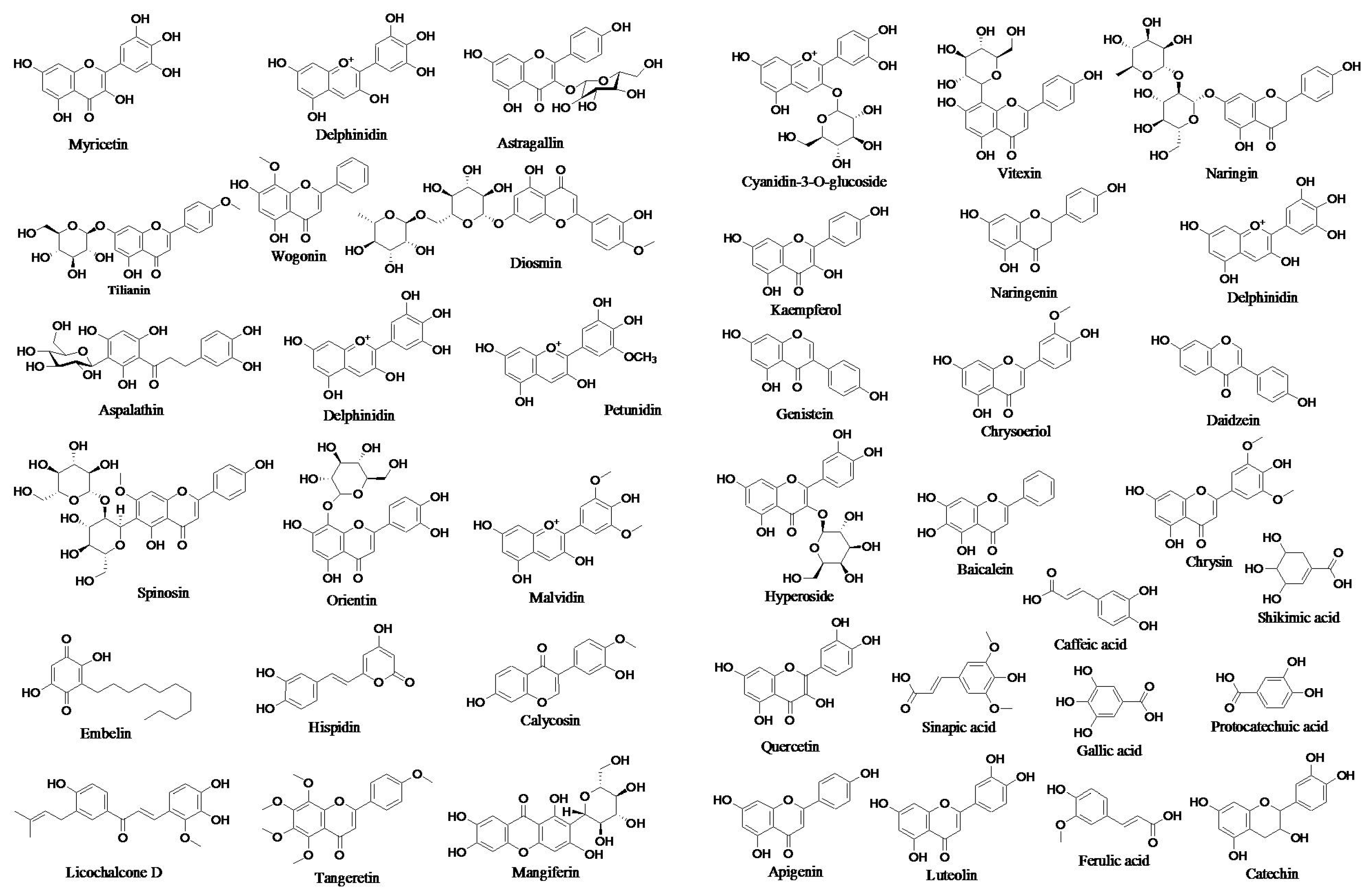
3. Polyphenols: Dietary Sources, Chemistry and Medicinal Importance
4. Protective Effects of Dietary Polyphenols in CS-Induced Diseases
4.1. Vaso-Protective Effect
4.2. Cardioprotective Effect
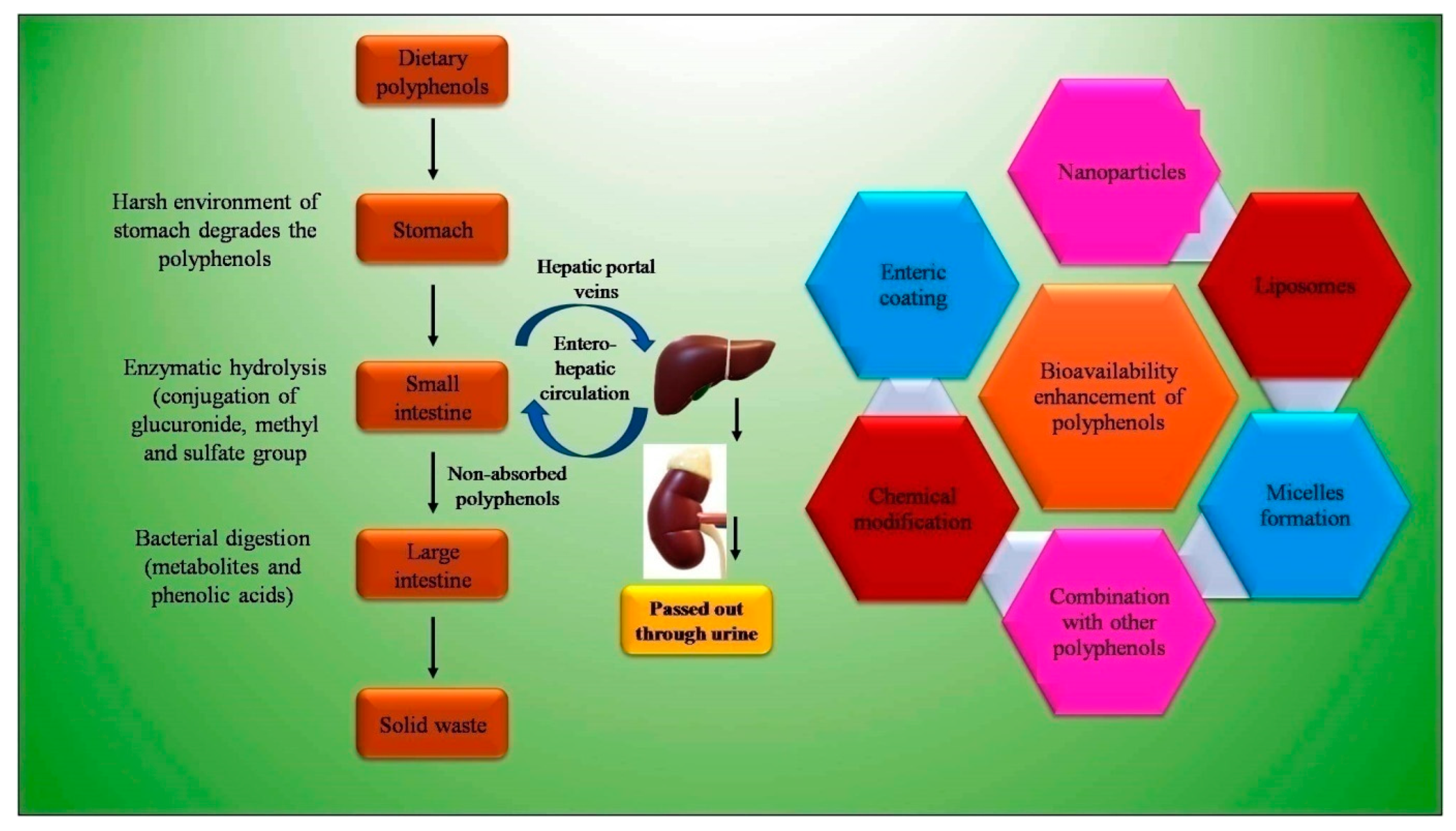
5. Bioavailability Issues of Dietary Polyphenols and Strategies to Overcome
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. WHO Framework Convention on Tobacco Control; WHO: Geneva, Switzerland, 2003; ISBN 9241591013.
- GBD 2015 Chronic Respiratory Disease Collaborators. Global, Regional, and National Deaths, Prevalence, Disability-Adjusted Life Years, and Years Lived with Disability for Chronic Obstructive Pulmonary Disease and Asthma, 1990–2015: A Systematic Analysis for the Global Burden of Disease Study 2015. Lancet. Respir. Med. 2017, 5, 691. [Google Scholar] [CrossRef] [Green Version]
- Gershon, A.S.; Warner, L.; Cascagnette, P.; Victor, J.C.; To, T. Lifetime Risk of Developing Chronic Obstructive Pulmonary Disease: A Longitudinal Population Study. Lancet 2011, 378, 991–996. [Google Scholar] [CrossRef]
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y. Global and Regional Mortality from 235 Causes of Death for 20 Age Groups in 1990 and 2010: A Systematic Analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Panda, K.; Chattopadhyay, R.; Ghosh, M.K.; Chattopadhyay, D.J.; Chatterjee, I.B. Vitamin C Prevents Cigarette Smoke Induced Oxidative Damage of Proteins and Increased Proteolysis. Free Radic. Biol. Med. 1999, 27, 1064–1079. [Google Scholar] [CrossRef]
- Church, D.F.; Pryor, W.A. Free-Radical Chemistry of Cigarette Smoke and Its Toxicological Implications. Environ. Health Perspect. 1985, 64, 111–126. [Google Scholar] [CrossRef]
- Frei, B.; Forte, T.M.; Ames, B.N.; Cross, C.E. Gas Phase Oxidants of Cigarette Smoke Induce Lipid Peroxidation and Changes in Lipoprotein Properties in Human Blood Plasma. Protective Effects of Ascorbic Acid. Biochem. J. 1991, 277, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Cross, C.E.; O’Neill, C.A.; Reznick, A.Z.; Hu, M.-L.; Marcocci, L.; Packer, L.; Frei, B. Cigarette Smoke Oxidation of Human Plasma Constituents. Ann. N. Y. Acad. Sci. 1993, 686, 72–89. [Google Scholar] [CrossRef]
- Ueyama, K.; Yokode, M.; Arai, H.; Nagano, Y.; Zhi-Xiang, L.; Cho, M.; Kita, T. Cholesterol Efflux Effect of High Density Lipoprotein Is Impaired by Whole Cigarette Smoke Extracts through Lipid Peroxidation. Free Radic. Biol. Med. 1998, 24, 182–190. [Google Scholar] [CrossRef]
- Shah, P.K.; Helfant, R.H. Smoking and Coronary Artery Disease. Chest 1988, 94, 449–452. [Google Scholar] [CrossRef]
- Bartecchi, C.E.; MacKenzie, T.D.; Schrier, R.W. The Human Costs of Tobacco Use. N. Engl. J. Med. 1994, 330, 907–912. [Google Scholar] [CrossRef]
- Becker, L.B. New Concepts in Reactive Oxygen Species and Cardiovascular Reperfusion Physiology. Cardiovasc. Res. 2004, 61, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; McLellan, L.I. Glutathione and Glutathione-Dependent Enzymes Represent a Co-Ordinately Regulated Defence against Oxidative Stress. Free Radic. Res. 1999, 31, 273–300. [Google Scholar] [CrossRef] [PubMed]
- Masella, R.; Di Benedetto, R.; Varì, R.; Filesi, C.; Giovannini, C. Novel Mechanisms of Natural Antioxidant Compounds in Biological Systems: Involvement of Glutathione and Glutathione-Related Enzymes. J. Nutr. Biochem. 2005, 16, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Hartman, R.E.; Shah, A.; Fagan, A.M.; Schwetye, K.E.; Parsadanian, M.; Schulman, R.N.; Finn, M.B.; Holtzman, D.M. Pomegranate Juice Decreases Amyloid Load and Improves Behavior in a Mouse Model of Alzheimer’s Disease. Neurobiol. Dis. 2006, 24, 506–515. [Google Scholar] [CrossRef]
- Hollman, P.C.H.; Van Trijp, J.M.P.; Buysman, M.N.C.P.; van der Gaag, M.S.; Mengelers, M.J.B.; De Vries, J.H.M.; Katan, M.B. Relative Bioavailability of the Antioxidant Flavonoid Quercetin from Various Foods in Man. FEBS Lett. 1997, 418, 152–156. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.-Q.; Zhang, Y.; Xiang, J.-J.; Xiong, C.-L. Protective Effect of Curcumin against Liver Warm Ischemia/Reperfusion Injury in Rat Model Is Associated with Regulation of Heat Shock Protein and Antioxidant Enzymes. World J. Gastroenterol. WJG 2007, 13, 1953. [Google Scholar] [CrossRef] [Green Version]
- Molina, M.F.; Sanchez-Reus, I.; Iglesias, I.; Benedi, J. Quercetin, a Flavonoid Antioxidant, Prevents and Protects against Ethanol-Induced Oxidative Stress in Mouse Liver. Biol. Pharm. Bull. 2003, 26, 1398–1402. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Yu, R.; Owuor, E.D.; Tony Kong, A.-N. Activation of Antioxidant-Response Element (ARE), Mitogen-Activated Protein Kinases (MAPKs) and Caspases by Major Green Tea Polyphenol Components during Cell Survival and Death. Arch. Pharm. Res. 2000, 23, 605–612. [Google Scholar] [CrossRef]
- Polia, F.; Pastor-Belda, M.; Martínez-Blázquez, A.; Horcajada, M.-N.; Tomás-Barberán, F.A.; García-Villalba, R. Technological and Biotechnological Processes to Enhance the Bioavailability of Dietary (Poly) Phenols in Humans. J. Agric. Food Chem. 2022, 70, 2092–2107. [Google Scholar] [CrossRef]
- Lawson, L.D.; Hunsaker, S.M. Allicin Bioavailability and Bioequivalence from Garlic Supplements and Garlic Foods. Nutrients 2018, 10, 812. [Google Scholar] [CrossRef] [Green Version]
- Dei Cas, M.; Ghidoni, R. Dietary Curcumin: Correlation between Bioavailability and Health Potential. Nutrients 2019, 11, 2147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, A.; FitzGerald, G.A. Oxidative Stress and Smoking-Induced Vascular Injury. Prog. Cardiovasc. Dis. 2003, 46, 79–90. [Google Scholar] [CrossRef]
- Repine, J.E.; Bast, A.; Lankhorst, I.D.A.; Group, O.S.S. Oxidative Stress in Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 1997, 156, 341–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paudel, K.R.; Panth, N.; Manandhar, B.; Singh, S.K.; Gupta, G.; Wich, P.R.; Nammi, S.; MacLoughlin, R.; Adams, J.; Warkiani, M.E.; et al. Attenuation of Cigarette-Smoke-Induced Oxidative Stress, Senescence, and Inflammation by Berberine-Loaded Liquid Crystalline Nanoparticles: In Vitro Study in 16HBE and RAW264.7 Cells. Antioxidants 2022, 11, 873. [Google Scholar] [CrossRef] [PubMed]
- Kouzarides, T. Histone Acetylases and Deacetylases in Cell Proliferation. Curr. Opin. Genet. Dev. 1999, 9, 40–48. [Google Scholar] [CrossRef]
- Chung, S.; Sundar, I.K.; Yao, H.; Ho, Y.-S.; Rahman, I. Glutaredoxin 1 Regulates Cigarette Smoke-Mediated Lung Inflammation through Differential Modulation of IκB Kinases in Mice: Impact on Histone Acetylation. Am. J. Physiol. Cell. Mol. Physiol. 2010, 299, L192–L203. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-Y.; Ito, K.; Hayashi, R.; Jazrawi, E.P.I.; Barnes, P.J.; Adcock, I.M. NF-kB and Activator Protein 1 Response Elements and the Role of Histone Modifications in IL-1β-Induced TGF-Β1 Gene Transcription. J. Immunol. 2006, 176, 603–615. [Google Scholar] [CrossRef] [Green Version]
- Adler, V.; Yin, Z.; Tew, K.D.; Ronai, Z. Role of Redox Potential and Reactive Oxygen Species in Stress Signaling. Oncogene 1999, 18, 6104–6111. [Google Scholar] [CrossRef] [Green Version]
- Rahman, I.; MacNee, W. Role of Transcription Factors in Inflammatory Lung Diseases. Thorax 1998, 53, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Marwick, J.A.; Kirkham, P.A.; Stevenson, C.S.; Danahay, H.; Giddings, J.; Butler, K.; Donaldson, K.; MacNee, W.; Rahman, I. Cigarette Smoke Alters Chromatin Remodeling and Induces Proinflammatory Genes in Rat Lungs. Am. J. Respir. Cell Mol. Biol. 2004, 31, 633–642. [Google Scholar] [CrossRef] [Green Version]
- Krieg, A.J.; Rankin, E.B.; Chan, D.; Razorenova, O.; Fernandez, S.; Giaccia, A.J. Regulation of the Histone Demethylase JMJD1A by Hypoxia-Inducible Factor 1α Enhances Hypoxic Gene Expression and Tumor Growth. Mol. Cell. Biol. 2010, 30, 344–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogg, J.C.; Timens, W. The Pathology of Chronic Obstructive Pulmonary Disease. Annu. Rev. Pathol. Mech. Dis. 2009, 4, 435–459. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; MacNee, W. Role of Oxidants/Antioxidants in Smoking-Induced Lung Diseases. Free Radic. Biol. Med. 1996, 21, 669–681. [Google Scholar] [CrossRef]
- Zhang, M.; Nomura, A.; Uchida, Y.; Iijima, H.; Sakamoto, T.; Iishii, Y.; Morishima, Y.; Mochizuki, M.; Masuyama, K.; Hirano, K. Ebselen Suppresses Late Airway Responses and Airway Inflammation in Guinea Pigs. Free Radic. Biol. Med. 2002, 32, 454–464. [Google Scholar] [CrossRef]
- Haddad, E.-B.; McCluskie, K.; Birrell, M.A.; Dabrowski, D.; Pecoraro, M.; Underwood, S.; Chen, B.; De Sanctis, G.T.; Webber, S.E.; Foster, M.L. Differential Effects of Ebselen on Neutrophil Recruitment, Chemokine, and Inflammatory Mediator Expression in a Rat Model of Lipopolysaccharide-Induced Pulmonary Inflammation. J. Immunol. 2002, 169, 974–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, P.J. The Cytokine Network in Chronic Obstructive Pulmonary Disease. Am. J. Respir. Cell Mol. Biol. 2009, 41, 631–638. [Google Scholar] [CrossRef]
- Gorowiec, M.R.; Borthwick, L.A.; Parker, S.M.; Kirby, J.A.; Saretzki, G.C.; Fisher, A.J. Free Radical Generation Induces Epithelial-to-Mesenchymal Transition in Lung Epithelium via a TGF-Β1-Dependent Mechanism. Free Radic. Biol. Med. 2012, 52, 1024–1032. [Google Scholar] [CrossRef]
- Sukkar, M.B.; Bhavsar, P.K.; Chung, K.F. TGF-Regulates Nox4, MnSOD and Catalase Expression, and IL-6 Release in Airway Smooth Muscle Cells. Am. J. Physiol. Cell. Mol. Physiol. 2011, 300, L295–L304. [Google Scholar]
- Barnes, P.J.; Baker, J.; Donnelly, L.E. Cellular Senescence as a Mechanism and Target in Chronic Lung Diseases. Am. J. Respir. Crit. Care Med. 2019, 200, 556–564. [Google Scholar] [CrossRef]
- Nakamaru, Y.; Vuppusetty, C.; Wada, H.; Milne, J.C.; Ito, M.; Rossios, C.; Elliot, M.; Hogg, J.; Kharitonov, S.; Goto, H. A Protein Deacetylase SIRT1 Is a Negative Regulator of Metalloproteinase-9. FASEB J. 2009, 23, 2810–2819. [Google Scholar] [CrossRef]
- Birch, J.; Barnes, P.J.; Passos, J.F. Mitochondria, Telomeres and Cell Senescence: Implications for Lung Ageing and Disease. Pharmacol. Ther. 2018, 183, 34–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feghali-Bostwick, C.A.; Gadgil, A.S.; Otterbein, L.E.; Pilewski, J.M.; Stoner, M.W.; Csizmadia, E.; Zhang, Y.; Sciurba, F.C.; Duncan, S.R. Autoantibodies in Patients with Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2008, 177, 156–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkham, P.A.; Caramori, G.; Casolari, P.; Papi, A.A.; Edwards, M.; Shamji, B.; Triantaphyllopoulos, K.; Hussain, F.; Pinart, M.; Khan, Y. Oxidative Stress–Induced Antibodies to Carbonyl-Modified Protein Correlate with Severity of Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2011, 184, 796–802. [Google Scholar] [CrossRef] [Green Version]
- Caramori, G.; Adcock, I.M.; Casolari, P.; Ito, K.; Jazrawi, E.; Tsaprouni, L.; Villetti, G.; Civelli, M.; Carnini, C.; Chung, K.F. Unbalanced Oxidant-Induced DNA Damage and Repair in COPD: A Link towards Lung Cancer. Thorax 2011, 66, 521–527. [Google Scholar] [CrossRef] [Green Version]
- Adcock, I.M.; Caramori, G.; Barnes, P.J. Chronic Obstructive Pulmonary Disease and Lung Cancer: New Molecular Insights. Respiration 2011, 81, 265–284. [Google Scholar] [CrossRef] [PubMed]
- Chisolm, G.M.; Steinberg, D. The Oxidative Modification Hypothesis of Atherogenesis: An Overview. Free Radic. Biol. Med. 2000, 28, 1815–1826. [Google Scholar] [CrossRef]
- Reilly, M.P.; Praticò, D.; Delanty, N.; DiMinno, G.; Tremoli, E.; Rader, D.; Kapoor, S.; Rokach, J.; Lawson, J.; FitzGerald, G.A. Increased Formation of Distinct F2 Isoprostanes in Hypercholesterolemia. Circulation 1998, 98, 2822–2828. [Google Scholar] [CrossRef] [Green Version]
- De Caterina, R.; Cipollone, F.; Filardo, F.P.; Zimarino, M.; Bernini, W.; Lazzerini, G.; Bucciarelli, T.; Falco, A.; Marchesani, P.; Muraro, R. Low-Density Lipoprotein Level Reduction by the 3-Hydroxy-3-Methylglutaryl Coenzyme-A Inhibitor Simvastatin Is Accompanied by a Related Reduction of F2-Isoprostane Formation in Hypercholesterolemic Subjects: No Further Effect of Vitamin E. Circulation 2002, 106, 2543–2549. [Google Scholar] [CrossRef] [Green Version]
- Kagota, S.; Yamaguchi, Y.; Shinozuka, K.; Kwon, Y.M.; Kunitomo, M. Cigarette Smoke-Modified Low Density Lipoprotein Impairs Endothelium-Dependent Relaxation in Isolated Rabbit Arteries. Gen. Pharmacol. 1996, 27, 477–481. [Google Scholar] [CrossRef]
- Gouazé, V.; Dousset, N.; Dousset, J.-C.; Valdiguié, P. Effect of Nicotine and Cotinine on the Susceptibility to in vitro Oxidation of LDL in Healthy Non Smokers and Smokers. Clin. Chim. Acta 1998, 277, 25–37. [Google Scholar] [CrossRef]
- Scheffler, E.; Wiest, E.; Woehrle, J.; Otto, I.; Schulz, I.; Huber, L.; Ziegler, R.; Dresel, H.A. Smoking Influences the Atherogenic Potential of Low-Density Lipoprotein. Clin. Investig. 1992, 70, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Harats, D.; Ben-Naim, M.; Dabach, Y.; Hollander, G.; Stein, O.; Stein, Y. Cigarette Smoking Renders LDL Susceptible to Peroxidative Modification and Enhanced Metabolism by Macrophages. Atherosclerosis 1989, 79, 245–252. [Google Scholar] [CrossRef]
- Santanam, N.; Sanchez, R.; Hendler, S.; Parthasarathy, S. Aqueous Extracts of Cigarette Smoke Promote the Oxidation of Low Density Lipoprotein by Peroxidases. FEBS Lett. 1997, 414, 549–551. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, A.; Kondo, K.; Sakamoto, Y.; Kurata, H.; Itakura, H.; Ikeda, Y. Smoking Cessation Increases the Resistance of Low-Density Lipoprotein to Oxidation. Atherosclerosis 1997, 130, 109–111. [Google Scholar] [CrossRef]
- Tsuchiya, M.; Asada, A.; Kasahara, E.; Sato, E.F.; Shindo, M.; Inoue, M. Smoking a Single Cigarette Rapidly Reduces Combined Concentrations of Nitrate and Nitrite and Concentrations of Antioxidants in Plasma. Circulation 2002, 105, 1155–1157. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 2015; ISBN 0198717482. [Google Scholar]
- Vayssier-Taussat, M.; Camilli, T.; Aron, Y.; Meplan, C.; Hainaut, P.; Polla, B.S.; Weksler, B. Effects of Tobacco Smoke and Benzo [a] Pyrene on Human Endothelial Cell and Monocyte Stress Responses. Am. J. Physiol. Circ. Physiol. 2001, 280, H1293–H1300. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Harrison, D.G. Endothelial Dysfunction in Cardiovascular Diseases: The Role of Oxidant Stress. Circ. Res. 2000, 87, 840–844. [Google Scholar] [CrossRef] [Green Version]
- Karim, Z.A.; Alshbool, F.Z.; Vemana, H.P.; Adhami, N.; Dhall, S.; Espinosa, E.V.P.; Martins-Green, M.; Khasawneh, F.T. Third-Hand Smoke: Impact on Hemostasis and Thrombogenesis. J. Cardiovasc. Pharmacol. 2015, 66, 177–182. [Google Scholar] [CrossRef]
- Levine, P.H. An Acute Effect of Cigarette Smoking on Platelet Function: A Possible Link between Smoking and Arterial Thrombosis. Circulation 1973, 48, 619–623. [Google Scholar] [CrossRef] [Green Version]
- Rångemark, C.; Ciabattoni, G.; Wennmalm, A. Excretion of Thromboxane Metabolites in Healthy Women after Cessation of Smoking. Arterioscler. Thromb. A J. Vasc. Biol. 1993, 13, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Mirshahi, F.; Vasse, M.; Vincent, L.; Trochon, V.; Pourtau, J.; Vannier, J.P.; Li, H.; Soria, J.; Soria, C. Fibrinogen: A Vascular Risk Factor, Why? Contributing Effect of Oncostatin M on Both Fibrinogen Biosynthesis by Hepatocytes and Participation in Atherothrombotic Risk Related to Modifications of Endothelial Cells. Ann. N. Y. Acad. Sci. 2001, 936, 621–624. [Google Scholar] [CrossRef] [PubMed]
- Wilhelmsen, L.; Svärdsudd, K.; Korsan-Bengtsen, K.; Larsson, B.O.; Welin, L.; Tibblin, G. Fibrinogen as a Risk Factor for Stroke and Myocardial Infarction. N. Engl. J. Med. 1984, 311, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Stone, M.C.; Thorp, J.M. Plasma Fibrinogen—A Major Coronary Risk Factor. J. R. Coll. Gen. Pract. 1985, 35, 565. [Google Scholar]
- Tuut, M.; Hense, H.-W. Smoking, Other Risk Factors and Fibrinogen Levels: Evidence of Effect Modification. Ann. Epidemiol. 2001, 11, 232–238. [Google Scholar] [CrossRef]
- Cook, D.G.; Cappuccio, F.P.; Atkinson, R.W.; Wicks, P.D.; Chitolie, A.; Nakandakare, E.R.; Sagnella, G.A.; Humphries, S.E. Ethnic Differences in Fibrinogen Levels: The Role of Environmental Factors and the β-Fibrinogen Gene. Am. J. Epidemiol. 2001, 153, 799–806. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.E.; Green, F.R.; Lamlum, H.; Humphries, S.E. The Association of Combined Alpha and Beta Fibrinogen Genotype on Plasma Fibrinogen Levels in Smokers and Non-Smokers. J. Med. Genet. 1995, 32, 585–589. [Google Scholar] [CrossRef]
- Gole, M.D.; Souza, J.M.; Choi, I.; Hertkorn, C.; Malcolm, S.; Foust III, R.F.; Finkel, B.; Lanken, P.N.; Ischiropoulos, H. Plasma Proteins Modified by Tyrosine Nitration in Acute Respiratory Distress Syndrome. Am. J. Physiol. Cell. Mol. Physiol. 2000, 278, L961–L967. [Google Scholar] [CrossRef]
- Barua, R.S.; Ambrose, J.A.; Saha, D.C.; Eales-Reynolds, L.-J. Smoking Is Associated with Altered Endothelial-Derived Fibrinolytic and Antithrombotic Factors: An in vitro Demonstration. Circulation 2002, 106, 905–908. [Google Scholar] [CrossRef] [Green Version]
- Sambola, A.; Osende, J.; Hathcock, J.; Degen, M.; Nemerson, Y.; Fuster, V.; Crandall, J.; Badimon, J.J. Role of Risk Factors in the Modulation of Tissue Factor Activity and Blood Thrombogenicity. Circulation 2003, 107, 973–977. [Google Scholar] [CrossRef] [Green Version]
- Busacca, M.; Balconi, G.; Pietra, A.; Vergara-Dauden, M.; de Gaetano, G.; Dejana, E. Maternal Smoking and Prostacyclin Production by Cultured Endothelial Cells from Umbilical Arteries. Am. J. Obstet. Gynecol. 1984, 148, 1127–1130. [Google Scholar] [CrossRef]
- Matetzky, S.; Tani, S.; Kangavari, S.; Dimayuga, P.; Yano, J.; Xu, H.; Chyu, K.-Y.; Fishbein, M.C.; Shah, P.K.; Cercek, B. Smoking Increases Tissue Factor Expression in Atherosclerotic Plaques: Implications for Plaque Thrombogenicity. Circulation 2000, 102, 602–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celermajer, D.S.; Sorensen, K.E.; Gooch, V.M.; Spiegelhalter, D.J.; Miller, O.I.; Sullivan, I.D.; Lloyd, J.K.; Deanfield, J.E. Non-Invasive Detection of Endothelial Dysfunction in Children and Adults at Risk of Atherosclerosis. Lancet 1992, 340, 1111–1115. [Google Scholar] [CrossRef]
- Furchgott, R.F.; Zawadzki, J. V The Obligatory Role of Endothelial Cells in the Relaxation of Arterial Smooth Muscle by Acetylcholine. Nature 1980, 288, 373–376. [Google Scholar] [CrossRef]
- Schächinger, V.; Britten, M.B.; Zeiher, A.M. Prognostic Impact of Coronary Vasodilator Dysfunction on Adverse Long-Term Outcome of Coronary Heart Disease. Circulation 2000, 101, 1899–1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barua, R.S.; Ambrose, J.A.; Eales-Reynolds, L.-J.; DeVoe, M.C.; Zervas, J.G.; Saha, D.C. Heavy and Light Cigarette Smokers Have Similar Dysfunction of Endothelial Vasoregulatory Activity: An in vivo and in vitro Correlation. J. Am. Coll. Cardiol. 2002, 39, 1758–1763. [Google Scholar] [CrossRef] [Green Version]
- Lekakis, J.; Papamichael, C.; Vemmos, C.; Nanas, J.; Kontoyannis, D.; Stamatelopoulos, S.; Moulopoulos, S. Effect of Acute Cigarette Smoking on Endothelium-Dependent Brachial Artery Dilatation in Healthy Individuals. Am. J. Cardiol. 1997, 79, 529–531. [Google Scholar] [CrossRef]
- Celermajer, D.S.; Adams, M.R.; Clarkson, P.; Robinson, J.; McCredie, R.; Donald, A.; Deanfield, J.E. Passive Smoking and Impaired Endothelium-Dependent Arterial Dilatation in Healthy Young Adults. N. Engl. J. Med. 1996, 334, 150–155. [Google Scholar] [CrossRef]
- Tresserra-Rimbau, A.; Lamuela-Raventos, R.M.; Moreno, J.J. Polyphenols, Food and Pharma. Current Knowledge and Directions for Future Research. Biochem. Pharmacol. 2018, 156, 186–195. [Google Scholar] [CrossRef]
- Ștefan, M.; Bîrsă, M.L. Flavonoids–An Amazing Group of Compounds with Potent Antimicrobial Properties. Mem. Sci. Sect. Rom. Acad. 2019, 42. [Google Scholar]
- Durazzo, A.; Lucarini, M.; Souto, E.B.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E.; Santini, A. Polyphenols: A Concise Overview on the Chemistry, Occurrence, and Human Health. Phyther. Res. 2019, 33, 2221–2243. [Google Scholar] [CrossRef] [Green Version]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; Deb, P.K.; Piya, S.; Medina, K.D.; Devi, R.; Walode, S.G.; Rudrapal, M. Dietary Flavonoids: Cardioprotective potential with antioxidant effects and their pharmacokinetic/therapeutic concerns. Molecules 2021, 26, 4021. [Google Scholar] [CrossRef] [PubMed]
- Rudrapal, M.; Khairnar, S.J.; Dukhyil, A.A.B.; Khan, J.; Alaidarous, M.; Palai, S.; Deb, P.K.; Bhattacharjee, S.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights into Protective Effects, Antioxidant Potential and Mechanism of Action. Front. Pharmacol. 2022, 13, 806470. [Google Scholar] [CrossRef]
- Rudrapal, M.; Khan, J.; Dukhyil, A.A.B.; Alarousy, R.M.I.I.; Emmanuel, I.A.; Sharma, T.; Khairnar, S.J.; Bendale, A.R. Chalcone Scaffolds, Precursors of Flavonoids, Chemistry, Bioactivity and Pharmacokinetics. Molecules 2021, 26, 7177. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, 162750. [Google Scholar] [CrossRef] [Green Version]
- Cook, N.C.; Samman, S. Flavonoids—Chemistry, Metabolism, Cardioprotective Effects, and Dietary Sources. J. Nutr. Biochem. 1996, 7, 66–76. [Google Scholar] [CrossRef]
- Santos, E.L.; Maia, B.; Ferriani, A.P.; Teixeira, S.D. Flavonoids: Classification, Biosynthesis and Chemical Ecology. In Flavonoids—From Biosynthesis to Human Health; InTechOpen: London, UK, 2017; Volume 13, pp. 78–94. [Google Scholar]
- Silva, A.S.; Reboredo-Rodríguez, P.; Süntar, I.; Sureda, A.; Belwal, T.; Loizzo, M.R.; Tundis, R.; Sobarzo-Sanchez, E.; Rastrelli, L.; Forbes-Hernandez, T.Y. Evaluation of the Status Quo of Polyphenols Analysis: Part I—Phytochemistry, Bioactivity, Interactions, and Industrial Uses. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3191–3218. [Google Scholar] [CrossRef]
- Corcoran, M.P.; McKay, D.L.; Blumberg, J.B. Flavonoid Basics: Chemistry, Sources, Mechanisms of Action, and Safety. J. Nutr. Gerontol. Geriatr. 2012, 31, 176–189. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and Anthocyanins: Colored Pigments as Food, Pharmaceutical Ingredients, and the Potential Health Benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, C.; Zhang, W.; Sheng, C.; Zhang, W.; Xing, C.; Miao, Z. Chalcone: A Privileged Structure in Medicinal Chemistry. Chem. Rev. 2017, 117, 7762–7810. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.M. A Review of Potential Health Benefits of Flavonoids. Plant Foods Hum. Nutr. 2008, 59, 113–122. [Google Scholar]
- Amic, D.; Davidovic-Amic, D.; Beslo, D.; Rastija, V.; Lucic, B.; Trinajstic, N. SAR and QSAR of the Antioxidant Activity of Flavonoids. Curr. Med. Chem. 2007, 14, 827–845. [Google Scholar] [CrossRef] [PubMed]
- Velagapudi, R.; El-Bakoush, A.; Olajide, O.A. Activation of Nrf2 Pathway Contributes to Neuroprotection by the Dietary Flavonoid Tiliroside. Mol. Neurobiol. 2018, 55, 8103–8123. [Google Scholar] [CrossRef] [Green Version]
- Romanova, D.; Vachalkova, A.; Cipak, L.; Ovesna, Z.; Rauko, P. Study of Antioxidant Effect of Apigenin, Luteolin and Quercetin by DNA Protective Method. Neoplasma 2001, 48, 104–107. [Google Scholar]
- Espley, R.V.; Butts, C.A.; Laing, W.A.; Martell, S.; Smith, H.; McGhie, T.K.; Zhang, J.; Paturi, G.; Hedderley, D.; Bovy, A. Dietary Flavonoids from Modified Apple Reduce Inflammation Markers and Modulate Gut Microbiota in Mice. J. Nutr. 2014, 144, 146–154. [Google Scholar] [CrossRef] [Green Version]
- Song, Q.; Chen, P.; Liu, X.-M. The Role of Cigarette Smoke-Induced Pulmonary Vascular Endothelial Cell Apoptosis in COPD. Respir. Res. 2021, 22, 1–15. [Google Scholar] [CrossRef]
- Barnes, P.J.; Di Shapiro, S.; Pauwels, R.A. Chronic Obstructive Pulmonary Disease: Molecular and Cellularmechanisms. Eur. Respir. J. 2003, 22, 672–688. [Google Scholar] [CrossRef]
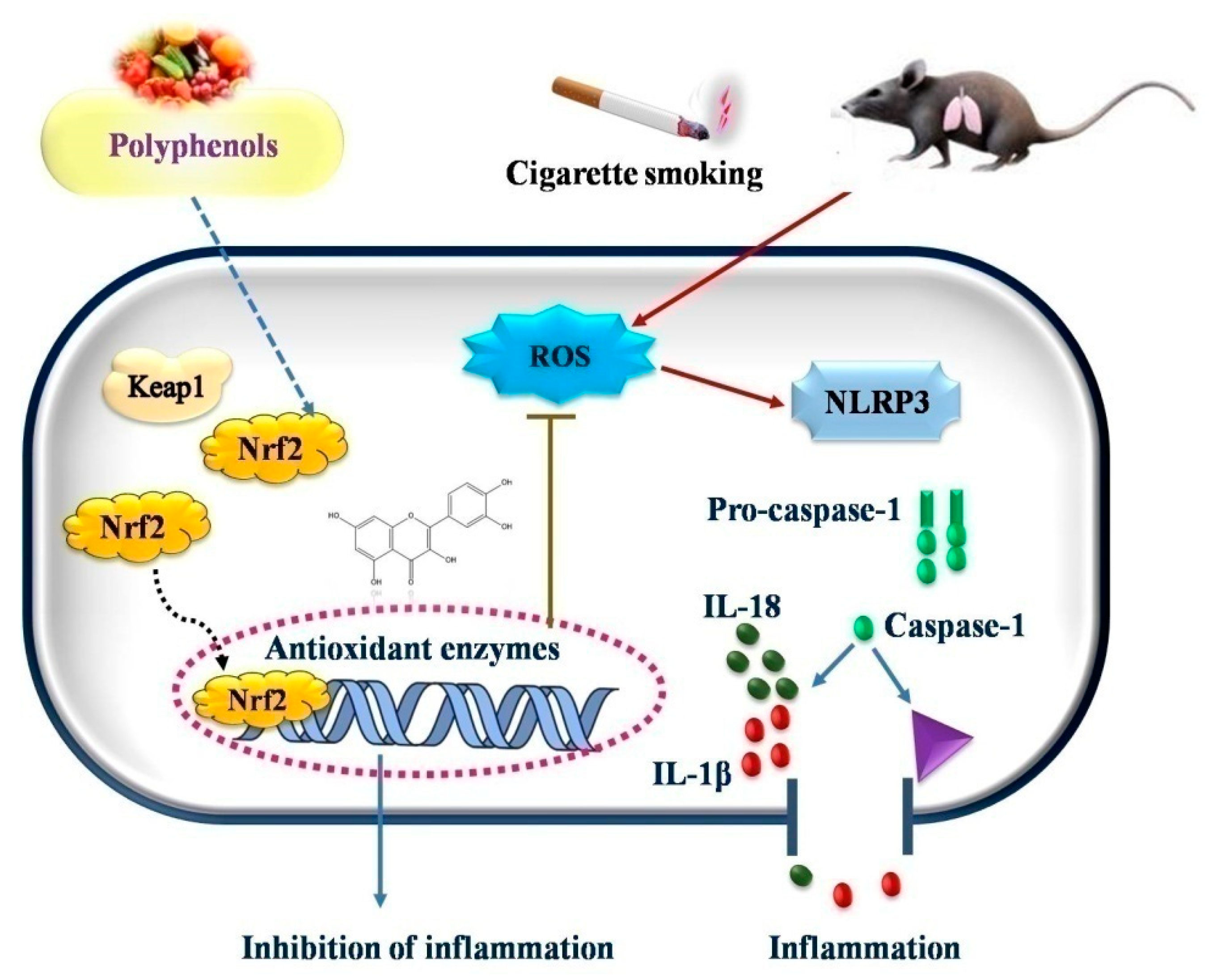
- Liu, X.; Ma, Y.; Luo, L.; Zong, D.; Li, H.; Zeng, Z.; Cui, Y.; Meng, W.; Chen, Y. Dihydroquercetin Suppresses Cigarette Smoke Induced Ferroptosis in the Pathogenesis of Chronic Obstructive Pulmonary Disease by Activating Nrf2-Mediated Pathway. Phytomedicine 2022, 96, 153894. [Google Scholar] [CrossRef]
- Tian, X.; Xue, Y.; Xie, G.; Zhou, Y.; Xiao, H.; Ding, F.; Zhang, M. (−)-Epicatechin Ameliorates Cigarette Smoke-Induced Lung Inflammation via Inhibiting ROS/NLRP3 Inflammasome Pathway in Rats with COPD. Toxicol. Appl. Pharmacol. 2021, 429, 115674. [Google Scholar] [CrossRef]
- Lakshmi, S.P.; Reddy, A.T.; Kodidhela, L.D.; Varadacharyulu, N.C. Epigallocatechin Gallate Diminishes Cigarette Smoke-Induced Oxidative Stress, Lipid Peroxidation, and Inflammation in Human Bronchial Epithelial Cells. Life Sci. 2020, 259, 118260. [Google Scholar] [CrossRef] [PubMed]
- Bao, M.-J.; Shen, J.; Jia, Y.-L.; Li, F.-F.; Ma, W.-J.; Shen, H.-J.; Shen, L.-L.; Lin, X.-X.; Zhang, L.-H.; Dong, X.-W. Apple Polyphenol Protects against Cigarette Smoke-Induced Acute Lung Injury. Nutrition 2013, 29, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhou, X.; Zhong, Y.; Ji, L.; Yu, W.; Fang, J.; Ying, H.; Li, C. Naringin Suppressed Airway Inflammation and Ameliorated Pulmonary Endothelial Hyperpermeability by Upregulating Aquaporin1 in Lipopolysaccharide/Cigarette Smoke-Induced Mice. Biomed. Pharmacother. 2022, 150, 113035. [Google Scholar] [CrossRef] [PubMed]
- Cutruzzolà, A.; Parise, M.; Vallelunga, R.; Lamanna, F.; Gnasso, A.; Irace, C. Effect of extra virgin olive oil and butter on endothelial function in type 1 diabetes. Nutrients 2021, 13, 2436. [Google Scholar] [CrossRef]
- dos Santos Jaques, J.A.; Doleski, P.H.; Castilhos, L.G.; da Rosa, M.M.; Souza, V.; Carvalho, F.B.; Marisco, P.; Thorstenberg, M.L.P.; Rezer, J.F.P.; Ruchel, J.B. Free and Nanoencapsulated Curcumin Prevents Cigarette Smoke-Induced Cognitive Impairment and Redox Imbalance. Neurobiol. Learn. Mem. 2013, 100, 98–107. [Google Scholar] [CrossRef]
- Scapagnini, G.; Colombrita, C.; Amadio, M.; D’Agata, V.; Arcelli, E.; Sapienza, M.; Quattrone, A.; Calabrese, V. Curcumin Activates Defensive Genes and Protects Neurons against Oxidative Stress. Antioxid. Redox Signal. 2006, 8, 395–403. [Google Scholar] [CrossRef]
- Prajapati, S.K.; Mishra, G.; Malaiya, A.; Jain, A.; Mody, N.; Raichur, A.M. Antimicrobial Application Potential of Phytoconstituents from Turmeric and Garlic. In Bioactive Natural Products for Pharmaceutical Applications; Springer: Berlin/Heidelberg, Germany, 2021; pp. 409–435. [Google Scholar]
- dos Santos Jaques, J.A.; Rezer, J.F.P.; Ruchel, J.B.; Becker, L.V.; da Rosa, C.S.; do Souza, V.C.G.; da Luz, S.C.A.; Gutierres, J.M.; Gonçalves, J.F.; Morsch, V.M. Lung and Blood Lymphocytes NTPDase and Acetylcholinesterase Activity in Cigarette Smoke-Exposed Rats Treated with Curcumin. Biomed. Prev. Nutr. 2011, 1, 109–115. [Google Scholar] [CrossRef]
- Qamar, W.; Sultana, S. Polyphenols from Juglans Regia L. (Walnut) Kernel Modulate Cigarette Smoke Extract Induced Acute Inflammation, Oxidative Stress and Lung Injury in Wistar Rats. Hum. Exp. Toxicol. 2011, 30, 499–506. [Google Scholar] [CrossRef]
- Rabbani, S.I.; Sajid, S.; Mani, V.; Afroz, S.M.; Khan, O.; Asdaq, S.M.B.; Yasmin, F.; Alamri, A.S.; Alsanie, W.F.; Alhomrani, M. Salvadora Persica Protects Libido by Reducing Corticosterone and Elevating the Testosterone Levels in Chronic Cigarette Smoke Exposure Rats. Saudi J. Biol. Sci. 2021, 28, 4931–4937. [Google Scholar] [CrossRef]
- Zong, D.D.; Liu, X.M.; Li, J.H.; Ouyang, R.Y.; Long, Y.J.; Chen, P.; Chen, Y. Resveratrol attenuates cigarette smoke induced endothelial apoptosis by activating Notch1 signaling mediated autophagy. Resp. Res. 2021, 22, 22. [Google Scholar] [CrossRef]
- Cheng, Y.-C.; Sheen, J.-M.; Hu, W.L.; Hung, Y.-C. Polyphenols and Oxidative Stress in Atherosclerosis-Related Ischemic Heart Disease and Stroke. Oxid. Med. Cell. Longev. 2017, 2017, 8526438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Förstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef] [PubMed]
- Pechánová, O.; Bernátová, I.; Babál, P.; Martínez, M.C.; Kyselá, S.; Stvrtina, S.; Andriantsitohaina, R. Red Wine Polyphenols Prevent Cardiovascular Alterations in L-NAME-Induced Hypertension. J. Hypertens. 2004, 22, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Rasouli, H.; Farzaei, M.H.; Khodarahmi, R. Polyphenols and Their Benefits: A Review. Int. J. Food Prop. 2017, 20, 1700–1741. [Google Scholar] [CrossRef] [Green Version]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and Computational Approaches to Estimate Solubility and Permeability in Drug Discovery and Development Settings. Adv. Drug Deliv. Rev. 1997, 23, 3, Erratum in Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Yang, C.S.; Sang, S.; Lambert, J.D.; Lee, M. Bioavailability Issues in Studying the Health Effects of Plant Polyphenolic Compounds. Mol. Nutr. Food Res. 2008, 52, S139–S151. [Google Scholar] [CrossRef]
- Yin, Z.; Zheng, T.; Ho, C.-T.; Huang, Q.; Wu, Q.; Zhang, M. Improving the Stability and Bioavailability of Tea Polyphenols by Encapsulations: A Review. Food Sci. Hum. Wellness 2022, 11, 537–556. [Google Scholar] [CrossRef]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of Intestinal Microbiota in the Bioavailability and Physiological Functions of Dietary Polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef] [Green Version]
- Pandareesh, M.D.; Mythri, R.B.; Bharath, M.M.S. Bioavailability of Dietary Polyphenols: Factors Contributing to Their Clinical Application in CNS Diseases. Neurochem. Int. 2015, 89, 198–208. [Google Scholar] [CrossRef]
- Hu, M. Commentary: Bioavailability of Flavonoids and Polyphenols: Call to Arms. Mol. Pharm. 2007, 4, 803–806. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Peng, S.; Li, Z.; Zou, L.; Liu, W.; Liu, C. Improved Bioavailability of Curcumin in Liposomes Prepared Using a PH-Driven, Organic Solvent-Free, Easily Scalable Process. RSC Adv. 2017, 7, 25978–25986. [Google Scholar] [CrossRef] [Green Version]
- Azzi, J.; Jraij, A.; Auezova, L.; Fourmentin, S.; Greige-Gerges, H. Novel Findings for Quercetin Encapsulation and Preservation with Cyclodextrins, Liposomes, and Drug-in-Cyclodextrin-in-Liposomes. Food Hydrocoll. 2018, 81, 328–340. [Google Scholar] [CrossRef]
- Chen, W.-T.; Wu, H.-T.; Chang, I.-C.; Chen, H.-W.; Fang, W.-P. Preparation of Curcumin-Loaded Liposome with High Bioavailability by a Novel Method of High Pressure Processing. Chem. Phys. Lipids 2022, 244, 105191. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, J.; Rao, Z.; Hu, J.; Wang, Q.; Sun, Y.; Lei, X.; Zhao, J.; Zeng, K.; Xu, Z. Study on the Stability and Oral Bioavailability of Curcumin Loaded (-)-Epigallocatechin-3-Gallate/Poly (N-Vinylpyrrolidone) Nanoparticles Based on Hydrogen Bonding-Driven Self-Assembly. Food Chem. 2022, 378, 132091. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Li, Z.; Yao, M.; McClements, D.J.; Xiao, H. Impact of Excipient Emulsions Made from Different Types of Oils on the Bioavailability and Metabolism of Curcumin in Gastrointestinal Tract. Food Chem. 2022, 370, 130980. [Google Scholar] [CrossRef]
- Ipar, V.S.; Singhal, R.S.; Devarajan, P. V An Innovative Approach Using Microencapsulated Turmeric Oleoresin to Develop Ready-to-Use Turmeric Milk Powder with Enhanced Oral Bioavailability. Food Chem. 2022, 373, 131400. [Google Scholar] [CrossRef] [PubMed]
- Brotons-Canto, A.; González-Navarro, C.J.; Gil, A.G.; Asin-Prieto, E.; Saiz, M.J.; Llabrés, J.M. Zein Nanoparticles Improve the Oral Bioavailability of Curcumin in Wistar Rats. Pharmaceutics 2021, 13, 361. [Google Scholar] [CrossRef]
- Panda, S.K.; Nirvanashetty, S.; Missamma, M.; Jackson-Michel, S. The Enhanced Bioavailability of Free Curcumin and Bioactive-Metabolite Tetrahydrocurcumin from a Dispersible, Oleoresin-Based Turmeric Formulation. Medicine 2021, 100, e26601. [Google Scholar] [CrossRef]
- Ng, S.W.; Selvarajah, G.T.; Hussein, M.Z.; Yeap, S.K.; Omar, A.R. In vitro Evaluation of Curcumin-Encapsulated Chitosan Nanoparticles against Feline Infectious Peritonitis Virus and Pharmacokinetics Study in Cats. Biomed Res. Int. 2020, 2020, 3012198. [Google Scholar] [CrossRef]
- Vijayan, U.K.; Varakumar, S.; Sole, S.; Singhal, R.S. Enhancement of Loading and Oral Bioavailability of Curcumin Loaded Self-Microemulsifying Lipid Carriers Using Curcuma Oleoresins. Drug Dev. Ind. Pharm. 2020, 46, 889–898. [Google Scholar] [CrossRef]
- Mahadev, M.; Nandini, H.S.; Ramu, R.; Gowda, D.V.; Almarhoon, Z.M.; Al-Ghorbani, M.; Mabkhot, Y.N. Fabrication and Evaluation of Quercetin Nanoemulsion: A Delivery System with Improved Bioavailability and Therapeutic Efficacy in Diabetes Mellitus. Pharmaceuticals 2022, 15, 70. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Chen, Z.; Zhang, Y.; Liu, E.; Han, S.; Gong, Z.; Xiao, W. Epigallocatechin-3-Gallate+ L-Theanine/β-Cyclodextrin Inclusion Complexes Enhance Epigallocatechin-3-Gallate Bioavailability and Its Lipid-Lowering and Weight Loss Effects. J. Funct. Foods 2022, 90, 104998. [Google Scholar] [CrossRef]
- Mazyed, E.A.; Helal, D.A.; Elkhoudary, M.M.; Abd Elhameed, A.G.; Yasser, M. Formulation and Optimization of Nanospanlastics for Improving the Bioavailability of Green Tea Epigallocatechin Gallate. Pharmaceuticals 2021, 14, 68. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, N.; Mandal, A.K.A. Pharmacokinetic, Toxicokinetic, and Bioavailability Studies of Epigallocatechin-3-Gallate Loaded Solid Lipid Nanoparticle in Rat Model. Drug Dev. Ind. Pharm. 2019, 45, 1506–1514. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhang, J. Enhanced Oral Bioavailability of EGCG Using PH-Sensitive Polymeric Nanoparticles: Characterization and in vivo Investigation on Nephrotic Syndrome Rats. Drug Des. Devel. Ther. 2018, 12, 2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Meng, Q.; Zhou, J.; Chen, B.; Xi, J.; Long, P.; Zhang, L.; Hou, R. Nanoemulsion Delivery System of Tea Polyphenols Enhanced the Bioavailability of Catechins in Rats. Food Chem. 2018, 242, 527–532. [Google Scholar] [CrossRef]
- Ezzat, H.M.; Elnaggar, Y.S.R.; Abdallah, O.Y. Improved Oral Bioavailability of the Anticancer Drug Catechin Using Chitosomes: Design, in-Vitro Appraisal and in-Vivo Studies. Int. J. Pharm. 2019, 565, 488–498. [Google Scholar] [CrossRef]
- Pardeshi, C.V.; Belgamwar, V.S. N, N, N-trimethyl Chitosan Modified Flaxseed Oil Based Mucoadhesive Neuronanoemulsions for Direct Nose to Brain Drug Delivery. Int. J. Biol. Macromol. 2018, 120, 2560–2571. [Google Scholar] [CrossRef]
- Patil, P.; Killedar, S. Improving Gallic Acid and Quercetin Bioavailability by Polymeric Nanoparticle Formulation. Drug Dev. Ind. Pharm. 2022, 1–8. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class of Polyphenols | Subclass | Prototype Compounds | Major Dietary Sources |
|---|---|---|---|
| Flavonoids | Flavone | Baicalein, apigenin, luteolin, chrysin | Herbal tea, fenugreek, onion, garlic, black pepper, citrus fruits, green leafy vegetables |
| Isoflavone | Genistein, daidzein, biochanin A, glycitein | Chickpea, peanut, dairy products, eggs, meat, seafood, soy products, legumes | |
| Flavonol | Rutin, quercetin, myricetin, fisetin | Tea, chocolate, cocoa, onions, scallions, kale, broccoli, apples, berries, | |
| Flavanonol | Taxifolin, aromadendrin, engeletin | Milk thistle seeds, citrus fruits | |
| Flavanol | (−)- epicatechin, (+)-catechin, (−)-epigallocatechin gallate (EGCG), theaflavins | Cocoa, chocolate, tea, grapes, apples | |
| Chalcone | Butein, xanthoangelol, 4-hydroxyderricin, cardamomin, isoliquiritigenin, isosalipurposide | Tomatoes, liquorice, shallots, bean sprouts | |
| Flavanone | Hesperidin, hesperetin, naringin, naringenin, eriodictyol | Pomegranate, citrus fruits, tomatoes, grape fruit | |
| Dihydrochalcones | Phlorizin, aspalathin, nothofagin | Apples and apple products, rooibos tea | |
| Anthocyanidins | Cyanidin, peonidin, delphinidin, petunidin, pelargonidin, malvidin | Red wines, cherries, red grapes, berries, flowers, oranges, black soybeans, hibiscus sp., purple/black rice, onions, red potatoes, purple cabbage | |
| Proanthocyanidins | Procyanidin B1, procyanidin B2, procyanidin B3 | Berries, cherries, red grapes, red wines, flowers, oranges, black soybeans, banana, cocoa, and apricot, cereals such as sorghum and barley | |
| Non-flavonoids | Phenolic acids | Caffeic acid, sinapic acid, gallic acid, protocatechuic acid, ferulic acid, p-coumaric acid | Green tea, citrus fruits, kiwi, coffee, berries, apples, rice bran, passion fruit, cherries, mangoes, wheat, corn flours |
| Stilbenes | Resveratrol | grapes (skin), mulberries, peanuts, red wine | |
| Lignans | Silymarin, sylibin, sesamin, syringaresinol, ecoisolariciresinol, matairesinol, medioresinol, pinoresinol, lariciresino | Flaxseed, soybeans, broccoli, cabbage, milk thistle, apricots, strawberries, etc. | |
| Coumarins | Dicumarol, osthole | Cinnamon, green tea, carrot, bison grass |
| Polyphenol | Bioavailability Issue | Delivery System | Subject | Result | Reference |
|---|---|---|---|---|---|
| Curcumin | Low bioavailability and degradation in solution form | Microencapsulation of curcumin in liposomes by the combination of ethanol injection and high-pressure processing | - | Effectively decreases the size of particle and PDI, which helps to cross the biological membrane. Sterilizes the bacterial, which prevent degradation in solution | [127] |
| Low bioavailability and rapid metabolism | Nanoparticle fabricated by EGCG and PVP | - | Bioavailability increased 12-fold through intestine EGCG Inhibit the metabolism of Cur, Shows high Caco-2 monolayer permeation and cellular uptake | [128] | |
| Low bioavailability and rapid metabolism | Emulsion was formed using different types of oils: corn oil, olive oil, and medium chain triglycerides (MCT) | - | Type of oil increased its transenterocyte absorption and reduced cellular metabolism | [129] | |
| Less physicochemical properties and oral bioavailability | Microencapsulating turmeric oleoresin with bioenhancers by spray drying using piperine and quercetin | - | Spray-dried powder with piperine (PIP) and quercetin (Quer) has higher permeability | [130] | |
| Low solubility and bioavailability | Zein-based nanoparticles | Wistar rats | Incresaed (9-fold) oral bioavailability with respect to the standard curcumin natural extract. | [131] | |
| Low bioavailability | Curcugen: dispersible, 98.5% turmeric-based curcuminoids formula | Randomized double-blind, 2-way cross over, single oral dose in humans | Auc-39 times and Cmax 16.1 times higher than of curcumin | [132] | |
| Low bioavailability | Curcumin-encapsulated chitosan (Cur-CS) nanoparticles | Crandell–Rees feline kidney of cat | Enhanced bioavailability, Cmax- 621.5 ng/mL three times more than normal curcumin | [133] | |
| Low bioavailability | Curcumin-loaded self-microemulsifying lipid carriers | Male Wistar rats | Higher bioavailability (29-fold) as compared to curcumin suspension | [134] | |
| Quercetin | Low bioavailability and less efficacy | Quercetin nano emulsion | Streptozocin-induced antidiabetic study in rats | Cmax of quercetin NE is 5962.74 ± 238.54 ng/mL and of quercetin pure drug is 1634.28 ± 70.18 ng/mL. AUC0-t and AUC0−∞ were 4.46 and 5.32 times higher than pure drug, respectively | [135] |
| Green tea (Epigallocatechin-3-gallate and L-theanine) | EGCG bioavailability is <5% | Preparation of EGCG + LTA/β-cyclodextrin (βCD) inclusion complexes by freeze-drying EGCG + LTA | Rats | EGCG bioavailability is improved through lipid lowerig and weight loss effects of EGCG (p < 0.05) | [136] |
| Low permeation and poor stability leads to low oral bioavailability | Nanospanlastic | Male Wistar rats | Cmax- niosomal formula (p < 0.05) and free EGCG dispersion (p < 0.001). AUC- niosomal formula (p < 0.01) and EGCG dispersion (p < 0.001) | [137] | |
| Low bioavailability and chemical instability | EGCG loaded solid lipid nanoparticles SLN | Male Wistar albino rats | Cmax of EGCG is 60.7 ± 1.07 * and EGCG loaded SLN 240 ± 16 * AUC of EGCG is 567 ± 14.5 * while EGCG loaded SL is 2329 ± 434.5 ** | [138] | |
| Poor oral bioavailability | Nanoparticles (NP) | Sprague Dawley rats | Cmax- EGCG NP 653.5 ± 181.3 * and EGCG powder 564.5 ± 121.7 * AUC 0–∞- EGCG NP5,241.6 ± 387.9 ** and EGCG powder 1321.6 ± 201.4 ** | [139] | |
| Poor bioaccesibility | Nanoemulsion | Sprague-Dawley (SD) rats | Cmax- nanoemulsion 166.7 ± 22.6 * and sol 258.8 ± 135.1 * AUC0-t- nanoemulsion 17.1 ± 0.1 ** and sol 13.3 ± 0.2 ** | [140] | |
| Green tea (Catechin) | Poor oral bioavailability | Catechin-loaded chitosan-tethered liposomes (Chitosomes) | Male Wistar rats | Cmax- Chitosomes 239.0 ± 35.27 * and sol 120.0 ± 3.97 * AUC0–24- Chitosomes 12,183 ± 1760.00 ** and sol 5739 ± 205.50 ** | [141] |
| Flaxseed | Poor efficacy | Flaxseed oil-based neuronanoemulsions (NNEs) | Balb/c mice | Plasma Cmax- NNE 24.09 ug/mL8 and pure drug suspension (PDS) 12.98 ug/ml * AUC0-12- NNE 96.38 ± 1.39 ** and PDS 18.10 ± 0.15 ** Brain Cmax- NNE 12.98 ± 0.05 * and PDS 1.67 ± 0.02 AUC0-12- NNE 107.58 ± 3.75 ** and PDS 13.18 ± 0.25 ** | [142] |
| Gallic, quercetin, amla, pomegranate | Poor bioavailability | Polyherbal nanoparticles and polyherbal extract following oral administration, pharmacokinetic parameters for polyherbal nanop | Male Wistar rats | GA and quercetin in polymeric nanoparticles improve their oral bioavailability | [143] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rudrapal, M.; Maji, S.; Prajapati, S.K.; Kesharwani, P.; Deb, P.K.; Khan, J.; Mohamed Ismail, R.; Kankate, R.S.; Sahoo, R.K.; Khairnar, S.J.; et al. Protective Effects of Diets Rich in Polyphenols in Cigarette Smoke (CS)-Induced Oxidative Damages and Associated Health Implications. Antioxidants 2022, 11, 1217. https://doi.org/10.3390/antiox11071217
Rudrapal M, Maji S, Prajapati SK, Kesharwani P, Deb PK, Khan J, Mohamed Ismail R, Kankate RS, Sahoo RK, Khairnar SJ, et al. Protective Effects of Diets Rich in Polyphenols in Cigarette Smoke (CS)-Induced Oxidative Damages and Associated Health Implications. Antioxidants. 2022; 11(7):1217. https://doi.org/10.3390/antiox11071217
Chicago/Turabian StyleRudrapal, Mithun, Siddhartha Maji, Shiv Kumar Prajapati, Payal Kesharwani, Prashanta Kumar Deb, Johra Khan, Randa Mohamed Ismail, Rani S. Kankate, Ranjan Kumar Sahoo, Shubham J. Khairnar, and et al. 2022. "Protective Effects of Diets Rich in Polyphenols in Cigarette Smoke (CS)-Induced Oxidative Damages and Associated Health Implications" Antioxidants 11, no. 7: 1217. https://doi.org/10.3390/antiox11071217