
The Prospects of Swietenia macrophylla King in Skin Care

 ,
,  ,
,  , , and
, , and 
Abstract
:
1. Introduction
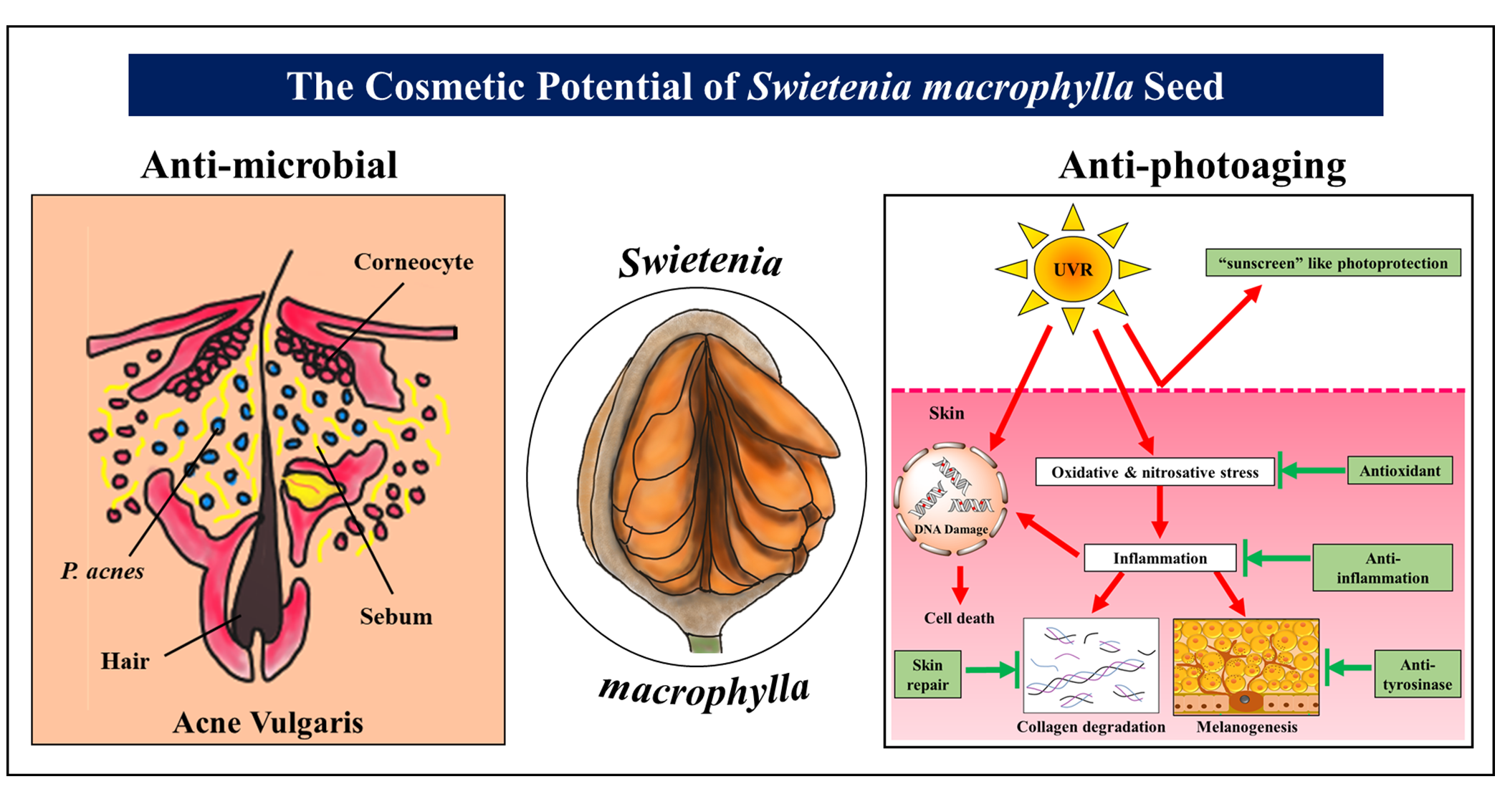
2. An Antimicrobial Remedy against Skin Disorders
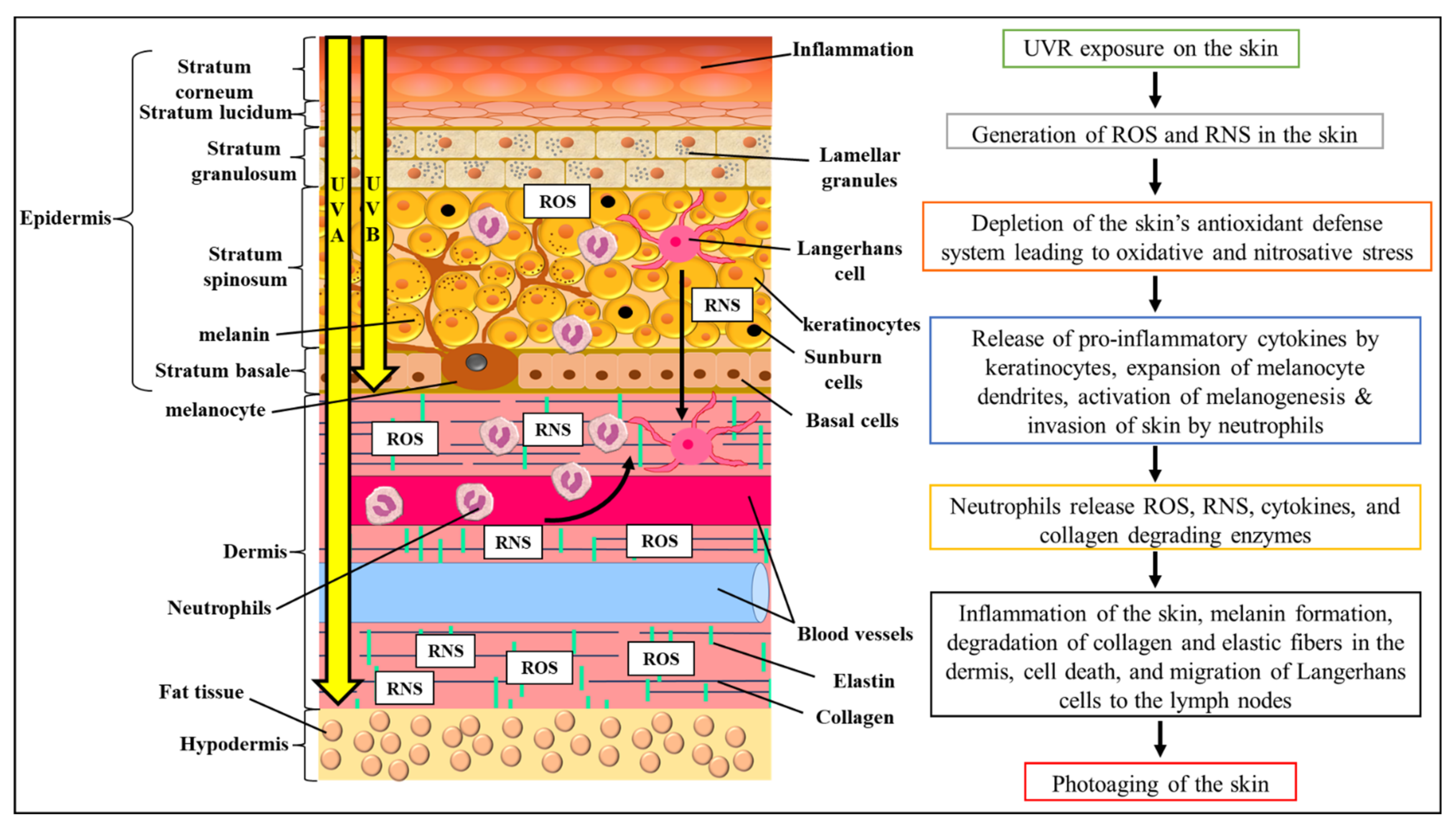
3. Photoprotective, Skin Whitening, and Skin Repair Cosmeceutical Prospects of S. macrophylla
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Haddara, M.; Hsieh, J.; Fagerstrøm, A.; Eriksson, N.; Sigurðsson, V. Exploring customer online reviews for new product development: The case of identifying reinforcers in the cosmetic industry. MDE Manag. Decis. Econ. 2019, 41, 250–273. [Google Scholar] [CrossRef]
- Feetham, H.J.; Jeong, H.S.; McKesey, J.; Wickless, H.; Jacobe, H. Skin care and cosmeceuticals: Attitudes and trends among trainees and educators. J. Cosmet. Dermatol. 2018, 17, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S. Exploratory analysis of global cosmetic industry: Major players, technology and market trends. Technovation 2005, 25, 1263–1272. [Google Scholar] [CrossRef]
- Dini, I.; Laneri, S. The new challenge of green cosmetics: Natural food ingredients for cosmetic formulations. Molecules 2021, 26, 3921. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.S.; Estanqueiro, M.; Oliveira, M.B.; Sousa Lobo, J.M. Main benefits and applicability of plant extracts in skin care products. Cosmetics 2015, 2, 48–65. [Google Scholar] [CrossRef] [Green Version]
- Ashawat, M.S.; Banchhor, M.; Saraf, S.; Saraf, S. Herbal cosmetics: Trends in skin care formulation. Pharmacogn. Rev. 2009, 3, 82–89. [Google Scholar]
- He, T.; Marco, J.; Soares, R.; Yin, Y.; Wiedenhoeft, A.C. Machine learning models with quantitative wood anatomy data can discriminate between Swietenia macrophylla and Swietenia mahagoni. Forests 2020, 11, 36. [Google Scholar] [CrossRef] [Green Version]
- Bergo, M.C.J.; Pastore, T.C.M.; Coradin, V.T.R.; Wiedenhoeft, A.C.; Braga, J.W.B. NIRS identification of Swietenia macrophylla is robust across specimens from 27 countries. IAWA J. 2016, 37, 420–430. [Google Scholar] [CrossRef] [Green Version]
- Moghadamtousi, S.Z.; Goh, B.H.; Chan, C.K.; Shabab, T.; Kadir, H.A. Biological activities and phytochemicals of Swietenia macrophylla King. Molecules 2013, 18, 10465–10483. [Google Scholar] [CrossRef]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R.; Green, E.D.; et al. Topographical and temporal diversity of the human skin microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.R.; Gómez, B.I.; McIntyre, M.K.; Dubick, M.A.; Christy, R.J.; Nicholson, S.E.; Burmeister, D.M. The cutaneous microbiome and wounds: New molecular targets to promote wound healing. Int. J. Mol. Sci. 2018, 19, 2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournière, M.; Latire, T.; Souak, D.; Feuilloley, M.G.J.; Bedoux, G. Staphylococcus epidermidis and Cutibacterium acnes: Two major sentinels of skin microbiota and the influence of cosmetics. Microorganisms 2020, 8, 1752. [Google Scholar] [CrossRef] [PubMed]
- Dréno, B. What is new in the pathophysiology of acne, an overview. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-J.; Kim, K.-P.; Choi, E.; Yim, J.-H.; Choi, C.; Yun, H.-S.; Ahn, H.-Y.; Oh, J.-Y.; Cho, Y. Effects of Lactobacillus plantarum CJLP55 on clinical improvement, skin condition and urine bacterial extracellular vesicles in patients with acne vulgaris: A Randomized, double-blind, placebo-controlled study. Nutrients 2021, 13, 1368. [Google Scholar] [CrossRef]
- Claudel, J.P.; Auffret, N.; Leccia, M.T.; Poli, F.; Corvec, S.; Dréno, B. Staphylococcus epidermidis: A potential new player in the physiopathology of acne? Dermatology 2019, 235, 287–294. [Google Scholar] [CrossRef]
- Suliman, M.B.; Nour, A.H.; Yusoff, M.; Nour, A.; Mazza, A.S. Growth inhibitory effect on bacteria of Swietenia macrophylla king seeds and leaves crude alkaloid extracts. Int. J. Pharm. Sci. Res. 2014, 5, 1000–1004. [Google Scholar] [CrossRef]
- Dharmalingam, K.; Tan, B.-K.; Mahmud, M.Z.; Sedek, S.A.M.; Majid, M.I.A.; Kuah, M.-K.; Sulaiman, S.F.; Ooi, K.L.; Khan, N.A.K.; Muhammad, T.S.T.; et al. Swietenia macrophylla extract promotes the ability of Caenorhabditis elegans to survive Pseudomonas aeruginosa infection. J. Ethnopharmacol. 2012, 139, 657–663. [Google Scholar] [CrossRef]
- Suliman, M.B.; Nour, A.; Yusoff, M.; Nour, A.; Kuppusamy, P.; Ar, Y.; Adam, M. Fatty acid composition and antibacterial activity of Swietenia macrophylla king seed oil. Afr. J. Plant Sci. 2013, 7, 300–303. [Google Scholar] [CrossRef]
- Gopalan, H.K.; Md Hanafiah, N.F.; Leong, C.R.; Tan, W.-N.; Wahidin, S.; Hway, T.S.; Yenn, T.W. Chemical composition and antimicrobial efficiency of Swietenia macrophylla seed extract on clinical wound pathogens. Nat. Prod. Sci. 2019, 25, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Durai, M.V.; Balamuniappan, G.; Geetha, S. Phytochemical screening and antimicrobial activity of leaf, seed and central-fruit-axis crude extract of Swietenia macrophylla King. J. Pharmacogn. Phytochem. 2016, 5, 181–186. [Google Scholar]
- Kong, C.; Yehye, W.A.; Abd Rahman, N.; Tan, M.W.; Nathan, S. Discovery of potential anti-infectives against Staphylococcus aureus using a Caenorhabditis elegans infection model. BMC Complement. Altern. Med. 2014, 14, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, A.K.M.S.; Chowdhury, A.K.A.; Ali, H.-A.; Raihan, S.Z.; Ali, M.S.; Nahar, L.; Sarker, S.D. Antibacterial activity of two limonoids from Swietenia mahagoni against multiple-drug-resistant (MDR) bacterial strains. J. Nat. Med. 2009, 63, 41–45. [Google Scholar] [CrossRef]
- Kader, M.A.; Haque, M.E.; Khondkar, P.; Islam, M.M.; Rahman, M.M. Antibacterial and Cytotoxic Limonoids from the Seeds of Swietenia mahagony. Dhaka Univ. J. Pharm. Sci. 2010, 8, 141–145. [Google Scholar] [CrossRef] [Green Version]
- Mootoo, B.S.; Ali, A.; Motilal, R.; Pingal, R.; Ramlal, A.; Khan, A.; Reynolds, W.F.; McLean, S. Limonoids from Swietenia macrophylla and S. aubrevilleana. J. Nat. Prod. 1999, 62, 1514–1517. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, T.S.R.; Passos, M.d.S.; Calixto, S.D.; Ventura, T.L.B.; Lassounskaia, E.; Muzitano, M.F.; Braz-Filho, R.; Vieira, I.J.C. Anti-Mycobacterium tuberculosis Activity of Compounds from Cedrela fissilis Vell Seeds (Meliaceae). Rev. Virtual De Quim. 2021, 13, 1116–1121. [Google Scholar] [CrossRef]
- Suliman, M.B. Preliminary phytochemical screening and thin layer chromatography analysis of Swietenia macrophylla King methanol extracts. Chem. Adv. Mater. 2018, 3, 1–7. [Google Scholar]
- García-Bores, A.M.; Espinosa-González, A.M.; Reyna-Campos, A.; Cruz-Toscano, S.; Benítez-Flores, J.C.; Hernández-Delgado, C.T.; Flores-Maya, S.; Urzúa-Meza, M.; Peñalosa-Castro, I.; Céspedes-Acuña, C.L.; et al. Lippia graveolens photochemopreventive effect against UVB radiation-induced skin carcinogenesis. J. Photochem. Photobiol. B 2017, 167, 72–81. [Google Scholar] [CrossRef]
- Herrling, T.; Jung, K.; Fuchs, J. Measurements of UV-generated free radicals/reactive oxygen species (ROS) in skin. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2006, 63, 840–845. [Google Scholar] [CrossRef]
- Terra, V.A.; Souza-Neto, F.P.; Pereira, R.C.; Silva, T.N.X.; Costa, A.C.C.; Luiz, R.C.; Cecchini, R.; Cecchini, A.L. Time-dependent reactive species formation and oxidative stress damage in the skin after UVB irradiation. J. Photochem. Photobiol. B 2012, 109, 34–41. [Google Scholar] [CrossRef]
- Mahendra, C.K.; Ser, H.-L.; Pusparajah, P.; Htar, T.T.; Chuah, L.-H.; Yap, W.H.; Tang, Y.-Q.; Zengin, G.; Tang, S.Y.; Lee, W.L.; et al. Cosmeceutical therapy: Engaging the repercussions of UVR photoaging on the skin’s circadian rhythm. Int. J. Mol. Sci. 2022, 23, 2884. [Google Scholar] [CrossRef]
- Pillai, S.; Oresajo, C.; Hayward, J. Ultraviolet radiation and skin aging: Roles of reactive oxygen species, inflammation and protease activation, and strategies for prevention of inflammation-induced matrix degradation—A review. Int. J. Cosmet. Sci. 2005, 27, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Muthusamy, V.; Piva, T.J. The UV response of the skin: A review of the MAPK, NFκB and TNFα signal transduction pathways. Arch. Dermatol. Res. 2009, 302, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.L.; van Weelden, H.; Bruijnzeel, P.L.B. Neutrophil Infiltration in normal human skin after exposure to different ultraviolet radiation sources. Photochem. Photobiol. 2008, 84, 1528–1534. [Google Scholar] [CrossRef]
- Takeuchi, H.; Gomi, T.; Shishido, M.; Watanabe, H.; Suenobu, N. Neutrophil elastase contributes to extracellular matrix damage induced by chronic low-dose UV irradiation in a hairless mouse photoaging model. J. Dermatol. Sci. 2010, 60, 151–158. [Google Scholar] [CrossRef]
- Rijken, F.; Bruijnzeel-Koomen, C.A.F.M. Photoaged skin: The role of neutrophils, preventive measures, and potential pharmacological targets. Clin. Pharmacol. Ther. 2011, 89, 120–124. [Google Scholar] [CrossRef]
- Tran, T.-N.T.; Schulman, J.; Fisher, D.E. UV and pigmentation: Molecular mechanisms and social controversies. Pigment Cell Melanoma Res. 2008, 21, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Hamakawa, M.; Sugihara, A.; Okamoto, H.; Horio, T. Ultraviolet B radiation suppresses Langerhans cell migration in the dermis by down-regulation of α4 integrin. Photodermatol. Photoimmunol. Photomed. 2006, 22, 116–123. [Google Scholar] [CrossRef]
- Wong, W.C.; Wu, J.Y.; Benzie, I.F.F. Photoprotective potential of Cordyceps polysaccharides against ultraviolet B radiation-induced DNA damage to human skin cells. Br. J. Dermatol. 2011, 164, 980–986. [Google Scholar] [CrossRef]
- Toriyama, E.; Masuda, H.; Torii, K.; Ikumi, K.; Morita, A. Time kinetics of cyclobutane pyrimidine dimer formation by narrowband and broadband UVB irradiation. J. Dermatol. Sci. 2021, 103, 151–155. [Google Scholar] [CrossRef]
- Mahendra, C.K.; Tan, L.T.-H.; Yap, W.H.; Chan, C.K.; Pusparajah, P.; Goh, B.H. An optimized cosmetic screening assay for ultraviolet B (UVB) protective property of natural products. Prog. Drug Discov. Biomed. Sci. 2019, 2, 1–6. [Google Scholar] [CrossRef]
- Smijs, T.G.; Pavel, S. Titanium dioxide and zinc oxide nanoparticles in sunscreens: Focus on their safety and effectiveness. Nanotechnol. Sci. Appl. 2011, 4, 95–112. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.-J.; Huang, S.-C.; Chen, Y.-P.; Chiueh, L.-C.; Shih, D.Y.-C. Analysis of titanium dioxide and zinc oxide nanoparticles in cosmetics. J. Food Drug Anal. 2015, 23, 587–594. [Google Scholar] [CrossRef] [Green Version]
- Scheuer, E.; Warshaw, E. Sunscreen allergy: A review of epidemiology, clinical characteristics, and responsible allergens. Dermatitis 2006, 17, 3–11. [Google Scholar] [CrossRef]
- Lee Granger, K.; Brown, P.R. The chemistry and HPLC analysis of chemical sunscreen filters in sunscreens and cosmetics. J. Liq. Chromatogr. Relat. Technol. 2001, 24, 2895–2924. [Google Scholar] [CrossRef]
- ISO. ISO 24444:2010 Cosmetics—Sun Protection Test Methods—In vivo Determination of the Sun Protection Factor (SPF). Available online: https://www.iso.org/obp/ui/#iso:std:iso:24444:ed-1:v1:en (accessed on 3 April 2022).
- FDA. Guidance for Industry Labelling and Effectiveness Testing: Sunscreen Drug Products for Over-the Counter Human Use—Small Entity Compliance Guide. Available online: https://www.fda.gov/drugs/guidancecomplianceregulatoryinformation/guidances/ucm330694.htm (accessed on 3 April 2022).
- Mahendra, C.K.; Mahendra, C.K.; Pusparajah, P.; Htar, T.-T.; Chuah, L.-H.; Duangjai, A.; Khan, T.M.; Yow, Y.Y.; Kumari, Y.; Goh, B.H. Simplified, cost effective, and accurate calculation of critical wavelength via the MATLAB software. Prog. Drug Discov. Biomed. Sci. 2021, 4, 1–16. [Google Scholar] [CrossRef]
- Osterwalder, U.; Herzog, B. Sun protection factors: World wide confusion. Br. J. Dermatol. 2009, 161, 13–24. [Google Scholar] [CrossRef]
- Bernstein, E.F.; Sarkas, H.W.; Boland, P.; Bouche, D. Beyond sun protection factor: An approach to environmental protection with novel mineral coatings in a vehicle containing a blend of skincare ingredients. J. Cosmet. Dermatol. 2020, 19, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Pandel, R.; Poljšak, B.; Godic, A.; Dahmane, R. Skin photoaging and the role of antioxidants in its prevention. ISRN Dermatol. 2013, 2013, 930164. [Google Scholar] [CrossRef]
- Cui, B.; Wang, Y.; Jin, J.; Yang, Z.; Guo, R.; Li, X.; Yang, L.; Li, Z. Resveratrol treats UVB-induced photoaging by anti-MMP expression, through anti-inflammatory, antioxidant, and antiapoptotic properties, and treats photoaging by upregulating VEGF-B expression. Oxid. Med. Cell Longev. 2022, 2022, 6037303. [Google Scholar] [CrossRef]
- Draelos, Z.D. Novel approach to the treatment of hyperpigmented photodamaged skin: 4% hydroquinone/0.3% retinol versus tretinoin 0.05% emollient cream. Dermatol. Surg. 2005, 31, 799–805. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Y.; Song, H.; He, J.; Li, G.; Zheng, Y.; Li, B. Collagen peptides promote photoaging skin cell repair by activating the TGF-β/Smad pathway and depressing collagen degradation. Food Funct. 2019, 10, 6121–6134. [Google Scholar] [CrossRef]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef]
- Levy, L.L.; Emer, J.J. Emotional benefit of cosmetic camouflage in the treatment of facial skin conditions: Personal experience and review. Clin. Cosmet. Investig. Dermatol. 2012, 5, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Mahendra, C.K.; Tan, L.T.H.; Mahendra, C.K.; Ser, H.-L.; Pusparajah, P.; Htar, T.T.; Chuah, L.-H.; Yap, W.H.; Tang, S.Y.; Ming, L.C.; et al. The potential of sky fruit as an anti-aging and wound healing cosmeceutical agent. Cosmetics 2021, 8, 79. [Google Scholar] [CrossRef]
- Mahendra, C.K.; Abidin, S.A.Z.; Htar, T.T.; Chuah, L.H.; Khan, S.U.; Ming, L.C.; Tang, S.Y.; Pusparajah, P.; Goh, B.H. Counteracting the ramifications of UVB irradiation and photoaging with Swietenia macrophylla king seed. Molecules 2021, 26, 2000. [Google Scholar] [CrossRef]
- Dutta, M.; Biswas, U.K.; Chakraborty, R.; Banerjee, P.; Maji, D.; Mondal, M.C.; Raychaudhuri, U. Antidiabetic and antioxidant effect of Swietenia macrophylla seeds in experimental type 2 diabetic rats. Int. J. Diabetes Dev. Ctries. 2013, 33, 60–65. [Google Scholar] [CrossRef]
- Wan Hassan, W.N.A.; Zulkifli, R.; Farediah, A.; Yunus, M. Antioxidant and tyrosinase inhibition activities of Eurycoma longifolia and Swietenia macrophylla. J. Appl. Pharm. Sci. 2015, 5, 006–010. [Google Scholar] [CrossRef] [Green Version]
- Coello, F.P.; Azuaje, D.R.; Catari, I.P.; Marrero, M.P.; Vargas, C.O. Evaluation of the antioxidant activity of aqueous extracts of leaves and seeds of Swietenia macrophylla King by chemical and biological methods. J. Drug Res. Dev. 2020, 6, 1–4. [Google Scholar] [CrossRef]
- Mak, K.-K.; Shiming, Z.; Balijepalli, M.K.; Dinkova-Kostova, A.T.; Epemolu, O.; Mohd, Z.; Pichika, M.R. Studies on the mechanism of anti-inflammatory action of swietenine, a tetranortriterpenoid isolated from Swietenia macrophylla seeds. Phytomed. Plus 2021, 1, 100018. [Google Scholar] [CrossRef]
- Chen, L.-C.; Liao, H.-R.; Chen, P.-Y.; Kuo, W.-L.; Chang, T.-H.; Sung, P.-J.; Wen, Z.-H.; Chen, J.-J. Limonoids from the seeds of Swietenia macrophylla and their anti-inflammatory activities. Molecules 2015, 20, 18551–18564. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-J.; Huang, S.-S.; Liao, C.-H.; Wei, D.-C.; Sung, P.-J.; Wang, T.-C.; Cheng, M.-J. A new phragmalin-type limonoid and anti-inflammatory constituents from the fruits of Swietenia macrophylla. Food Chem. 2010, 120, 379–384. [Google Scholar] [CrossRef]
- Ma, Y.-Q.; Jiang, K.; Deng, Y.; Guo, L.; Wan, Y.-Q.; Tan, C.-H. Mexicanolide-type limonoids from the seeds of Swietenia macrophylla. J. Asian Nat. Prod. Res. 2018, 20, 299–305. [Google Scholar] [CrossRef]
- Eid, A.M.; El Enshasy, H.A.; Aziz, R.; Elmarzugi, N.A. Preparation, characterization and anti-inflammatory activity of Swietenia macrophylla nanoemulgel. J. Nanomed. Nanotechnol. 2014, 5, 1–10. [Google Scholar] [CrossRef]
- Eid, A.M.; El-Enshasy, H.A.; Aziz, R.; Elmarzugi, N.A. The preparation and evaluation of self-nanoemulsifying systems containing Swietenia oil and an examination of its anti-inflammatory effects. Int. J. Nanomed. 2014, 9, 4685–4695. [Google Scholar] [CrossRef] [Green Version]
- Dolinska, M.B.; Wingfield, P.T.; Young, K.L.; Sergeev, Y.V. The TYRP1-mediated protection of human tyrosinase activity does not involve stable interactions of tyrosinase domains. Pigment Cell Melanoma Res. 2019, 32, 753–765. [Google Scholar] [CrossRef] [PubMed]
- Nilugal, K.C.; Fattepur, S.; Asmani, F.M.; Abdullah, I.; Vijendren, S.; Ugandar, R.E. Evaluation of wound healing activity of Swietenia macrophylla (Meliaceae) seed extract in albino rats. Am. J. PharmTech Res. 2017, 7, 113–124. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Seed Extract | Extraction Method | Antimicrobial Strains | Antimicrobial Activity | References |
|---|---|---|---|---|
| Alkaloid crude extract Rate of alkaloid extracted from the seeds: 2.85% | (A) The seeds were cleaned, sun-dried and grinded to fine powder. (B) Ethanol (300 mL; 70%) was mixed with 20 g powder of the seeds in an orbital shaker at 150 rpm for 24 h at 25 °C. (C) The extract was then evaporated with a rotary evaporator until one fifth of the initial volume was reached, before adding 20 mL of 0.1 N hydrochloric acid. (D) The extract was filtered and extracted twice with 20 mL of chloroform. (E) The extract was treated twice with 10 mL of 0.1 N hydrochloric acid and 0.1 N ammonia was added to obtain pH 9, before adding 30 mL of chloroform. (F) The operation was repeated three times, before evaporating and dissolving the residue in 20 mL of 0.02 N hydrochloric acid. (G) The extract was titrated with 0.02 N potassium hydroxide, with methyl red as an indicator. | S. aureus ATCC1026, E. coli ATCC10536, P. aeruginosa ATCC15442. | (A) Inhibition activity (disk diffusion method): (i) S. aureus: 17 mm (50 mg/mL); 21 mm (100 mg/mL); (ii) E. coli: 10 mm (50 mg/mL); 12 mm (100 mg/mL); (iii) P. aeruginosa: 12 mm (50 mg/mL); 15 mm (100 mg/mL). (B) MIC and MBC value: (i) S. aureus: not available (MIC); 12.5 mg/mL (MBC); (ii) E. coli: 25 mg/mL (MIC); 50 mg/mL (MBC); (iii) P. aeruginosa: 25 mg/mL (MIC); 50 mg/mL (MBC). | [16] |
| Methanol, ethyl acetate and butanol crude extract | The sample was air-dried, powdered and extracted with methanol, ethyl acetate and butanol. | C. elegans infected with MRSA ATCC33591 or methicillin-sensitive S. aureus (MSSA) NCTC83254. | (A) Survival of 72 h S. aureus infected C. elegans after treatment (200 μg/mL): (i) Methanol: 70% survival (3-fold change vs. untreated); (ii) Ethyl acetate: 65% (2.8-fold change vs. untreated); (iii) Butanol: 96.7% (4.2-fold change vs. untreated). (B) None of the extracts had bacteriostatic or bactericidal activity on S. aureus growth in vitro. (C) All three extracts significantly increased the survival of infected C. elegans but did not affect replication of S. aureus. There was no difference in CFU of the intestinal bacterial loads. (D) All three extracts promoted survival in 72 h MRSA infected C. elegans (>70% survival). | [21] |
| Methanol extract and ethyl acetate extract | (A) The sample was air-dried, powdered and extracted with methanol. (B) The crude extract was “defattened” using hexane and then further extracted with ethyl acetate. | C. elegans infected with P. aeruginosa (Bacteria strain was not specified). | (A) Ethyl acetate extract (200 and 400 μg/mL) and methanol extract (200 μg/mL) significantly improved the survival of C. elegans 48 h after P. aeruginosa infection. (B) Treatment with 200 μg/mL of ethyl acetate extract had the highest survival of 59.5 ± 1.65%. (C) MIC assay demonstrated that the ethyl acetate extract (25–1000 μg/mL) had no inhibition against P. aeruginosa. (D) There was no significant difference in CFU of P. aeruginosa in C. elegans intestines after both extract treatment. © Ethyl acetate extract (200 μg/mL) increased expression of lys-7 gene that was suppressed by P. aeruginosa infection. | [17] |
| Seed oil extract by diethyl ether (39%) and n-hexane (42.7%) | (A) The seeds were dried and ground to small pieces. (B) The seeds (10 g) were extracted using n-hexane and diethyl ether to recover at least 10 mL of oil using the Soxhlet apparatus. (C) The solvent was evaporated using the rotary evaporator, before further drying under the open air. | S. aureus, E. coli, P. aeruginosa, Salmonella Typhimurium. (All bacteria are in-house bacteria strains). | (A) Antibacterial activity of seed oil (disk diffusion method): (i) 10 μg/mL: 5 mm (S. aureus); 4 mm (Salmonella Typhimurium); 5 mm (P. aeruginosa); (ii) 20 μg/mL: 8 mm (S. aureus); 6 mm (Salmonella Typhimurium); 5 mm (P. aeruginosa); (iii) 50 μg/mL: 8 mm (S. aureus); 9 mm (Salmonella Typhimurium); 6 mm (P. aeruginosa); (iv) 100 μg/mL: 9 mm (S. aureus); 10 mm (Salmonella Typhimurium); 6 mm (P. aeruginosa); (v) 1000 μg/mL: 11 mm (S. aureus); 20 mm (Salmonella Typhimurium); 11 mm (P. aeruginosa); (vi) There was no effect on E. coli after the seed oil treatment. | [18] |
| Methanol extract | (A) The seeds were washed and dried at 50 °C for 3 days, before grinding to a coarse powder. (B) The powder was then soaked in methanol at a ratio of 1:5 (w/v) for 24 h. (C) The extract was filtered and concentrated using the rotary evaporator at 60 °C. | Gram-positive: S. aureus, B. cereus, B. subtilis, MRSA Gram-negative: Proteus mirabilis, Yersinia sp., E. coli, Klebsiella pneumoniae, S. boydii, A. anitratus Fungal: Aspergillus niger, Microsporum fulvum, Rhizopus sp. Yeast: Candida utilis. (Microbial strains are clinical isolates). | (A) Antimicrobial activity of methanol extract (disk diffusion method): (i) S. aureus: 13.0 ± 0.6 mm; (ii) B. cereus: 15.0 ± 0.6 mm; (iii) B. subtilis: 15.0 ± 1.5 mm; (iv) S. boydii: 15.0 ± 1.0 mm; (v) A. anitratus: 16.0 ± 1.2 mm; (vi) C. utilis: 10.0 ± 1.2 mm; (vii) No inhibitory activity was observed for MRSA, P. mirabilis, Yersinia sp., E. coli, K. pneumoniae, A. niger, M. fulvum, or Rhizopus sp. (B) MIC and MBC of methanol extract: (i) B. cereus: 3.13 mg/mL (MIC); 6.25 mg/mL (MBC); (ii) S. aureus: 6.25 mg/mL (MIC); 25 mg/mL(MBC); (iii) B. subtilis: 1.56 mg/mL (MIC); 3.13 mg/mL (MBC); (iv) S. boydii: 12.5 mg/mL (MIC); 25 mg/mL (MBC); (v) A. anitratus: 12.5 mg/mL (MIC); 25 mg/mL (MBC); (vi) C. utilis: 12.5 mg/mL (MIC); 25 mg/mL (MBC). | [19] |
| Methanol extract | (A) The seeds were dried for 2 days and pulverized to powder form. (B) The powdered seeds (30 g) were then extracted by reflux using Soxhlet apparatus for 10 h with successive organic solvent (80% methanol) and concentrated through oven drying. (C) Each fraction was collected, distilled and dried in the incubator. | Gram-positive: S. aureus Gram-negative: E. coli Fungi: Fusarium sp, Helminthosporium sp, Alternaria sp. (Bacteria and fungi are lab strains). | (A) Agar diffusion method (inhibition zone): (i) S. aureus:
(i) Fusarium sp.:
| [20] |
| Seed Extract | Bioproperties | Extraction Method | In Vitro/In Vivo Studies | Bioactivity | References |
|---|---|---|---|---|---|
| Aqueous extract | Antioxidant | (A) The seeds were washed, dried at room temperature, powdered, and sieved through 40 meshes. (B) For 200 mg of powder, 1 mL of distilled water was added, before centrifuging for 15 min at 3000 rpm. (C) The supernatant was filtered and collected as the extract. | (A) Winstar rats (200–250 g) injected intraperitoneally with 65 mg/kg of streptozotocin in a volume of 1 mL/kg body weight to induce diabetes. The rats were fed with glucose 30 min prior to treatment with the extract. Blood was drawn from the tail. (B) Antioxidant testing on the extract itself. | (A) Antioxidant level in the blood demonstrated dose-dependent increase in antioxidant activity, using modified free oxygen radical defense (FORD) assay. (B) Antioxidant activity of pure aqueous extract also showed increasing antioxidant activity dose-dependently via modified FORD assay. | [58] |
| Aqueous extract | Antioxidant | (A) The seeds were washed, dried at room temperature, powdered, and sieved. (B) The powdered seeds were added to 200 mL of boiled distilled water, before centrifuging at 3500 rpm for 8 min and filtered. | (A) FORD assay. (B) Free radical 2,2-diphenyl-1-picrylhydrazyl (DPPH) + H2 assay. (C) Ferric-reducing antioxidant power (FRAP) assay. (D) Oxidative stress test on Saccharomyces cerevisiae (S. cerevisiae). | (A) FORD assay: increase in antioxidant activity over time. (B) DPPH assay: 56.2 ± 0.97%. (C) FRAP assay: 34.8 ± 0.13 μmol Fe + 2/g PM. (D) Oxidative stress test: I extract promotes the growth of S. cerevisiae over time, even in the presence of hydrogen peroxide that causes oxidative stress. | [60] |
| Ethyl acetate extract; methanol extract | Antioxidant and anti-pigmentation | (A) The seeds were dried and then ground to powder, before being subjected to extraction. (B) The powdered seeds were extracted with ethyl acetate and methanol in increasing order of their polarity. (C) Each extract was concentrated using a rotary evaporator at 40–50 °C. | (A) DPPH assay. (B) Tyrosinase inhibitory activity. | (A) DPPH radical scavenging capacity (500 μg/mL): (i) Ethyl acetate extract: 30.30 ± 1.63%; IC50 detected; (ii) Methanol extract: 56.82 ± 2.67%; IC50: 200 μg/mL. (B) Percentage of inhibition on tyrosinase activity: (i) Ethyl acetate extract: 14.44 ± 2.45%; (ii) Methanol extract: 15.95 ± 1.27%. | [59] |
| Ethanol extract (SMCE); hexane fraction (SMHF); ethyl acetate fraction (SMEAF); aqueous fraction (SMWF) | Photoprotection, antioxidant, wound healing | (A) The seeds (3 kg) were finely grounded and soaked in ethanol for 72 h at room temperature. (B) The extract was filtered and concentrated with a rotary evaporator at 40 °C to obtain SMCE. (C) SMCE was then dissolved in hexane to obtain the hexane fraction. The supernatant was dried with anhydrous sodium sulphate, before concentrating with a rotary vacuum evaporator to obtain SMHF. (D) The insoluble residues of hexane were subjected to ethyl acetate and water portioning in a 1:1 ratio. (E) The ethyl acetate fraction was dried via rotary evaporation to obtain SMEAF, while the water fraction was freeze-dried to obtain SMWF. | (A) DPPH radical scavenging assay. (B) 2′azino-bis (3, -ethylbenzothaizoline-6-sulfonic acid). (ABTS) radical scavenging assay. (C) Ferrous ion chelating assay. (D) Critical wavelength measurement. (E) Scratch wound assay on HaCaT cells. | (A) Antioxidant assays: (i) SMCE: not significant for DPPH, ABTS and ion chelating activity; (ii) SMHF: significant only for iron chelating activity at 2000 μg/mL with 14.073 ± 0.18% activity. Not significant for DPPH and ABTS assays; (iii) SMEAF: not significant for DPPH, ABTS and ion chelating activity; (iv) SMWF: (a) Significant for DPPH activity: 6.332 ± 0.80% at 2000 μg/mL; (b) Significant and dose-dependent increase for ABTS activity: 12.796 ± 2.01% at 125 μg/mL; (c) Significant and dose-dependent increase for iron chelating activity: 8.014 ± 2.51% at 125 μg/mL. (B) Critical wavelength of SMCE, SMHF, SMEAF, and SMWF are 347.6, 345, 341.6, and 362.4 nm, respectively. (C) Percentage of wound closure after 24 h of treatment: (i) SMCE (6.25 μg/mL): 54.10 ± 2.59%; (ii) SMHF (100 μg/mL): 59.45 ± 5.72%; (iii) SMEAF (12.5 μg/mL): 41.48 ± 3.91%; (iv) SMWF (50 μg/mL): 74.68 ± 5.16%. | [56] |
| SMCE, SMHF, SMEAF, SMWF | Photoprotection against UVB irradiation | (A) The seeds (3 kg) were finely ground and soaked in ethanol for 72 h at room temperature. (B) The extract was filtered and concentrated with a rotary evaporator at 40 °C to obtain SMCE. (C) SMCE was then dissolved in hexane to obtain the hexane fraction. The supernatant was dried with anhydrous sodium sulphate, before concentrating with a rotary vacuum evaporator to obtain SMHF. (D) The insoluble residues of hexane were subjected to ethyl acetate and water portioning in a 1:1 ratio. (E) The ethyl acetate fraction was dried via rotary evaporation to obtain SMEAF, while the water fraction was freeze-dried to obtain SMWF. | HaCaT cells treated with SMCE (6.25 μg/mL), SMHF (100 μg/mL), SMEAF (12.5 μg/mL), and SMWF (50 μg/mL) in PBS, while being exposed to 50 mJ/cm2 UVB. Cells were then rinsed and incubated for 24 h at 37 °C, 5% CO2. Protein and gene expression changes were taken 24 h post exposure. Negative control: non-irradiated cells. Inducer control: irradiated but non-treated cells. | Comparison of treatment with negative and inducer controls: (A) SMCE: (i) Gene expression changes: downregulation of TNF-α and MMP-1 (vs inducer control); (ii) Protein expression changes: downregulation of ribosomes and Filamin Bβ (vs negative control). (B) SMHF: (i) Gene expression changes: downregulation of NF-κB and cyclin D1 (vs inducer control); (ii) Protein expression changes: multiple changes across the redox system, RNA to protein processing, DNA maintenance and repair, glycolysis process, and cell growth, proliferation and migration. All changes demonstrated reversal against UVB induced damage. (C) SMEAF: (i) Gene expression changes: downregulation of TNF-α, NF-κB, MMP-1 and Bax; (ii) Protein expression changes: downregulation of PRDX-3, PDI-A3, and fascin (vs negative control). (D) SMWF: (i) Gene expression changes: no significant changes in TNF-α, NF-κB, COX-2, MMP-1, cyclin D1 and Bax; (ii) Protein expression changes: multiple changes across the redox system, RNA to protein processing, DNA maintenance and repair, glycolysis process, and cell growth, proliferation and migration, in which majority are opposite to SMHF. | [57] |
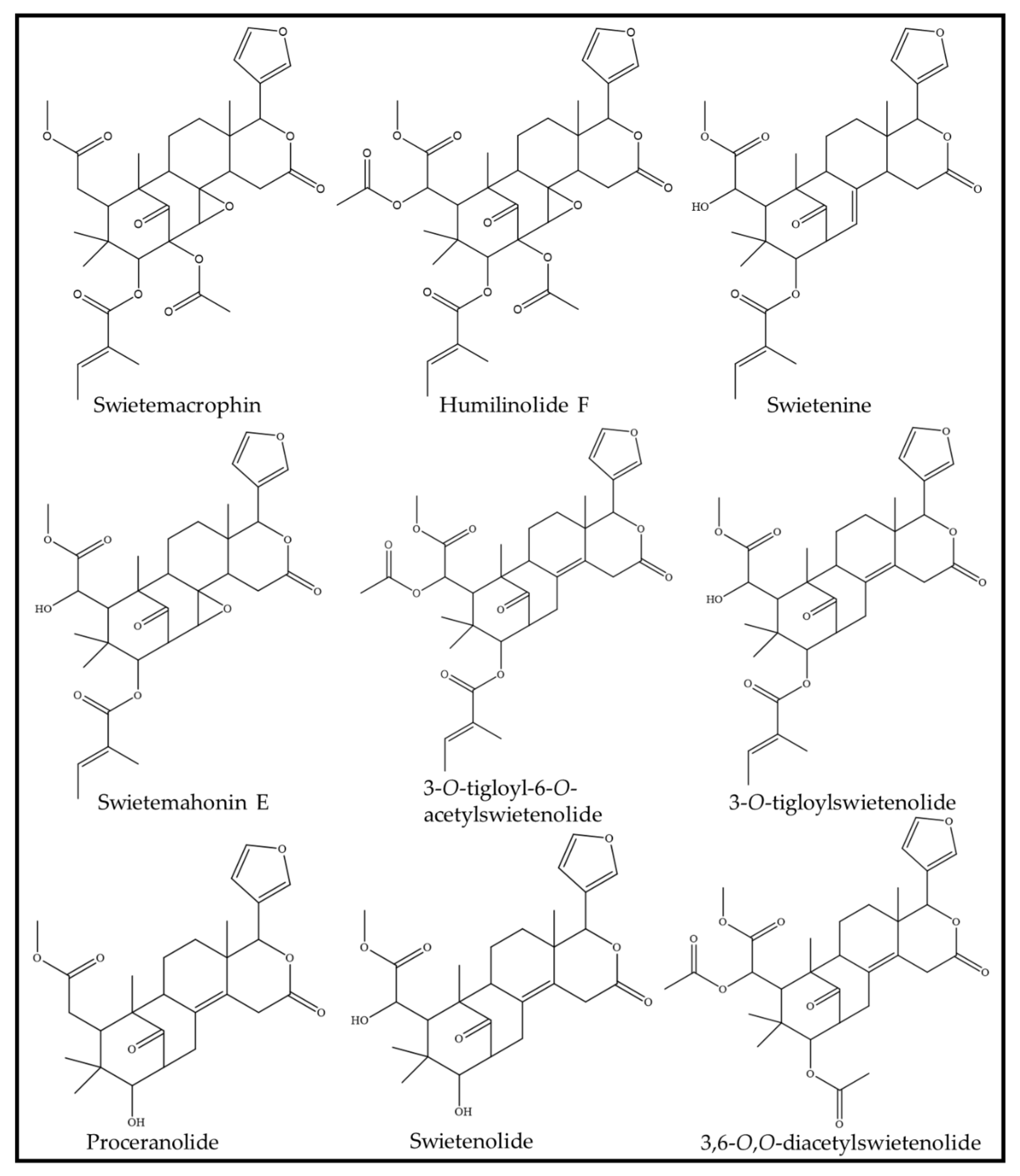
| Pure compounds (i) swietemacrophin (ii) humilinolide F (iii) 3,6-O,O-diacetylswietenolide (iv) 3-O-tigloylswietenolide (v) swietemahonin E; (vi) swietenine | Anti-inflammation | (A) Dried seeds (380 g) were pulverized and extracted with methanol for 3 days at room temperature. (B) The extract was concentrated at 35 °C with reduced pressure, before being partitioned between ethyl acetate and water in a 1:1 ratio. (C) The water fraction was further extracted with n-butanol to produce a butanol soluble fraction and water fraction. (D) The ethyl acetate fraction was further fractionated and purified to produce six pure compounds. | (A) Human neutrophils obtained from the venous blood of healthy, adult volunteers aged 20–30 years old. (B) RAW264.7 (murine macrophage) cells. | (A) Suppression of superoxide anion generation by human neutrophils (IC50): (i) Swietemacrophin: 45.44 ± 3.76 μM (p < 0.05); (ii) Humilinolide F: 27.13 ± 1.82 μM (p < 0.01); (iii) 3,6-O,O-diacetylswietenolide: 29.36 ± 1.75 μM (p < 0.05); (iv) 3-O-tigloylswietenolide: 35.58 ± 2.12 μM; (v) Swietemahonin E: 33.64 ± 2.05 μM (p < 0.05); (vi) Swietenine: >100. (B) Inhibition of NO generation by RAW264.7 cells: (i) Swietemacrophin: 33.45 ± 1.88 μM (p < 0.01); (ii) Humilinolide F: 49.36 ± 4.01 μM; (iii) 3,6-O,O-diacetylswietenolide: 64.21 ± 5.67 μM; (iv) 3-O-tigloylswietenolide: 32.62 ± 3.27μM (p < 0.01); (v) Swietemahonin E: 29.70 ± 2.11μM (p < 0.05); (vi) Swietenine: 36.32 ± 2.84. | [62] |
| Swietenine | Antioxidant and anti-inflammation | (A) The seeds were dried for 24 h in a drying oven at 30 °C. (B) Oil from the seeds were removed using an oil press machine. (C) The pressed seed was then sequentially extracted with hexane, ethyl acetate and methanol via a Soxhlet extractor. (D) The ethyl acetate extract was concentrated with a rotary evaporator and dried in a vacuum dryer, before further purification to isolate swietenine. | (A) RAW264.7 cells induced by lipopolysaccharide. (B) Hepa1c1c7 (murine hepatoma) cells. | (A) Swietenine dose-dependently inhibited NO production in induced RAW264.7 cells with 65.97 ± 0.7% at 0.78 μM and 21.03 ± 1.4% at 25 μM. (B) Swietenine dose-dependently significantly inhibited production of pro-inflammatory cytokine IL-1β, IFN-γ, TNF-α, and IL-6 in induced RAW264.7 cells. At 25 μM, RAW264.7 experienced a reduction in fold change in IL-1β by 1.3 ± 0.13, IFN-γ by 3.40 ± 0.07, TNF-α by 1.45 ± 0.06, and IL-6 by 1.60 ± 0.20. (C) Swietenine dose-dependently inhibited the expression of COX-2 and NF-κB of induced RAW264.7 cells. At 25μM, RAW264.7 experienced a reduction in fold change in COX-2 by 1.73 ± 0.06 and NF-κB by 2.90 ± 0.09. (D) Swietenine dose-dependently upregulated NRF2 and HO-1 in induced RAW264.7 cells. At 25μM, RAW264.7 experienced an increase in fold change in NRF2 by 2.57 ± 0.02 μM and HO-1 by 2.46 ± 0.03. (E) Swietenine induced NQO1 activity in Hepa1c1c7 cells. The CD value of swietenine was 15.8 ± 0.23 μM. | [61] |
| (A) Swietenia oil extracted from Swietenia macrophylla (B) Nanoemulsion Swietenia oil (C) Nanoemulgel of Swietenia oil | Anti-inflammation | Not available | Male Sprague–Dawley rats (180–200 g) with induced edema in the right hind paw. The rats were treated with the Swietenia oil, before being induced to have an edema. | (A) Significant dose-dependent inhibition of inflammation in the paw of the rats across 4 h of Swietenia oil treatment. (B) Nanoemulsion of Swietenia oil improved the percentage of inflammation inhibition from 54% to 76.4%, at 4 h of 4 mg/kg treatment. (C) Nanoemulgel of Swietenia oil improved the percentage of inflammation inhibition from 27% to 69.6%, at 4 h of 20% concentration treatment. | [65] |
| Ethanol extract in the form of ointment (10% w/w) | Wound healing | (A) The seeds were dried and homogenized before extracting with 95% ethanol at room temperature for 6 days. (B) The extract was filtered and concentrated. | Adult male Sprague–Dawley albino rats (200–250 g) were excised on the shaved dorsal thoracic region. The wound size was 200 mm and 2 mm deep. The wound was then blotted with a cotton swab soaked in normal saline to achieve hemostasis, before leaving it open. | (A) The wound area was closed by the ethanolic ointment by day 15 as compared to the control, which took 21 days. (B) The ethanolic ointment demonstrated higher fibroblast proliferation and increased formation of blood capillaries. There was also presence of collagen fibers and collagen deposition. As compared to the control, the control sample had disorganized fibroblasts, fewer blood capillaries and reduced collagen deposition. | [68] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahendra, C.K.; Goh, K.W.; Ming, L.C.; Zengin, G.; Low, L.E.; Ser, H.-L.; Goh, B.H. The Prospects of Swietenia macrophylla King in Skin Care. Antioxidants 2022, 11, 913. https://doi.org/10.3390/antiox11050913
Mahendra CK, Goh KW, Ming LC, Zengin G, Low LE, Ser H-L, Goh BH. The Prospects of Swietenia macrophylla King in Skin Care. Antioxidants. 2022; 11(5):913. https://doi.org/10.3390/antiox11050913
Chicago/Turabian StyleMahendra, Camille Keisha, Khang Wen Goh, Long Chiau Ming, Gokhan Zengin, Liang Ee Low, Hooi-Leng Ser, and Bey Hing Goh. 2022. "The Prospects of Swietenia macrophylla King in Skin Care" Antioxidants 11, no. 5: 913. https://doi.org/10.3390/antiox11050913