
Anti-Inflammatory and Anti-Acne Effects of Hamamelis virginiana Bark in Human Keratinocytes

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phytochemical Characterization
2.2. Analysis of Condensed Tannins
2.3. Folin–Ciocalteu Assay
2.4. ORAC Test
2.5. DPPH Test
2.6. Cell Culture
2.7. Cutibacterium Acnes Culture and Infection
2.8. Measurement of the Antibacterial Activity
2.9. Cell Viability
2.10. HaCaT Cell Treatment
2.11. Measurement of Intracellular ROS
2.12. Measurement of CAT Activity
2.13. Measurement of the Release of Inflammatory Mediators: IL-8 and VEGF-A
2.14. Measurement of NO Release
2.15. Measurement of the NF-κB-Driven Transcription
2.16. Statistical Analysis
3. Results
3.1. Phytochemical Analysis of HVE
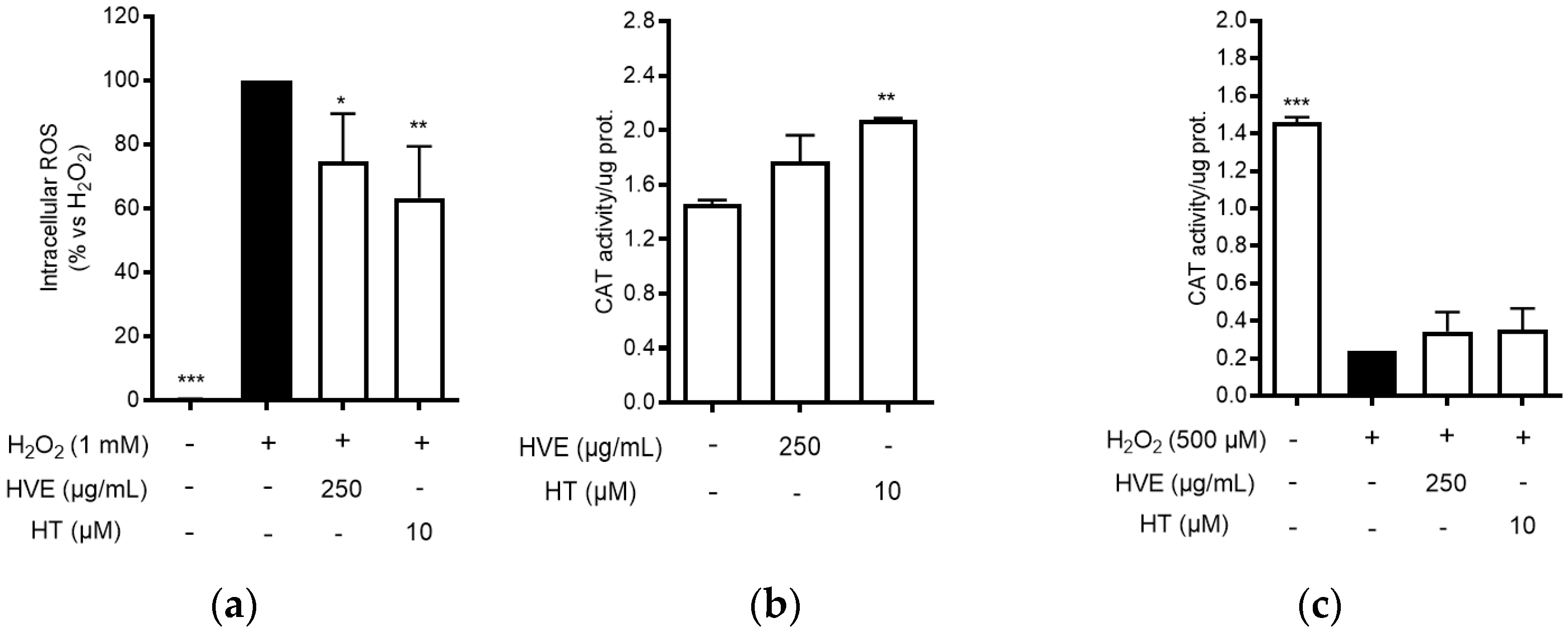
3.2. HVE Inhibits Intracellular ROS Production
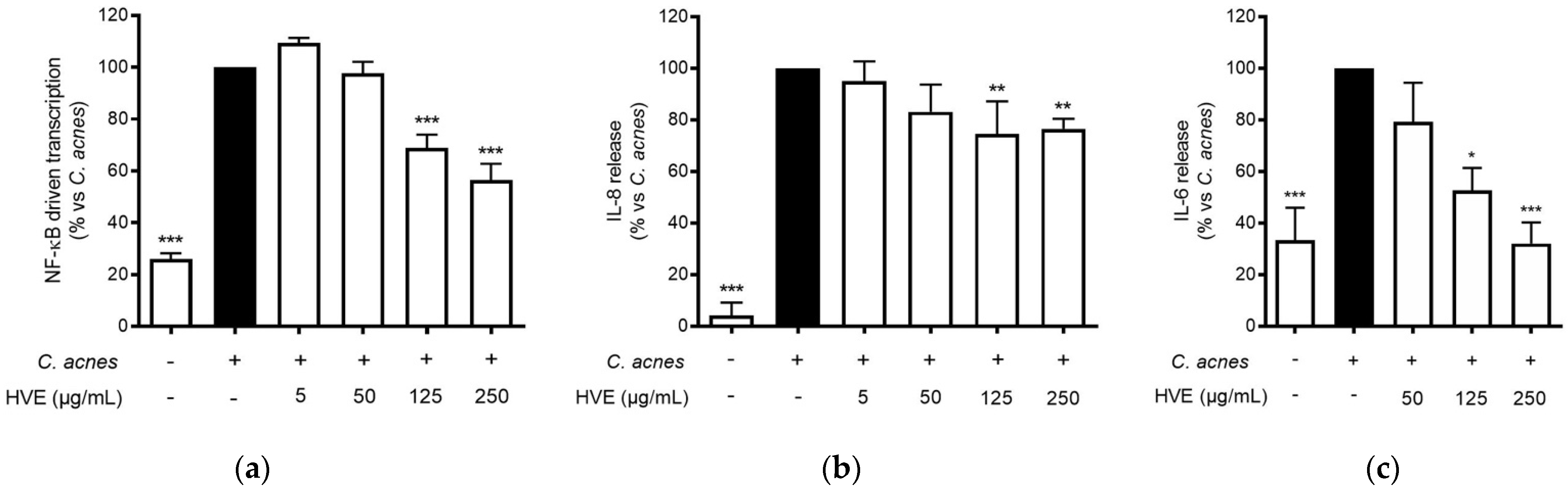
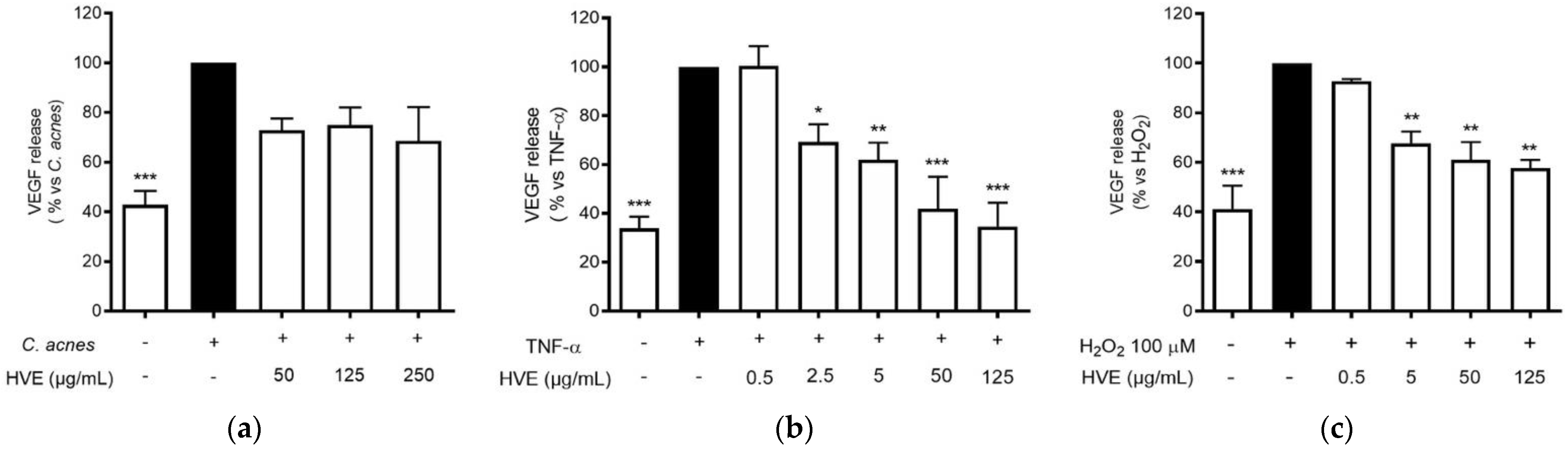
3.3. HVE Inhibits C. acnes-Induced Inflammation in HaCaT Cells
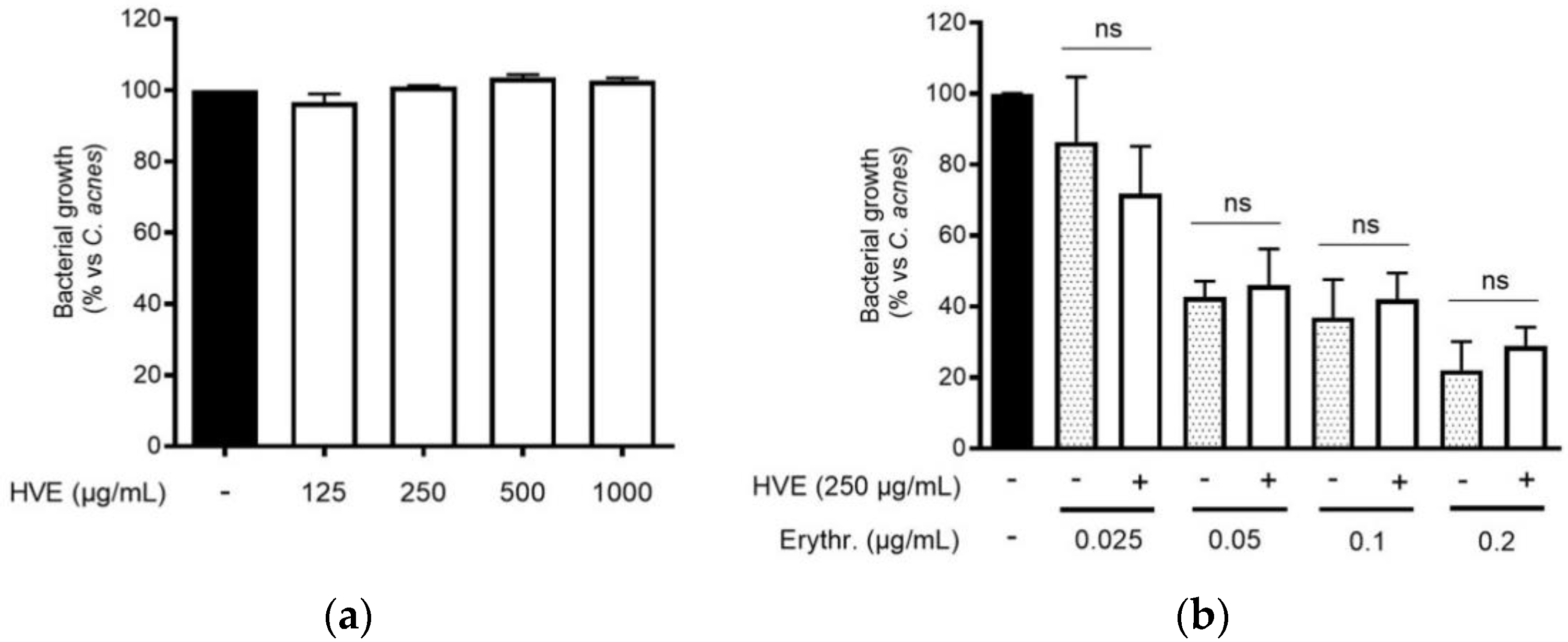
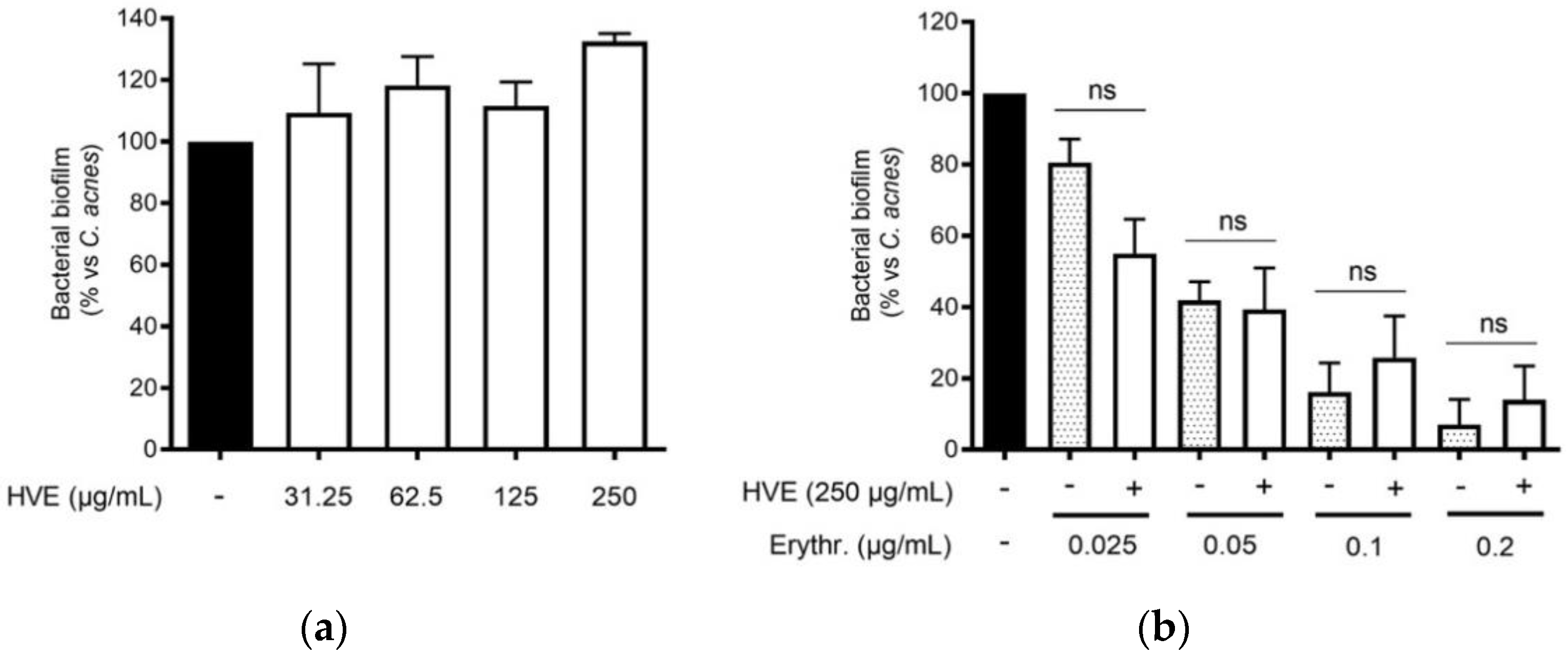
3.4. The Bioactivity of HVE Is Independent of the Antibacterial Effect on C. acnes
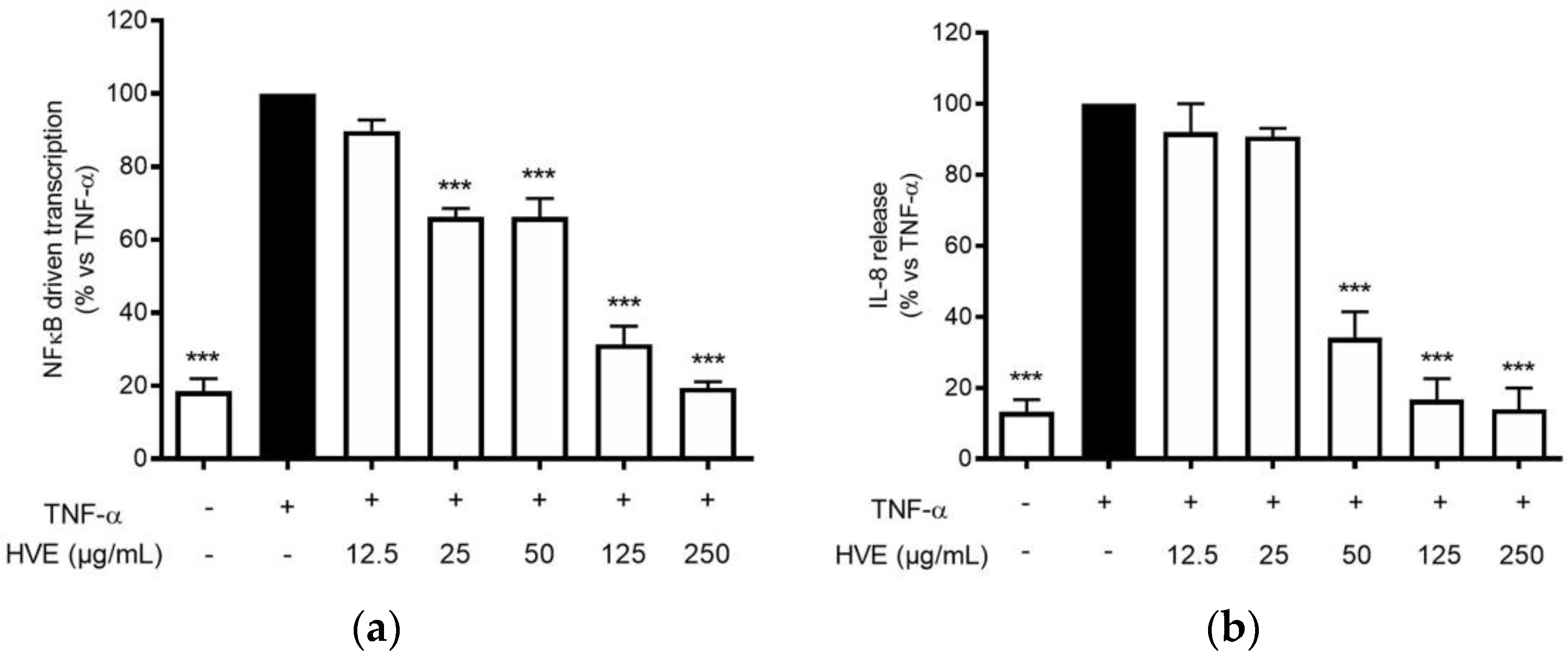
3.5. HVE Inhibits TNF-α-Induced Inflammation in HaCaT Cells
3.6. Comparison of Pro-Inflammatory and Pro-Oxidant Stimuli on the Effect of HVE
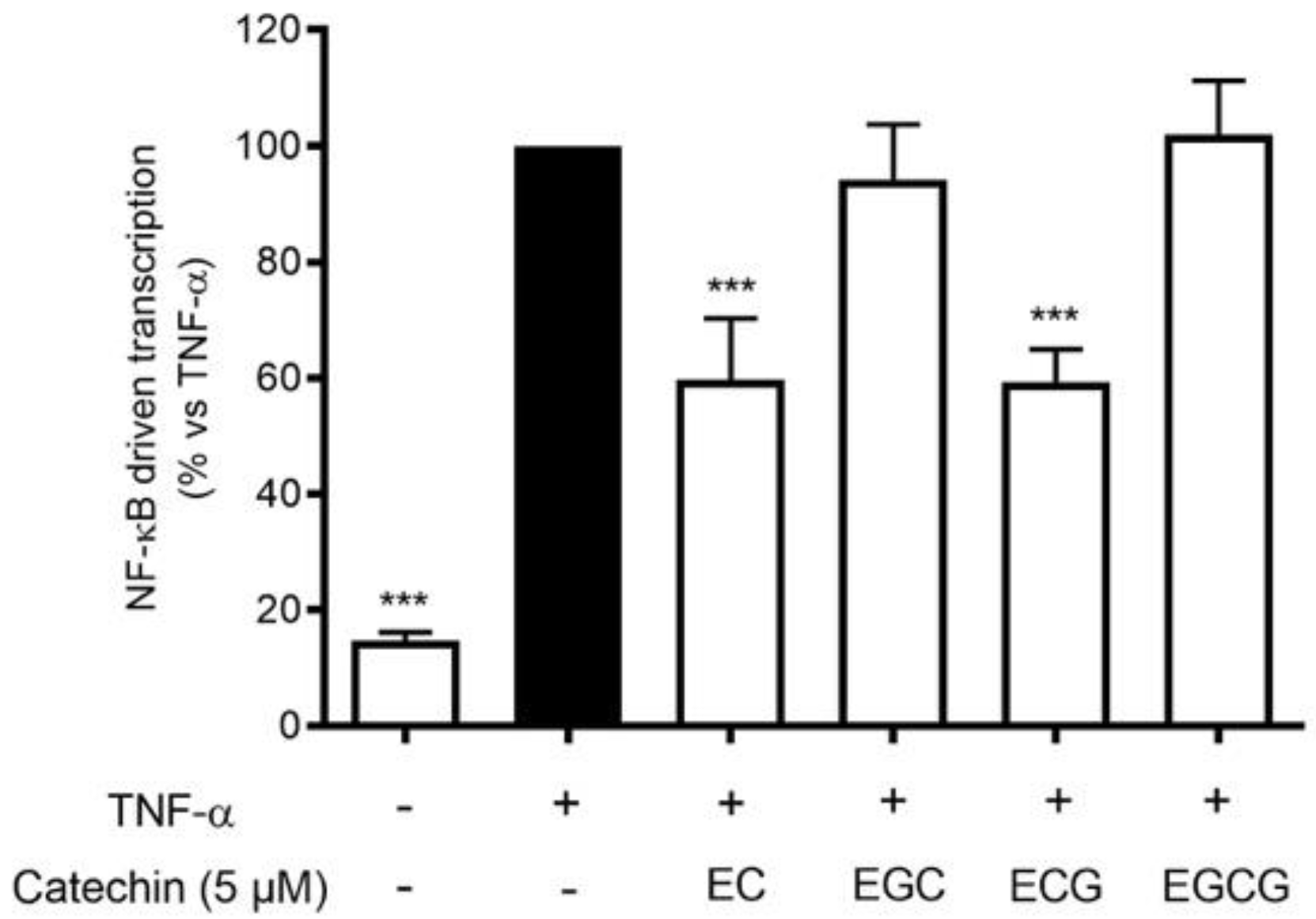
3.7. Contribution of Catechins to the NF-κB Inhibitory Activity of HVE
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhate, K.; Williams, H.C. Epidemiology of acne vulgaris. Br. J. Dermatol. 2013, 168, 474–485. [Google Scholar] [CrossRef] [PubMed]
- Kim, J. Review of the innate immune response in acne vulgaris: Activation of Toll-like receptor 2 in acne triggers inflammatory cytokine responses. Dermatology 2005, 211, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.F.; Hsieh, Y.D.; Lin, Y.C.; Two, A.; Shu, C.W.; Huang, C.M. Propionibacterium acnes in the Pathogenesis and Immunotherapy of Acne Vulgaris. Curr. Drug Metab. 2015, 16, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Azimi, H.; Fallah-Tafti, M.; Khakshur, A.A.; Abdollahi, M. A review of phytotherapy of acne vulgaris: Perspective of new pharmacological treatments. Fitoterapia 2012, 83, 1306–1317. [Google Scholar] [CrossRef]
- Moerman, D.E. Medicinal Plants of Native America; University of Michigan Press: Ann Arbor, MI, USA, 1986; Volume 1. [Google Scholar]
- Committee on Herbal Medicinal Products. Final Community Herbal Monograph on Hamamelis virginiana L., Folium; Doc. Ref.: EMA/HMPC/114586/2008; European Medicines Agency: London, UK, 2010.
- Committee on Herbal Medicinal Products. Final Community Herbal Monograph on Hamamelis Virginiana L., Cortex; Doc. Ref.: EMA/HMPC/114583/2008; European Medicines Agency: London, UK, 2019.
- Committee on Herbal Medicinal Products. Community Herbal Monograph on Hamamelis virginiana L., Folium et Cortex aut Ramunculus Destillatum; Doc. Ref.: EMA/HMPC/114584/2008; European Medicines Agency: London, UK, 2010.
- European Scientific Cooperative on Phytotherapy (ESCOP). Hamamelidis cortex (Hamamelis bark); European Scientific Cooperative on Phytotherapy (ESCOP): Exeter, UK, 2012; p. 15. [Google Scholar]
- Committee on Herbal Medicinal Products. Assessment Report on Hamamelis Virginiana l., Cortex Hamamelis Virginiana l., Folium Hamamelis Virginiana l., Folium et Cortex aut Ramunculus Destillatum; Doc. Ref.: EMA/HMPC/114585/2008; European Medicines Agency: London, UK, 2010.
- Wolff, H.H.; Kieser, M. Hamamelis in children with skin disorders and skin injuries: Results of an observational study. Eur. J. Pediatr. 2007, 166, 943–948. [Google Scholar] [CrossRef]
- Hughes-Formella, B.J.; Filbry, A.; Gassmueller, J.; Rippke, F. Anti-inflammatory efficacy of topical preparations with 10% hamamelis distillate in a UV erythema test. Ski. Pharm. Appl Ski. Physiol. 2002, 15, 125–132. [Google Scholar] [CrossRef]
- Hughes-Formella, B.J.; Bohnsack, K.; Rippke, F.; Benner, G.; Rudolph, M.; Tausch, I.; Gassmueller, J. Anti-inflammatory effect of hamamelis lotion in a UVB erythema test. Dermatology 1998, 196, 316–322. [Google Scholar] [CrossRef]
- Korting, H.C.; Schäfer-Korting, M.; Klövekorn, W.; Klövekorn, G.; Martin, C.; Laux, P. Comparative efficacy of hamamelis distillate and hydrocortisone cream in atopic eczema. Eur. J. Clin. Pharm. 1995, 48, 461–465. [Google Scholar] [CrossRef]
- Korting, H.C.; Schäfer-Korting, M.; Hart, H.; Laux, P.; Schmid, M. Anti-inflammatory activity of hamamelis distillate applied topically to the skin. Influence of vehicle and dose. Eur. J. Clin. Pharm. 1993, 44, 315–318. [Google Scholar] [CrossRef]
- Deters, A.; Dauer, A.; Schnetz, E.; Fartasch, M.; Hensel, A. High molecular compounds (polysaccharides and proanthocyanidins) from Hamamelis virginiana bark: Influence on human skin keratinocyte proliferation and differentiation and influence on irritated skin. Phytochemistry 2001, 58, 949–958. [Google Scholar] [CrossRef]
- Erdelmeier, C.A.; Cinatl, J., Jr.; Rabenau, H.; Doerr, H.W.; Biber, A.; Koch, E. Antiviral and antiphlogistic activities of Hamamelis virginiana bark. Planta Med. 1996, 62, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Hartisch, C.; Kolodziej, H.; von Bruchhausen, F. Dual inhibitory activities of tannins from Hamamelis virginiana and related polyphenols on 5-lipoxygenase and lyso-PAF: Acetyl-CoA acetyltransferase. Planta Med. 1997, 63, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Masaki, H.; Atsumi, T.; Sakurai, H. Evaluation of superoxide scavenging activities of hamamelis extract and hamamelitannin. Free. Radic. Res. Commun. 1993, 19, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Hamamelitannin from Hamamelis virginiana inhibits the tumour necrosis factor-alpha (TNF)-induced endothelial cell death in vitro. Toxicon 2002, 40, 83–88. [Google Scholar] [CrossRef]
- Touriño, S.; Lizárraga, D.; Carreras, A.; Lorenzo, S.; Ugartondo, V.; Mitjans, M.; Vinardell, M.P.; Juliá, L.; Cascante, M.; Torres, J.L. Highly galloylated tannin fractions from witch hazel (Hamamelis virginiana) bark: Electron transfer capacity, in vitro antioxidant activity, and effects on skin-related cells. Chem. Res. Toxicol. 2008, 21, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Cheesman, M.J.; Alcorn, S.; Verma, V.; Cock, I.E. An assessment of the growth inhibition profiles of Hamamelis virginiana L. extracts against Streptococcus and Staphylococcus spp. J. Tradit. Complement Med. 2021, 11, 457–465. [Google Scholar] [CrossRef]
- Iauk, L.; Lo Bue, A.M.; Milazzo, I.; Rapisarda, A.; Blandino, G. Antibacterial activity of medicinal plant extracts against periodontopathic bacteria. Phytother Res 2003, 17, 599–604. [Google Scholar] [CrossRef]
- Vrhovsek, U.; Masuero, D.; Gasperotti, M.; Franceschi, P.; Caputi, L.; Viola, R.; Mattivi, F. A Versatile Targeted Metabolomics Method for the Rapid Quantification of Multiple Classes of Phenolics in Fruits and Beverages. J. Agr. Food Chem. 2012, 60, 8831–8840. [Google Scholar] [CrossRef]
- Gasperotti, M.; Masuero, D.; Vrhovsek, U.; Guella, G.; Mattivi, F. Profiling and Accurate Quantification of Rubus Ellagitannins and Ellagic Acid Conjugates Using Direct UPLC-Q-TOF HDMS and HPLC-DAD Analysis. J. Agr. Food Chem. 2010, 58, 4602–4616. [Google Scholar] [CrossRef]
- Arapitsas, P.; Perenzoni, D.; Guella, G.; Mattivi, F. Improving the Phloroglucinolysis Protocol and Characterization of Sagrantino Wines Proanthocyanidins. Molecules 2021, 26, 1087. [Google Scholar] [CrossRef]
- Nwakiban, A.P.A.; Fumagalli, M.; Piazza, S.; Magnavacca, A.; Martinelli, G.; Beretta, G.; Magni, P.; Tchamgoue, A.D.; Agbor, G.A.; Kuiate, J.R.; et al. Dietary Cameroonian Plants Exhibit Anti-Inflammatory Activity in Human Gastric Epithelial Cells. Nutrients 2020, 12, 3787. [Google Scholar] [CrossRef]
- Fracassetti, D.; Gabrielli, M.; Costa, C.; Tomas-Barberan, F.A.; Tirelli, A. Characterization and suitability of polyphenols-based formulas to replace sulfur dioxide for storage of sparkling white wine. Food Control 2016, 60, 606–614. [Google Scholar] [CrossRef]
- Ng, N.S.; Ooi, L. A Simple Microplate Assay for Reactive Oxygen Species Generation and Rapid Cellular Protein Normalization. Bio Protoc. 2021, 11, e3877. [Google Scholar] [CrossRef]
- Suvanprakorn, P.; Tongyen, T.; Prakhongcheep, O.; Laoratthaphong, P.; Chanvorachote, P. Establishment of an Anti-acne Vulgaris Evaluation Method Based on TLR2 and TLR4-mediated Interleukin-8 Production. In Vivo 2019, 33, 1929–1934. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Ochoa, M.T.; Krutzik, S.R.; Takeuchi, O.; Uematsu, S.; Legaspi, A.J.; Brightbill, H.D.; Holland, D.; Cunliffe, W.J.; Akira, S.; et al. Activation of toll-like receptor 2 in acne triggers inflammatory cytokine responses. J. Immunol. 2002, 169, 1535–1541. [Google Scholar] [CrossRef] [Green Version]
- Zeng, R.; Xu, H.; Liu, Y.; Du, L.; Duan, Z.; Tong, J.; He, Y.; Chen, Q.; Chen, X.; Li, M. miR-146a Inhibits Biofilm-Derived Cutibacterium acnes-Induced Inflammatory Reactions in Human Keratinocytes. J. Invest. Dermatol. 2019, 139, 2488–2496.e4. [Google Scholar] [CrossRef]
- Abd El-Hamid, M.I.; ES, Y.E.-N.; T, M.K.; Hegazy, W.A.H.; Mosbah, R.A.; Nassar, M.S.; Bakhrebah, M.A.; Abdulaal, W.H.; Alhakamy, N.A.; Bendary, M.M. Promising Antibiofilm Agents: Recent Breakthrough against Biofilm Producing Methicillin-Resistant Staphylococcus aureus. Antibiotics 2020, 9, 667. [Google Scholar] [CrossRef]
- Toyoda, M.; Morohashi, M. Pathogenesis of acne. Med. Electron Microsc. 2001, 34, 29–40. [Google Scholar] [CrossRef]
- Dreno, B. What is new in the pathophysiology of acne, an overview. J. Eur. Acad. Dermatol. Venereol. 2017, 31 (Suppl. 5), 8–12. [Google Scholar] [CrossRef] [PubMed]
- Colombo, I.; Sangiovanni, E.; Maggio, R.; Mattozzi, C.; Zava, S.; Corbett, Y.; Fumagalli, M.; Carlino, C.; Corsetto, P.A.; Scaccabarozzi, D.; et al. HaCaT Cells as a Reliable In Vitro Differentiation Model to Dissect the Inflammatory/Repair Response of Human Keratinocytes. Mediat. Inflamm 2017, 2017, 7435621. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.H.; Lee, W.R.; Wang, P.H.; Cheng, K.T.; Chen, Y.C.; Shen, S.C. Propionibacterium acnes-induced iNOS and COX-2 protein expression via ROS-dependent NF-kappa B and AP-1 activation in macrophages. J. Dermatol. Sci. 2013, 69, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Mina, O.M.D.; Kartelishev, A.V.; Karpova, E.I.; Danischuk, O.I. Role of Cytokines in the Pathogenesis of Acne. Int. J. Biomed. 2017, 7, 37–40. [Google Scholar] [CrossRef]
- Sangiovanni, E.; di Lorenzo, C.; Piazza, S.; Manzoni, Y.; Brunelli, C.; Fumagalli, M.; Magnavacca, A.; Martinelli, G.; Colombo, F.; Casiraghi, A.; et al. Vitis vinifera L. Leaf Extract Inhibits In Vitro Mediators of Inflammation and Oxidative Stress Involved in Inflammatory-Based Skin Diseases. Antioxidants 2019, 8, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Compounds | Amount | |
|---|---|---|---|
| Phenolic acids | 4-hydroxybenzoic acid | µg/g | 2.68 |
| Vanillin | µg/g | 0.11 | |
| Vanillic acid | µg/g | 0.87 | |
| Protocatechuic acid | µg/g | 0.56 | |
| Methyl gallate | µg/g | 0.07 | |
| Ferulic acid | µg/g | 0.45 | |
| Ellagic acid | µg/g | 76.00 | |
| Flavonoids | Naringenin | µg/g | 0.22 |
| Quercetin-3-O-glucoside | µg/g | 0.56 | |
| Isorhamnetin-3-O-glucoside | µg/g | 0.56 | |
| Flavan-3-ols (catechins) | Catechin (free) | µg/g | 143.30 |
| Epicatechin (free) | µg/g | 0.67 | |
| Total flavanols monomers | µg/g | 230.13 | |
| Gallocatechin (free) | µg/g | 76.44 | |
| Epigallocatechin (free) | µg/g | 0.67 | |
| Gallotannins | Hamamelitannin | µg/g | 2910.21 |
| Procyanidins dimers | Procyanidin B1 | µg/g | 18.08 |
| Procyanidin B2 | µg/g | 0.00 | |
| Total procyanidins dimers | µg/g | 18.08 | |
| Oligomeric and polymeric proanthocyanidins | Catechin (after Phl.) | µg/g | 221.53 |
| Catechin (terminal units) | µg/g | 78.23 | |
| Epicatechin (after Phl.) | µg/g | 13.79 | |
| Epicatechin (terminal units) | µg/g | 13.12 | |
| Gallocatechin (after Phl.) | µg/g | 127.79 | |
| Gallocatechin (terminal units) | µg/g | 51.34 | |
| Epigallocatechin (after Phl.) | µg/g | 2.46 | |
| Epigallocatechin (terminal units) | µg/g | 1.79 | |
| Catechin Gallate + | |||
| Epicatechin Gallate (free) | µg/g | 9.03 | |
| Catechin Gallate + | |||
| Epicatechin Gallate (after Phl.) | µg/g | 20.21 | |
| Catechin Gallate + | |||
| Epicatechin Gallate (terminal) | µg/g | 11.15 | |
| Catechin-Phl. | µg/g | 0.00 | |
| Catechin + Epicatechin-Phl. | µg/g | 298.64 | |
| Epigallocatechin-Phl | µg/g | 310.02 | |
| Epicatechin Gallate-Phl | µg/g | 2213.26 | |
| % Galloylation (terminal units) | % | 3.21 | |
| % Galloylation (upper and extension units) | % | 78.43 | |
| Total terminal units | µg/g | 155.64 | |
| Total upper units | µg/g | 2821.92 | |
| Total polymeric proanthocyanidins | µg/g | 2977.56 | |
| aDP (average degree of polymerization) | 19.1 |
| ORAC Test | mmol Trolox eq./g HVE | ±SEM |
|---|---|---|
| HVE | 5.519 | 0.512 |
| HT | 0.389 | 0.002 |
| DPPH Test | µmol Trolox eq./g HVE | ±SEM |
|---|---|---|
| HVE | 90.223 | 0.284 |
| HT | 16.760 | 0.976 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piazza, S.; Martinelli, G.; Vrhovsek, U.; Masuero, D.; Fumagalli, M.; Magnavacca, A.; Pozzoli, C.; Canilli, L.; Terno, M.; Angarano, M.; et al. Anti-Inflammatory and Anti-Acne Effects of Hamamelis virginiana Bark in Human Keratinocytes. Antioxidants 2022, 11, 1119. https://doi.org/10.3390/antiox11061119
Piazza S, Martinelli G, Vrhovsek U, Masuero D, Fumagalli M, Magnavacca A, Pozzoli C, Canilli L, Terno M, Angarano M, et al. Anti-Inflammatory and Anti-Acne Effects of Hamamelis virginiana Bark in Human Keratinocytes. Antioxidants. 2022; 11(6):1119. https://doi.org/10.3390/antiox11061119
Chicago/Turabian StylePiazza, Stefano, Giulia Martinelli, Urska Vrhovsek, Domenico Masuero, Marco Fumagalli, Andrea Magnavacca, Carola Pozzoli, Luisa Canilli, Massimo Terno, Marco Angarano, and et al. 2022. "Anti-Inflammatory and Anti-Acne Effects of Hamamelis virginiana Bark in Human Keratinocytes" Antioxidants 11, no. 6: 1119. https://doi.org/10.3390/antiox11061119