
The ArsH Protein Product of the Paracoccus denitrificans ars Operon Has an Activity of Organoarsenic Reductase and Is Regulated by a Redox-Responsive Repressor

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Protein Production and Purification
2.3. Determination of Protein Concentration
2.4. Protein Crystallization
2.5. X-ray Data Collection and Processing
2.6. Structure Solution and Refinement
2.7. arsH Gene Transcript Analysis
2.8. Enzyme Assays
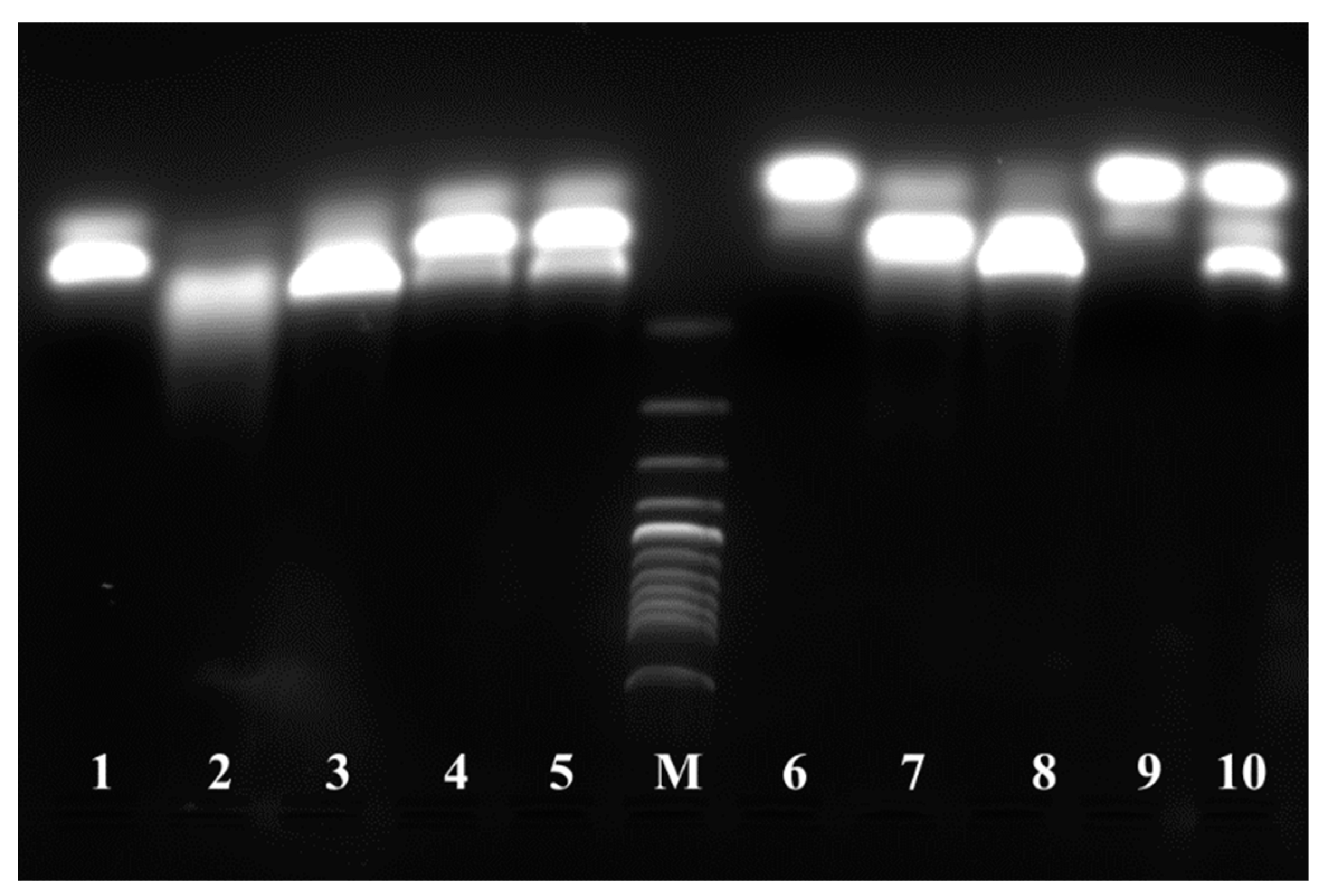
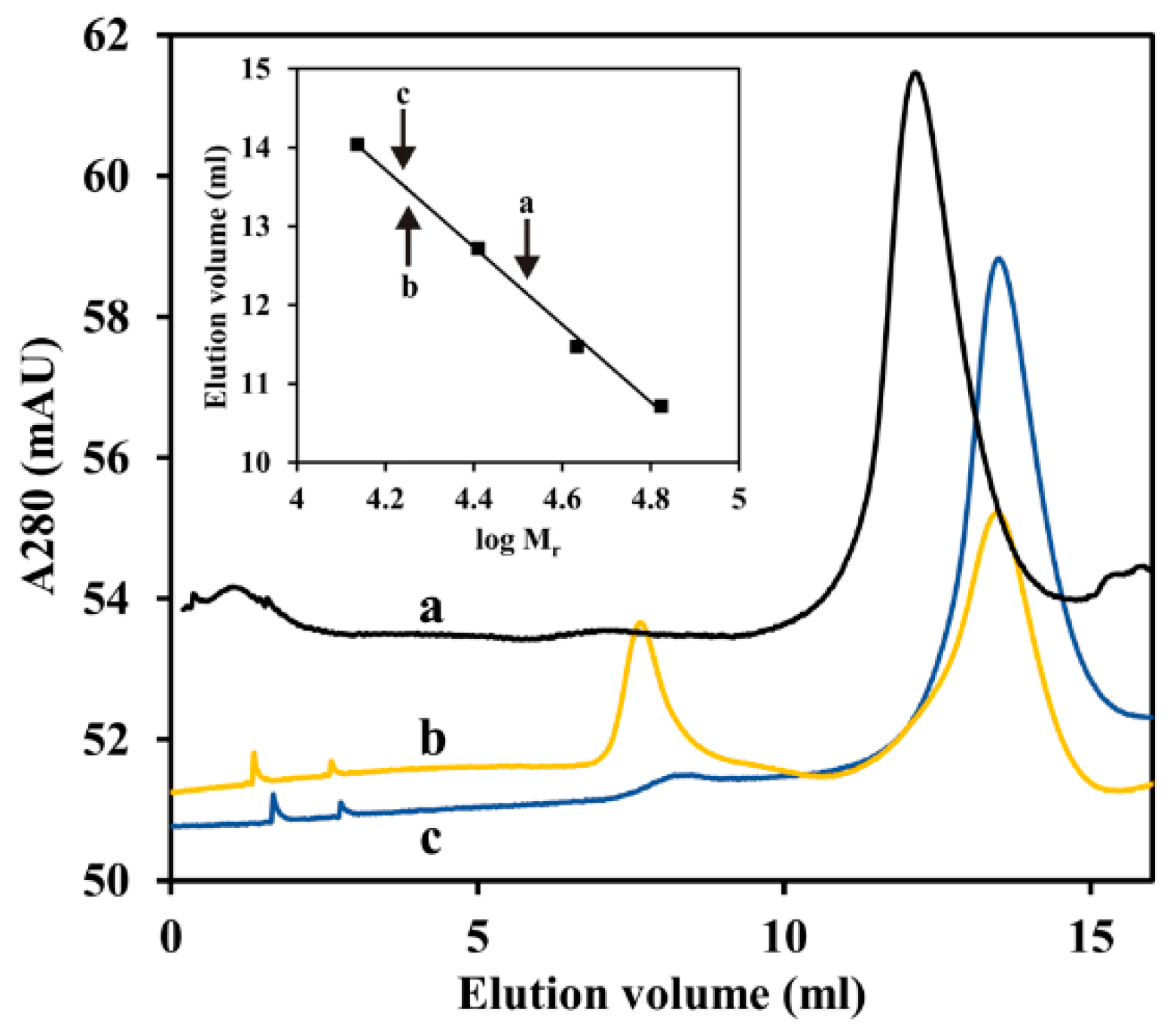
2.9. ArsR Molecular Mass Determination
2.10. Electromobility Shift Assay
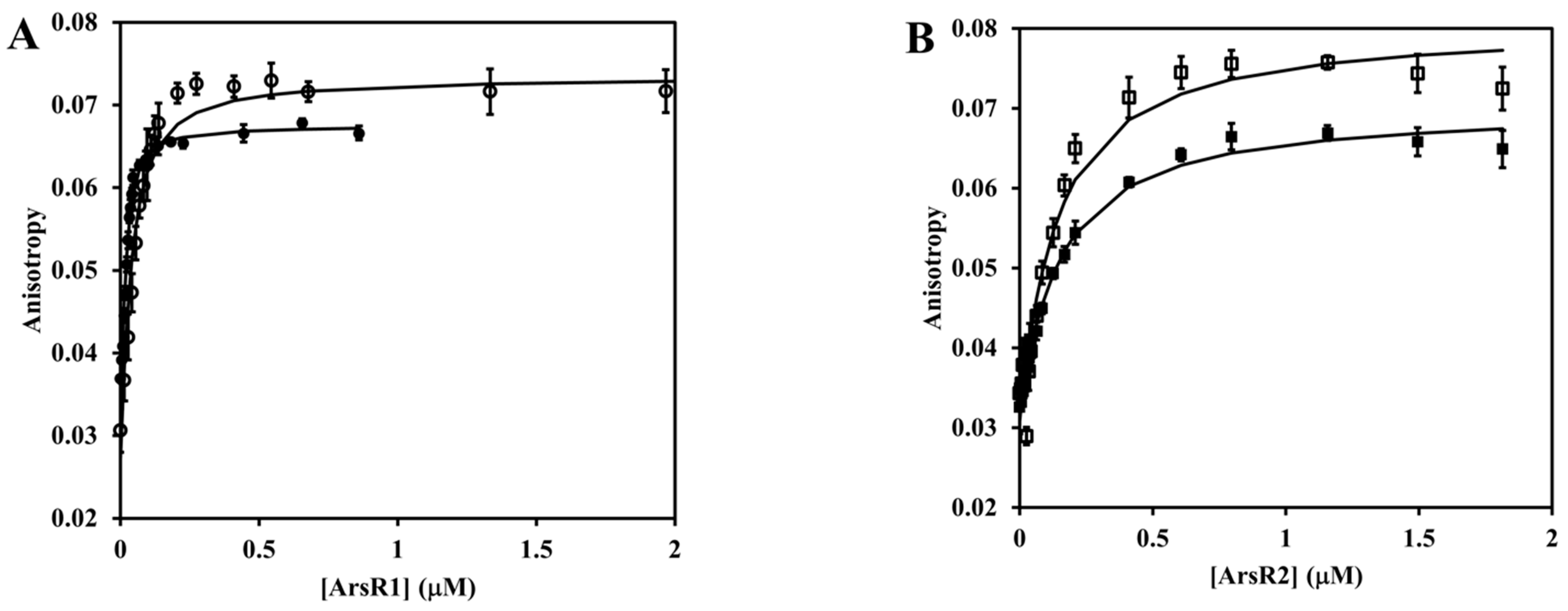
2.11. Fluorescence Anisotropy Measurement
3. Results
3.1. arsH Genes in the Genus Paracoccus
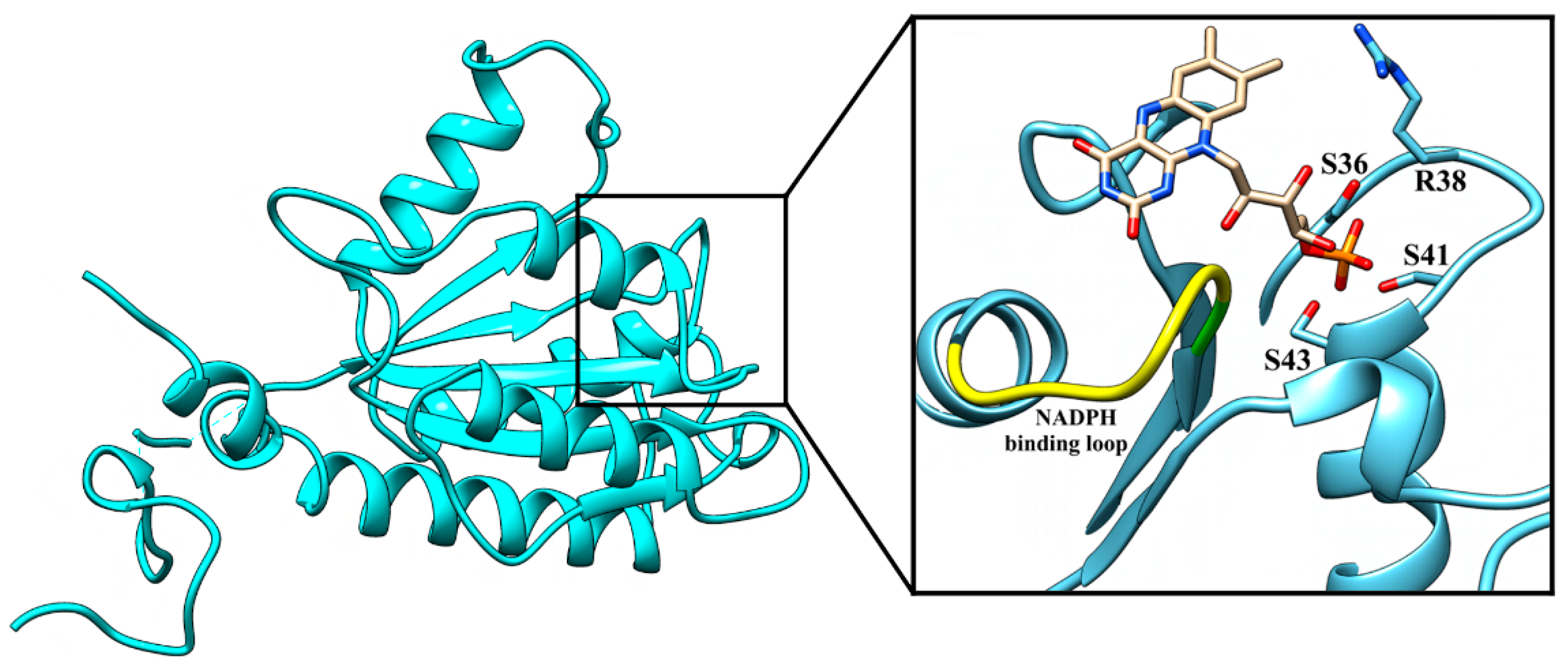
3.2. ArsH Structure
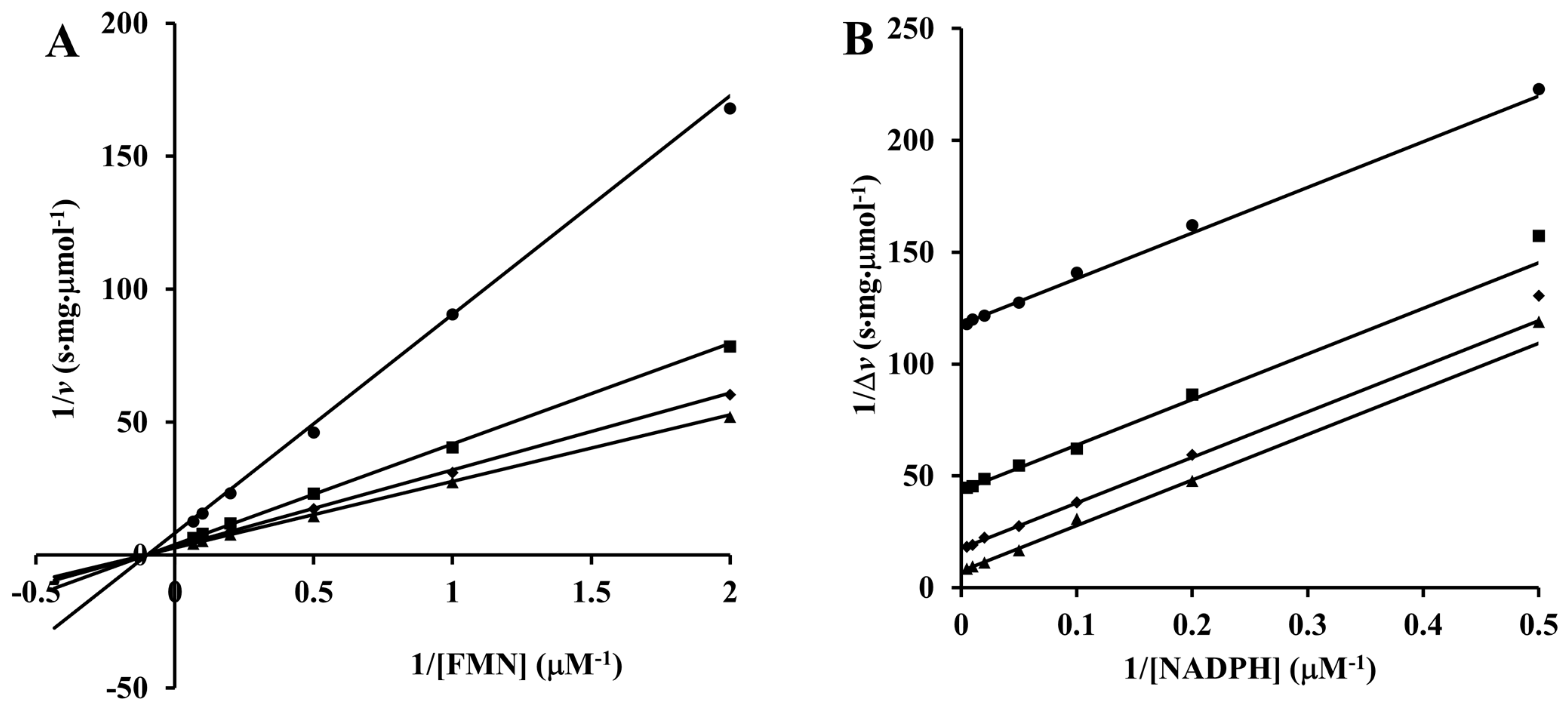
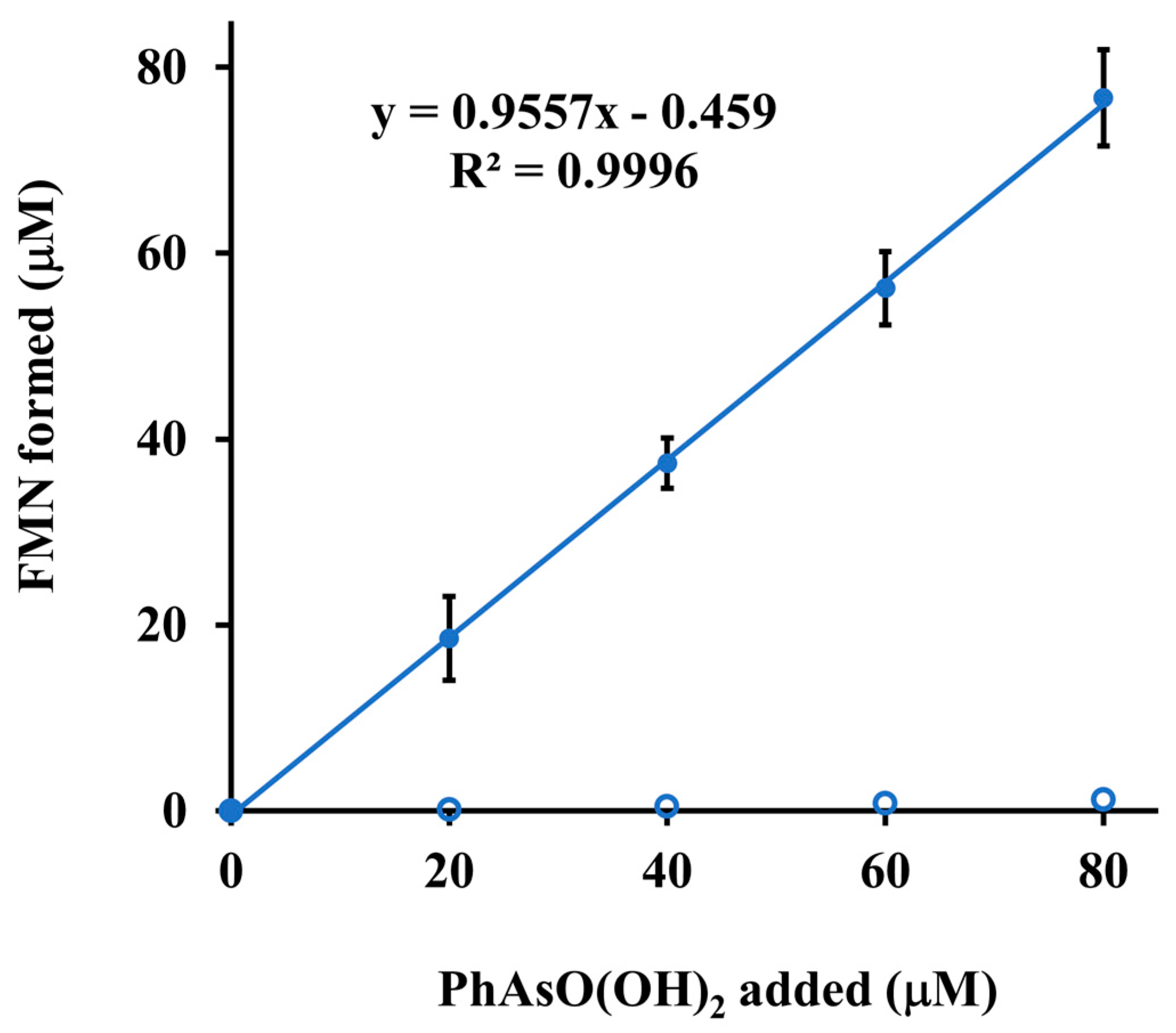
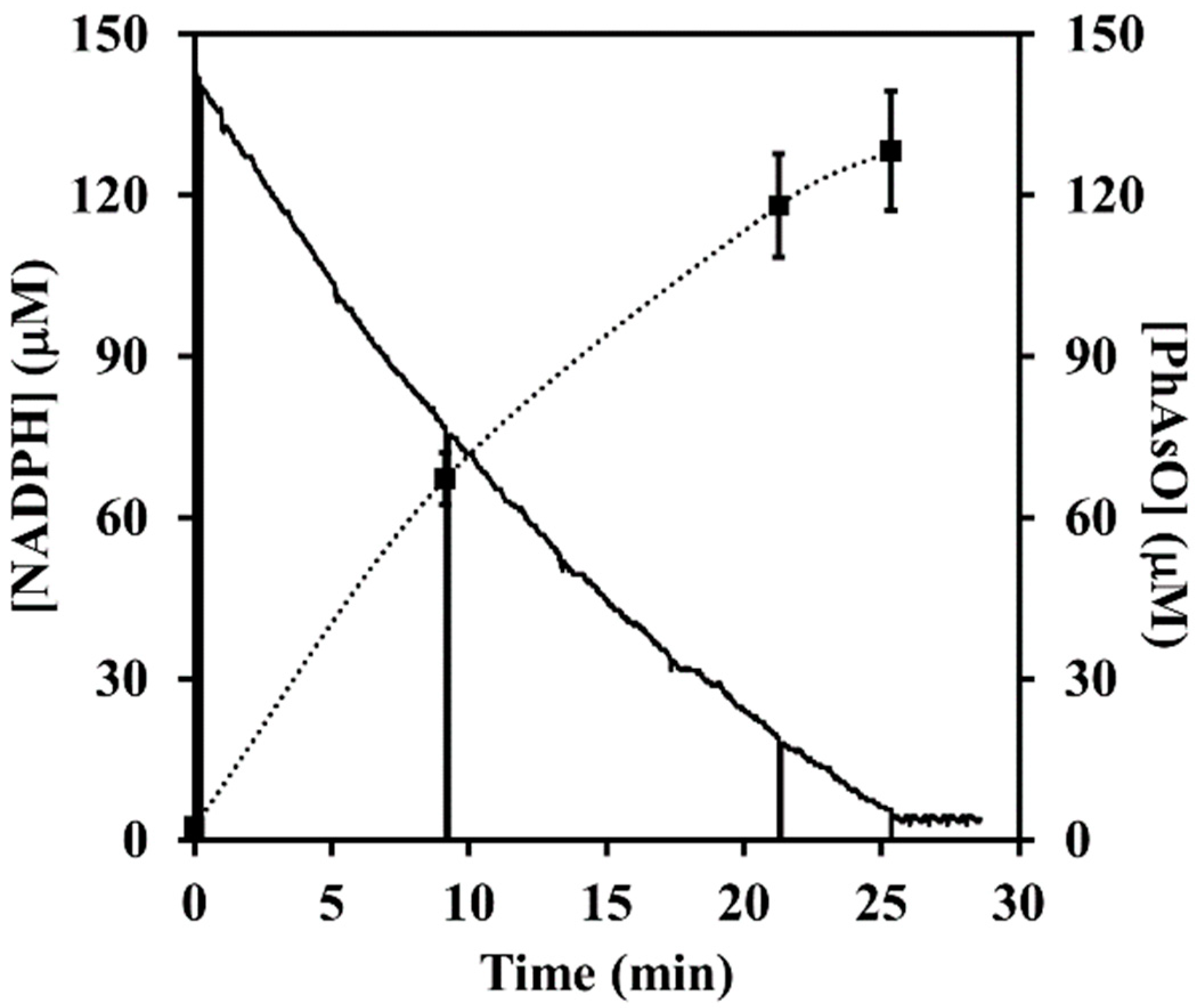
3.3. Enzymatic Activities of ArsH
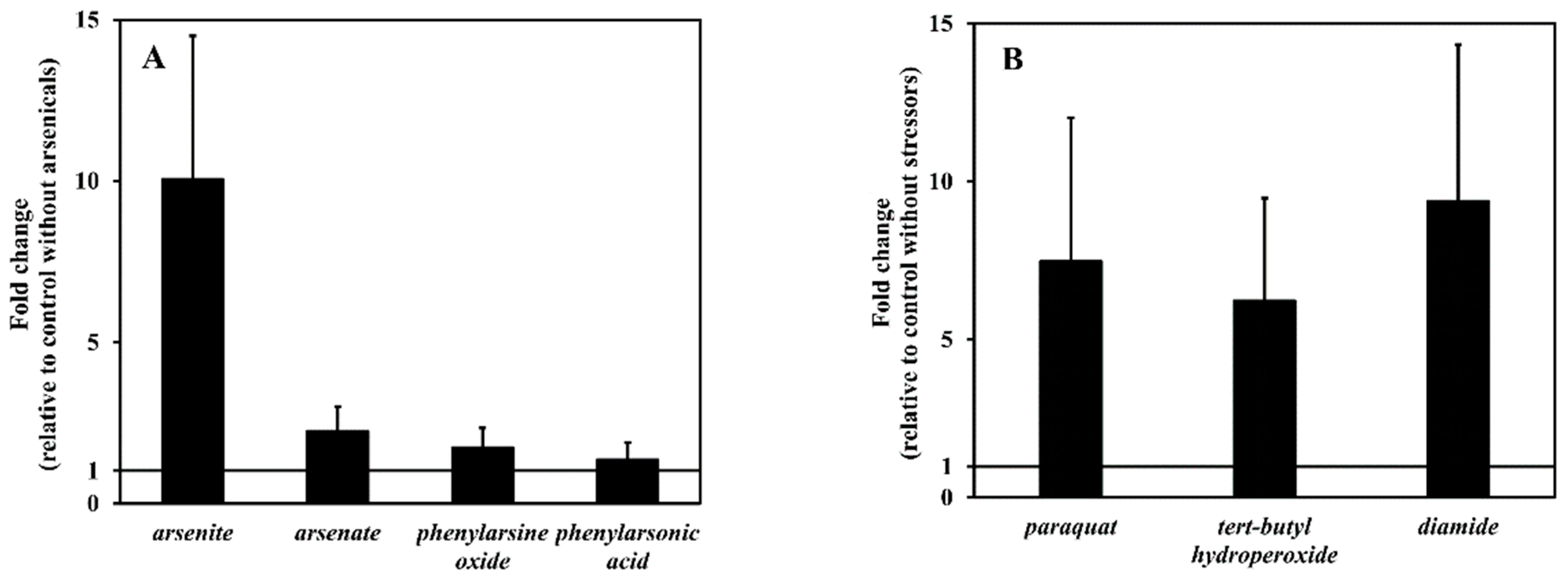
3.4. arsH Gene Expression Elicited by Arsenicals
3.5. arsH Gene Expression Elicited by Redox Stressors
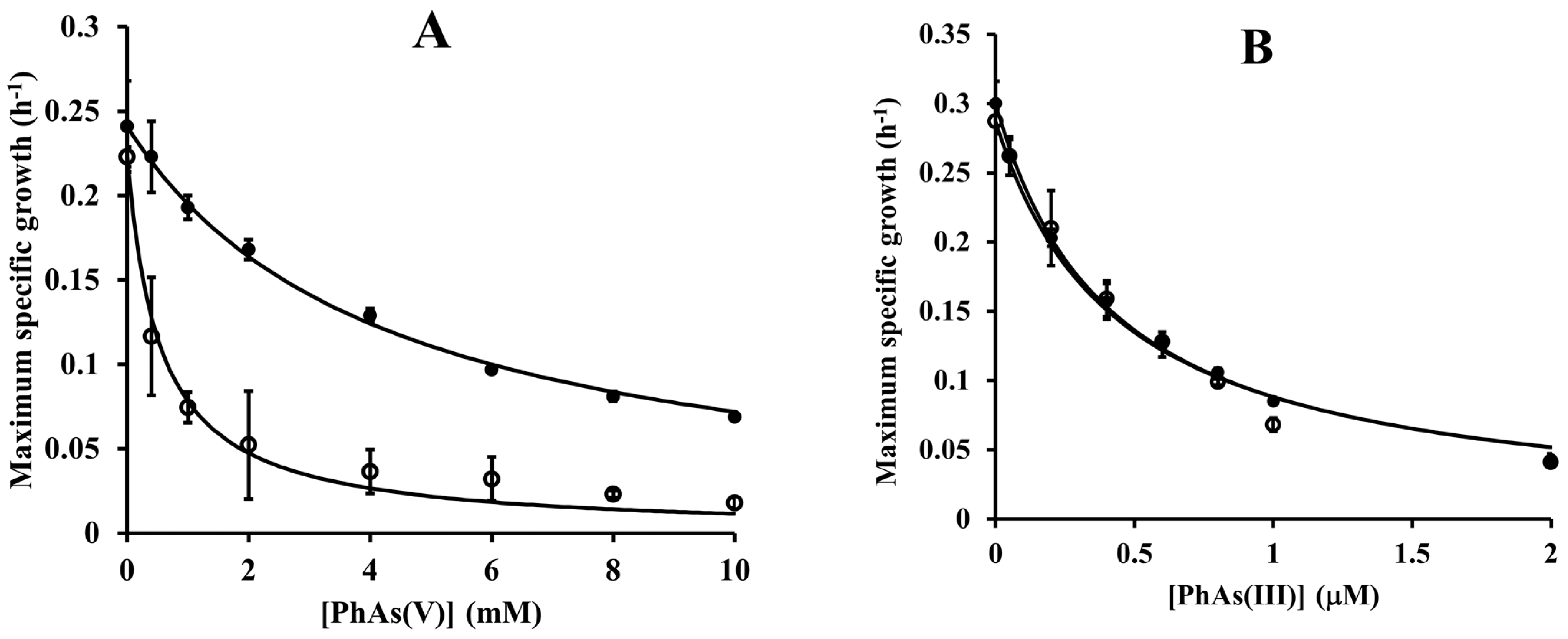
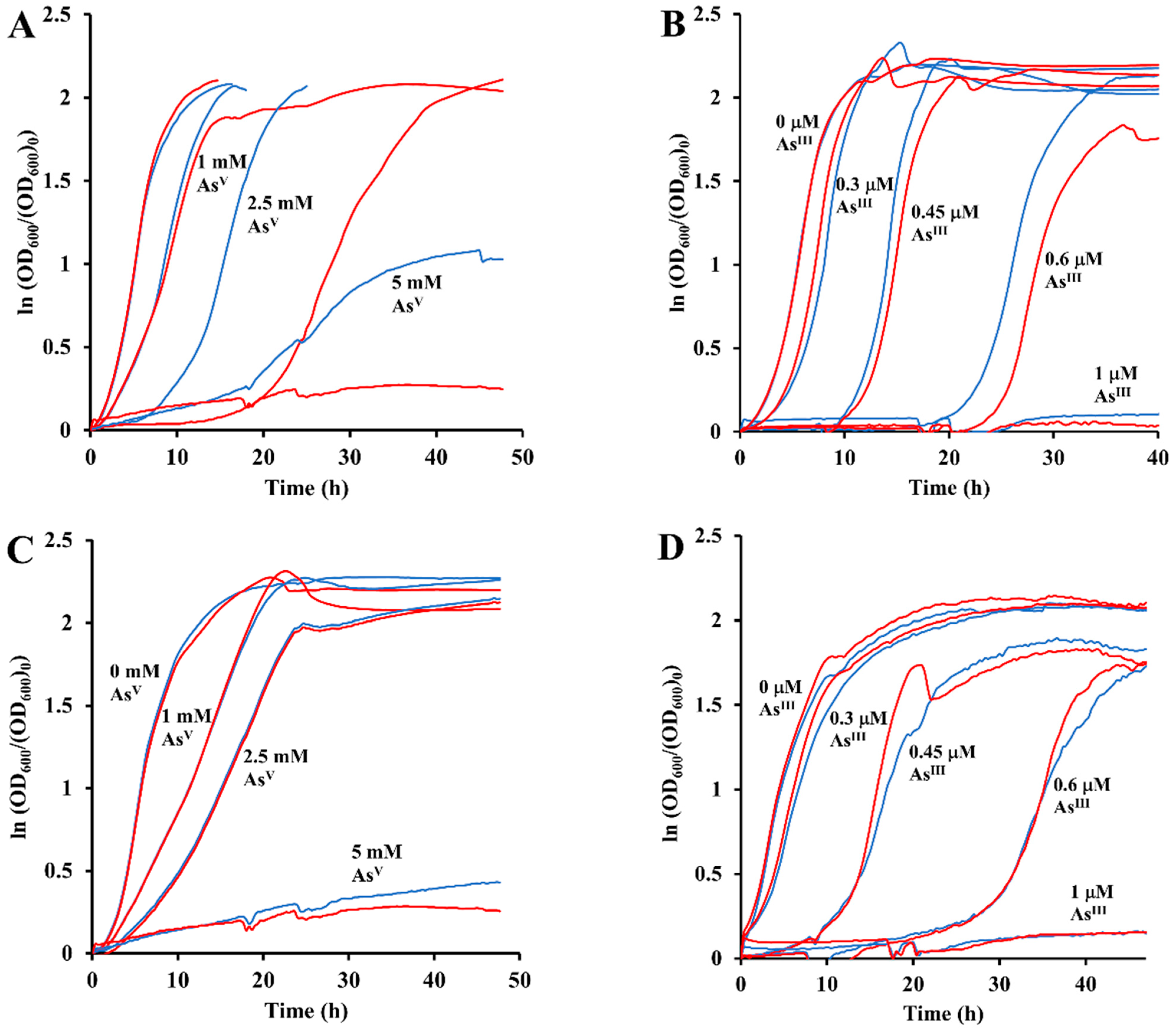
3.6. Synergistic Growth Inhibitory Effects
3.7. Effect of Heterologously Expressed arsH Gene
3.8. ArsR Repressor Functioning
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baker, S.C.; Ferguson, S.J.; Ludwig, B.; Page, M.D.; Richter, O.M.; van Spanning, R.J. Molecular genetics of the genus Paracoccus: Metabolically versatile bacteria with bioenergetic flexibility. Microbiol. Mol. Biol. Rev. 1998, 62, 1046–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedlacek, V.; Klumpler, T.; Marek, J.; Kucera, I. The structural and functional basis of catalysis mediated by NAD(P)H:acceptor oxidoreductase (FerB) of Paracoccus denitrificans. PLoS ONE 2014, 9, e96262. [Google Scholar] [CrossRef] [PubMed]
- Mazoch, J.; Tesarik, R.; Sedlacek, V.; Kucera, I.; Turanek, J. Isolation and biochemical characterization of two soluble iron(III) reductases from Paracoccus denitrificans. Eur. J. Biochem. 2004, 271, 553–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedlacek, V.; van Spanning, R.J.M.; Kucera, I. Characterization of the quinone reductase activity of the ferric reductase B protein from Paracoccus denitrificans. Arch. Biochem. Biophys. 2009, 483, 29–36. [Google Scholar] [CrossRef]
- Sedlacek, V.; Ptackova, N.; Rejmontova, P.; Kucera, I. The flavoprotein FerB of Paracoccus denitrificans binds to membranes, reduces ubiquinone and superoxide, and acts as an in vivo antioxidant. FEBS J. 2015, 282, 283–296. [Google Scholar] [CrossRef]
- Sedlacek, V.; Kucera, I. Chromate reductase activity of the Paracoccus denitrificans ferric reductase B (FerB) protein and its physiological relevance. Arch. Microbiol. 2010, 192, 919–926. [Google Scholar] [CrossRef]
- Sedlacek, V.; Kucera, I. Arginine-95 is important for recruiting superoxide to the active site of the FerB flavoenzyme of Paracoccus denitrificans. FEBS Lett. 2019, 593, 697–702. [Google Scholar] [CrossRef]
- Pernikarova, V.; Sedlacek, V.; Potesil, D.; Prochazkova, I.; Zdrahal, Z.; Bouchal, P.; Kucera, I. Proteomic responses to a methyl viologen-induced oxidative stress in the wild type and FerB mutant strains of Paracoccus denitrificans. J. Proteom. 2015, 125, 68–75. [Google Scholar] [CrossRef]
- Sedlacek, V.; Kucera, I. Functional and mechanistic characterization of an atypical flavin reductase encoded by the pden_5119 gene in Paracoccus denitrificans. Mol. Microbiol. 2019, 112, 166–183. [Google Scholar] [CrossRef]
- Paez-Espino, D.; Tamames, J.; de Lorenzo, V.; Canovas, D. Microbial responses to environmental arsenic. Biometals 2009, 22, 117–130. [Google Scholar] [CrossRef]
- Mo, H.; Chen, Q.; Du, J.; Tang, L.; Qin, F.; Miao, B.; Wu, X.; Zeng, J. Ferric reductase activity of the ArsH protein from Acidithiobacillus ferrooxidans. J. Microbiol. Biotechnol. 2011, 21, 464–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, X.M.; Yan, Y.; Xu, H.J.; Wang, N.; Zhang, X.; Ye, J. ArsH from Synechocystis sp. PCC 6803 reduces chromate and ferric iron. FEMS Microbiol. Lett. 2014, 356, 105–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hervas, M.; Lopez-Maury, L.; Leon, P.; Sanchez-Riego, A.M.; Florencio, F.J.; Navarro, J.A. ArsH from the cyanobacterium Synechocystis sp. PCC 6803 is an efficient NADPH-dependent quinone reductase. Biochemistry 2012, 51, 1178–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorontsov, I.I.; Minasov, G.; Brunzelle, J.S.; Shuvalova, L.; Kiryukhina, O.; Collart, F.R.; Anderson, W.F. Crystal structure of an apo form of Shigella flexneri ArsH protein with an NADPH-dependent FMN reductase activity. Protein Sci. 2007, 16, 2483–2490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Yang, H.C.; Rosen, B.P.; Bhattacharjee, H. Crystal structure of the flavoprotein ArsH from Sinorhizobium meliloti. FEBS Lett. 2007, 581, 3996–4000. [Google Scholar] [CrossRef] [Green Version]
- Crescente, V.; Holland, S.M.; Kashyap, S.; Polycarpou, E.; Sim, E.; Ryan, A. Identification of novel members of the bacterial azoreductase family in Pseudomonas aeruginosa. Biochem. J. 2016, 473, 549–558. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Bhattacharjee, H.; Rosen, B.P. ArsH is an organoarsenical oxidase that confers resistance to trivalent forms of the herbicide monosodium methylarsenate and the poultry growth promoter roxarsone. Mol. Microbiol. 2015, 96, 1042–1052. [Google Scholar] [CrossRef] [Green Version]
- Paez-Espino, A.D.; Nikel, P.I.; Chavarria, M.; de Lorenzo, V. ArsH protects Pseudomonas putida from oxidative damage caused by exposure to arsenic. Environ. Microbiol. 2020, 22, 2230–2242. [Google Scholar] [CrossRef]
- Wijtzes, T.; de Wit, J.C.; In, H.; Van’t, R.; Zwietering, M.H. Modelling bacterial growth of Lactobacillus curvatus as a function of acidity and temperature. Appl. Environ. Microbiol. 1995, 61, 2533–2539. [Google Scholar] [CrossRef] [Green Version]
- Tesarik, R.; Sedlacek, V.; Plockova, J.; Wimmerova, M.; Turanek, J.; Kucera, I. Heterologous expression and molecular characterization of the NAD(P)H:acceptor oxidoreductase (FerB) of Paracoccus denitrificans. Protein Expres. Purif. 2009, 68, 233–238. [Google Scholar] [CrossRef]
- Battye, T.G.; Kontogiannis, L.; Johnson, O.; Powell, H.R.; Leslie, A.G. iMOSFLM: A new graphical interface for diffraction-image processing with MOSFLM. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 271–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krissinel, E. Ccp4 software suite: History, evolution, content, challenges and future developments. Arbor 2015, 191, a220. [Google Scholar] [CrossRef] [Green Version]
- Long, F.; Vagin, A.A.; Young, P.; Murshudov, G.N. BALBES: A molecular-replacement pipeline. Acta Crystallogr. D Biol. Crystallogr. 2008, 64, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Vagin, A.A.; Steiner, R.A.; Lebedev, A.A.; Potterton, L.; McNicholas, S.; Long, F.; Murshudov, G.N. REFMAC5 dictionary: Organization of prior chemical knowledge and guidelines for its use. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2184–2195. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Horecker, B.L.; Kornberg, A. The extinction coefficients of the reduced band of pyridine nucleotides. J. Biol. Chem. 1948, 175, 385–390. [Google Scholar] [CrossRef]
- Cleland, W.W. Kinetics of enzyme-catalyzed reactions with 2 or more substrates or products. I. Nomenclature and rate equations. Biochim. Biophys. Acta 1963, 67, 104–137. [Google Scholar] [CrossRef]
- Whitby, L.G. A new nethod for preparing flavin-adenine dinucleotide. Biochem. J. 1953, 54, 437–442. [Google Scholar] [CrossRef] [Green Version]
- Heyduk, T.; Lee, J.C. Application of fluorescence energy-transfer and polarization to monitor Escherichia coli cAMP receptor protein and lac promoter interaction. Proc. Natl. Acad. Sci. USA 1990, 87, 1744–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, R.; Bonanno, J.B.; Burley, S.K.; Swaminathan, S. Structure determination of an FMN reductase from Pseudomonas aeruginosa PA01 using sulfur anomalous signal. Acta Crystallogr. D 2006, 62, 383–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, Y.H.; Wu, C.Y.; Sargsyan, K.; Lim, C. Sequence-motif detection of NAD(P)-binding proteins: Discovery of a unique antibacterial drug target. Sci. Rep. 2014, 4, 6471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.H.; Rosen, B.P. Metalloregulated expression of the ars operon. J. Biol. Chem. 1993, 268, 52–58. [Google Scholar] [CrossRef]
- Prabaharan, C.; Kandavelu, P.; Packianathan, C.; Rosen, B.P.; Thiyagarajana, S. Structures of two ArsR As(III)-responsive transcriptional repressors: Implications for the mechanism of derepression. J. Struct. Biol. 2019, 207, 209–217. [Google Scholar] [CrossRef]
- Wang, L.P.; Jeon, B.W.; Sahin, O.; Zhang, Q.J. Identification of an arsenic resistance and arsenic-sensing system in Campylobacter jejuni. Appl. Environ. Microb. 2009, 75, 5064–5073. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Rosen, B.P. Dimerization is essential for DNA binding and repression by the ArsR metalloregulatory protein of Escherichia coli. J. Biol. Chem. 1997, 272, 15734–15738. [Google Scholar] [CrossRef] [Green Version]
- Kretzschmar, J.; Brendler, E.; Wagler, J.; Schmidt, A.C. Kinetics and activation parameters of the reaction of organoarsenic(V) compounds with glutathione. J. Hazard. Mater. 2014, 280, 734–740. [Google Scholar] [CrossRef]
- Nuallain, C.O.; Cinneide, S.O. Thermodynamic ionization constants of aromatic arsonic acids. J. Inorg. Nucl. Chem. 1973, 35, 2871–2881. [Google Scholar] [CrossRef]
- Millis, K.K.; Weaver, K.H.; Rabenstein, D.L. Oxidation/reduction potential of glutathione. J. Org. Chem. 1993, 58, 4144–4146. [Google Scholar] [CrossRef]
- Knowles, F.C. Reactions of lipoamide dehydrogenase and glutathione reductase with arsonic acids and arsonous acids. Arch. Biochem. Biophys. 1985, 242, 1–10. [Google Scholar] [CrossRef]
- Walsh, C.T.; Wencewicz, T.A. Flavoenzymes: Versatile catalysts in biosynthetic pathways. Nat. Prod. Rep. 2013, 30, 175–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayhew, S.G. The effects of pH and semiquinone formation on the oxidation-reduction potentials of flavin mononucleotide. A reappraisal. Eur. J. Biochem. 1999, 265, 698–702. [Google Scholar] [CrossRef]
- Pi, K.F.; Markelova, E.; Zhang, P.; van Cappellen, P. Arsenic oxidation by flavin-derived reactive species under oxic and anoxic conditions: Oxidant Formation and pH Dependence. Environ. Sci. Technol. 2019, 53, 10897–10905. [Google Scholar] [CrossRef]
- Hirose, K.; Ezaki, B.; Liu, T.; Nakashima, S. Diamide stress induces a metallothionein BmtA through a repressor BxmR and is modulated by Zn-inducible BmtA in the cyanobacterium Oscillatoria brevis. Toxicol. Lett. 2006, 163, 250–256. [Google Scholar] [CrossRef]
- Ehira, S.; Ohmori, M. The redox-sensing transcriptional regulator RexT controls expression of thioredoxin A2 in the Cyanobacterium anabaena sp. strain PCC 7120. J. Biol. Chem. 2012, 287, 40433–40440. [Google Scholar] [CrossRef] [Green Version]
- Palm, G.J.; Chi, B.K.; Waack, P.; Gronau, K.; Becher, D.; Albrecht, D.; Hinrichs, W.; Read, R.J.; Antelmann, H. Structural insights into the redox-switch mechanism of the MarR/DUF24-type regulator HypR. Nucleic Acids Res. 2012, 40, 4178–4192. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes, B.G.; Barbosa, R.L.; Soprano, A.S.; Campos, B.M.; de Souza, T.A.; Tonoli, C.C.C.; Leme, A.F.P.; Murakami, M.T.; Benedetti, C.E. Plant pathogenic bacteria utilize biofilm growth-associated repressor (BigR), a novel winged-helix redox switch, to control hydrogen sulfide detoxification under hypoxia. J. Biol. Chem. 2011, 286, 26148–26157. [Google Scholar] [CrossRef] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sedláček, V.; Kryl, M.; Kučera, I. The ArsH Protein Product of the Paracoccus denitrificans ars Operon Has an Activity of Organoarsenic Reductase and Is Regulated by a Redox-Responsive Repressor. Antioxidants 2022, 11, 902. https://doi.org/10.3390/antiox11050902
Sedláček V, Kryl M, Kučera I. The ArsH Protein Product of the Paracoccus denitrificans ars Operon Has an Activity of Organoarsenic Reductase and Is Regulated by a Redox-Responsive Repressor. Antioxidants. 2022; 11(5):902. https://doi.org/10.3390/antiox11050902
Chicago/Turabian StyleSedláček, Vojtěch, Martin Kryl, and Igor Kučera. 2022. "The ArsH Protein Product of the Paracoccus denitrificans ars Operon Has an Activity of Organoarsenic Reductase and Is Regulated by a Redox-Responsive Repressor" Antioxidants 11, no. 5: 902. https://doi.org/10.3390/antiox11050902