
Biochemical Discrimination of the Down Syndrome-Related Metabolic and Oxidative/Nitrosative Stress Alterations from the Physiologic Age-Related Changes through the Targeted Metabolomic Analysis of Serum

 ,
,  ,
,  , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Inclusion and Exclusion Criteria
2.3. Blood Withdrawal and Processing for Metabolite Analyses
2.4. HPLC Analysis of Purines, Pyrimidines, Antioxidants and Nitrosative Stress Biomarkers
2.5. HPLC Analysis of Amino Acids
2.6. Spectrophotometric Analysis of Serum Lactate
2.7. Statistical Analysis
3. Results
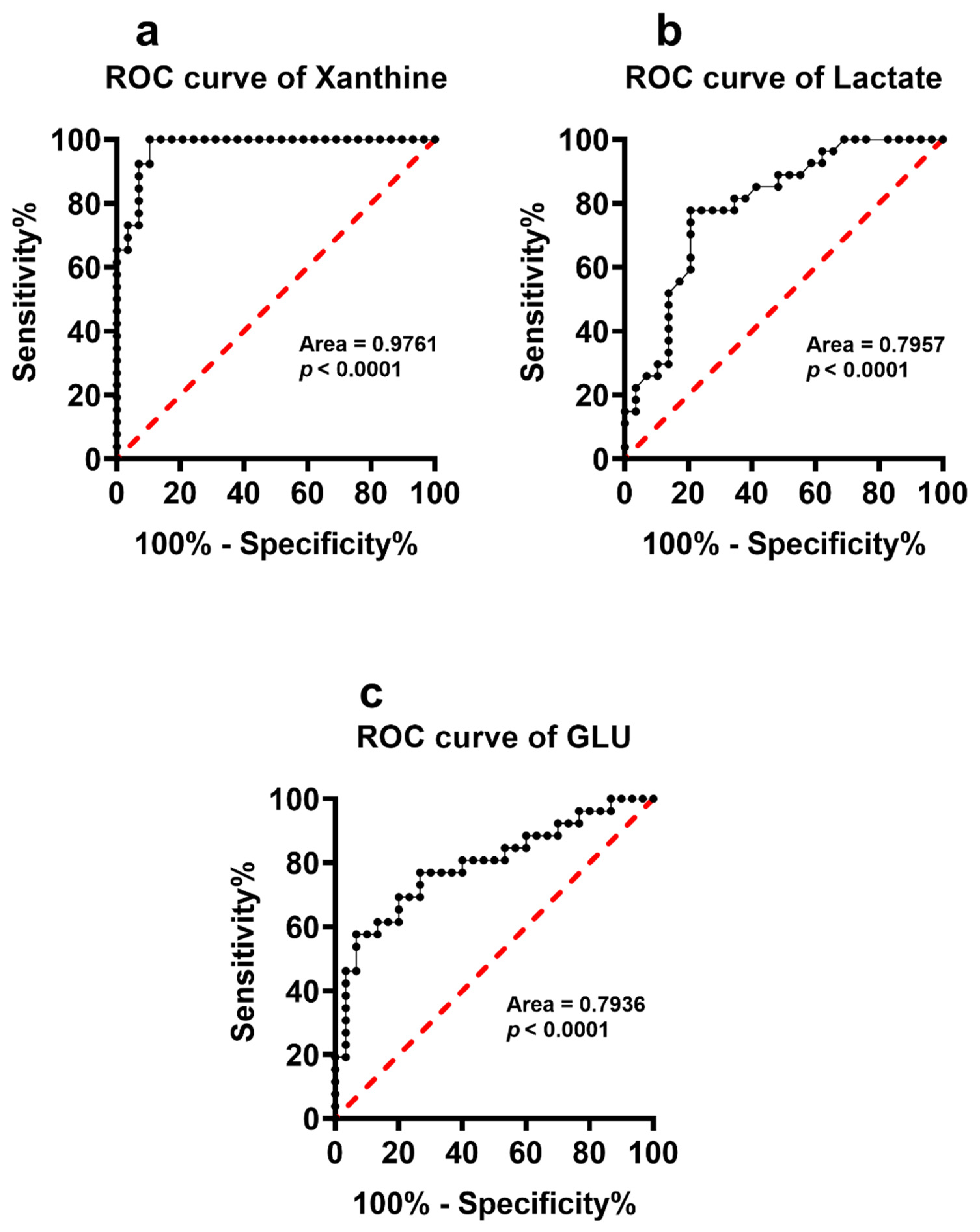
3.1. Serum Metabolites in Pooled Non-DS and Pooled DS Patients
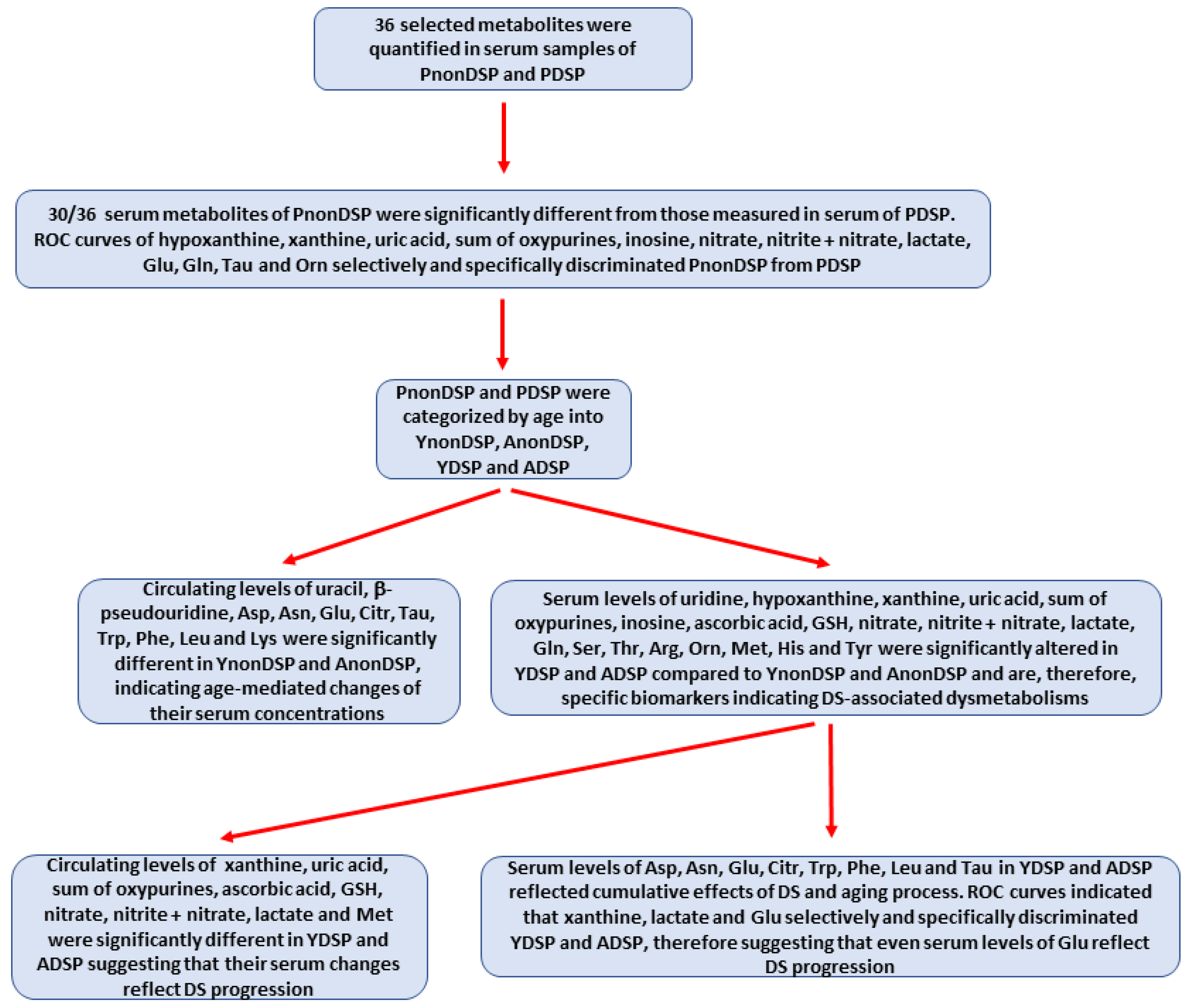
3.2. Serum Metabolites in Non-DS and DS Patients Categorized According to the Age of the Subjects
3.3. Serum Metabolites Specific of DS
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carfì, A.; Romano, A.; Zaccaria, G.; Villani, E.R.; Manes Gravina, E.; Vetrano, D.L.; Bernabei, R.; Onder, G. The burden of chronic disease, multimorbidity, and polypharmacy in adults with Down Syndrome. Am. J. Med. Genet. A 2020, 182, 1735–1743. [Google Scholar] [CrossRef] [PubMed]
- Zigman, W.B. Atypical aging in Down Syndrome. Dev. Disabil. Res. Rev. 2013, 18, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Dekker, A.D.; Ulgiati, A.M.; Groen, H.; Boxelaar, V.A.; Sacco, S.; Falquero, S.; Carfi, A.; di Paola, A.; Benejam, B.; Valldeneu, S.; et al. The Behavioral and Psychological Symptoms of Dementia in Down Syndrome Scale (BPSD-DS II): Optimization and Further Validation. J. Alzheimer’s Dis. 2021, 82, 1371. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, A.M.; Yuan, P.; Rivera, B.M.; Schaaf, W.; Mladinov, M.; Prasher, V.P.; Robinson, A.C.; DeGrado, W.F.; Condello, C. Emergence of distinct and heterogeneous strains of amyloid beta with advanced Alzheimer’s disease pathology in Down Syndrome. Acta Neuropathol. Commun. 2021, 9, 201. [Google Scholar] [CrossRef]
- Cenini, G.; Fiorini, A.; Sultana, R.; Perluigi, M.; Cai, J.; Klein, J.B.; Head, E.; Butterfield, D.A. An investigation of the molecular mechanisms engaged before and after the development of Alzheimer disease neuropathology in Down Syndrome: A proteomics approach. Free Radic. Biol. Med. 2014, 76, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Song, B.; Li, G.; Cai, F.; Wu, M.; Zhao, Y.; Jiang, L.; Guo, T.; Shen, M.; Hou, H.; et al. USP25 inhibition ameliorates Alzheimer’s pathology through the regulation of APP processing and Abeta generation. J. Clin. Investig. 2022, 132, e152170. [Google Scholar] [CrossRef]
- Wiseman, F.K.; Al-Janabi, T.; Hardy, J.; Karmiloff-Smith, A.; Nizetic, D.; Tybulewicz, V.L.; Fisher, E.M.; Strydom, A. A genetic cause of Alzheimer disease: Mechanistic insights from Down Syndrome. Nat. Rev. Neurosci. 2015, 16, 564–574. [Google Scholar] [CrossRef] [Green Version]
- Lejeune, J. Pathogenesis of mental deficiency in trisomy 21. Am. J. Med. Genet. Suppl. 1990, 7, 20–30. [Google Scholar] [CrossRef]
- Pagano, G.; Castello, G. Oxidative stress and mitochondrial dysfunction in Down Syndrome. Adv. Exp. Med. Biol. 2012, 724, 291–299. [Google Scholar]
- Ferrari, M.; Stagi, S. Oxidative Stress in Down and Williams-Beuren Syndromes: An Overview. Molecules 2021, 26, 3139. [Google Scholar] [CrossRef]
- Pogribna, M.; Melnyk, S.; Pogribny, I.; Chang, A.P.; James, S.J. Homocysteine metabolism in children with Down Syndrome: In vitro modulation. Am. J. Hum. Genet. 2001, 69, 88–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Infantino, V.; Castegna, A.; Iacobazzi, F.; Spera, I.; Scala, I.; Andria, G.; Iacobazzi, V. Impairment of methyl cycle affects mitochondrial methyl availability and glutathione level in Down’s syndrome. Mol. Genet. Metab. 2011, 102, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Garlet, T.R.; Parisotto, E.B.; de Medeiros Gda, S.; Pereira, L.C.; Moreira, E.A.; Dalmarco, E.M.; Dalmarco, J.B.; Filho, D.W. Systemic oxidative stress in children and teenagers with Down Syndrome. Life Sci. 2013, 93, 558–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montoliu-Gaya, L.; Strydom, A.; Blennow, K.; Zetterberg, H.; Ashton, N.J. Blood Biomarkers for Alzheimer’s Disease in Down Syndrome. J. Clin. Med. 2021, 10, 3639. [Google Scholar] [CrossRef] [PubMed]
- Obeid, R.; Hartmuth, K.; Herrmann, W.; Gortner, L.; Rohrer, T.R.; Geisel, J.; Reed, M.C.; Nijhout, H.F. Blood biomarkers of methylation in Down Syndrome and metabolic simulations using a mathematical model. Mol. Nutr. Food Res. 2012, 56, 1582–1589. [Google Scholar] [CrossRef]
- Antonaros, F.; Lanfranchi, S.; Locatelli, C.; Martelli, A.; Olivucci, G.; Cicchini, E.; Carosi Diatricch, L.; Mannini, E.; Vione, B.; Feliciello, A.; et al. One-carbon pathway and cognitive skills in children with Down Syndrome. Sci. Rep. 2021, 11, 4225. [Google Scholar] [CrossRef]
- Culp-Hill, R.; Zheng, C.; Reisz, J.A.; Smith, K.; Rachubinski, A.; Nemkov, T.; Butcher, E.; Granrath, R.; Hansen, K.C.; Espinosa, J.M.; et al. Red blood cell metabolism in Down Syndrome: Hints on metabolic derangements in aging. Blood Adv. 2017, 1, 2776–2780. [Google Scholar] [CrossRef] [Green Version]
- Barone, E.; Arena, A.; Head, E.; Butterfield, D.A.; Perluigi, M. Disturbance of redox homeostasis in Down Syndrome: Role of iron dysmetabolism. Free Radic. Biol. Med. 2018, 114, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Caracausi, M.; Ghini, V.; Locatelli, C.; Mericio, M.; Piovesan, A.; Antonaros, F.; Pelleri, M.C.; Vitale, L.; Vacca, R.A.; Bedetti, F.; et al. Plasma and urinary metabolomic profiles of Down Syndrome correlate with alteration of mitochondrial metabolism. Sci. Rep. 2018, 8, 2977. [Google Scholar] [CrossRef] [Green Version]
- Campos, C.; Guzmán, R.; López-Fernández, E.; Casado, A. Evaluation of urinary biomarkers of oxidative/nitrosative stress in children with Down Syndrome. Life Sci. 2011, 89, 655–661. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, J.; The, J.P.Y.; Cheon, B.K.; Yang, Y.; Schlundt, J.; Wang, Y.; Conway, P.L. Comparative Blood and Urine Metabolomics Analysis of Healthy Elderly and Young Male Singaporeans. J. Proteome Res. 2020, 19, 3264–3275. [Google Scholar] [CrossRef] [PubMed]
- Lazzarino, G.; Amorini, A.M.; Fazzina, G.; Vagnozzi, R.; Signoretti, S.; Donzelli, S.; Di Stasio, E.; Giardina, B.; Tavazzi, B. Single-sample preparation for simultaneous cellular redox and energy state determination. Anal. Biochem. 2003, 322, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Tavazzi, B.; Lazzarino, G.; Leone, P.; Amorini, A.M.; Bellia, F.; Janson, C.G.; Di Pietro, V.; Ceccarelli, L.; Donzelli, S.; Francis, J.S.; et al. Simultaneous high performance liquid chromatographic separation of purines, pyrimidines, N-acetylated amino acids, and dicarboxylic acids for the chemical diagnosis of inborn errors of metabolism. Clin. Biochem. 2005, 38, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Bracko, O.; Di Pietro, V.; Lazzarino, G.; Amorini, A.M.; Tavazzi, B.; Artmann, J.; Wong, E.C.; Buxton, R.B.; Weller, M.; Luft, A.R.; et al. 3-Nitropropionic acid-induced ischemia tolerance in the rat brain is mediated by reduced metabolic activity and cerebral blood flow. J. Cereb. Blood Flow Metab. 2014, 34, 1522–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazzarino, G.; Listorti, I.; Bilotta, G.; Capozzolo, T.; Amorini, A.M.; Longo, S.; Caruso, G.; Lazzarino, G.; Tavazzi, B.; Bilotta, P. Water- and Fat-Soluble Antioxidants in Human Seminal Plasma and Serum of Fertile Males. Antioxidants 2019, 8, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amorini, A.M.; Lazzarino, G.; Di Pietro, V.; Signoretti, S.; Lazzarino, G.; Belli, A.; Tavazzi, B. Severity of experimental traumatic brain injury modulates changes in concentrations of cerebral free amino acids. J. Cell. Mol. Med. 2017, 21, 530–542. [Google Scholar] [CrossRef]
- Artiss, J.D.; Karcher, R.E.; Cavanagh, K.T.; Collins, S.L.; Peterson, V.J.; Varma, S.; Zak, B. A liquid-stable reagent for lactic acid levels. Application to the Hitachi 911 and Beckman CX7. Am. J. Clin. Pathol. 2000, 114, 139–143. [Google Scholar] [CrossRef] [Green Version]
- Pecze, L.; Randi, E.B.; Szabo, C. Meta-analysis of metabolites involved in bioenergetic pathways reveals a pseudohypoxic state in Down Syndrome. Mol. Med. 2020, 26, 102. [Google Scholar] [CrossRef]
- Fuller, R.W.; Luce, M.W.; Mertz, E.T. Serum uric acid in mongolism. Science 1962, 137, 868–869. [Google Scholar] [CrossRef]
- Mertz, E.T.; Fuller, R.W.; Fuller, R.W.; Concon, J.M. Serum uric acid in young mongoloids. Science 1963, 141, 535. [Google Scholar] [CrossRef]
- Chapman, M.; Stern, J. Uric acid in Downs disease. J. Intellect. Disabil. Res. 1964, 8, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Rosner, F.; Ong, B.H.; Paine, R.S.; Mahanand, D. Biochemical differentiation of trisomic Down’s syndrome (mongolism) from that due to translocation. N. Engl. J. Med. 1965, 273, 1356–1361. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, J.M.; O’Brien, W.M. Hyperuricemia in mongolism. N. Engl. J. Med. 1967, 276, 953–956. [Google Scholar] [CrossRef] [PubMed]
- Howell, A.; Mason, A.S.; Brown, E.; Watts, R.W.; Chanarin, I.; McPherson, K.; Ridler, M.A. Red cell size and uric acid in Down’s syndrome. Scand. J. Haematol. 1973, 11, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Pant, S.S.; Moser, H.W.; Krane, S.M. Hyperuricemia in Down’s syndrome. J. Clin. Endocrinol. Metab. 1968, 28, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Zitnanová, I.; Korytár, P.; Aruoma, O.I.; Sustrová, M.; Garaiová, I.; Muchová, J.; Kalnovicová, T.; Pueschel, S.; Duracková, Z. Uric acid and allantoin levels in Down Syndrome: Antioxidant and oxidative stress mechanisms? Clin. Chim. Acta 2004, 341, 139–146. [Google Scholar] [CrossRef]
- Tiano, L.; Padella, L.; Carnevali, P.; Gabrielli, O.; Bruge, F.; Principi, F.; Littarru, G.P. Coenzyme Q10 and oxidative imbalance in Down Syndrome: Biochemical and clinical aspects. Biofactors 2008, 32, 161–167. [Google Scholar] [CrossRef]
- Powers, R.K.; Culp-Hill, R.; Ludwig, M.P.; Smith, K.P.; Waugh, K.A.; Minter, R.; Tuttle, K.D.; Lewis, H.C.; Rachubinski, A.L.; Granrath, R.E.; et al. Trisomy 21 activates the kynurenine pathway via increased dosage of interferon receptors. Nat. Commun. 2019, 101, 4766. [Google Scholar] [CrossRef] [Green Version]
- Orozco, J.S.; Hertz-Picciotto, I.; Abbeduto, L.; Slupsky, C.M. Metabolomics analysis of children with autism, idiopathic-developmental delays, and Down Syndrome. Transl. Psychiatry 2019, 9, 243. [Google Scholar] [CrossRef] [Green Version]
- Antonaros, F.; Ghini, V.; Pulina, F.; Ramacieri, G.; Cicchini, E.; Mannini, E.; Martelli, A.; Feliciello, A.; Lanfranchi, S.; Onnivello, S.; et al. Plasma metabolome and cognitive skills in Down Syndrome. Sci. Rep. 2020, 10, 10491. [Google Scholar] [CrossRef]
- Schulz, E.; Wenzel, P.; Münzel, T.; Daiber, A. Mitochondrial redox signaling: Interaction of mitochondrial reactive oxygen species with other sources of oxidative stress. Antioxid. Redox Signal. 2014, 20, 308–324. [Google Scholar] [CrossRef] [PubMed]
- Lazzarino, G.; Amorini, A.M.; Petzold, A.; Gasperini, C.; Ruggieri, S.; Quartuccio, M.E.; Lazzarino, G.; Di Stasio, E.; Tavazzi, B. Serum Compounds of Energy Metabolism Impairment Are Related to Disability, Disease Course and Neuroimaging in Multiple Sclerosis. Mol. Neurobiol. 2017, 54, 7520–7533. [Google Scholar] [CrossRef] [PubMed]
- Sadeghian, M.; Mastrolia, V.; Rezaei Haddad, A.; Mosley, A.; Mullali, G.; Schiza, D.; Sajic, M.; Hargreaves, I.; Heales, S.; Duchen, M.R.; et al. Mitochondrial dysfunction is an important cause of neurological deficits in an inflammatory model of multiple sclerosis. Sci. Rep. 2016, 6, 33249. [Google Scholar] [CrossRef]
- Carpenter, K.L.; Jalloh, I.; Hutchinson, P.J. Glycolysis and the significance of lactate in traumatic brain injury. Front. Neurosci. 2015, 9, 112. [Google Scholar] [CrossRef] [PubMed]
- Esmael, A.; Talaat, M.; Egila, H.; Eltoukhy, K. Mitochondrial dysfunction and serum lactate as a biomarker for the progression and disability in MS and its correlation with the radiological findings. Neurol. Res. 2021, 43, 582–590. [Google Scholar] [CrossRef]
- Amorini, A.M.; Nociti, V.; Petzold, A.; Gasperini, C.; Quartuccio, E.; Lazzarino, G.; Di Pietro, V.; Belli, A.; Signoretti, S.; Vagnozzi, R.; et al. Serum lactate as a novel potential biomarker in multiple sclerosis. Biochim. Biophys. Acta 2014, 1842, 1137–1143. [Google Scholar] [CrossRef] [Green Version]
- Alldred, M.J.; Martini, A.C.; Patterson, D.; Hendrix, J.; Granholm, A.C. Aging with Down Syndrome-Where Are We Now and Where Are We Going? J. Clin. Med. 2021, 10, 4687. [Google Scholar] [CrossRef]
- Martinez, J.L.; Zammit, M.D.; West, N.R.; Christian, B.T.; Bhattacharyya, A. Basal Forebrain Cholinergic Neurons: Linking Down Syndrome and Alzheimer’s Disease. Front. Aging Neurosci. 2021, 13, 703876. [Google Scholar] [CrossRef]
- Talebi, M.; Majdi, A.; Nasiri, E.; Naseri, A.; Sadigh-Eteghad, S. The correlation between circulating inflammatory, oxidative stress, and neurotrophic factors level with the cognitive outcomes in multiple sclerosis patients. Neurol. Sci. 2021, 42, 2291–2300. [Google Scholar] [CrossRef]
- Li, L.L.; Ma, Y.H.; Bi, Y.L.; Sun, F.R.; Hu, H.; Hou, X.H.; Xu, W.; Shen, X.N.; Dong, Q.; Tan, L.; et al. Serum Uric Acid May Aggravate Alzheimer’s Disease Risk by Affecting Amyloidosis in Cognitively Intact Older Adults: The CABLE Study. J. Alzheimer’s Dis. 2021, 81, 389–401. [Google Scholar] [CrossRef]
- González-Domínguez, R.; García-Barrera, T.; Gómez-Ariza, J.L. Metabolite profiling for the identification of altered metabolic pathways in Alzheimer’s disease. J. Pharm. Biomed. Anal. 2015, 107, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Socha, K.; Klimiuk, K.; Naliwajko, S.K.; Soroczyńska, J.; Puścion-Jakubik, A.; Markiewicz-Żukowska, R.; Kochanowicz, J. Dietary Habits, Selenium, Copper, Zinc and Total Antioxidant Status in Serum in Relation to Cognitive Functions of Patients with Alzheimer’s Disease. Nutrients 2021, 13, 287. [Google Scholar] [CrossRef] [PubMed]
- Dildar, K.; Sinem, F.; Gökhan, E.; Orhan, Y.; Filiz, M. Serum nitrosative stress levels are increased in Alzheimer disease but not in vascular dementia. Alzheimer’s Dis. Assoc. Disord. 2010, 24, 194–197. [Google Scholar] [CrossRef] [PubMed]
- Schirinzi, T.; Di Lazzaro, G.; Sancesario, G.M.; Summa, S.; Petrucci, S.; Colona, V.L.; Bernardini, S.; Pierantozzi, M.; Stefani, A.; Mercuri, N.B.; et al. Young-onset and late-onset Parkinson’s disease exhibit a different profile of fluid biomarkers and clinical features. Neurobiol. Aging 2020, 90, 119–124. [Google Scholar] [CrossRef]
- Babu, G.N.; Gupta, M.; Paliwal, V.K.; Singh, S.; Chatterji, T.; Roy, R. Serum metabolomics study in a group of Parkinson’s disease patients from northern India. Clin. Chim. Acta 2018, 480, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Karahalil, B.; Salihoğlu, E.M.; Elkama, A.; Orhan, G.; Saygın, E.; Akaydin, S.Y. Individual susceptibility has a major impact on strong association between oxidative stress, defence systems and Parkinson’s disease. Basic Clin. Pharmacol. Toxicol. 2022, 130, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Kouti, L.; Noroozian, M.; Akhondzadeh, S.; Abdollahi, M.; Javadi, M.R.; Faramarzi, M.A.; Mousavi, S.; Ghaeli, P. Nitric oxide and peroxynitrite serum levels in Parkinson’s disease: Correlation of oxidative stress and the severity of the disease. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 964–970. [Google Scholar]
- Sun, J.; Lv, X.; Gao, X.; Chen, Z.; Wei, D.; Ling, Y.; Zhang, J.; Gu, Q.; Liu, J.; Chen, W.; et al. The association between serum uric acid level and the risk of cognitive impairment after ischemic stroke. Neurosci. Lett. 2020, 734, 135098. [Google Scholar] [CrossRef]
- Oh, T.K.; Song, I.A.; Bae, H.J.; Jeon, Y.T. Serum lactate level upon admission to the neuro-intensive care unit and 90-day mortality: A retrospective study. J. Clin. Neurosci. 2019, 70, 173–177. [Google Scholar] [CrossRef]
- Shaafi, S.; Hadisi, F.; Mahmoudinezhad, M.; Razmi, H.; Nejadghaderi, S.A.; Khalili, M. The significance of the oxidative stress markers in the one-year prognosis of patients with acute ischemic stroke: A case-control study. BMC Neurol. 2021, 21, 258. [Google Scholar] [CrossRef]
- Ciancarelli, I.; Di Massimo, C.; De Amicis, D.; Carolei, A.; Tozzi Ciancarelli, M.G. Evidence of redox unbalance in post-acute ischemic stroke patients. Curr. Neurovasc. Res. 2012, 9, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Mandurino-Mirizzi, A.; Cornara, S.; Somaschini, A.; Demarchi, A.; Galazzi, M.; Puccio, S.; Montalto, C.; Crimi, G.; Ferlini, M.; Camporotondo, R.; et al. Elevated serum uric acid is associated with a greater inflammatory response and with short- and long-term mortality in patients with ST-segment elevation myocardial infarction undergoing primary percutaneous coronary intervention. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Lazzarino, G.; Raatikainen, P.; Nuutinen, M.; Nissinen, J.; Tavazzi, B.; Di Pierro, D.; Giardina, B.; Peuhkurinen, K. Myocardial release of malondialdehyde and purine compounds during coronary bypass surgery. Circulation 1994, 90, 291–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ham, P.B., 3rd; Raju, R. Mitochondrial function in hypoxic ischemic injury and influence of aging. Prog. Neurobiol. 2017, 157, 92–116. [Google Scholar] [CrossRef]
- Tomandlova, M.; Parenica, J.; Lokaj, P.; Ondrus, T.; Kala, P.; Miklikova, M.; Helanova, K.; Helan, M.; Malaska, J.; Benesova, K.; et al. Prognostic value of oxidative stress in patients with acute myocardial infarction complicated by cardiogenic shock: A prospective cohort study. Free Radic. Biol. Med. 2021, 17, 66–72. [Google Scholar] [CrossRef]
- Abdelzaher, W.Y.; Ahmed, S.M.; Welson, N.N.; Alsharif, K.F.; Batiha, G.E.; Labib, D.A.A. Dapsone Ameliorates Isoproterenol-Induced Myocardial Infarction via Nrf2/ HO-1; TLR4/ TNF-α Signaling Pathways and the Suppression of Oxidative Stress, Inflammation, and Apoptosis in Rats. Front. Pharmacol. 2021, 12, 669679. [Google Scholar] [CrossRef]
- Marcovecchio, G.E.; Ferruam, F.; Fontana, E.; Beretta, S.; Genua, M.; Bortolomai, I.; Conti, A.; Montin, D.; Cascarano, M.T.; Bergante, S.; et al. Premature Senescence and Increased Oxidative Stress in the Thymus of Down Syndrome Patients. Front. Immunol. 2021, 12, 669893. [Google Scholar] [CrossRef]
- Edgin, J.O.; Anand, P.; Rosser, T.; Pierpont, E.I.; Figueroa, C.; Hamilton, D.; Huddleston, L.; Mason, G.; Spanò, G.; Toole, L.; et al. The Arizona Cognitive Test Battery for Down Syndrome: Test-Retest Reliability and Practice Effects. Am. J. Intellect. Dev. Disabil. 2017, 122, 215–234. [Google Scholar] [CrossRef]
- Sullivan, K.D.; Lewis, H.C.; Hill, A.A.; Pandey, A.; Jackson, L.P.; Cabral, J.M.; Smith, K.P.; Liggett, L.A.; Gomez, E.B.; Galbraith, M.D.; et al. Trisomy 21 consistently activates the interferon response. eLife 2016, 5, e16220. [Google Scholar] [CrossRef]
- Valenti, D.; De Rasmo, D.; Signorile, A.; Rossi, L.; de Bari, L.; Scala, I.; Granese, B.; Papa, S.; Vacca, R.A. Epigallocatechin-3-gallate prevents oxidative phosphorylation deficit and promotes mitochondrial biogenesis in human cells from subjects with Down’s syndrome. Biochim. Biophys. Acta 2013, 1832, 542–552. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Sureda, V.; Vilches, Á.; Sánchez, O.; Audí, L.; Domínguez, C. Intracellular oxidant activity, antioxidant enzyme defense system, and cell senescence in fibroblasts with trisomy 21. Oxidative Med. Cell. Longev. 2015, 2015, 509241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izzo, A.; Nitti, M.; Mollo, N.; Paladino, S.; Procaccini, C.; Faicchia, D.; Calì, G.; Genesio, R.; Bonfiglio, F.; Cicatiello, R.; et al. Metformin restores the mitochondrial network and reverses mitochondrial dysfunction in Down Syndrome cells. Hum. Mol. Genet. 2017, 26, 1056–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helguera, P.; Seiglie, J.; Rodriguez, J.; Hanna, M.; Helguera, G.; Busciglio, J. Adaptive downregulation of mitochondrial function in Down Syndrome. Cell Metab. 2013, 17, 132–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, A.; Fabbrini, F.; D’Agostino, P.; Negri, R.; Greco, D.; Genesio, R.; D’Armiento, M.; Olla, C.; Paladini, D.; Zannini, M.; et al. Altered expression of mitochondrial and extracellular matrix genes in the heart of human fetuses with chromosome 21 trisomy. BMC Genom. 2007, 8, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amorini, A.M.; Giorlandino, C.; Longo, S.; D’Urso, S.; Mesoraca, A.; Santoro, M.L.; Picardi, M.; Gullotta, S.; Cignini, P.; Lazzarino, D.; et al. Metabolic profile of amniotic fluid as a biochemical tool to screen for inborn errors of metabolism and fetal anomalies. Mol. Cell. Biochem. 2012, 359, 205–216. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Concentrations in Serum of PnonDSP (n = 102) μmol/L Serum | Concentrations in Serum of PDSP (n = 56) μmol/L Serum | Significantly Different from PnonDSP q-Values | Correlation with the Age of the Subject, Values of the Spearman’s Correlation Coefficient r of PnonDSP | p-Values of r of PnonDSP | Correlation with the Age of the Subject, Values of the Spearman’s Correlation Coefficient r of PDSP | p-Values of r of PDSP |
|---|---|---|---|---|---|---|---|
| Uracil | 1.05 ± 0.76 | 0.59 ± 0.36 | <0.0001 | 0.456 | <0.0001 | 0.378 | <0.005 |
| β-pseudouridine | 2.50± 1.26 | 4.03 ± 1.54 | <0.0001 | 0.359 | <0.0002 | 0.011 | N.S. |
| Uridine | 6.39 ± 3.23 | 12.64 ± 8.20 | <0.0001 | 0.063 | N.S. | 0.104 | N.S. |
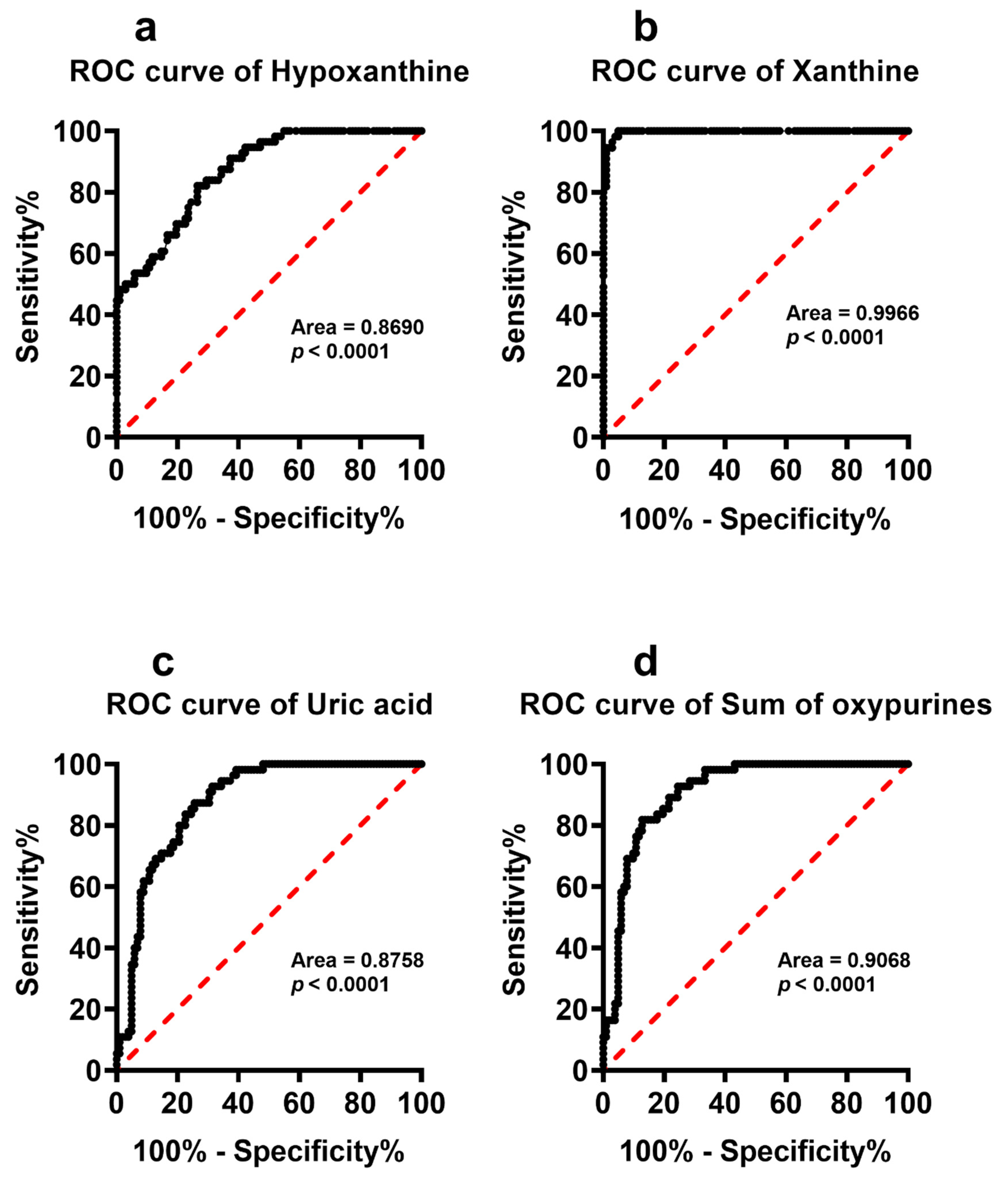
| Hypoxanthine | 5.40 ± 2.40 | 10.73 ± 4.22 | <0.0001 | 0.076 | N.S. | 0.125 | N.S. |
| Xanthine | 2.86 ± 1.89 | 19.18 ± 7.76 | <0.0001 | −0.028 | N.S. | 0.299 | <0.05 |
| Uric acid | 280.25 ± 101.71 | 410.40 ± 67.72 | <0.0001 | −0.086 | N.S. | 0.248 | N.S. |
| Sum of oxypurines | 290.96 ± 104.67 | 439.65 ± 70.63 | <0.0001 | −0.059 | N.S. | 0.243 | N.S. |
| Inosine | 0.77 ± 0.66 | 2.15 ± 1.55 | <0.0001 | −0.055 | N.S. | 0.044 | N.S. |
| Ascorbic acid | 52.50 ± 20.33 | 42.72 ± 17.02 | <0.002 | 0.010 | N.S. | −0.175 | N.S. |
| GSH | 25.10 ± 7.83 | 19.67 ± 8.75 | <0.001 | −0.146 | N.S. | −0.392 | <0.005 |
| Nitrite | 1.72 ± 0.84 | 2.38 ± 1.32 | <0.002 | 0.049 | N.S. | 0.031 | N.S. |
| Nitrate | 31.37 ± 14.09 | 83.30 ± 29.70 | <0.0001 | 0.044 | N.S. | 0.392 | <0.005 |
| Nitrite + Nitrate | 33.09 ± 14.23 | 85.68 ± 29.70 | <0.0001 | 0.037 | N.S. | 0.392 | <0.005 |
| Lactate | 1.05 ± 0.23 | 1.91 ± 0.65 | <0.0001 | 0.192 | N.S. | 0.529 | <0.0001 |
| Creatinine | 69.89 ± 26.81 | 75.91 ± 18.41 | N.S. | 0.101 | N.S. | 0.054 | N.S. |
| Compound | Concentrations in Serum of PnonDSP (n = 102) μmol/L serum | Concentrations in Serum of PDSP (n = 56) μmol/L serum | Significantly Different from PnonDSP q-values | Correlation with the Age of the Subject, Values of the Spearman’s Correlation Coefficient r of PnonDSP | p-Values of r of PnonDSP | Correlation with the Age of the Subject, Values of the Spearman’s Correlation Coefficient r of PDSP | p-Values of r of PDSP |
|---|---|---|---|---|---|---|---|
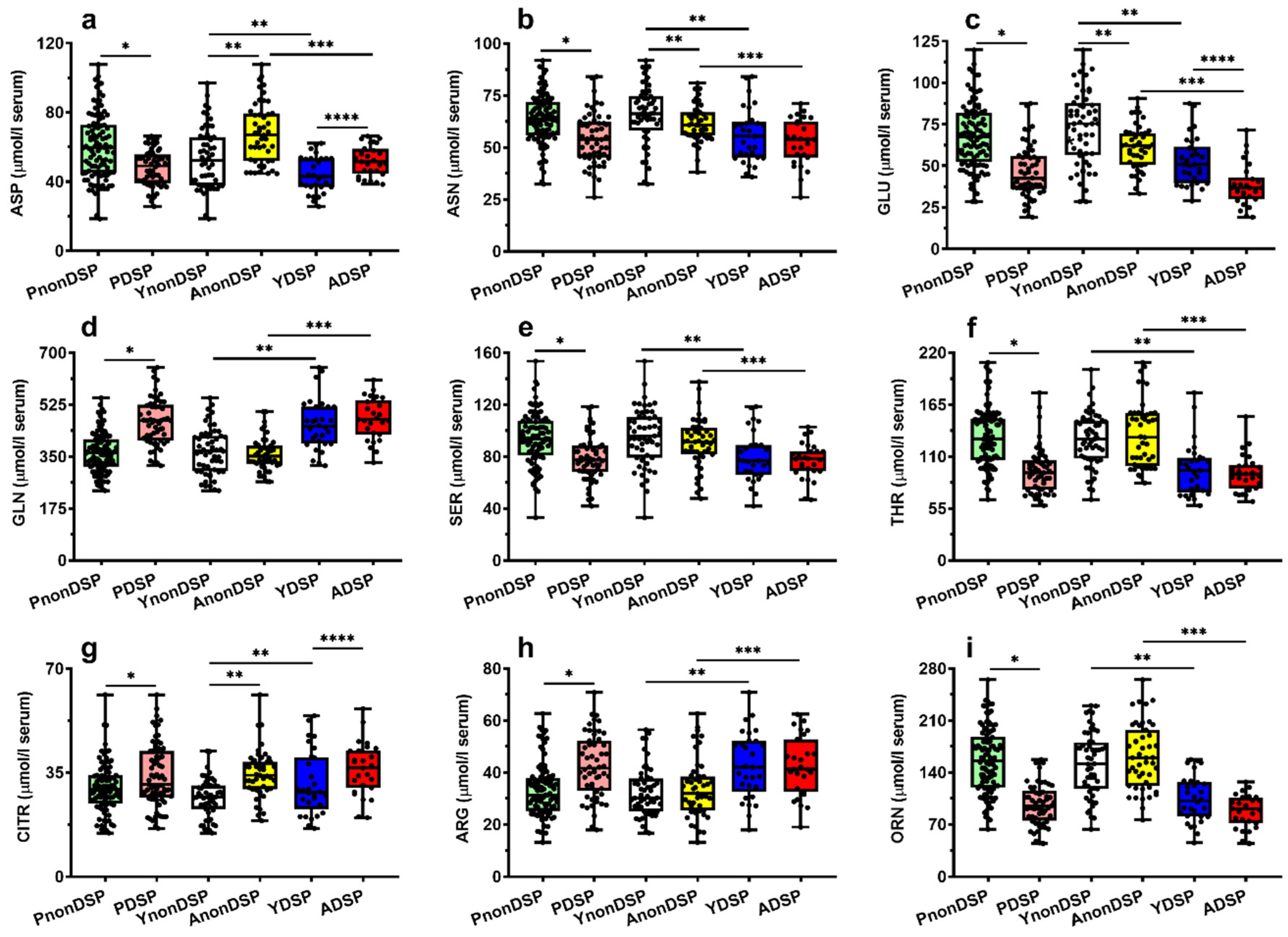
| ASP | 60.15 ± 18.88 | 47.85 ± 10.38 | <0.0001 | 0.316 | <0.002 | 0.318 | <0.02 |
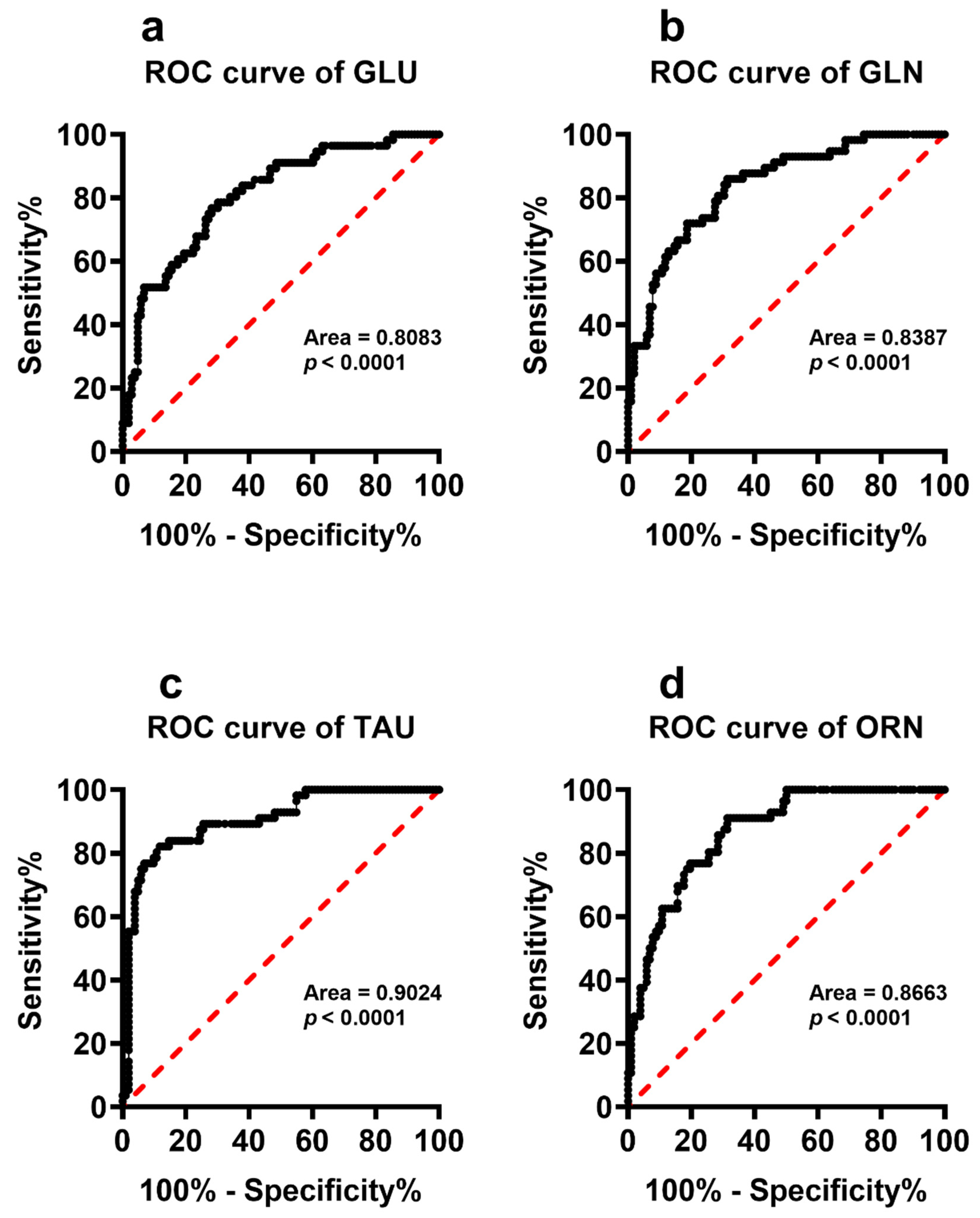
| GLU | 67.45± 19.22 | 46.08 ± 15.55 | <0.0001 | −0.275 | <0.002 | −0.447 | <0.001 |
| ASN | 63.65 ± 11.43 | 54.19 ± 12.25 | <0.0001 | −0.118 | N.S. | −0.156 | N.S. |
| SER | 93.03 ± 20.74 | 78.75 ± 17.42 | <0.0001 | −0.030 | N.S. | −0.088 | N.S. |
| GLN | 363.09 ± 68.14 | 467.40 ± 78.58 | <0.0001 | −0.203 | N.S. | 0.229 | N.S. |
| HIS | 73.68± 14.89 | 62.11 ± 14.04 | <0.0001 | −0.077 | N.S. | −0.257 | N.S. |
| GLY | 220.41 ± 79.08 | 212.07 ± 49.26 | N.S. | −0.161 | N.S. | 0.181 | N.S. |
| THR | 131.08 ± 30.53 | 95.68 ± 25.24 | <0.0001 | −0.082 | N.S. | −0.121 | N.S. |
| CITR | 29.86 ± 8.52 | 33.89 ± 10.44 | <0.02 | 0.414 | <0.0001 | 0.227 | N.S. |
| ARG | 33.11 ± 10.34 | 42.48 ± 12.01 | <0.0001 | 0.102 | N.S. | −0.076 | N.S. |
| ALA | 371.96 ± 96.49 | 382.45 ± 106.78 | N.S. | −0.136 | N.S. | 0.221 | N.S. |
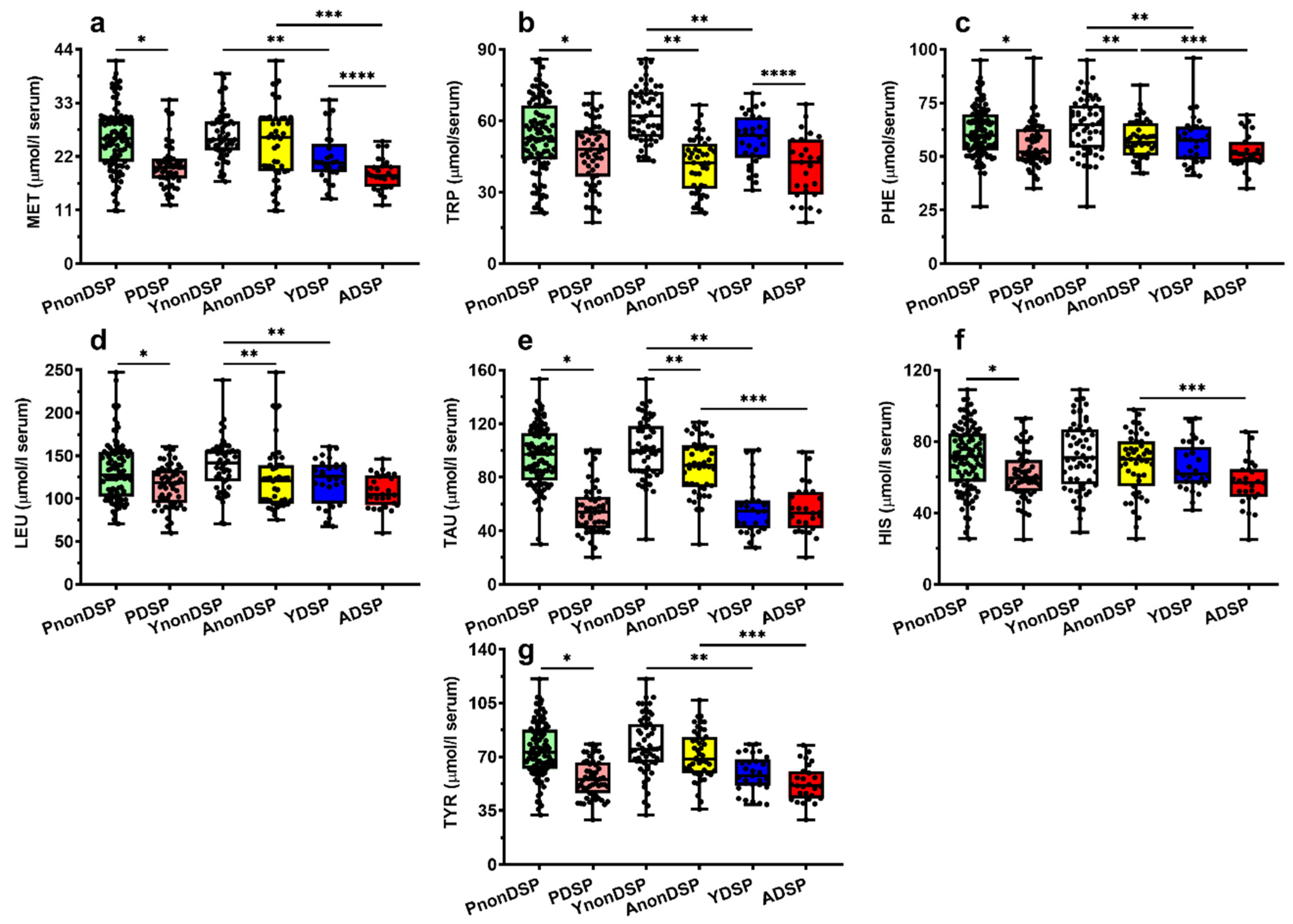
| TAU | 95.23 ± 22.49 | 56.50 ± 19.03 | <0.0001 | −0.178 | N.S. | −0.033 | N.S. |
| TYR | 74.30± 17.82 | 56.29 ± 12.33 | <0.01 | −0.203 | <0.05 | −0.260 | N.S. |
| VAL | 263.10 ± 59.97 | 248.92 ± 46.79 | N.S. | −0.474 | <0.0001 | −0.213 | N.S. |
| MET | 25.58 ± 6.40 | 20.10 ± 4.60 | <0.0001 | −0.164 | N.S. | −0.277 | <0.05 |
| TRP | 53.45± 15.68 | 46.71± 13.26 | <0.0001 | −0.636 | <0.0001 | −0.517 | <0.0001 |
| PHE | 61.44± 11.62 | 54.99± 10.32 | <0.0001 | −0.244 | <0.02 | −0.289 | <0.05 |
| ILE | 69.63 ± 20.66 | 63.84 ± 13.99 | N.S. | −0.266 | <0.01 | −0.299 | <0.03 |
| LEU | 132.76 ± 34.49 | 115.73 ± 24.23 | <0.02 | −0.396 | <0.0001 | −0.283 | <0.05 |
| ORN | 156.00 ±43.57 | 96.76 ±28.38 | <0.0001 | 0.077 | N.S. | −0.163 | N.S. |
| LYS | 204.68 ± 47.71 | 209.80 ± 45.10 | N.S. | −0.072 | N.S. | −0.096 | N.S. |
| Compound | Concentrations in Serum of YnonDSP (n = 55) μmol/L Serum | Concentrations in Serum of AnonDSP (n = 47) μmol/L serum | Significantly Different from YnonDSP q-Values | Concentrations in Seraum of YDSP (n = 29) μmol/L Serum | Concentrations in Serum of ADSP (n = 27) μmol/L Serum | Significantly Different from YDSP q-Values |
|---|---|---|---|---|---|---|
| Uracil | 0.69 ± 0.54 | 3.01 ± 1.34 | <0.0001 | 0.44 ± 0.26 | 0.75 ± 0.39 | <0.002 |
| β-pseudouridine | 2.13 ± 1.04 | 1.53 ± 1.07 | <0.005 | 4.17 ± 1.58 | 3.89 ± 1.53 | N.S. |
| Uridine | 5.71 ± 2.38 | 7.07 ± 4.01 | N.S. | 12.78 ± 8.38 | 12.48 ± 8.16 | N.S. |
| Hypoxanthine | 4.95 ± 2.09 | 5.74 ± 2.66 | N.S. | 10.81 ± 4.39 | 10.64 ± 4.12 | N.S. |
| Xanthine | 2.74 ± 1.57 | 3.00 ±2.21 | N.S. | 13.24 ± 3.86 | 25.80 ± 5.18 | <0.02 |
| Uric acid | 285.02 ± 55.91 | 272.40 ± 136.47 | N.S. | 391.26 ± 69.54 | 430.95 ± 60.38 | <0.03 |
| Sum of oxypurines | 292.20 ± 56.55 | 286.87 ± 141.34 | N.S. | 417.45 ± 72.74 | 463.49 ± 60.95 | <0.02 |
| Inosine | 0.76 ± 0.51 | 0.79 ± 0.81 | N.S. | 2.32 ± 1.57 | 1.97 ± 1.53 | N.S. |
| Ascorbic acid | 51.08 ± 18.67 | 53.22 ± 21.37 | N.S. | 47.27 ± 17.19 | 37.83 ± 15.71 | <0.05 |
| GSH | 26.13 ± 6.96 | 23.97 ± 8.40 | N.S. | 22.26 ± 8.75 | 16.88 ± 8.00 | <0.02 |
| Nitrite | 1.64 ± 0.70 | 1.82 ± 0.93 | N.S. | 2.49 ± 1.58 | 2.27 ± 0.99 | N.S. |
| Nitrate | 30.35 ± 12.72 | 33.06 ± 15.22 | N.S. | 74.70 ± 31.57 | 92.53 ± 24.93 | <0.001 |
| Nitrite + Nitrate | 31.99 ± 12.78 | 34.88 ± 15.42 | N.S. | 77.19 ± 31.87 | 94.80 ± 24.59 | <0.01 |
| Lactate | 1.13 ± 0.30 | 1.05 ± 0.22 | N.S. | 1.62 ± 0.58 | 2.16 ± 0.62 | <0.001 |
| Creatinine | 63.47 ± 16.20 | 75.88 ± 34.95 | <0.03 | 73.15 ± 20.46 | 78.86 ± 15.77 | N.S. |
| Compound | Concentrations in Serum of YnonDSP (n = 55) μmol/L Serum | Concentrations in Serum of AnonDSP (n = 47) μmol/L Serum | Significantly Different from YnonDSP q-Values | Concentrations in Serum of YDSP (n = 29) μmol/L Serum | Concentrations in Serum of ADSP (n = 27) μmol/l serum | Significantly Different from YDSP q-Values |
|---|---|---|---|---|---|---|
| ASP | 52.65 ± 17.43 | 67.92 ± 17.47 | <0.001 | 44.06 ± 10.47 | 51.91 ± 8.76 | <0.005 |
| GLU | 71.92 ± 21.95 | 61.30 ± 13.14 | <0.001 | 53.42 ± 14.55 | 38.20 ± 12.61 | <0.001 |
| ASN | 66.21 ± 12.93 | 60.66 ± 8.59 | <0.02 | 55.94 ± 13.19 | 53.54 ± 11.82 | N.S. |
| SER | 95.37 ± 22.16 | 90.29 ± 18.81 | N.S. | 80.81 ± 19.90 | 76.64± 13.65 | N.S. |
| GLN | 369.80 ± 77.15 | 358.85 ± 56.05 | N.S. | 458.10 ± 85.40 | 477.70 ± 70.40 | N.S. |
| HIS | 73.14 ± 19.21 | 67.51 ± 16.94 | N.S. | 65.61 ± 13.26 | 58.35 ± 14.11 | N.S. |
| GLY | 225.76 ± 79.84 | 212.83 ± 74.99 | N.S. | 202.71 ± 40.90 | 223.50± 70.19 | N.S. |
| THR | 129.48 ± 28.37 | 132.76 ± 34.48 | N.S. | 96.03 ± 28.21 | 95.29 ± 22.15 | N.S. |
| CITR | 26.50 ± 8.52 | 34.57 ± 8.05 | <0.0001 | 31.31 ± 10.87 | 36.06 ± 8.89 | <0.025 |
| ARG | 32.34 ± 9.15 | 33.80 ± 11.37 | N.S. | 42.81 ± 12.90 | 42.14 ± 11.20 | N.S. |
| ALA | 383.78 ± 102.42 | 358.14 ± 89.11 | N.S. | 366.34 ± 122.14 | 399.67 ± 115.63 | N.S. |
| TAU | 100.99 ± 22.60 | 87.88 ± 20.38 | <0.0001 | 56.93 ± 20.20 | 56.04 ± 18.06 | N.S. |
| TYR | 77.11 ± 19.16 | 71.00 ± 15.69 | N.S. | 58.85 ± 11.87 | 52.96 ± 11.88 | N.S. |
| VAL | 274.21 ± 62.28 | 250.24 ± 47.47 | N.S. | 255.29 ± 54.88 | 239.58 ± 35.19 | N.S. |
| MET | 26.27 ± 5.35 | 24.60 ± 7.61 | N.S. | 21.62 ± 5.13 | 18.39 ± 3.24 | <0.02. |
| TRP | 64.42 ± 13.00 | 41.86 ± 11.52 | <0.0001 | 52.04 ± 10.70 | 40.79 ± 13.48 | <0.005 |
| PHE | 65.26 ± 13.00 | 57.95 ± 8.66 | <0.0001 | 57.50 ± 11.49 | 52.99 ± 8.16 | N.S. |
| ILE | 73.18 ± 18.37 | 65.48 ± 22.56 | N.S. | 65.79 ± 15.26 | 59.31 ± 11.21 | N.S. |
| LEU | 139.22 ± 32.14 | 123.19 ± 37.32 | <0.002 | 121.02 ± 27.11 | 110.04 ± 19.64 | N.S. |
| ORN | 151.40 ± 43.03 | 161.30 ± 44.04 | N.S. | 104.29 ± 31.20 | 88.14 ± 21.92 | N.S. |
| LYS | 203.09 ± 42.86 | 206.97 ± 53.70 | N.S. | 216.54 ± 49.25 | 199.33 ± 38.64 | N.S. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazzarino, G.; Amorini, A.M.; Mangione, R.; Saab, M.W.; Di Stasio, E.; Di Rosa, M.; Tavazzi, B.; Lazzarino, G.; Onder, G.; Carfì, A. Biochemical Discrimination of the Down Syndrome-Related Metabolic and Oxidative/Nitrosative Stress Alterations from the Physiologic Age-Related Changes through the Targeted Metabolomic Analysis of Serum. Antioxidants 2022, 11, 1208. https://doi.org/10.3390/antiox11061208
Lazzarino G, Amorini AM, Mangione R, Saab MW, Di Stasio E, Di Rosa M, Tavazzi B, Lazzarino G, Onder G, Carfì A. Biochemical Discrimination of the Down Syndrome-Related Metabolic and Oxidative/Nitrosative Stress Alterations from the Physiologic Age-Related Changes through the Targeted Metabolomic Analysis of Serum. Antioxidants. 2022; 11(6):1208. https://doi.org/10.3390/antiox11061208
Chicago/Turabian StyleLazzarino, Giacomo, Angela M. Amorini, Renata Mangione, Miriam Wissam Saab, Enrico Di Stasio, Michelino Di Rosa, Barbara Tavazzi, Giuseppe Lazzarino, Graziano Onder, and Angelo Carfì. 2022. "Biochemical Discrimination of the Down Syndrome-Related Metabolic and Oxidative/Nitrosative Stress Alterations from the Physiologic Age-Related Changes through the Targeted Metabolomic Analysis of Serum" Antioxidants 11, no. 6: 1208. https://doi.org/10.3390/antiox11061208