
Low-Intensity Blue Light Supplemented during Photoperiod in Controlled Environment Induces Flowering and Antioxidant Production in Kalanchoe




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
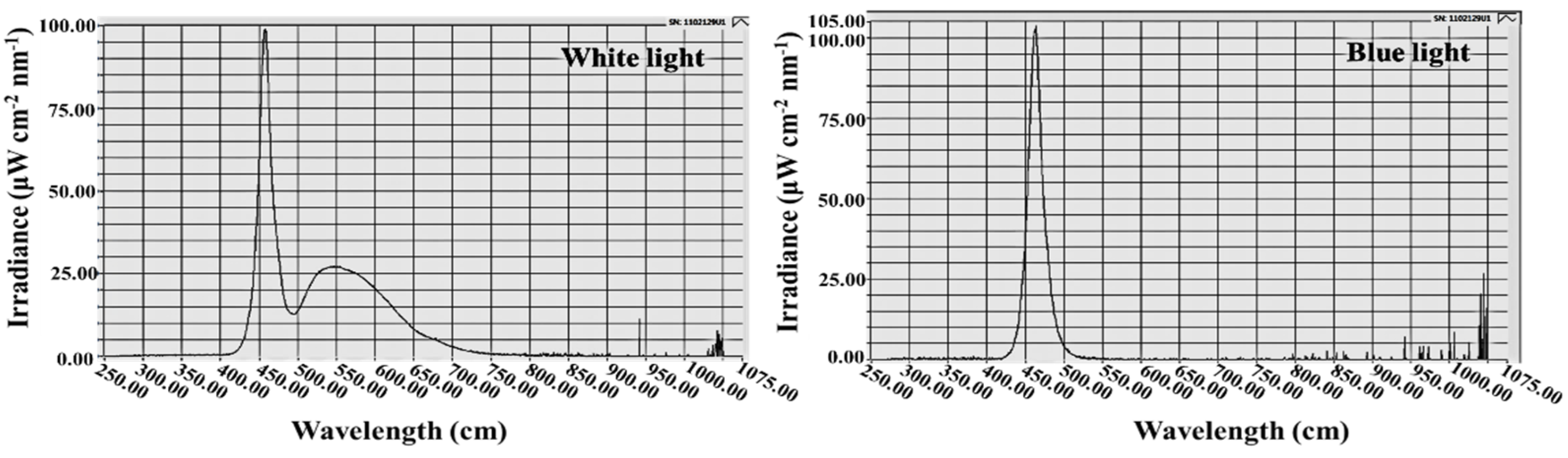
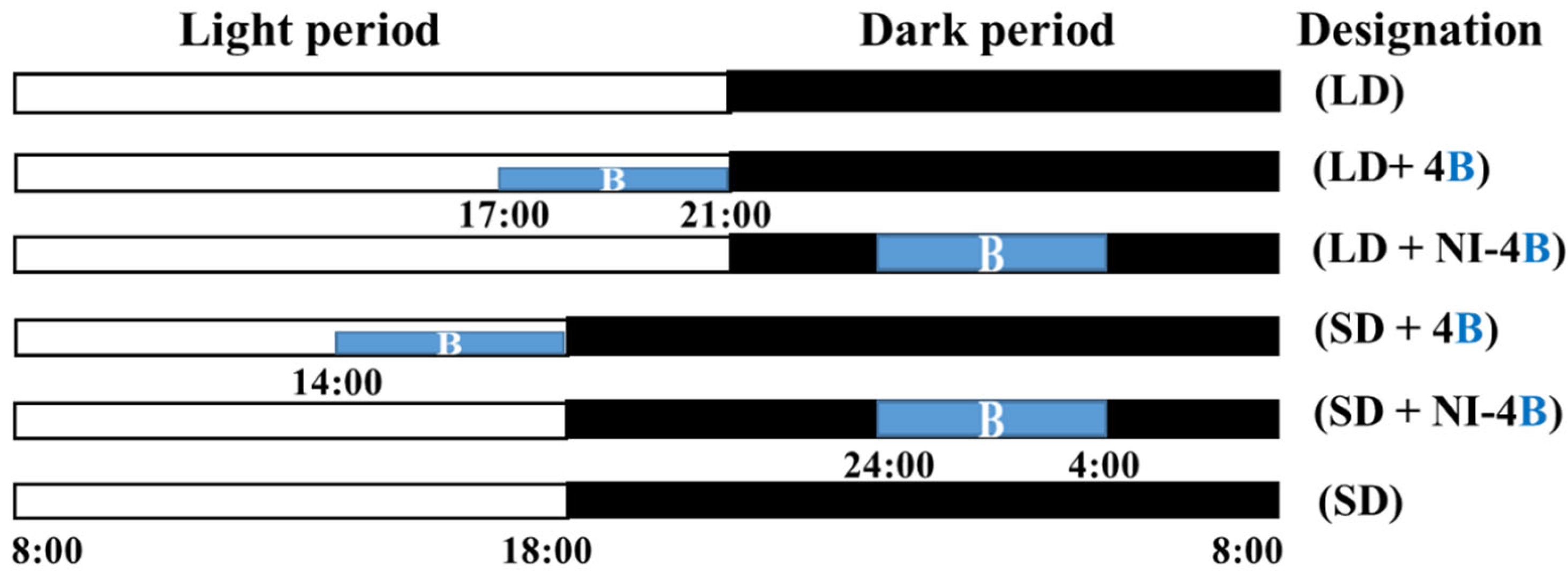
2.2. Photoperiodic Light Treatments
2.3. Measurements of the Morphological Parameters
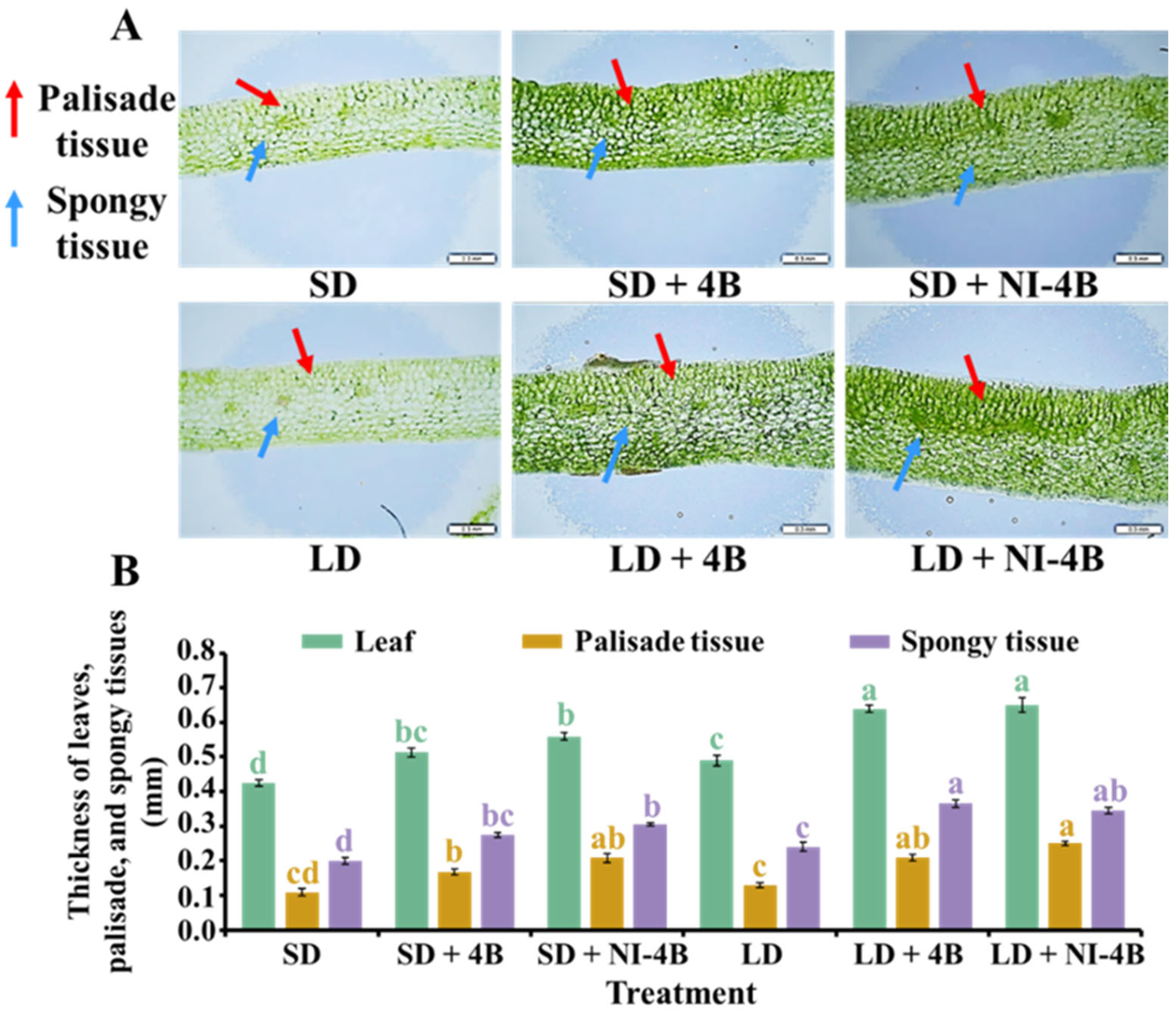
2.4. Leaf Anatomical Features and Chloroplast Distribution
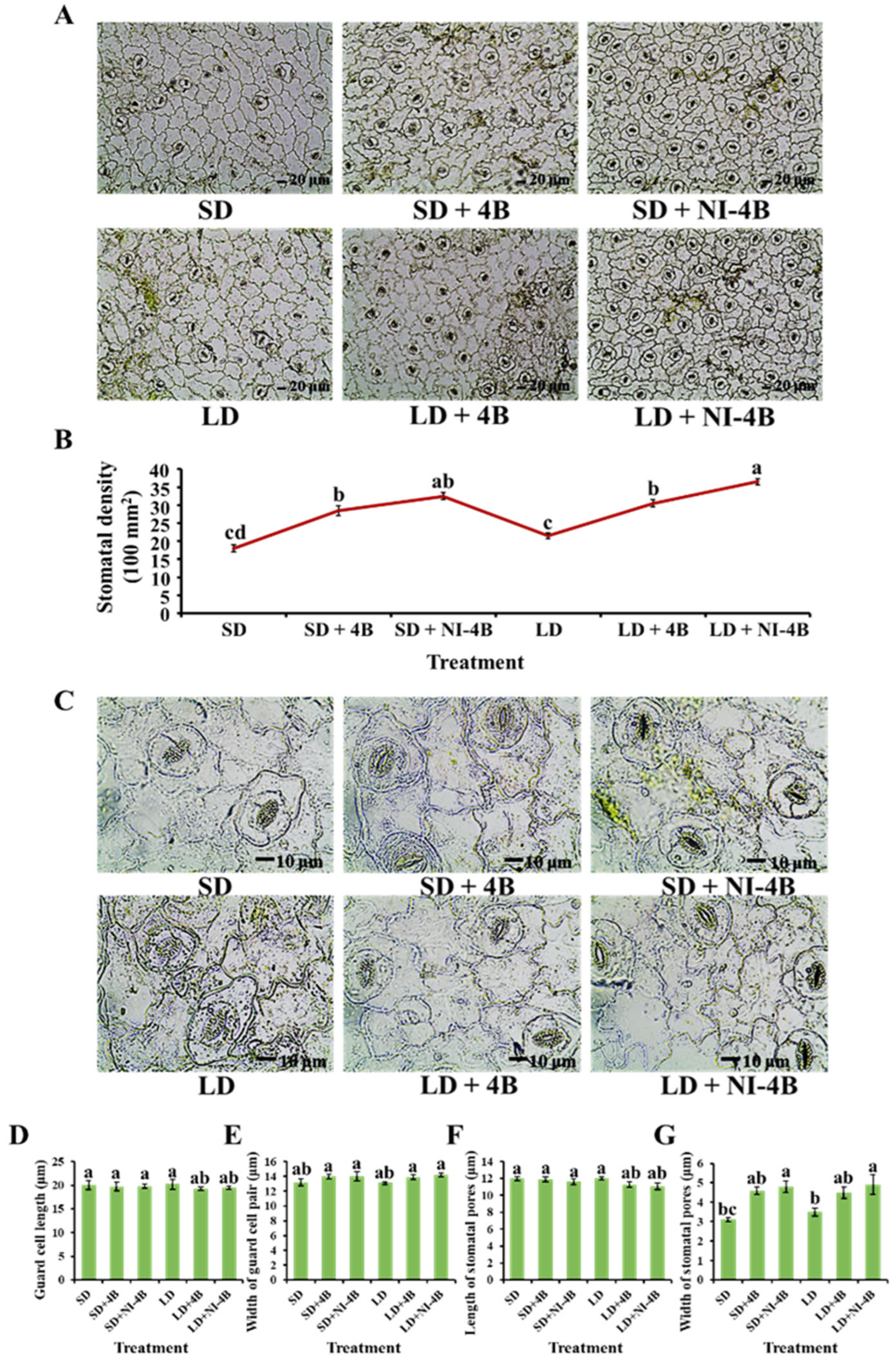
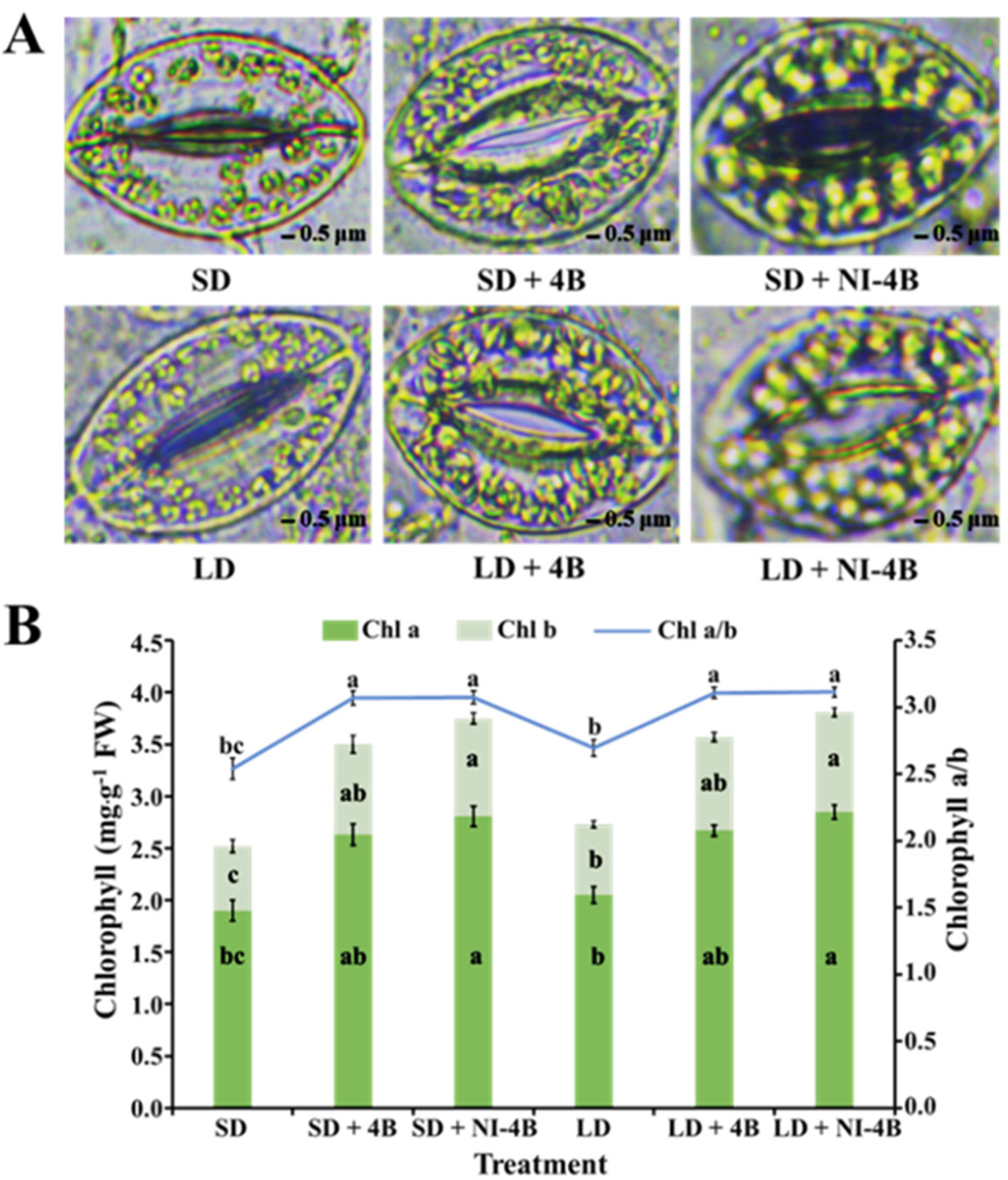
2.5. Stomatal Density and Morphological Characteristics
2.6. Chlorophyll Content
2.7. Measurements of Photosynthesis and Chlorophyll Fluorescence
2.8. Accumulation of Carbohydrates and Soluble Proteins
2.9. Enzyme Activities
2.10. Real-Time Quantitative PCR Verification
2.11. Statistical Analysis
3. Results
3.1. Morphological Analyses
3.2. Anatomical Features of Leaves
3.3. Morphological Characteristics of the Stomata
3.4. Chloroplast Distribution and Chlorophyll Content
3.5. Photosynthetic and Chlorophyll Fluorescence Characteristics
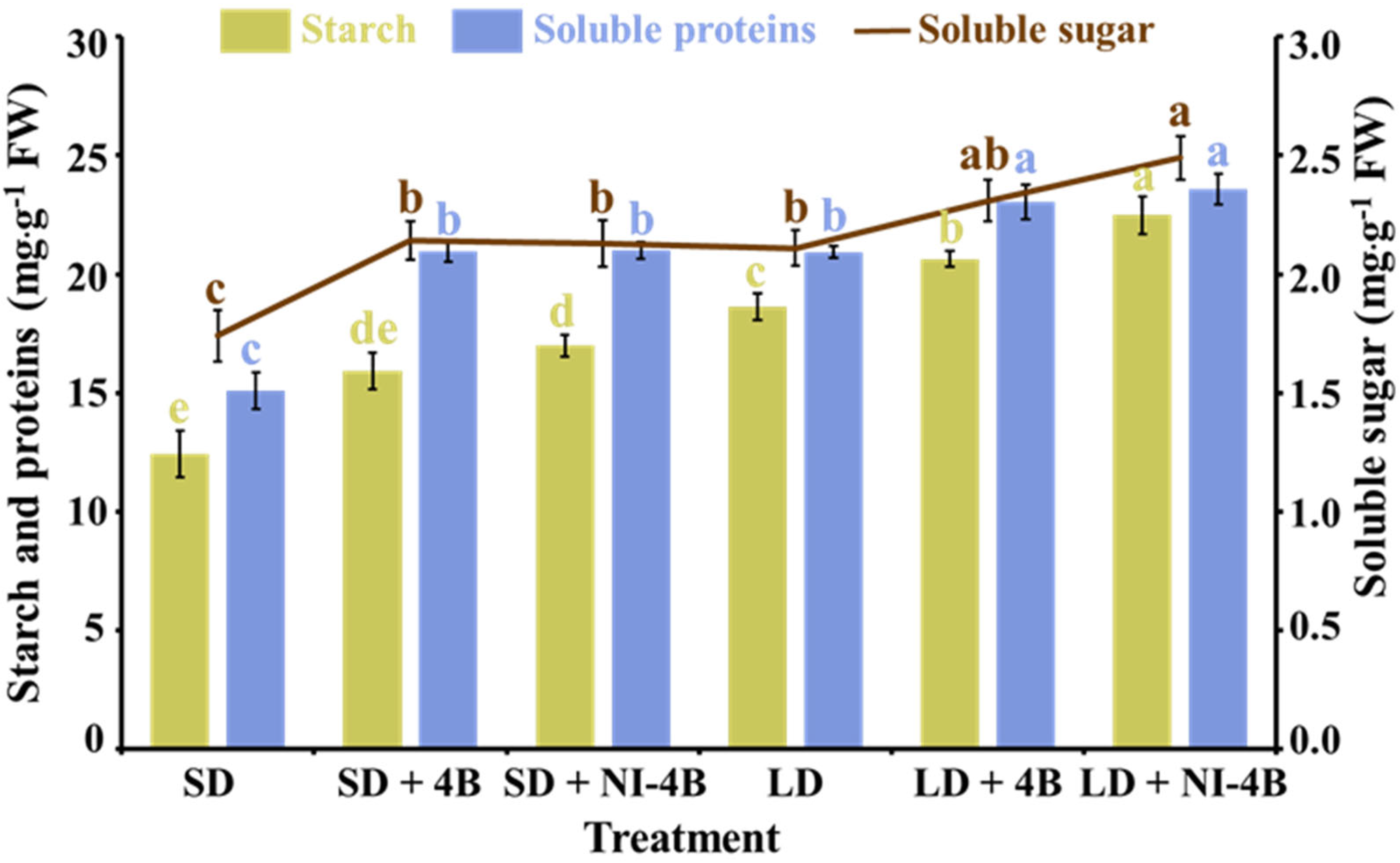
3.6. Accumulation of Carbohydrates and Soluble Proteins
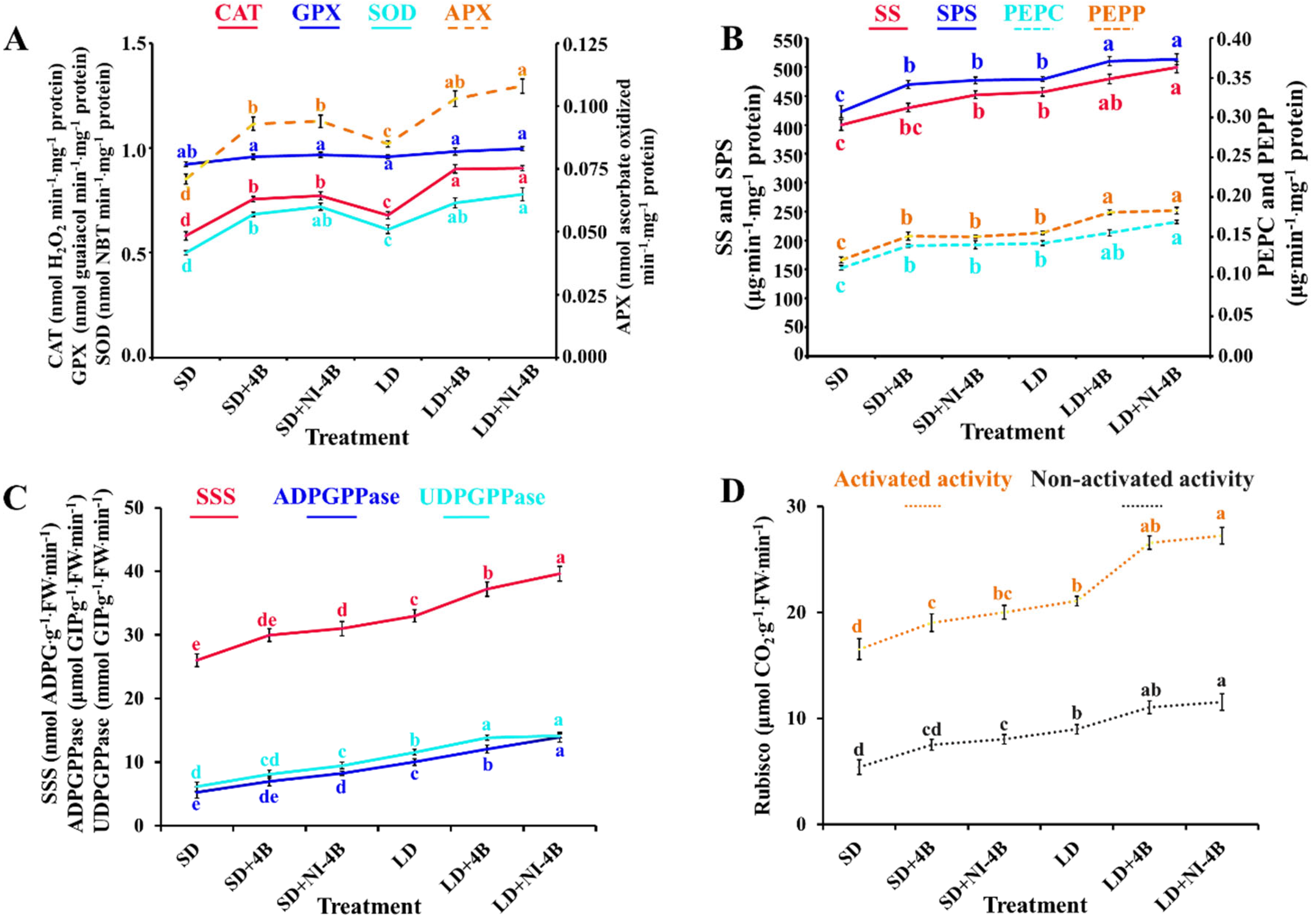
3.7. Enzymatic Activities
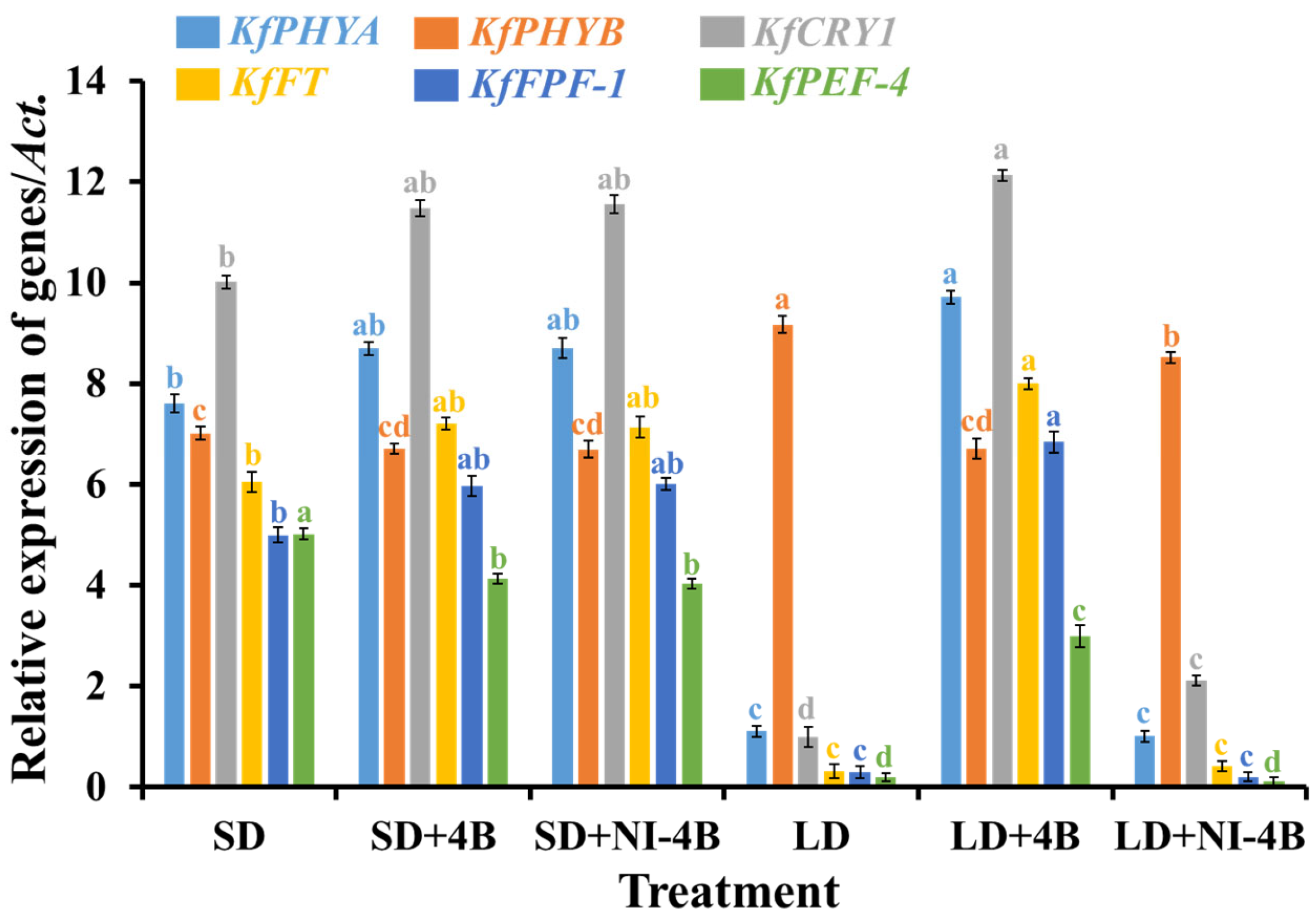
3.8. Gene Expressions
4. Discussion
4.1. Growth and Morphology of Kalanchoes Were Influenced by Low-Intensity Supplementary and Night-Interrupting Blue Light in the Photoperiodic Light Treatments
4.1.1. Low-Intensity Supplementary and Night-Interrupting Blue Light in Photoperiodic Light Treatments Synthetically Affected the Development of Plant Shoots
4.1.2. Low-Intensity Supplementary and Night-Interrupting Blue Light in Photoperiodic Light Treatments Increased the Leaf Number and Decreased the Leaf Size
4.1.3. Low-Intensity Supplementary and Night-Interrupting Blue Light in Photoperiodic Light Treatments Affected the Flower Color, Delayed the Formation of Flower Buds, and Improved the Number of Inflorescences by Controlling Flowering-Related Genes
4.2. Photosynthesis, Physiology, and Internal Structures of Kalanchoes Were Influenced by Low-Intensity Supplementary and Night-Interrupting Blue Light in Photoperiodic Light Treatments
4.2.1. Low-Intensity Supplementary and Night-Interrupting Blue Light in Photoperiodic Light Treatments Enhanced Leaf Structures, Chloroplast Distribution, and Chlorophyll Accumulation in Kalanchoes
4.2.2. Low-Intensity Supplementary and Night-Interrupting Blue Light in Photoperiodic Light Treatments Induced Excellent Stomatal, Photosynthetic, and Chlorophyll Fluorescence Characteristics
4.2.3. Low-Intensity Supplementary and Night-Interrupting Blue Light in Photoperiodic Light Treatments Directed the Accumulation of Carbohydrates and Proteins in Kalanchoes
4.2.4. Low-Intensity Supplementary and Night-Interrupting Blue Light in Photoperiodic Light Treatments Observably Activated the Enzymatic Activities in Kalanchoes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Demotes-Mainard, S.; Péron, T.; Corot, A.; Bertheloot, J.; Le Gourrierec, J.; Pelleschi-Travier, S.; Crespel, L.; Morel, P.; Huché-Thélier, L.; Boumaza, R.; et al. Plant responses to red and far-red lights, applications in horticulture. Environ. Exp. Bot. 2016, 121, 4–21. [Google Scholar] [CrossRef]
- Haliapas, S.; Yupsanis, T.A.; Syros, T.D.; Kofidis, G.; Economou, A.S. Petunia × hybrida during transition to flowering as affected by light intensity and quality treatments. Acta Physiol. Plant. 2008, 30, 807–815. [Google Scholar] [CrossRef]
- Fan, X.X.; Xu, Z.G.; Liu, X.Y.; Tang, C.M.; Wang, L.W.; Han, X.L. Effects of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light. Sci. Hortic. 2013, 153, 50–55. [Google Scholar] [CrossRef]
- Kami, C.; Lorrain, S.; Hornitschek, P.; Fankhauser, C. Light-regulated plant growth and development. Curr. Top. Dev. Biol. 2010, 91, 29–66. [Google Scholar]
- Galvão, V.C.; Fankhauser, C. Sensing the light environment in plants: Photoreceptors and early signaling steps. Curr. Opin. Neurobiol. 2015, 34, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Huché-Thélier, L.; Crespel, L.; Le Gourrierec, J.; Morel, P.; Sakr, S.; Leduc, N. Light signaling and plant responses to blue and UV radiations—Perspectives for applications in horticulture. Environ. Exp. Bot. 2016, 121, 22–38. [Google Scholar] [CrossRef]
- Cashmore, A.R.; Jarillo, J.A.; Wu, Y.J.; Liu, D. Cryptochromes: Blue light receptors for plants and animals. Science 1999, 284, 760–765. [Google Scholar] [CrossRef]
- Clack, T.; Mathews, S.; Sharrock, R.A. The phytochrome apoprotein family in Arabidopsis is encoded by five genes: The se-quences and expression of PHYD and PHYE. Plant Mol. Biol. 1994, 25, 413–427. [Google Scholar] [CrossRef]
- Sharrock, R.A.; Quail, P.H. Novel phytochrome sequences in Arabidopsis thaliana: Structure, evolution, and differential expression of a plant regulatory photoreceptor family. Genes Dev. 1989, 3, 1745–1757. [Google Scholar] [CrossRef] [Green Version]
- Runkle, E.; Heins, R. Manipulating the light environment to control flowering and morphogenesis of herbaceous plants. Acta Hortic. 2006, 711, 51–60. [Google Scholar] [CrossRef]
- Zheng, Q.; Weng, Q.; Huang, L.; Wang, K.; Deng, J.; Jiang, R.; Ye, Z.; Gan, M. A new source of multi-spectral high spatial resolution night-time light imagery—JL1-3B. Remote Sens. Environ. 2018, 215, 300–312. [Google Scholar] [CrossRef]
- Yamada, A.; Tanigawa, T.; Suyama, T.; Matsuno, T.; Kunitake, T. Night break treatment using different light sources promotes or delays growth and flowering of Eustoma grandiflorum (Raf.) Shinn. J. Japan. Soc. Hortic. Sci. 2008, 77, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.G.; Muneer, S.; Jeong, B.R. Morphogenesis, flowering, and gene expression of Dendranthema grandiflorum in response to shift in light quality of night interruption. Int. J. Mol. Sci. 2015, 16, 16497–16513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamamoto, H.; Shimaji, H.; Higashide, T. Budding response of horticultural crops to night break with red light on alternate days. Environ. Control Biol. 2005, 43, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Adams, S.; Valdes, V.; Langton, F. Why does low intensity, long-day lighting promote growth in Petunia, Impatiens, and tomato? J. Hortic. Sci. Biotechnol. 2008, 83, 609–615. [Google Scholar] [CrossRef]
- Kim, Y.J.; Lee, H.J.; Kim, K.S. Night interruption promotes vegetative growth and flowering of Cymbidium. Sci. Hortic. 2011, 130, 887–893. [Google Scholar] [CrossRef]
- Hogewoning, S.; Trouwborst, G.; Maljaars, H.; Poorter, H.; Van Ieperen, W.; Harbinson, J. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef] [PubMed]
- Appelgren, M. Effects of light quality on stem elongation of Pelargonium in vitro. Sci. Hortic. 1991, 45, 345–351. [Google Scholar] [CrossRef]
- Dougher, T.A.; Bugbee, B. Long-term blue light effects on the histology of lettuce and soybean leaves and stems. J. Am. Soc. Hortic. Sci. 2004, 129, 467–472. [Google Scholar] [CrossRef] [Green Version]
- Senger, H. The effect of blue light on plants and microorganisms. Photochem. Photobiol. 1982, 35, 911–920. [Google Scholar] [CrossRef]
- Fankhauser, C.; Ulm, R. Light-regulated interactions with SPA proteins underlie cryptochrome-mediated gene expression. Genes Dev. 2011, 25, 1004–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirose, F.; Shinomura, T.; Tanabata, T.; Shimada, H.; Takano, M. Involvement of rice cryptochromes in de-etiolation responses and flowering. Plant Cell Physiol. 2006, 47, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.W.; Hogewoning, S.; van Ieperen, W. Responses of supplemental blue light on flowering and stem extension growth of cut chrysanthemum. Sci. Hortic. 2014, 165, 69–74. [Google Scholar] [CrossRef]
- Higuchi, Y.; Sumitomo, K.; Oda, A.; Shimizu, H.; Hisamatsu, T. Day light quality affects the night-break response in the short-day plant chrysanthemum, suggesting differential phytochrome-mediated regulation of flowering. J. Plant Physiol. 2012, 169, 1789–1796. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.G.; Jeong, B.R. Night interruption light quality changes morphogenesis, flowering, and gene expression in Dendranthema grandiflorum. Hortic. Environ. Biotechnol. 2019, 60, 167–173. [Google Scholar] [CrossRef]
- Park, Y.G.; Jeong, B.R. How supplementary or night-interrupting low-intensity blue light affects the flower induction in chry-santhemum, a qualitative short-day plant. Plants 2020, 9, 1694. [Google Scholar] [CrossRef]
- Kang, D.I.; Jeong, H.K.; Park, Y.G.; Jeong, B.R. Flowering and morphogenesis of kalanchoe in response to quality and intensity of night interruption light. Plants 2019, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Cope, K.R.; Bugbee, B. Spectral effects of three types of white light-emitting diodes on plant growth and development: Absolute versus relative amounts of blue light. HortScience 2013, 48, 504–509. [Google Scholar] [CrossRef]
- Nanya, K.; Ishigami, Y.; Hikosaka, S.; Goto, E. Effects of blue and red light on stem elongation and flowering of tomato seedlings. Acta Hortic. 2012, 261–266. [Google Scholar] [CrossRef]
- Prochazkova, D.; Sairam, R.; Srivastava, G.; Singh, D. Oxidative stress and antioxidant activity as the basis of senescence in maize leaves. Plant Sci. 2001, 161, 765–771. [Google Scholar] [CrossRef]
- Mastropasqua, L.; Borraccino, G.; Bianco, L.; Paciolla, C. Light qualities and dose influence ascorbate pool size in detached oat leaves. Plant Sci. 2012, 183, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, L.B.; Leal-Costa, M.V.; Coutinho, M.A.; Moreira, N.D.S.; Lage, C.L.; Barbi, N.D.S.; Costa, S.S.; Tavares, E.S. In-creased antioxidant activity and changes in phenolic profile of Kalanchoe pinnata (Lamarck) persoon (Crassulaceae) specimens grown under supplemental blue light. Photochem. Photobiol. 2013, 89, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Kim, J.H.; Jeong, S.M.; Kim, D.R.; Ha, J.U.; Nam, K.C.; Ahn, D.U. Effect of far-infrared radiation on the antioxidant activity of rice hulls. J. Agric. Food Chem. 2003, 51, 4400–4403. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.C.; Hou, C.Y.; Jiang, C.M.; Wang, Y.T.; Wang, C.Y.; Chen, H.H.; Chang, H.M. A novel approach of LED light radiation improves the antioxidant activity of pea seedlings. Food Chem. 2007, 101, 1753–1758. [Google Scholar] [CrossRef]
- Fan, Y.; Chen, J.; Cheng, Y.; Raza, M.A.; Wu, X.; Wang, Z.; Liu, Q.; Wang, R.; Wang, X.; Yong, T.; et al. Effect of shading and light recovery on the growth, leaf structure, and photosynthetic performance of soybean in a maize-soybean relay-strip intercropping system. PLoS ONE 2018, 13, e0198159. [Google Scholar] [CrossRef] [PubMed]
- Sack, L.; Buckley, T.N. The developmental basis of stomatal density and flux. Plant Physiol. 2016, 171, 2358–2363. [Google Scholar] [CrossRef] [Green Version]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N.; Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence-a practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Vasseur, F.; Pantin, F.; Vile, D. Changes in light intensity reveal a major role for carbon balance in Arabidopsis responses to high temperature. Plant Cell Environ. 2011, 34, 1563–1576. [Google Scholar] [CrossRef]
- Ren, X.X.; Xue, J.Q.; Wang, S.; Xue, Y.Q.; Zhang, P.; Jiang, H.D.; Zhang, X.X. Proteomic analysis of tree peony (Paeonia ostii ‘Feng Dan’) seed germination affected by low temperature. J. Plant Physiol. 2017, 224, 56–67. [Google Scholar] [CrossRef]
- Song, J.; Li, Y.; Hu, J.; Lee, J.; Jeong, B.R. Pre- and/or postharvest silicon application prolongs the vase life and enhances the quality of cut peony (Paeonia lactiflora Pall.) flowers. Plants 2021, 10, 1742. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Soundararajan, P.; Jeong, B.R. Proteomic and antioxidant analysis elucidates the underlying mechanism of tolerance to hyperhydricity stress in in vitro shoot cultures of Dianthus caryophyllus. J. Plant Growth Regul. 2016, 35, 667–679. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Aeibi, H.; Bergmeyer, H. Methods in Enzymatic Analysis; Academic Press: New York, NY, USA, 1974; Volume 3, p. 673. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Castillo, F.J.; Penel, C.; Greppin, H. Peroxidase release induced by ozone in sedum album leaves: Involvement of Ca2+. Plant Physiol. 1984, 74, 846–851. [Google Scholar] [CrossRef] [Green Version]
- Becana, M.; Tejo, P.A.; Irigoyen, J.J.; Sa’nchez-Dı’az, M. Some enzymes of hydrogen peroxide metabolism in leaves and root nodules of Medicago sativa. Plant Physiol. 1986, 82, 1169–1171. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Raza, M.A.; Li, Z.; Chen, Y.; Bin Khalid, M.H.; Du, J.; Liu, W.; Wu, X.; Song, C.; Yu, L.; et al. The influence of light intensity and leaf movement on photosynthesis characteristics and carbon balance of soybean. Front. Plant Sci. 2019, 9, e01952. [Google Scholar] [CrossRef]
- Yang, L.T.; Chen, L.S.; Peng, H.Y.; Guo, P.; Wang, P.; Ma, C.L. Organic acid metabolism in Citrus grandis leaves and roots is differently affected by nitric oxide and aluminum interactions. Sci. Hortic. 2012, 133, 40–46. [Google Scholar] [CrossRef]
- Doehlert, D.C.; Kuo, T.M.; Felker, F.C. Enzymes of sucrose and hexose metabolism in developing kernels of two inbreds of maize. Plant Physiol. 1988, 86, 1013–1019. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.S.; Cao, X.; Xu, S.; Zhu, Q.; Song, P. Studies on the relationship between the grain sink strength and its starch accu-mulation in rice (O. sativa). Acta Agron. Sin. 1994, 20, 685–691. [Google Scholar]
- Yu, X.; Liu, H.; Klejnot, J.; Lin, C. The cryptochrome blue light receptors. Arab. Book 2010, 8, e0135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, B. Specific effects of blue light on plant growth and development. Plants Daylight Spectr. 1981, 1981, 443–459. [Google Scholar]
- Xu, D.; Gao, W.; Ruan, J. Effects of light quality on plant growth and development. Plant Physiol. J. 2015, 51, 1217–1234. [Google Scholar]
- Martínez-García, J.F.; García-Martínez, J.L. Interaction of gibberellins and phytochrome in the control of cowpea epicotyl elongation. Physiol. Plant. 1992, 86, 236–244. [Google Scholar] [CrossRef]
- Folta, K.M.; Pontin, M.A.; Karlin-Neumann, G.; Bottini, R.; Spalding, E.P. Genomic and physiological studies of early crypto-chrome 1 action demonstrate roles for auxin and gibberellin in the control of hypocotyl growth by blue light. Plant J. 2003, 36, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, F.; Mao, J.F.; Wang, J.; Zhang, S.; Li, Y. Transcriptome analysis reveals that red and blue light regulate growth and phytohormone metabolism in norway spruce [Picea abies (L.) Karst.]. PLoS ONE 2015, 10, e0127896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volmaro, C.; Pontin, M.; Luna, V.; Baraldi, R.; Bottini, R. Blue light control of hypocotyl elongation in etiolated seedlings of Lactuca sativa (L.) cv. grand rapids related to exogenous growth regulators and endogenous IAA, GA3 and abscisic acid. Plant Growth Regul. 1998, 26, 165–173. [Google Scholar] [CrossRef]
- Lin, K.H.; Huang, M.Y.; Huang, W.D.; Hsu, M.H.; Yang, Z.W.; Yang, C.M. The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Khattak, A.M.; Pearson, S. Spectral filters and temperature effects on the growth and development of chrysanthemums under low light integral. Plant Growth Regul. 2006, 49, 61–68. [Google Scholar] [CrossRef]
- Xu, Y.L.; Gage, D.A.; Zeevaart, J. Gibberellins and stem growth in Arabidopsis thaliana (effects of photoperiod on expression of the GA4 and GA5 loci). Plant Physiol. 1997, 114, 1471–1476. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.Y.; Yu, X.H.; Liu, X.; Lin, C.T. Light regulation of gibberellins metabolism in seedling development. J. Integr. Plant Biol. 2007, 49, 21–27. [Google Scholar] [CrossRef]
- Huang, N.Z.; Zhao, Z.G.; Fu, C.M.; Tang, F.L.; Huang, Z.M. Effect of different wavelength lights on photosynthesis and growth of Siraitia grosvenorrii. Guang Xi Zhi Wu 2008, 28, 251–255. (In Chinese) [Google Scholar]
- Menard, C.; Dorais, M.; Hovi, T.; Gosselin, A. Developmental and physiological responses of tomato and cucumber to additional blue light. Acta Hortic. 2006, 291–296. [Google Scholar] [CrossRef]
- Gautier, H.; Varlet-Grancher, C.; Baudry, N. Effects of blue light on the vertical colonization of space by white clover and their consequences for dry matter distribution. Ann. Bot. 1997, 80, 665–671. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Guo, Y.P.; Zhang, S.L.; Dai, W.S.; Fu, Q.G. Effect of light quality on plant growth and fruiting of Toyonoka strawberry (Fragaria × ananassa) cultivar. J. Fruit Sci. 2006, 23, 818–824. (In Chinese) [Google Scholar]
- Kong, Y.; Schiestel, K.; Zheng, Y. Maximum elongation growth promoted as a shade-avoidance response by blue light is related to deactivated phytochrome: A comparison with red light in four microgreen species. Can. J. Plant Sci. 2020, 100, 314–326. [Google Scholar] [CrossRef]
- Mockler, T.; Yang, H.; Yu, X.; Parikh, D.; Cheng, Y.C.; Dolan, S.; Lin, C. Regulation of photoperiodic flowering by Arabidopsis photoreceptors. Proc. Natl. Acad. Sci. USA 2003, 100, 2140–2145. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.; Bradley, M.; Harberd, N.P.; Whitelam, G.C. Photoresponses of light-grown phyA mutants of Arabidopsis (phyto-chrome A is required for the perception of daylength extensions). Plant Physiol. 1994, 105, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Mockler, T.C.; Guo, H.; Yang, H.; Lin, D.C. Antagonistic actions of Arabidopsis cryptochromes and phytochrome B in the reg-ulation of floral induction. Development 1999, 126, 2073–2082. [Google Scholar] [CrossRef]
- Franklin, K.A.; Praekelt, U.; Stoddart, W.M.; Billingham, O.E.; Halliday, K.; Whitelam, G.C. Phytochromes B, D, and E act redundantly to control multiple physiological responses in Arabidopsis. Plant Physiol. 2003, 131, 1340–1346. [Google Scholar] [CrossRef] [Green Version]
- Shinomura, T.; Nagatani, A.; Hanzawa, H.; Kubota, M.; Watanabe, M.; Furuya, M. Action spectra for phytochrome A and B specific photoinduction of seed germination in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1996, 93, 8129–8133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usami, T.; Mochizuki, N.; Kondo, M.; Nishimura, M.; Nagatani, A. Cryptochromes and phytochromes synergistically regulate Arabidopsis root greening under blue light. Plant Cell Physiol. 2004, 45, 1798–1808. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, R.; Shinomura, T.; Takano, M.; Shimamoto, K. Phytochrome dependent quantitative control of Hd3a transcription is the basis of the night break effect in rice flowering. Genes Genet. Syst. 2009, 84, 179–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliday, K.J.; Salter, M.G.; Thingnaes, E.; Whitelam, G.C. Phytochrome control of flowering is temperature sensitive and cor-relates with expression of the floral integrator FT. Plant J. 2003, 33, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Cerdán, P.D.; Chory, J. Regulation of flowering time by light quality. Nature 2003, 423, 881–885. [Google Scholar] [CrossRef] [PubMed]
- Devlin, P.F.; Halliday, K.J.; Harberd, N.P.; Whitelam, G.C. The rosette habit of Arabidopsis thaliana is dependent upon phyto-chrome action: Novel phytochromes control internode elongation and flowering time. Plant J. 1996, 10, 1127–1134. [Google Scholar] [CrossRef]
- Sessa, G.; Carabelli, M.; Sassi, M.; Ciolfi, A.; Possenti, M.; Mittempergher, F.; Becker, J.; Morelli, G.; Ruberti, I. A dynamic balance between gene activation and repression regulates the shade avoidance response in Arabidopsis. Genes Dev. 2005, 19, 2811–2815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, S.; Allen, T.; Whitelam, G.C. Interaction between the light quality and flowering time pathways in Arabidopsis. Plant J. 2009, 60, 257–267. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Yu, X.; Michaels, S.D. Regulation of CONSTANS and FLOWERING LOCUS T expression in response to changing light quality. Plant Physiol. 2008, 148, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Wollenberg, A.C.; Strasser, B.; Cerdán, P.D.; Amasino, R.M. Acceleration of flowering during shade avoidance in Arabidopsis alters the balance between FLOWERING LOCUS C-mediated repression and photoperiodic induction of flowering. Plant Physiol. 2008, 148, 1681–1694. [Google Scholar] [CrossRef] [Green Version]
- Iñigo, S.; Alvarez, M.J.; Strasser, B.; Califano, A.; Cerdán, P.D. PFT1, the MED25 subunit of the plant Mediator complex, promotes flowering through CONSTANS dependent and independent mechanisms in Arabidopsis. Plant J. 2011, 69, 601–612. [Google Scholar] [CrossRef]
- Su, L.; Shan, J.X.; Gao, J.P.; Lin, H.X. OsHAL3, a blue light-responsive protein, interacts with the floral regulator Hd1 to activate flowering in rice. Mol. Plant 2015, 9, 233–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Wang, X.J. Effects of blue light and sucrose on anthocyanin accumulation and chalcone synthase gene expression in Arabidopsis. J. Trop. Subtrop. Bot. 2004, 12, 252–256. [Google Scholar]
- Zheng, L.; Van Labeke, M.C. Long-term effects of red-and blue-light emitting diodes on leaf anatomy and photosynthetic effi-ciency of three ornamental pot plants. Front. Plant Sci. 2017, 8, 917. [Google Scholar] [CrossRef] [Green Version]
- Sæbø, A.; Krekling, T.; Appelgren, M. Light quality affects photosynthesis and leaf anatomy of birch plantlets in vitro. Plant Cell Tissue Organ Cult. 1995, 41, 177–185. [Google Scholar] [CrossRef]
- Bornman, J.F.; Vogelmann, T.C.; Martin, G. Measurement of chlorophyll fluorescence within leaves using a fibreoptic microprobe. Plant Cell Environ. 1991, 14, 719–725. [Google Scholar] [CrossRef]
- Terashima, I.; Saeki, T. A new model for leaf photosynthesis incorporating the gradients of light environment and of photo-synthetic properties of chloroplasts within a leaf. Ann. Bot. 1985, 56, 489–499. [Google Scholar] [CrossRef]
- Klem, K.; Gargallo-Garriga, A.; Rattanapichai, W.; Oravec, M.; Holub, P.; Veselá, B.; Sardans, J.; Peñuelas, J.; Urban, O. Distinct morphological, physiological, and biochemical responses to light quality in barley leaves and roots. Front. Plant Sci. 2019, 10, 1026. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Zang, J.; Xu, Z.; Guo, S.; Jiao, X.; Liu, X.; Gao, Y. Effects of different light quality on growth, chlorophyll concentration and chlorophyll biosynthesis precursors of non-heading Chinese cabbage (Brassica campestris L.). Acta Physiol. Plant. 2013, 35, 2721–2726. [Google Scholar] [CrossRef]
- Naznin, M.T.; Lefsrud, M.; Gravel, V.; Azad, O.K. Blue light added with red LEDs enhance growth characteristics, pigments content, and antioxidant capacity in lettuce, spinach, kale, basil, and sweet pepper in a controlled environment. Plants 2019, 8, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Li, Y.; Zhong, S. Interplay between light and plant hormones in the control of Arabidopsis seedling chlorophyll biosyn-thesis. Front. Plant Sci. 2017, 8, 1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno, T.; Amaki, W.; Watanabe, H. Effects of monochromatic light irradiation by led on the growth and anthocyanin contents in leaves of cabbage seedlings. Acta Hortic. 2011, 179–184. [Google Scholar] [CrossRef]
- Li, H.; Tang, C.; Xu, Z.; Liu, X.; Han, X. Effects of different light sources on the growth of non-heading Chinese cabbage (Brassica campestris L.). J. Agric. Sci. 2012, 4, 262. [Google Scholar] [CrossRef] [Green Version]
- Snowden, M.C.; Cope, K.; Bugbee, B. Sensitivity of seven diverse species to blue and green light: Interactions with photon flux. PLoS ONE 2016, 11, e0163121. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, A.; Mehrjerdi, M.Z.; Aliniaeifard, S.; Seif, M. Photosynthetic and growth responses of green and purple basil plants under different spectral compositions. Physiol. Mol. Biol. Plants 2019, 25, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Marten, H.; Hedrich, R.; Roelfsema, M.R.G. Blue light inhibits guard cell plasma membrane anion channels in a phototropin-dependent manner. Plant J. 2007, 50, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Gu, M.; Cui, J.; Shi, K.; Zhou, Y.; Yu, J. Effects of light quality on CO2 assimilation, chlorophyll-fluorescence quenching, expression of calvin cycle genes and carbohydrate accumulation in Cucumis sativus. J. Photochem. Photobiol. B Biol. 2009, 96, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Ong, B.L. Effect of radiation quality on growth and photosynthesis of Acacia mangium seedlings. Photosynthetica 2003, 41, 349–355. [Google Scholar] [CrossRef]
- Ohashi-Kaneko, K.; Matsuda, R.; Goto, E.; Fujiwara, K.; Kurata, K. Growth of rice plants under red light with or without supplemental blue light. Soil Sci. Plant Nutr. 2006, 52, 444–452. [Google Scholar] [CrossRef]
- Krause, A.G.; Weis, E. Chlorophyll fluorescence and photosynthesis: The basics. Ann. Rev. Plant Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Schreiber, U. Chlorophyll fluorescence yield changes as a tool in plant physiology I. The measuring system. Photosynth. Res. 1983, 4, 361–373. [Google Scholar] [CrossRef]
- Ouzounis, T.; Fretté, X.; Ottosen, C.O.; Rosenqvist, E. Spectral effects of leds on chlorophyll fluorescence and pigmentation in Phalaenopsis ‘Vivien’ and ‘Purple Star’. Physiol. Plant. 2015, 154, 314–327. [Google Scholar] [CrossRef]
- Samuolienė, G.; Brazaitytė, A.; Duchovskis, P.; Viršilė, A.; Jankauskienė, J.; Sirtautas, R.; Novičkovas, A.; Sakalauskienė, S.; Sakalauskaitė, J. Cultivation of vegetable transplants using solid-state lamps for the short-wavelength supplementary lighting in greenhouses. In Proceedings of the International Symposium on Advanced Technologies and Management towards Sustainable Greenhouse Eco-Systems—Greensys 2011, Athens, Greece, 5–10 June 2011; pp. 885–892. [Google Scholar]
- Aalifar, M.; Aliniaeifard, S.; Arab, M.; Zare Mehrjerdi, M.; Dianati Daylami, S.; Serek, M.; Woltering, E.; Li, T. Blue light im-proves vase life of carnation cut flowers through its effect on the antioxidant defense system. Front. Plant Sci. 2020, 11, 511. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Liu, Y.; Song, L.; Jacobs, D.; Du, X.; Ying, Y.; Shao, Q.; Wu, J. Effect of differential light quality on morphology, photosynthesis, and antioxidant enzyme activity in Camptotheca acuminata seedlings. J. Plant Growth Regul. 2016, 36, 148–160. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Kim, K.; Kook, H.S.; Jang, Y.J.; Lee, W.H.; Kamala-Kannan, S.; Chae, J.C.; Lee, K.J. The effect of blue-light-emitting diodes on antioxidant properties and resistance to Botrytis cinerea in tomato. J. Plant Pathol. Microbiol. 2013, 4, 10.4172. [Google Scholar]
- Bartoli, C.G.; Yu, J.; Gómez, F.; Fernández, L.; McIntosh, L.; Foyer, C.H. Inter-relationships between light and respiration in the control of ascorbic acid synthesis and accumulation in Arabidopsis thaliana leaves. J. Exp. Bot. 2006, 57, 1621–1631. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Zhang, L.; Kato, M.; Yamawaki, K.; Kiriiwa, Y.; Yahata, M.; Ikoma, Y.; Matsumoto, H. Effect of blue and red LED Light irradiation on β-cryptoxanthin accumulation in the flavedo of citrus fruits. J. Agric. Food Chem. 2011, 60, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ma, G.; Yamawaki, K.; Ikoma, Y.; Matsumoto, H.; Yoshioka, T.; Ohta, S.; Kato, M. Regulation of ascorbic acid metabolism by blue led light irradiation in citrus juice sacs. Plant Sci. 2015, 233, 134–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojciechowska, R.; Długosz-Grochowska, O.; Kołton, A.; Żupnik, M. Effects of led supplemental lighting on yield and some quality parameters of lamb’s lettuce grown in two winter cycles. Sci. Horticul. 2015, 187, 80–86. [Google Scholar] [CrossRef]
- Simlat, M.; Ślęzak, P.; Moś, M.; Warchoł, M.; Skrzypek, E.; Ptak, A. The effect of light quality on seed germination, seedling growth and selected biochemical properties of Stevia rebaudiana Bertoni. Sci. Hortic. 2016, 211, 295–304. [Google Scholar] [CrossRef]
- Roscher, E.; Zetsche, K. The effects of light quality and intensity on the synthesis of ribulose-1,5-bisphosphate carboxylase and its mRNAs in the green alga Chlorogonium elongatum. Planta 1986, 167, 582–586. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D.; Seemann, J.R.; Berry, J.A. Regulation of ribulose-1,5-bisphosphate carboxylase activity in response to changing partial pressure of O2 and light in Phaseolus vulgaris. Plant Physiol. 1986, 81, 788–791. [Google Scholar] [CrossRef] [Green Version]
- Humbeck, K.; Schumann, R.; Senger, H. The influence of blue light on the formation of chlorophyll-protein complexes in Scenedesmus. In Blue Light Effects in Biological Systems; Springer: Berlin/Heidelberg, Germany, 1984; pp. 359–365. [Google Scholar]
- Kamiya, A.; Miyachi, S. Blue light-induced formation of phosphoenolpyruvate carboxylase in colorless Chlorella mutant cells. Plant Cell Physiol. 1975, 16, 729–736. [Google Scholar]
- Conradt, W.; Ruyters, G. Blue light-effects on enzymes of the carbohydrate metabolism in chlorella 2. Glyceraldehyde 3-phosphate dehydrogenase (NADP-dependent). In The Blue Light Syndrome; Springer: Berlin/Heidelberg, Germany, 1980; pp. 368–371. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene Identifier | Description | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|---|---|
| KfACTIN | Kaladp0071s0282 | As reference gene | TTCGAGACCTTCAATGCTCCT | GATGGCTGGAAAAGCACCTCA |
| KfPHYA | Kaladp0034s0172 | Phytochrome A, flowering promoted gene [Kalanchoe fedtschenkoi] | TTCAGCCATTTGGGTGTTTGT | CGTATCACACAGCTTGTCCAG |
| KfPHYB | Kaladp0039s0298 | Phytochrome B, flowering suppressor gene [Kalanchoe fedtschenkoi] | CACATGCTCTCTCTGACTCCC | AAAAGCTGAATGTCCCCTCCA |
| KfCRY1 | Kaladp0071s0308 | Cryptochrome 1, flowering promoted gene [Kalanchoe fedtschenkoi] | TCTTGGCGCCAGTTTCATCA | TGTCAGCATTGCTCCATCCA |
| KfFT | Kaladp0099s0141 | PROTEIN FLOWERING LOCUS T (FT), regulation of flower development [Kalanchoe fedtschenkoi] | AGGGAAGTTGCAAATGCGTG | GGAAGTTCTGACGCCATCCC |
| KfFPF-1 | Kaladp1115s0001 | FLOWERING-PROMOTING FACTOR 1-like PROTEIN 1-related [Kalanchoe fedtschenkoi] | AGCGCTGATCAAAACGGTAGA | GAAAAATCGCGAGGAAGGGA |
| KfPEF-4 | Kaladp0059s0037 | PROTEIN EARLY FLOWERING 4 [Kalanchoe fedtschenkoi] | GGCGGCGGAGATATTTGTGA | TGGCAGCACGATGATGAAAG |
| Photoperiod (I) | Blue Light (II) | Shoot | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Plant Height (cm) | Canopy Diameter (cm) | No. of Nodes | No. of Branches | Length of Top 4th Internode (mm) | Stem Diameter (mm) | Fresh Weight (g) | Dry Weight (g) | ||
| Short day 10 h (SD) | None | 13.1 ± 0.17 d 1 | 13.0 ± 0.24 e | 7.4 ± 0.13 c | 1.3 ± 0.04 d | 14.9 ± 0.51 b | 6.1 ± 0.11 d | 66.6 ± 1.17 f | 2.8 ± 0.05 f |
| + 4B | 15.1 ± 0.20 c | 14.7 ± 0.23 d | 10.6 ± 0.15 b | 2.9 ± 0.04 cd | 13.3 ± 0.47 c | 7.8 ± 0.17 c | 98.3 ± 1.03 e | 3.9 ± 0.05 e | |
| + NI-4B | 15.2 ± 0.23 c | 16.6 ± 0.15 c | 10.9 ± 0.15 b | 3.4 ± 0.05 c | 13.1 ± 0.38 c | 7.6 ± 0.20 c | 121.1 ± 1.98 d | 5.0 ± 0.04d | |
| Long day 13 h (LD) | None | 17.7 ± 0.16 b | 17.7 ± 0.20 bc | 10.8 ± 0.17 b | 6.6 ± 0.12 b | 15.9 ± 0.27 a | 8.5 ± 0.14 b | 135.9 ± 2.03 c | 6.0 ± 0.09 c |
| + 4B | 18.6 ± 0.23 ab | 18.6 ± 0.32 b | 13.1 ± 0.24 a | 5.8 ± 0.10 bc | 14.0 ± 0.36 b | 9.7 ± 0.24 a | 155.2 ± 2.17 b | 7.2 ± 0.09 b | |
| + NI-4B | 20.7 ± 0.27 a | 21.5 ± 0.26 a | 13.7 ± 0.21 a | 8.7 ± 0.25 a | 14.2 ± 0.41 b | 9.9 ± 0.23 a | 169.5 ± 1.74 a | 8.0 ± 0.08 a | |
| F-test | I | *** | *** | *** | *** | *** | *** | *** | *** |
| II | *** | *** | *** | ** | ** | ** | *** | *** | |
| I × II | *** | *** | *** | *** | *** | *** | *** | *** | |
| Photoperiod (I) | Blue light (II) | Leaf | Flower | Root | |||||
| Number | Length (cm) | Width (cm) | Days to visible flower buds | No. of inflorescences | Length (cm) | Fresh weight (g) | Dry weight (g) | ||
| Short day 10 h (SD) | None | 18.3 ± 1.34 e | 7.1 ± 0.34 b | 6.2 ± 0.20 ab | 17.0 ± 1.17 c | 6.7 ± 0.98 b | 12.7 ± 0.57 d | 4.8 ± 0.06 c | 0.5 ± 0.01 c |
| + 4B | 24.7 ± 2.43 d | 6.4 ± 0.41 bc | 5.2 ± 0.23 b | 24.7 ± 1.13 b | 11.3 ± 1.01 ab | 15.3 ± 0.71 cd | 8.5 ± 0.07 bc | 0.9 ± 0.02 bc | |
| + NI-4B | 30.7 ± 2.17 c | 6.6 ± 0.52 bc | 5.5 ± 0.32 b | 24.3 ± 1.03 b | 12.3 ± 1.12 ab | 16.0 ± 0.67 c | 9.1 ± 0.05 b | 1.1 ± 0.02 b | |
| Long day 13 h (LD) | None | 51.8 ± 5.44 b | 8.4 ± 0.43 a | 7.2 ± 0.24 a | - | 0.0 ± 0.00 c | 17.2 ± 0.54 b | 16.1 ± 0.10 a | 2.1 ± 0.02 a |
| + 4B | 53.3 ± 6.27 b | 7.6 ± 0.37 ab | 6.4 ± 0.31 ab | 33.3 ± 1.24 a | 14.3 ± 1.17 a | 18.4 ± 0.63 ab | 15.9 ± 0.10 a | 1.9 ± 0.02 a | |
| + NI-4B | 68.1 ± 5.72 a | 7.5 ± 0.31 ab | 6.5 ± 0.27 ab | - | 0.0 ± 0.00 c | 19.21 ± 0.91 a | 16.0 ± 0.09 a | 2.0 ± 0.01 a | |
| F-test | I | *** | *** | ** | *** | *** | *** | *** | *** |
| II | *** | ** | ** | *** | *** | *** | ** | ** | |
| I × II | *** | ** | ** | ** | *** | *** | *** | *** | |
| Photoperiod (I) | Blue Light (II) | Pn 2 (μmol CO2 m−2s−1) | Tr 3 (mmol H2O m−2s−1) | Gs 4 (mol H2O m−2s−1) | Ci 5 (μmol CO2 mol−1) | Fv/Fm 6 | Fv’/Fm’ 7 |
|---|---|---|---|---|---|---|---|
| Short day 10 h (SD) | None | 13.0 ± 0.63 e 1 | 1.59 ± 0.014 cd | 0.39 ± 0.013 cd | 357.5 ± 10.21 cd | 0.75 ± 0.017 bc | 0.44 ± 0.011 bc |
| + 4B | 16.3 ± 0.57 d | 2.03 ± 0.011 b | 0.76 ± 0.014 b | 406.9 ± 10.24 b | 0.80 ± 0.013 ab | 0.52 ± 0.023 ab | |
| + NI-4B | 17.2 ± 0.87 cd | 2.18 ± 0.082 a | 0.81 ± 0.010 ab | 487.7 ± 17.21 a | 0.85 ± 0.016 a | 0.58 ± 0.019 a | |
| Long day 13 h (LD) | None | 17.8 ± 0.52 c | 1.76 ± 0.034 c | 0.48 ± 0.013 c | 380.2 ± 11.37 c | 0.78 ± 0.010 b | 0.48 + 0.013 b |
| + 4B | 19.9 ± 0.86 b | 2.08 ± 0.024 b | 0.77 ± 0.011 b | 413.9 ± 11.56 b | 0.81 ± 0.011 ab | 0.52 ± 0.024 ab | |
| + NI-4B | 21.9 ± 0.91 a | 2.22 ± 0.083 a | 0.86 ± 0.012 a | 499.2 ± 15.33 a | 0.86 ± 0.015 a | 0.59 ± 0.021 a | |
| F-test | I | *** | * | * | * | ** | ** |
| II | *** | *** | *** | *** | ** | ** | |
| I × II | *** | * | ** | * | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Song, J.; Jeong, B.R. Low-Intensity Blue Light Supplemented during Photoperiod in Controlled Environment Induces Flowering and Antioxidant Production in Kalanchoe. Antioxidants 2022, 11, 811. https://doi.org/10.3390/antiox11050811
Yang J, Song J, Jeong BR. Low-Intensity Blue Light Supplemented during Photoperiod in Controlled Environment Induces Flowering and Antioxidant Production in Kalanchoe. Antioxidants. 2022; 11(5):811. https://doi.org/10.3390/antiox11050811
Chicago/Turabian StyleYang, Jingli, Jinnan Song, and Byoung Ryong Jeong. 2022. "Low-Intensity Blue Light Supplemented during Photoperiod in Controlled Environment Induces Flowering and Antioxidant Production in Kalanchoe" Antioxidants 11, no. 5: 811. https://doi.org/10.3390/antiox11050811