
Ginseng, Tribulus Extracts and Pollen Grains Supplementation Improves Sexual State, Testes Redox Status, and Testicular Histology in Nile Tilapia Males

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish and Experimental Facilities
2.2. Experimental Design
2.3. Experimental Diets
2.4. Samples Collection
2.5. Measured Parameters
2.5.1. Testes Somatic Index
2.5.2. Sexual Hormones
- Luteinizing hormone determination
- Testosterone determination
2.5.3. Antioxidant Status Assays
- The glutathione S-transferases
- Glutathione peroxidase
- Catalase
- Superoxide dismutase
- Reduced glutathione
- Thiobarbituric acid-reactive substances
2.5.4. Histological Examination
2.6. Statistical Analysis
3. Results
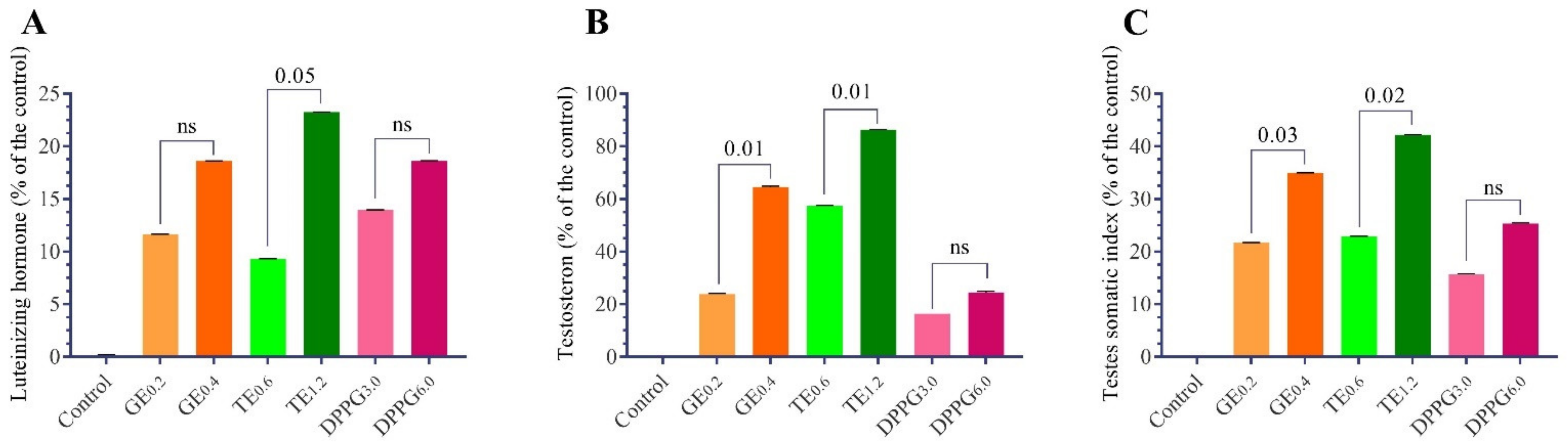
3.1. Sexual Hormones and Testis Somatic Index
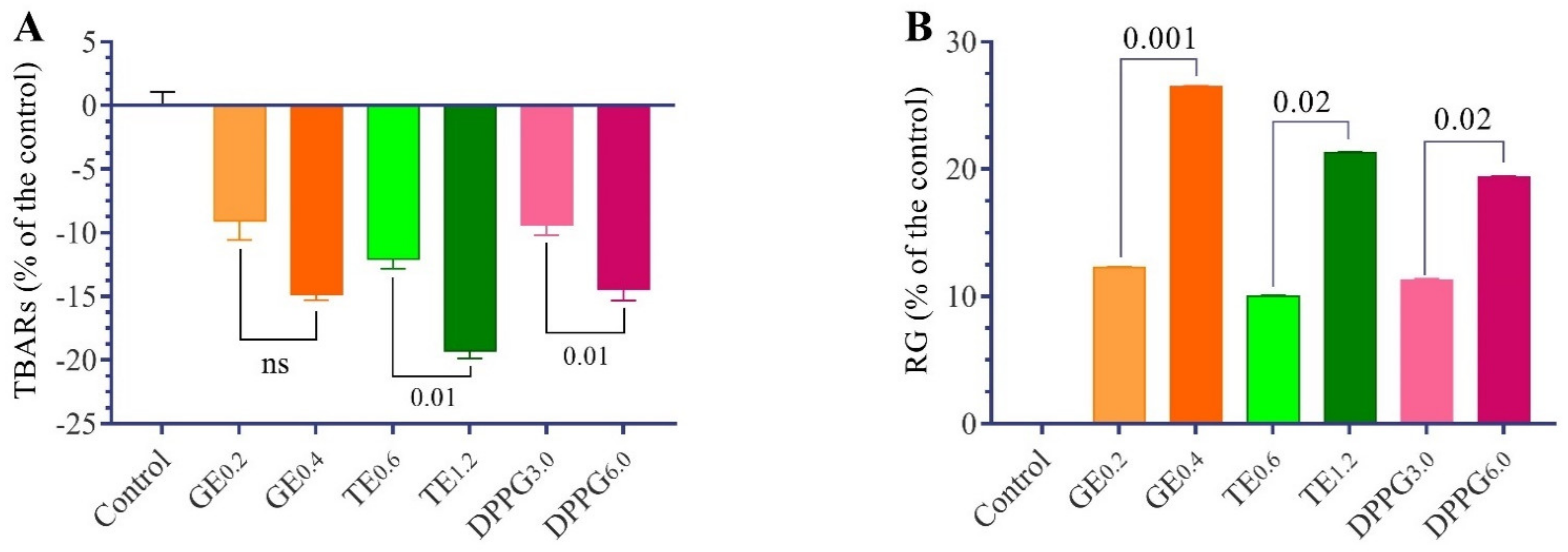
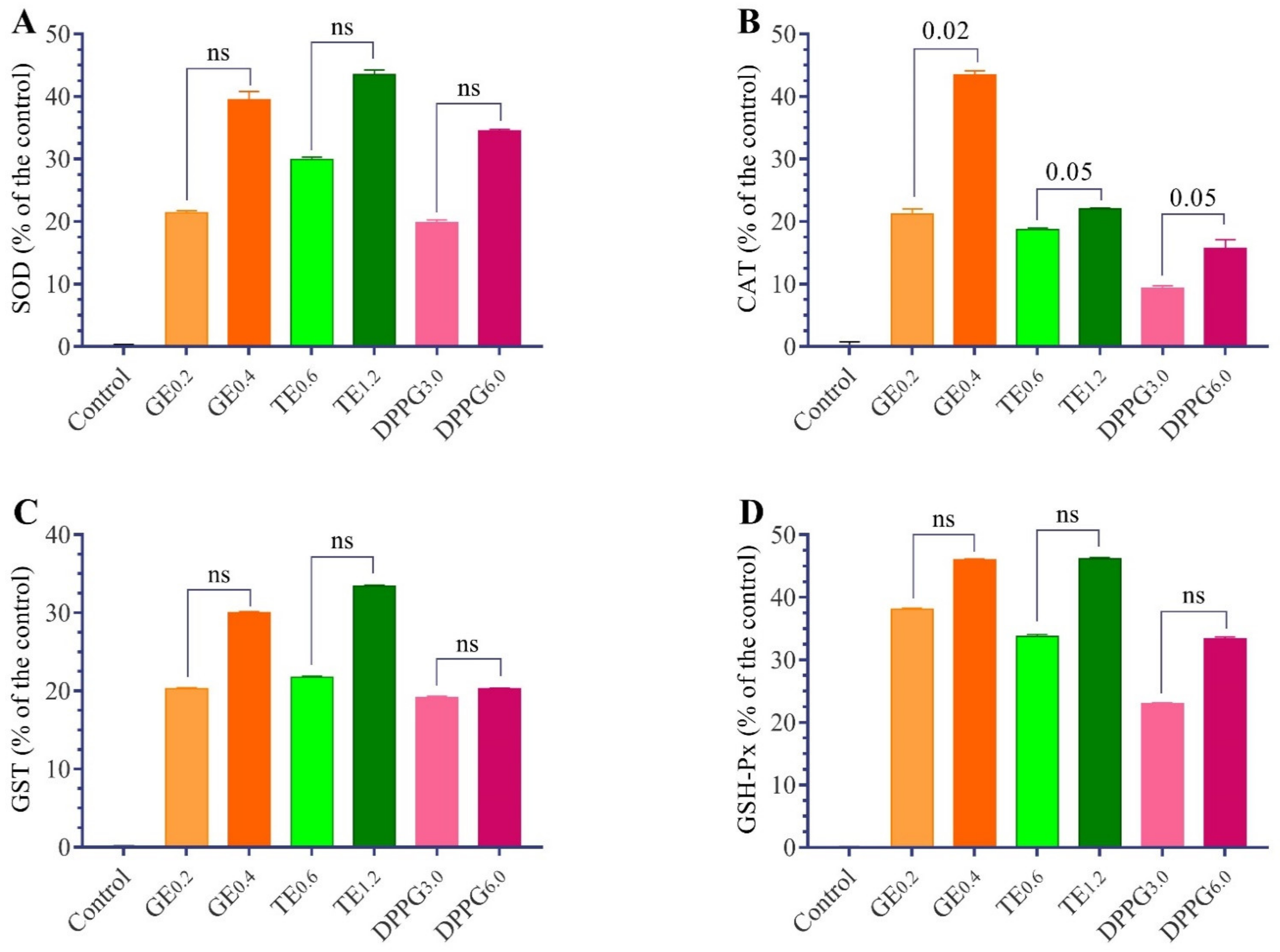
3.2. Antioxidant Status
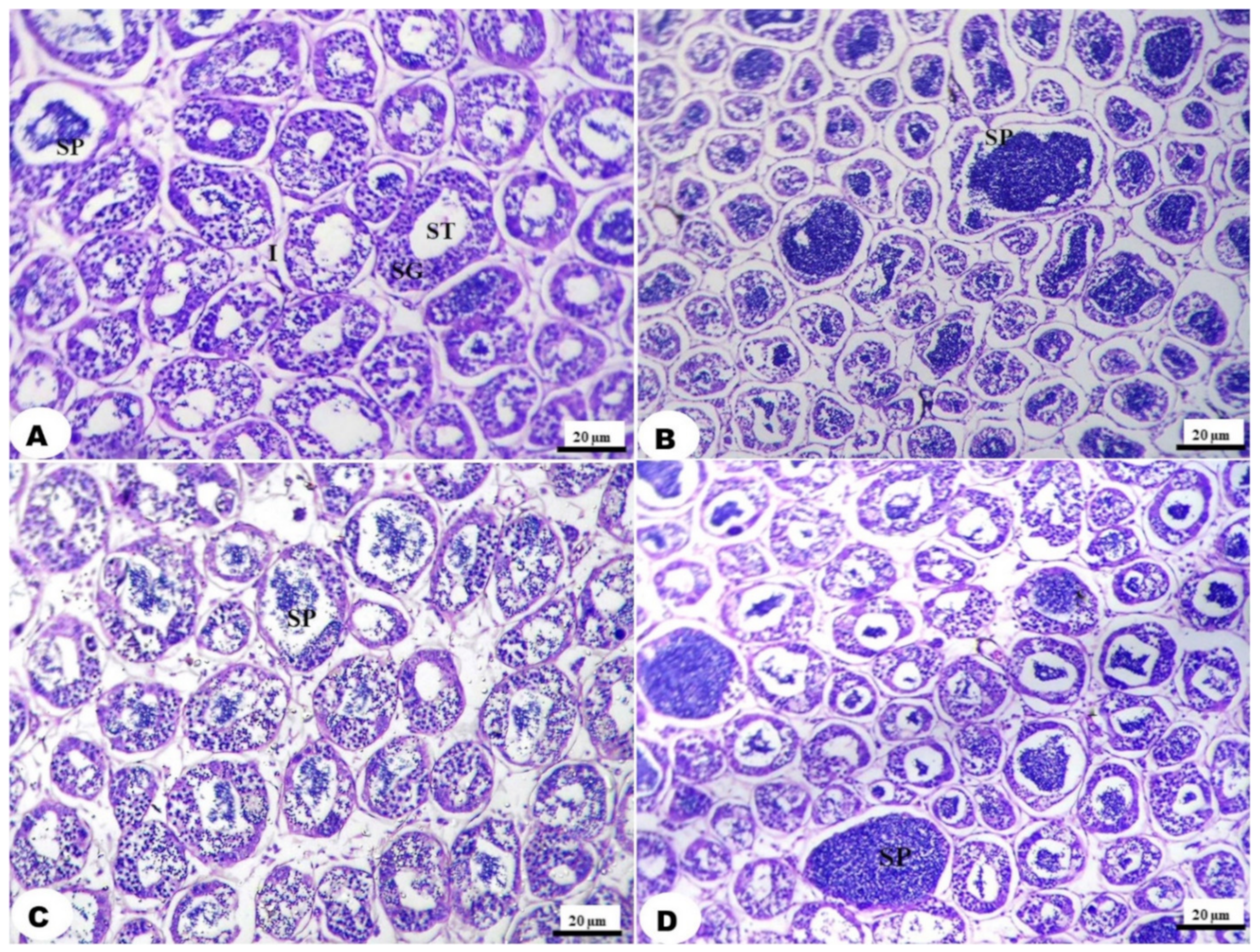
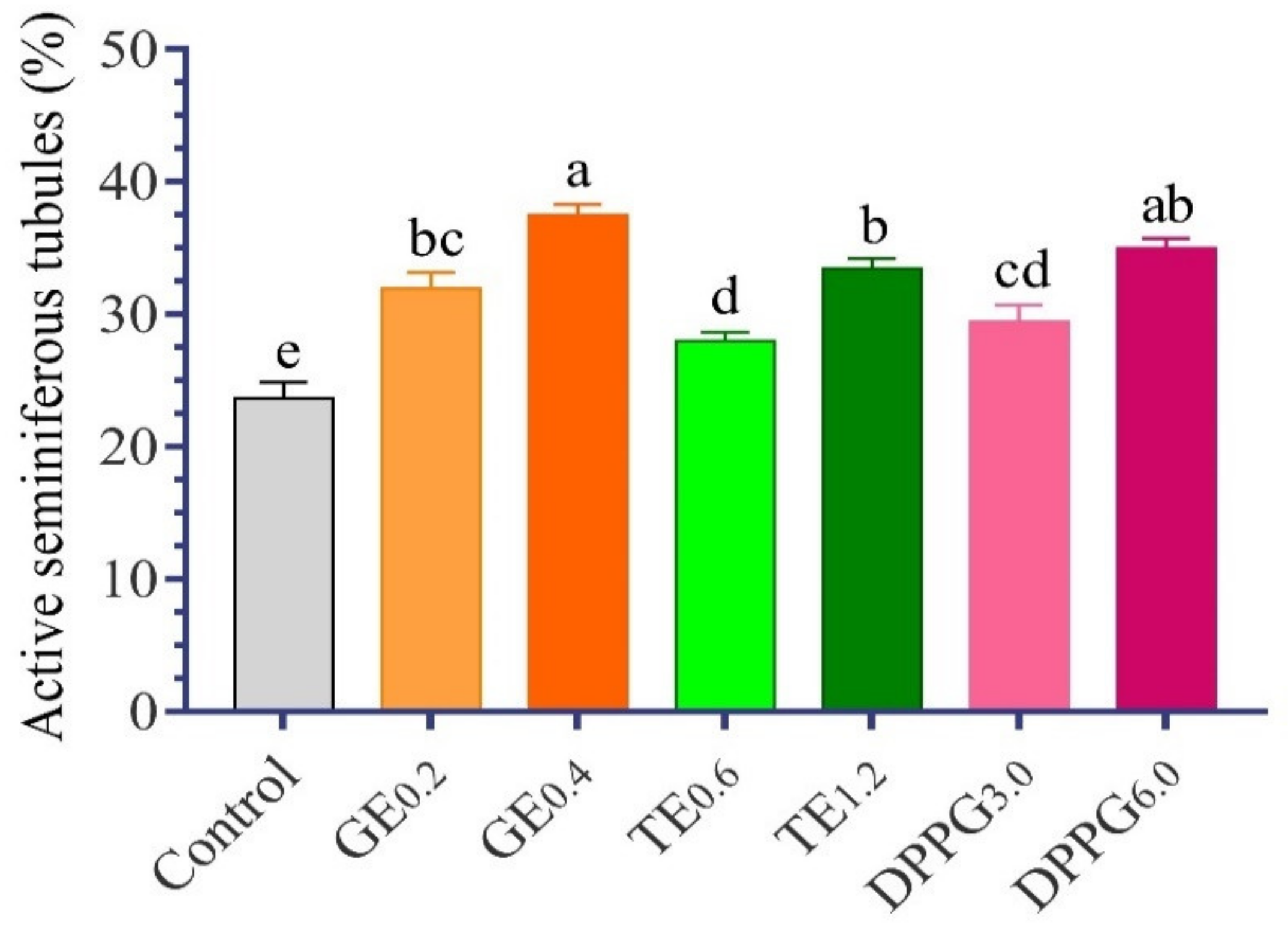
3.3. Histological Examination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halliwell, B. Oxidative stress and neurodegeneration: Where are we now? J. Neurochem. 2006, 97, 1634–1658. [Google Scholar] [CrossRef] [PubMed]
- Asadi, N.; Bahmani, M.; Kheradmand, A.; Rafieian-Kopaei, M. The impact of oxidative stress on testicular function and the role of antioxidants in improving it: A review. J. Clin. Diagn. Res. 2017, 11, IE01–IE05. [Google Scholar] [CrossRef] [PubMed]
- Mora-Esteves, C.; Shin, D. Nutrient supplementation: Improving male fertility fourfold. Semin. Reprod. Med. 2013, 31, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Cabrita, E.; Martínez-Páramo, S.; Gavaia, P.J.; Riesco, M.F.; Valcarce, D.; Sarasquete, C.; Herráez, M.; Robles, V. Factors enhancing fish sperm quality and emerging tools for sperm analysis. Aquaculture 2014, 432, 389–401. [Google Scholar] [CrossRef]
- Schreiber, E.; Garcia, T.; Sharma, R.P.; Torrente, M.; Domingo, J.L.; Gómez, M. Oxidative stress in testes of rats exposed to n-butylparaben. Food Chem. Toxicol. 2019, 131, 110573. [Google Scholar] [CrossRef]
- Sonmez, A.; Ozdemir, R.; Bilen, S.; Kadak, A. Effect of Ginseng Root (Araliaceae sp.) Extracts on Sperm Quality Parameters and Reproductive Performance in Rainbow Trout (Oncorhynchus mykiss). Isr. J. Aquac. 2019, 71, 1570–1576. [Google Scholar] [CrossRef]
- Yeganeh, S.; Sotoudeh, A.; Movaffagh, A.N. Effects of Tribulus terrestris extract on growth and reproductive performance of male convict cichlid (Cichlasoma nigrofasciatum). Turk. J. Fish. Aquat. Sci. 2017, 17, 1003–1007. [Google Scholar] [CrossRef]
- Hassona, N.N.; Zayed, M.M.; Eltras, W.F.; Mohamed, R.A. Dietary supplementation of Tribulus terrestris extract improves growth and reproductive performances of the male Nile tilapia (Oreochromis niloticus). Aquac. Res. 2020, 51, 4245–4254. [Google Scholar] [CrossRef]
- Chakraborty, S.B.; Horn, P.; Hancz, C. Application of phytochemicals as growth-promoters and endocrine modulators in fish culture. Rev. Aquac. 2014, 6, 1–19. [Google Scholar] [CrossRef]
- Shakya, S.R. Effect of herbs and herbal products feed supplements on growth in fishes: A review. Nepal J. Biotechnol. 2017, 5, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Shahrajabian, M.H.; Sun, W.; Cheng, Q. A review of ginseng species in different regions as a multipurpose herb in traditional Chinese medicine, modern herbology and pharmacological science. J. Med. Plants Res. 2019, 13, 213–226. [Google Scholar]
- Guo, Q.; Cui, S.W.; Kang, J.; Ding, H.; Wang, Q.; Wang, C. Non-starch polysaccharides from American ginseng: Physicochemical investigation and structural characterization. Food Hydrocoll. 2015, 44, 320–327. [Google Scholar] [CrossRef]
- Patel, S.; Rauf, A. Adaptogenic herb ginseng (Panax) as medical food: Status quo and future prospects. Biomed. Pharmacother. 2017, 85, 120–127. [Google Scholar] [CrossRef]
- Sun, L.; Ropartz, D.; Cui, L.; Shi, H.; Ralet, M.-C.; Zhou, Y. Structural characterization of rhamnogalacturonan domains from Panax ginseng CA Meyer. Carbohydr. Polym. 2019, 203, 119–127. [Google Scholar] [CrossRef]
- Chhatre, S.; Nesari, T.; Somani, G.; Kanchan, D.; Sathaye, S. Phytopharmacological overview of Tribulus terrestris. Pharmacogn. Rev. 2014, 8, 45. [Google Scholar] [CrossRef] [Green Version]
- Kostova, I.; Dinchev, D. Saponins in Tribulus terrestris—Chemistry and bioactivity. Phytochem. Rev. 2005, 4, 111–137. [Google Scholar] [CrossRef]
- Khare, C.P. Indian Medicinal Plants: An Illustrated Dictionary; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Singh, S.; Nair, V.; Gupta, Y.K. Evaluation of the aphrodisiac activity of Tribulus terrestris Linn. in sexually sluggish male albino rats. J. Pharmacol. Pharmacother. 2012, 3, 43. [Google Scholar] [CrossRef] [Green Version]
- Kavitha, P.; Subramanian, P. Influence of Tribulus terrestris on testicular enzyme in fresh water ornamental fish Poecilia latipinna. Fish Physiol. Biochem. 2011, 37, 801–807. [Google Scholar] [CrossRef]
- Gharaei, A.; Ebrahimi Jorjani, H.; Mirdar Harijani, J.; Kolangi Miandare, H. Effects of Tribullus terrestris extract on masculinization, growth indices, sex deteminationreversal and steroid hormones level in Zebra fish (Danio rerio). Int. Aquat. Res. 2020, 12, 22–29. [Google Scholar]
- Zohary, D.; Hopf, M. Date Palm Phoenix Dactylifera. In Domestication of Plants in the Old World, 2nd ed.; Clarendon: Oxford, UK, 1993; Volume 42, pp. 1077–1082. [Google Scholar]
- Hassan, H.M. Chemical composition and nutritional value of palm pollen grains. Glob. J. Biotechnol. Biochem. 2011, 6, 1–7. [Google Scholar]
- Daoud, A.; Malika, D.; Bakari, S.; Hfaiedh, N.; Mnafgui, K.; Kadri, A.; Gharsallah, N. Assessment of polyphenol composition, antioxidant and antimicrobial properties of various extracts of Date Palm Pollen (DPP) from two Tunisian cultivars. Arab. J. Chem. 2019, 12, 3075–3086. [Google Scholar] [CrossRef] [Green Version]
- Bennett, R.D.; Ko, S.-T.; Heftmann, E. Isolation of estrone and cholesterol from the date palm, Phoenix dactylifera L. Phytochemistry 1966, 5, 231–235. [Google Scholar] [CrossRef]
- Tatar, T.; Akdevelioğlu, Y. Effect of pollen, pit powder, and gemmule extract of date palm on male infertility: A systematic review. J. Am. Coll. Nutr. 2018, 37, 154–160. [Google Scholar] [CrossRef] [PubMed]
- El Hadrami, A.; Al-Khayri, J.M. Socioeconomic and traditional importance of date palm. Emir. J. Food Agric. 2012, 24, 371. [Google Scholar]
- Elberry, A.A.; Mufti, S.T.; Al-Maghrabi, J.A.; Abdel-Sattar, E.A.; Ashour, O.M.; Ghareib, S.A.; Mosli, H.A. Anti-inflammatory and antiproliferative activities of date palm pollen (Phoenix dactylifera) on experimentally-induced atypical prostatic hyperplasia in rats. J. Inflamm. 2011, 8, 40. [Google Scholar] [CrossRef] [Green Version]
- Bahmanpour, S.; Panjeh, S.M.; Talaei, T.; Vojdani, Z.; Poust, P.A.; Zareei, S.; Ghaemian, M. Effect of Phoenix dactylifera pollen on sperm parameters and reproductive system of adult male rats. Iran. J. Med. Sci. 2006, 31, 208–212. [Google Scholar]
- Uzbekova, D.; Makarova, V.; Khvoynitskaya, L.; Slepnev, A. Evaluation of bee-collected pollen influence on lipid peroxidation, antioxidant system and liver function in old animals. J. Hepatol. 2003, 38, 203. [Google Scholar] [CrossRef]
- Mansour, A.; Omar, E.; Srour, T.; Yousef, M. Effect of three natural phytochemicals supplementation on growth performance, testosterone level and feed utilization of Nile tilapia (Oreochromis niloticus). Aquacult. Nutr. 2018, 24, 408–415. [Google Scholar] [CrossRef]
- Gauthaman, K.; Ganesan, A.P. The hormonal effects of Tribulus terrestris and its role in the management of male erectile dysfunction—An evaluation using primates, rabbit and rat. Phytomedicine 2008, 15, 44–54. [Google Scholar] [CrossRef]
- Goda, A.M.S. Effect of dietary Ginseng herb (Ginsana® G115) supplementation on growth, feed utilization, and hematological indices of Nile Tilapia, Oreochromis niloticus (L.), fingerlings. J. World Aquac. Soc. 2008, 39, 205–214. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011.
- Beitins, I.; O’loughlin, K.; Ostrea, T.; McArthur, J. Gonadotropin determinations in timed 3-hour urine collections during the menstrual cycle and LHRH testing. J. Clin. Endocrinol. Metab. 1976, 43, 46–55. [Google Scholar] [CrossRef]
- Abraham, G.E. Radioimmunoassay of steroids. In Handbook of Radioimmunoassay; Abraham, G., Ed.; Dekker: New York, NY, USA, 1977. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Flohé, L.; Günzler, W.A. Assays of glutathione peroxidase. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 114–120. [Google Scholar]
- Luck, H. Catalase. In Method of Enzymatic Analysis; Bergmayer, M.V., Ed.; Verlag Chemic/Academic Press: New York, NY, USA, 1974; p. 885. [Google Scholar]
- Misra, H.; Fridovich, I. The role of superoxide anion in the autooxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Beutler, E.; Duron, O.; Kelly, B.M. An improved method for the detection of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar]
- Tappel, A.; Zalkin, H. Inhibition of lipide peroxidation in mitochondria by vitamin E. Arch. Biochem. Biophys. 1959, 80, 333–336. [Google Scholar] [CrossRef]
- Culling, C.F. Handbook of Histopathologic and Histochemical Staining, 3rd ed.; Buterworth: London, UK, 1983. [Google Scholar]
- Duncan, D. Multiple range and multiple F test. Biometric 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Aitken, R.J.; Roman, S.D. Antioxidant systems and oxidative stress in the testes. In Molecular Mechanisms in Spermatogenesis; Springer: New York, NY, USA, 2009; pp. 154–171. [Google Scholar]
- Huang, H.; Wang, Y.; An, Y.; Jiao, W.; Xu, Y.; Han, Q.; Teng, X.; Teng, X. Selenium alleviates oxidative stress and autophagy in lead-treated chicken testes. Theriogenology 2019, 131, 146–152. [Google Scholar] [CrossRef]
- Peña-Mendoza, B.; Gómez-Márquez, J.L.; Salgado-Ugarte, I.; Ramírez-Noguera, D. Reproductive biology of Oreochromis niloticus (Perciformes: Cichlidae) at Emiliano Zapata dam, Morelos, Mexico. Rev. Biol. Trop. 2005, 53, 515–522. [Google Scholar] [CrossRef] [Green Version]
- Kopalli, S.R.; Hwang, S.-Y.; Won, Y.-J.; Kim, S.-W.; Cha, K.-M.; Han, C.-K.; Hong, J.-Y.; Kim, S.-K. Korean red ginseng extract rejuvenates testicular ineffectiveness and sperm maturation process in aged rats by regulating redox proteins and oxidative defense mechanisms. Exp. Gerontol. 2015, 69, 94–102. [Google Scholar] [CrossRef]
- Ramesh, T.; Kim, S.-W.; Sung, J.-H.; Hwang, S.-Y.; Sohn, S.-H.; Yoo, S.-K.; Kim, S.-K. Effect of fermented Panax ginseng extract (GINST) on oxidative stress and antioxidant activities in major organs of aged rats. Exp. Gerontol. 2012, 47, 77–84. [Google Scholar] [CrossRef]
- Sun, Z.; Tan, X.; Ye, H.; Zou, C.; Ye, C.; Wang, A. Effects of dietary Panax notoginseng extract on growth performance, fish composition, immune responses, intestinal histology and immune related genes expression of hybrid grouper (Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀) fed high lipid diets. Fish Shellfish. Immunol. 2018, 73, 234–244. [Google Scholar] [PubMed]
- Ok, S.; Kang, J.S.; Kim, K.M. Testicular antioxidant mechanism of cultivated wild ginseng extracts. Mol. Cell. Toxicol. 2016, 12, 149–158. [Google Scholar] [CrossRef]
- Usha, R.; Pugazhendy, K.; Tamizhazhagan, V.; Sakthidasan, V.; Jayanthi, C. Potential efficacy of tribulus terrtri aginst toxic impact of chlorpyrifos on haematological alteration in the fresh water fish Oreochrommis mossambicus. Int. J. Zool. Appl. Biosci. 2017, 2, 232–240. [Google Scholar]
- Li, Z.; Li, M.; Liu, S.; Che, L.; Jiang, X.; Guo, R. Effects of Tribulus terrestris extract on growth performance and serum antioxidant indexes of weaned piglets. Chin. J. Anim. Nutr. 2018, 30, 3662–3669. [Google Scholar]
- Ramnath, V.; Rekha, P.; Sujatha, K. Amelioration of heat stress induced disturbances of antioxidant defense system in chicken by Brahma Rasayana. Evid.-Based Complement. Altern. Med. 2008, 5, 77–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dakshayini, P.; Mahaboob Basha, P. Phytochemical screening and in vitro antioxidant potential of Tribulus terrestris fruit and Mesua ferrea flower extracts: A comparative study. Int. J. Pharm. Pharm. Sci. 2018, 10, 70–75. [Google Scholar]
- Mohamed, N.A.; Ahmed, O.M.; Hozayen, W.G.; Ahmed, M.A. Ameliorative effects of bee pollen and date palm pollen on the glycemic state and male sexual dysfunctions in streptozotocin-induced diabetic wistar rats. Biomed. Pharmacother. 2018, 97, 9–18. [Google Scholar] [CrossRef]
- Hassan, W.A.; El-kashlan, A.M.; Mohamed, N.A. Egyptian date palm pollen ameliorates testicular dysfunction induced by cadmium chloride in adult male rats. J. Am. Sci. 2012, 8, 659–669. [Google Scholar]
- El-Neweshy, M.; El-Maddawy, Z.; El-Sayed, Y. Therapeutic effects of date palm (Phoenix dactylifera L.) pollen extract on cadmium-induced testicular toxicity. Andrologia 2013, 45, 369–378. [Google Scholar] [CrossRef]
- Khider, M.; Elbanna, K.; Mahmoud, A.; Owayss, A.A. Egyptian honeybee pollen as antimicrobial, antioxidant agents, and dietary food supplements. Food Sci. Biotechnol. 2013, 22, 1–9. [Google Scholar] [CrossRef]
- Shiva, M.; Gautam, A.K.; Verma, Y.; Shivgotra, V.; Doshi, H.; Kumar, S. Association between sperm quality, oxidative stress, and seminal antioxidant activity. Clin. Biochem. 2011, 44, 319–324. [Google Scholar] [CrossRef]
- Yun, S.J.; Bae, G.-S.; Park, J.H.; Song, T.H.; Choi, A.; Ryu, B.-Y.; Pang, M.-G.; Kim, E.J.; Yoon, M.; Chang, M.B. Antioxidant effects of cultured wild ginseng root extracts on the male reproductive function of boars and guinea pigs. Anim. Reprod. Sci. 2016, 170, 51–60. [Google Scholar] [CrossRef]
- Tsai, S.-C.; Chiao, Y.-C.; Lu, C.-C.; Wang, P.S. Stimulation of the secretion of luteinizing hormone by ginsenoside-Rb1 in male rats. Chin. J. Physiol. 2003, 46, 1–7. [Google Scholar]
- Antonio, J.; Uelmen, J.; Rodriguez, R.; Earnest, C. The effects of Tribulus terrestris on body composition and exercise performance in resistance-trained males. Int. J. Sport Nutr. Exerc. Metab. 2000, 10, 208–215. [Google Scholar] [CrossRef]
- Kim, I.-H.; Kim, S.-K.; Kim, E.-H.; Kim, S.-W.; Sohn, S.-H.; Lee, S.C.; Choi, S.; Pyo, S.; Rhee, D.-K. Korean red ginseng up-regulates C21-steroid hormone metabolism via Cyp11a1 gene in senescent rat testes. J. Ginseng. Res. 2011, 35, 272. [Google Scholar] [CrossRef] [Green Version]
- Selmani, C.; Chabane, D.; Bouguedoura, N. Ethnobotanical survey of Phoenix dactylifera L. pollen used for the treatment of infertility problems in Algerian oases. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Iftikhar, S.; Bashir, A.; Anwar, M.S.; Mastoi, S.M.; Shahzad, M. Effect of date palm pollen (DPP) on serum testosterone levels in prepubertal albino rats. Pak. J. Med. Health Sci. 2011, 6, 639–644. [Google Scholar]
- Abbass, A.A.; El-Asely, A.M.; Kandiel, M.M. Effects of dietary propolis and pollen on growth performance, fecundity and some hematological parameters of Oreochromis niloticus. Turk. J. Fish. Aquat. Sci. 2012, 12, 851–859. [Google Scholar]
- Sharma, P.; Huq, A.U.; Singh, R. Cypermethrin induced reproductive toxicity in male Wistar rats: Protective role of Tribulus terrestris. J. Environ. Biol. 2013, 34, 857. [Google Scholar]
- Mahmoud, O.M.; Al Badawi, M.H.; Salem, N.A. Role of Ginseng on mercury chloride-induced testicular lesions in adult albino rat: A histological and immunohistochemical study. Egypt. J. Histol. 2014, 37, 506–513. [Google Scholar] [CrossRef]
- El-Tantawy, W.H.; Temraz, A.; El-Gindi, O.D. Free serum testosterone level in male rats treated with Tribulus alatus extracts. Int. Braz. J. Urol. 2007, 33, 554–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.Z.; Luo, L. Ginseng on hyperglycemia: Effects and mechanisms. Evid.-Based Complement. Altern. Med. 2009, 6, 423–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cek, S.; Turan, F.; Atik, E. The effects of Gokshura, Tribulus terrestris on sex reversal of guppy, Poecilia reticulata. Pak. J. Biol. Sci. 2007, 10, 718. [Google Scholar] [CrossRef] [PubMed]
- Babahajiani, P.; Shokrollahi, B.; Gharibkhani, M. The effect of gokshura (Tribulus terrestris) and nettle root (Urtica dioica) extracts on growth rate and sex reversal in convict cichlid (Cichlasoma nigrofasciatum). Iran. J. Fish. Sci. 2018, 17, 620–628. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | g kg−1 |
|---|---|
| Fish meal (herring; 72%) | 260 |
| Corn gluten | 150 |
| Yellow corn | 390 |
| Rice bran | 100 |
| Wheat flour | 50 |
| Corn oil | 24 |
| Vitamins and Minerals primex 1 | 20 |
| Calcium-mono phosphate | 6 |
| Proximate chemical composition | |
| Dry matter (DM) | 922.00 |
| Crude protein (CP) | 323.80 |
| Ether extract (EE) | 72.00 |
| Nitrogen-free extract (NFE) 2 | 489.80 |
| Crude fiber (CF) | 46.10 |
| Ash | 68.30 |
| Gross energy (GE; kj g−1 DM) 3 | 18.90 |
| P/E ratio (mg CP: kj) 4 | 17.13 |
| Items | Luteinizing Hormone (IU/L) | Testosterone (ng/mL) [30] | Testes Somatic Index (%) [30] |
|---|---|---|---|
| Control | 0.43 ± 0.02 b | 4.15 ± 0.13 d | 0.83 ± 0.03 c |
| GE0.2 | 0.48 ± 0.03 a,b | 5.14 ± 0.26 c | 1.01 ± 0.03 a,b,c |
| GE0.4 | 0.50 ± 0.02 a | 6.83 ± 0.30 b | 1.12 ± 0.04 a,b |
| TT0.6 | 0.47 ± 0.02 a,b | 6.53 ± 0.18 b | 1.02 ± 0.04 a,b,c |
| TT1.2 | 0.53 ± 0.01 a | 7.73 ± 0.16 a | 1.18 ± 0.05 a |
| DPPG3.0 | 0.49 ± 0.01 a,b | 4.83 ± 0.03 c | 0.96 ± 0.10 b,c |
| DPPG6.0 | 0.51 ± 0.03 a | 5.17 ± 0.28 c | 1.04 ± 0.12 a,b |
| Items | Thiobarbituric Acid-Reactive Substances (nmol/mL) | Reduced Glutathione (µmol/mL) | Superoxide Dismutase (U/mL) | Catalase (U/mL) | Glutathione S-Transferase (µmol/h) | Glutathione Peroxidase (U/mL) |
|---|---|---|---|---|---|---|
| Control | 49.97 ± 1.05 a | 3.10 ± 0.09 d | 8.92 ± 0.33 b | 20.22 ± 0.79 d | 3.50 ± 0.18 b | 5.96 ± 0.14 c |
| GE 0.2 | 45.44 ± 1.50 b | 3.48 ± 0.06 b,c | 10.83 ± 0.32 a,b | 24.50 ± 0.84 b | 4.21 ± 0.12 a | 8.23 ± 0.20 a |
| GE 0.4 | 42.56 ± 0.47 b,c | 3.92 ± 0.08 a | 12.44 ± 1.37 a | 29.00 ± 0.68 a | 4.55 ± 0.16 a | 8.70 ± 0.19 a |
| TT 0.6 | 43.94 ± 0.74 b | 3.41 ± 0.10 c | 11.59 ± 0.37 a | 24.00 ± 0.26 b,c | 4.26 ± 0.16 a | 7.97 ± 0.39 a,b |
| TT 1.2 | 40.34 ± 0.59 c | 3.76 ± 0.07 a | 12.80 ± 0.74 a | 24.67 ± 0.15 b | 4.67 ± 0.13 a | 8.71 ± 0.25 a |
| DPPG 3 | 45.29 ± 0.82 b | 3.45 ± 0.09 c | 10.69 ± 0.41 a,b | 22.11 ± 0.37 c,d | 4.17 ± 0.18 b | 7.33 ± 0.15 b |
| DPPG 6 | 42.75 ± 0.86 b,c | 3.70 ± 0.08 a,b | 12.00 ± 0.24 a | 23.40 ± 1.36 b,c | 4.21 ± 0.07 a | 7.95 ± 0.31 a,b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mansour, A.T.; Alsaqufi, A.S.; Omar, E.A.; El-Beltagi, H.S.; Srour, T.M.; Yousef, M.I. Ginseng, Tribulus Extracts and Pollen Grains Supplementation Improves Sexual State, Testes Redox Status, and Testicular Histology in Nile Tilapia Males. Antioxidants 2022, 11, 875. https://doi.org/10.3390/antiox11050875
Mansour AT, Alsaqufi AS, Omar EA, El-Beltagi HS, Srour TM, Yousef MI. Ginseng, Tribulus Extracts and Pollen Grains Supplementation Improves Sexual State, Testes Redox Status, and Testicular Histology in Nile Tilapia Males. Antioxidants. 2022; 11(5):875. https://doi.org/10.3390/antiox11050875
Chicago/Turabian StyleMansour, Abdallah Tageldein, Ahmed Saud Alsaqufi, Eglal Ali Omar, Hossam S. El-Beltagi, Tarek Mohamed Srour, and Mokhtar Ibrahim Yousef. 2022. "Ginseng, Tribulus Extracts and Pollen Grains Supplementation Improves Sexual State, Testes Redox Status, and Testicular Histology in Nile Tilapia Males" Antioxidants 11, no. 5: 875. https://doi.org/10.3390/antiox11050875