
PPARδ Inhibits Hyperglycemia-Triggered Senescence of Retinal Pigment Epithelial Cells by Upregulating SIRT1


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. SA β-Gal Staining
2.4. Western Blot Analysis
2.5. Confocal Immunofluorescence Microscopy
2.6. Generation of ARPE-19 Cells Stably Expressing shRNA
2.7. ROS Measurement
2.8. RNA Extraction and Real-Time PCR Analysis
2.9. Statistical Analysis
3. Results
3.1. Glucose Induces Premature Senescence of ARPE-19 Cells
3.2. Activation of PPARδ, but Not of PPARα or PPARγ, Suppresses HG-Triggered Premature Senescence of ARPE-19 Cells
3.3. GW501516-Activated PPARδ Attenuates HG-Triggered Generation of ROS
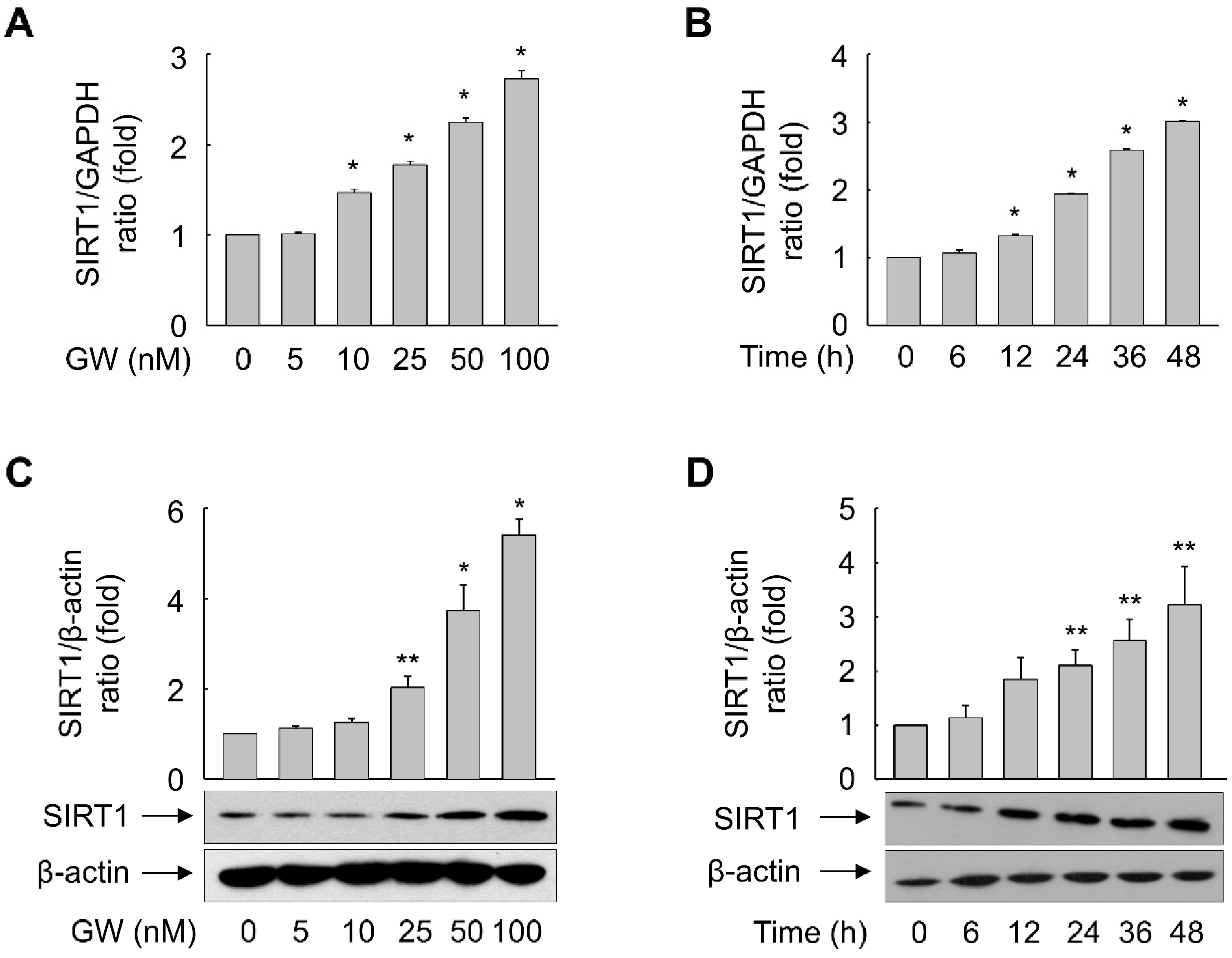
3.4. GW501516-Activated PPARδ Increases mRNA and Protein Expression of SIRT1 in ARPE-19 Cells
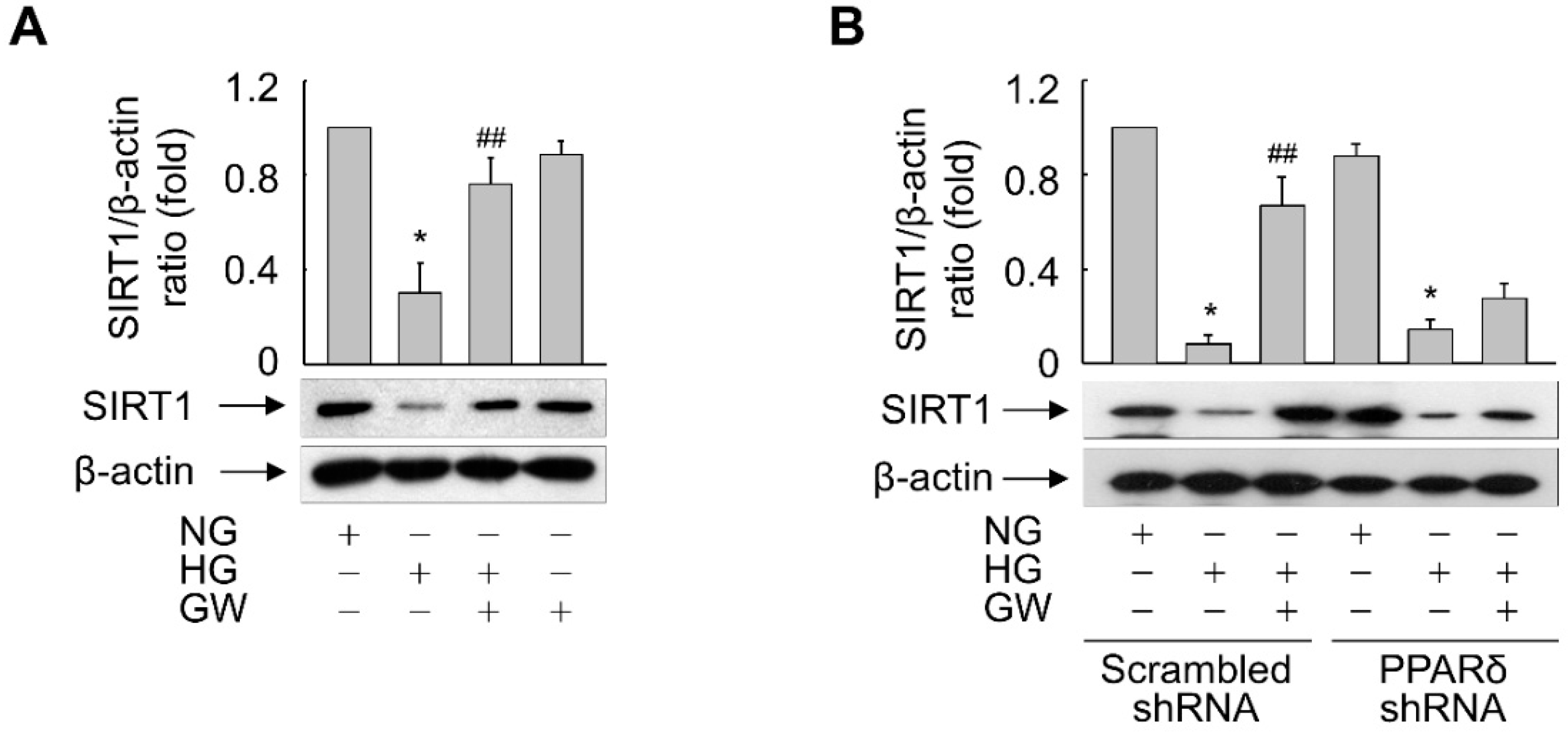
3.5. SIRT1 Is Essential for PPARδ-Mediated Suppression of HG-Triggered Premature Senescence of ARPE-19 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bojic, L.A.; Huff, M.W. Peroxisome proliferator-activated receptor δ: A multifaceted metabolic player. Curr. Opin. Lipidol. 2013, 24, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Ham, S.A.; Kim, M.Y.; Hwang, J.S.; Lee, H.; Kang, E.S.; Yoo, T.; Woo, I.S.; Yabe-Nishimura, C.; Paek, K.S.; et al. PPARδ coordinates angiotensin II-induced senescence in vascular smooth muscle cells through PTEN-mediated inhibition of superoxide generation. J. Biol. Chem. 2011, 286, 44585–44593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.Y.; Kang, E.S.; Ham, S.A.; Hwang, J.S.; Yoo, T.S.; Lee, H.; Paek, K.S.; Park, C.; Lee, H.T.; Kim, J.-H.; et al. The PPARδ-mediated inhibition of angiotensin II-induced premature senescence in human endothelial cells is SIRT1-dependent. Biochem. Pharm. 2012, 84, 1627–1634. [Google Scholar] [CrossRef] [PubMed]
- Higashiyama, H.; Billin, A.N.; Okamoto, Y.; Kinoshita, M.; Asano, S. Expression profiling of peroxisome proliferator-activated receptor-delta (PPAR-delta) in mouse tissues using tissue microarray. Histochem. Cell Biol. 2007, 127, 485–494. [Google Scholar] [CrossRef]
- Kim, H.J.; Ham, S.A.; Paek, K.S.; Hwang, J.S.; Jung, S.Y.; Kim, M.Y.; Jin, H.; Kang, E.S.; Woo, I.S.; Kim, H.J.; et al. Transcriptional up-regulation of antioxidant genes by PPARδ inhibits angiotensin II-induced premature senescence in vascular smooth muscle cells. Biochem. Biophys. Res. Commun. 2011, 406, 564–569. [Google Scholar] [CrossRef]
- Ham, S.A.; Hwang, J.S.; Yoo, T.; Lee, H.; Kang, E.S.; Park, C.; Oh, J.-W.; Lee, H.T.; Min, G.; Kim, J.-H.; et al. Ligand-activated PPARδ inhibits UVB-induced senescence of human keratinocytes via PTEN-mediated inhibition of superoxide production. Biochem. J. 2012, 444, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Simó, R.; Villarroel, M.; Corraliza, L.; Hernández, C.; Garcia-Ramírez, M. The retinal pigment epithelium: Something more than a constituent of the blood-retinal barrier--implications for the pathogenesis of diabetic retinopathy. J. Biomed. Biotechnol. 2010, 2010, 190724. [Google Scholar] [CrossRef]
- Shao, J.; Yao, Y. Repression of retinal microvascular endothelial cells by transthyretin under simulated diabetic retinopathy conditions. Int. J. Ophthalmol. 2016, 9, 809–815. [Google Scholar] [CrossRef]
- Packer, L.; Kraemer, K.; Rimbach, G. Molecular aspects of lipoic acid in the prevention of diabetes complications. Nutrition 2001, 17, 888–895. [Google Scholar] [CrossRef]
- Zhong, Q.; Mishra, M.; Kowluru, R.A. Transcription factor Nrf2-mediated antioxidant defense system in the development of diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2013, 54, 3941–3948. [Google Scholar] [CrossRef]
- Yu, A.L.; Fuchshofer, R.; Kook, D.; Kampik, A.; Bloemendal, H.; Welge-Lüssen, U. Subtoxic oxidative stress induces senescence in retinal pigment epithelial cells via TGF-beta release. Investig. Ophthalmol. Vis. Sci. 2009, 50, 926–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuge, C.-C.; Xu, J.-Y.; Zhang, J.; Li, W.; Li, P.; Li, Z.; Chen, L.; Liu, X.; Shang, P.; Xu, H.; et al. Fullerenol protects retinal pigment epithelial cells from oxidative stress-induced premature senescence via activating SIRT1. Investig. Ophthalmol. Vis. Sci. 2014, 55, 4628–4638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Wan, Y.; Zhou, S.; Lu, Y.; Zhang, Z.; Zhang, R.; Chen, F.; Hao, D.; Zhao, X.; Guo, Z.; et al. Endothelium-specific SIRT1 overexpression inhibits hyperglycemia-induced upregulation of vascular cell senescence. Sci. China Life Sci. 2012, 55, 467–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senthil, K.K.J.; Gokila, V.M.; Mau, J.-L.; Lin, C.-C.; Chu, F.-H.; Wei, C.-C.; Liao, V.H.-C.; Wang, S.-Y. A steroid like phytochemical Antcin M is an anti-aging reagent that eliminates hyperglycemia-accelerated premature senescence in dermal fibroblasts by direct activation of Nrf2 and SIRT-1. Oncotarget 2016, 7, 62836–62861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.-I.; Park, M.-J.; Choi, J.-H.; Lim, S.-K.; Choi, H.-J.; Park, S.-H. Hyperglycemia-induced GLP-1R downregulation causes RPE cell apoptosis. Int. J. Biochem. Cell Biol. 2015, 59, 41–51. [Google Scholar] [CrossRef]
- Michan, S.; Sinclair, D. Sirtuins in mammals: Insights into their biological function. Biochem. J. 2007, 404, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Vaquero, A.; Scher, M.; Lee, D.; Erdjument-Bromage, H.; Tempst, P.; Reinberg, D. Human SirT1 interacts with histone H1 and promotes formation of facultative heterochromatin. Mol. Cell 2004, 16, 93–105. [Google Scholar] [CrossRef]
- Vaziri, H.; Dessain, S.K.; Ng Eaton, E.; Imai, S.I.; Frye, R.A.; Pandita, T.K.; Guarente, L.; Weinberg, R.A. hSIR2(SIRT1) functions as an NAD-dependent p53 deacetylase. Cell 2001, 107, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Zhou, N.; Lin, X.; Dong, W.; Huang, W.; Jiang, W.; Lin, L.; Qiu, Q.; Zhang, X.; Shen, J.; Song, Z.; et al. SIRT1 alleviates senescence of degenerative human intervertebral disc cartilage endo-plate cells via the p53/p21 pathway. Sci. Rep. 2016, 6, 22628. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-I.; Park, M.-J.; Choi, J.-H.; Kim, I.-S.; Han, H.-J.; Yoon, K.-C.; Park, S.-W.; Lee, M.-Y.; Oh, K.-S.; Park, S.-H. PRMT1 and PRMT4 Regulate Oxidative Stress-Induced Retinal Pigment Epithelial Cell Damage in SIRT1-Dependent and SIRT1-Independent Manners. Oxid. Med. Cell Longev. 2015, 2015, 617919. [Google Scholar] [CrossRef] [Green Version]
- Okazaki, M.; Iwasaki, Y.; Nishiyama, M.; Taguchi, T.; Tsugita, M.; Nakayama, S.; Kambayashi, M.; Hashimoto, K.; Terada, Y. PPARbeta/delta regulates the human SIRT1 gene transcription via Sp1. Endocr. J. 2010, 57, 403–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, T.; Ham, S.A.; Lee, W.J.; Hwang, S.I.; Park, J.-A.; Hwang, J.S.; Hur, J.; Shin, H.-C.; Han, S.G.; Lee, C.-H.; et al. Ligand-Dependent Interaction of PPARδ With T-Cell Protein Tyrosine Phosphatase 45 Enhances Insulin Signaling. Diabetes 2018, 67, 360–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, T.; Ham, S.A.; Hwang, J.S.; Lee, W.J.; Paek, K.S.; Oh, J.W.; Kim, J.H.; Do, J.T.; Han, C.W.; Kim, J.H.; et al. Peroxisome proliferator-activated receptor δ inhibits Porphyromonas gingivalis lipopolysaccharide-induced activation of matrix metalloproteinase-2 by downregulating NADPH oxidase 4 in human gingival fibroblasts. Mol. Oral Microbiol. 2016, 31, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Ham, S.A.; Kim, E.; Yoo, T.; Lee, W.J.; Youn, J.H.; Choi, M.-J.; Han, S.G.; Lee, C.-H.; Paek, K.S.; Hwang, J.S.; et al. Ligand-activated interaction of PPARδ with c-Myc governs the tumorigenicity of breast cancer. Int. J. Cancer 2018, 143, 2985–2996. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.S.; Choi, H.S.; Ham, S.A.; Yoo, T.; Lee, W.J.; Paek, K.S.; Seo, H.G. Deacetylation-mediated interaction of SIRT1-HMGB1 improves survival in a mouse model of endotoxemia. Sci. Rep. 2015, 5, 15971. [Google Scholar] [CrossRef] [Green Version]
- Campisi, J. Cellular senescence as a tumor-suppressor mechanism. Trends Cell Biol. 2001, 11, S27–S31. [Google Scholar] [CrossRef] [Green Version]
- Panebianco, C.; Oben, J.A.; Vinciguerra, M.; Pazienza, V. Senescence in hepatic stellate cells as a mechanism of liver fibrosis reversal: A putative synergy between retinoic acid and PPAR-gamma signalings. Clin. Exp. Med. 2017, 17, 269–280. [Google Scholar] [CrossRef]
- Jung, K.J.; Ishigami, A.; Maruyama, N.; Takahashi, R.; Goto, S.; Yu, B.P.; Chung, H.Y. Modulation of gene expression of SMP-30 by LPS and calorie restriction during aging process. Exp. Gerontol. 2004, 39, 1169–1177. [Google Scholar] [CrossRef]
- Lawless, C.; Wang, C.; Jurk, D.; Merz, A.; Zglinicki, T.v.; Passos, J.F. Quantitative assessment of markers for cell senescence. Exp. Gerontol. 2010, 45, 772–778. [Google Scholar] [CrossRef]
- Miyazaki, R.; Ichiki, T.; Hashimoto, T.; Inanaga, K.; Imayama, I.; Sadoshima, J.; Sunagawa, K. SIRT1, a longevity gene, downregulates angiotensin II type 1 receptor expression in vascular smooth muscle cells. Arter. Thromb. Vasc. Biol. 2008, 28, 1263–1269. [Google Scholar] [CrossRef] [Green Version]
- Imanishi, T.; Kobayashi, K.; Kuroi, A.; Ikejima, H.; Akasaka, T. Pioglitazone inhibits angiotensin II-induced senescence of endothelial progenitor cell. Hypertens. Res. 2008, 31, 757–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, Q.; Huang, J.; Zhou, R.; Niu, J.; Zhu, X.; Wang, J.; Zhang, Z.; Tong, T. PPAR{gamma} accelerates cellular senescence by inducing p16INK4{alpha} expression in human diploid fibroblasts. J. Cell Sci. 2008, 121, 2235–2245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustapha, N.M.; Tarr, J.M.; Kohner, E.M.; Chibber, R. NADPH Oxidase versus Mitochondria-Derived ROS in Glucose-Induced Apoptosis of Pericytes in Early Diabetic Retinopathy. J. Ophthalmol. 2010, 2010, 746978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, H.Y.; Lavu, S.; Bitterman, K.J.; Hekking, B.; Imahiyerobo, T.A.; Miller, C.; Frye, R.; Ploegh, H.; Kessler, B.M.; Sinclair, D.A. Acetylation of the C terminus of Ku70 by CBP and PCAF controls Bax-mediated apoptosis. Mol. Cell 2004, 13, 627–638. [Google Scholar] [CrossRef]
- Langley, E.; Pearson, M.; Faretta, M.; Bauer, U.-M.; Frye, R.A.; Minucci, S.; Pelicci, P.G.; Kouzarides, T. Human SIR2 deacetylates p53 and antagonizes PML/p53-induced cellular senescence. EMBO J. 2002, 21, 2383–2396. [Google Scholar] [CrossRef] [Green Version]
- Boily, G.; Seifert, E.L.; Bevilacqua, L.; He, X.H.; Sabourin, G.; Estey, C.; Moffat, C.; Crawford, S.; Saliba, S.; Jardine, K.; et al. SirT1 regulates energy metabolism and response to caloric restriction in mice. PLoS ONE 2008, 3, e1759. [Google Scholar] [CrossRef]
- Cohen, H.Y.; Miller, C.; Bitterman, K.J.; Wall, N.R.; Hekking, B.; Kessler, B.; Howitz, K.T.; Gorospe, M.; de Cabo, R.; Sinclair, D.A. Calorie restriction promotes mammalian cell survival by inducing the SIRT1 deacetylase. Science 2004, 305, 390–392. [Google Scholar] [CrossRef] [Green Version]
- Coll, T.; Rodrïguez-Calvo, R.; Barroso, E.; Serrano, L.; Eyre, E.; Palomer, X.; Vázquez-Carrera, M. Peroxisome proliferator-activated receptor (PPAR) beta/delta: A new potential therapeutic target for the treatment of metabolic syndrome. Curr. Mol. Pharm. 2009, 2, 46–55. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, E.J.; Won, J.P.; Lee, H.G.; Kim, E.; Hur, J.; Lee, W.J.; Hwang, J.S.; Seo, H.G. PPARδ Inhibits Hyperglycemia-Triggered Senescence of Retinal Pigment Epithelial Cells by Upregulating SIRT1. Antioxidants 2022, 11, 1207. https://doi.org/10.3390/antiox11061207
Lee EJ, Won JP, Lee HG, Kim E, Hur J, Lee WJ, Hwang JS, Seo HG. PPARδ Inhibits Hyperglycemia-Triggered Senescence of Retinal Pigment Epithelial Cells by Upregulating SIRT1. Antioxidants. 2022; 11(6):1207. https://doi.org/10.3390/antiox11061207
Chicago/Turabian StyleLee, Eun Ji, Jun Pil Won, Hyuk Gyoon Lee, Eunsu Kim, Jinwoo Hur, Won Jin Lee, Jung Seok Hwang, and Han Geuk Seo. 2022. "PPARδ Inhibits Hyperglycemia-Triggered Senescence of Retinal Pigment Epithelial Cells by Upregulating SIRT1" Antioxidants 11, no. 6: 1207. https://doi.org/10.3390/antiox11061207