
2,3,5,4′-Tetrahydroxystilbene-2-O-β-glucoside Attenuates Reactive Oxygen Species-Dependent Inflammation and Apoptosis in Porphyromonas gingivalis-Infected Brain Endothelial Cells

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Primary Cell Isolation and Culture
2.3. Bacterial Culture and Preparation
2.4. 2,3,5,4′-Tetrahydroxystilbene-2-O-β-glucoside (THSG) Preparation
2.5. Antioxidant Treatment and Bacterial Infection
2.6. MTT Assay
2.7. Nuclear Staining
2.8. FITC Annexin V and PI Staining
2.9. Western Blot Analysis
2.10. NF-κB p65 Transcription Factor Assay
2.11. Measurement of Reactive Oxygen Species (ROS) Generation
2.12. Statistical Analysis
3. Results
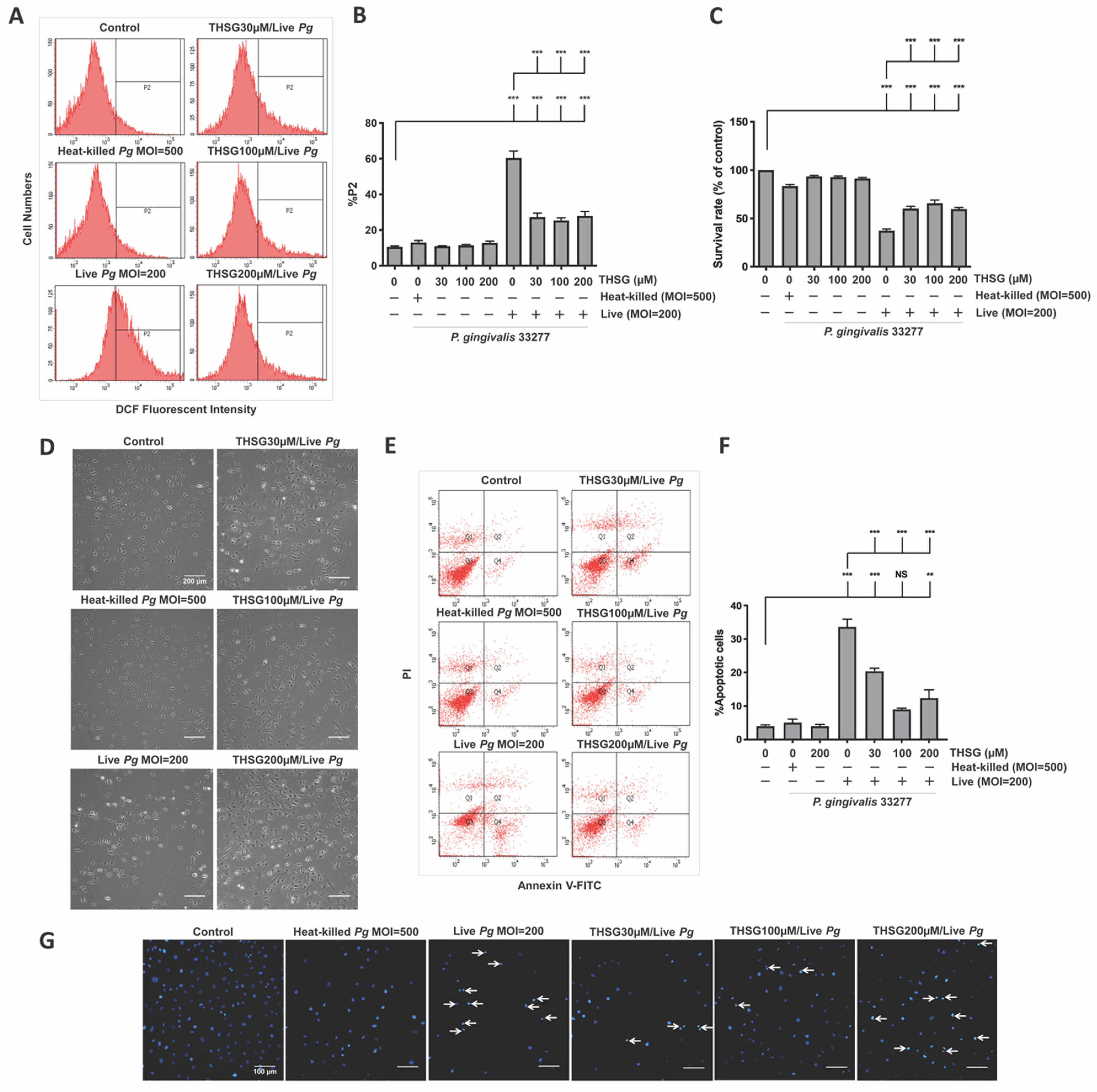
3.1. THSG Reduces Intracellular ROS Production and Increases Cell-Survival Rate in P. gingivalis-Infected Brain Endothelial Cells
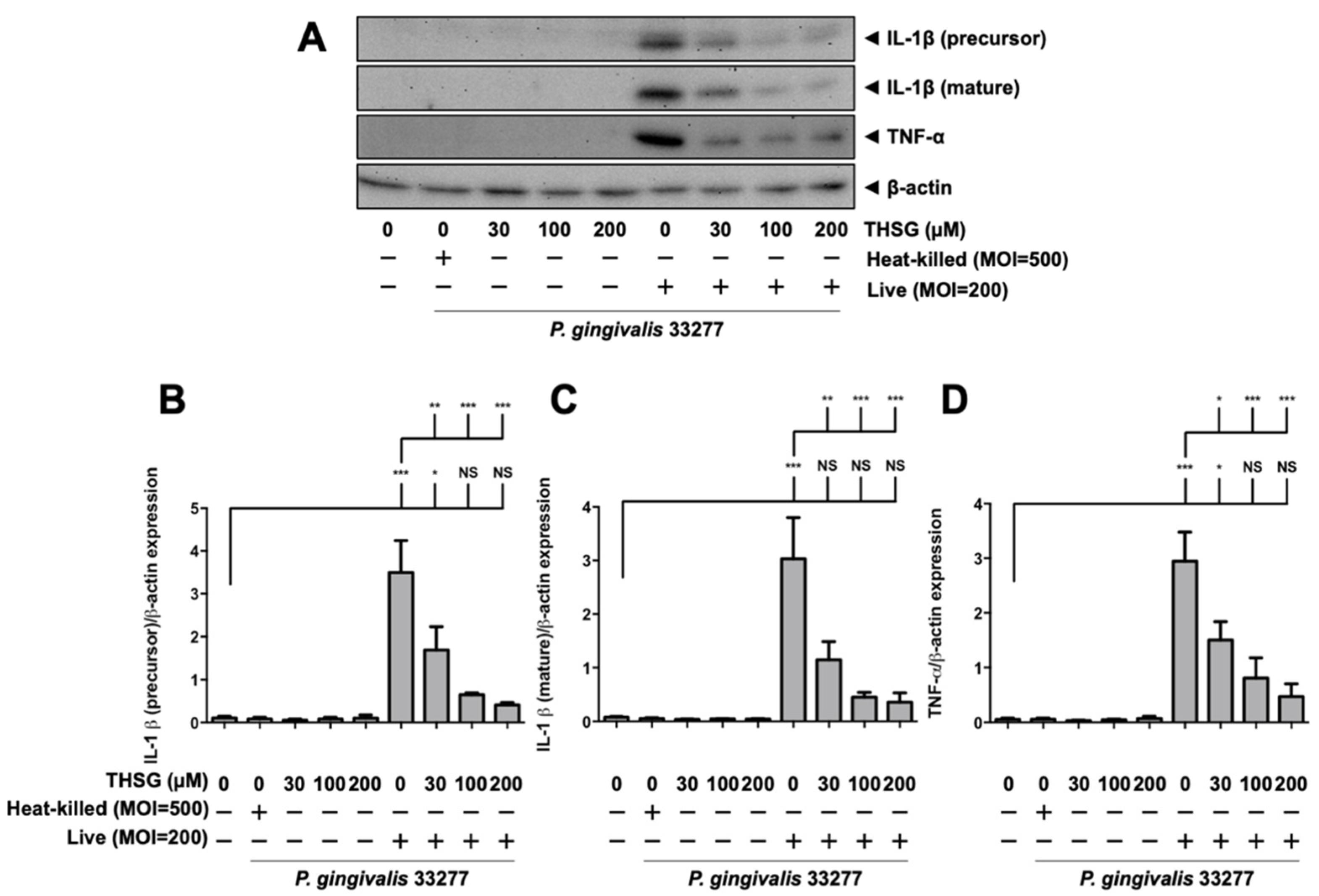
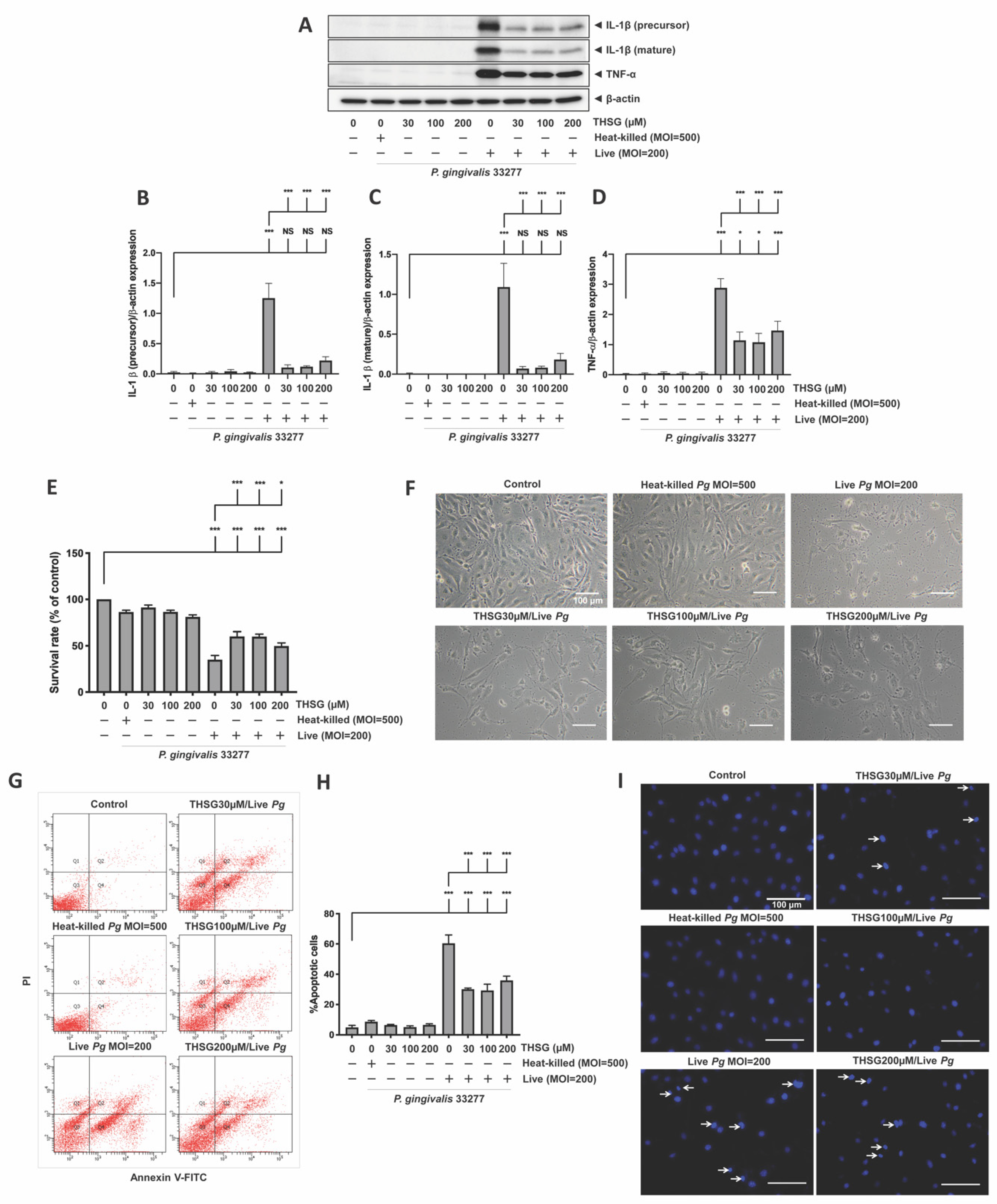
3.2. THSG Reduces the Upregulation of Proinflammatory Cytokines in P. gingivalis-Stimulated Brain Endothelial Cells
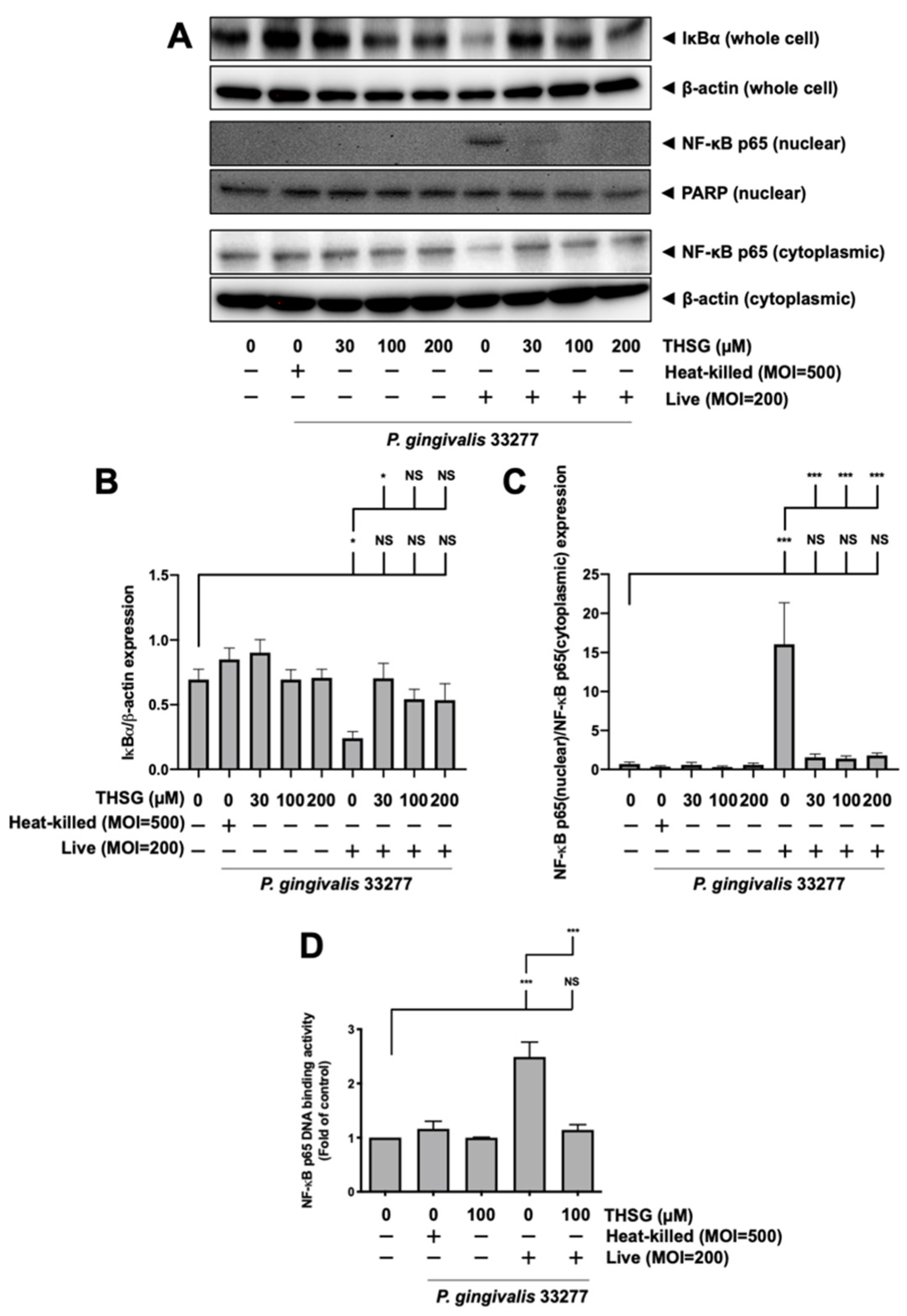
3.3. Treatment of THSG Inhibits NF-κB Signal Transduction Stimulated by P. gingivalis in Brain Endothelial Cells
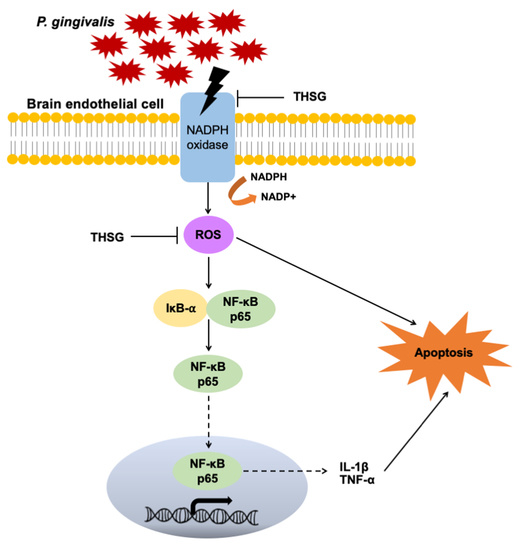
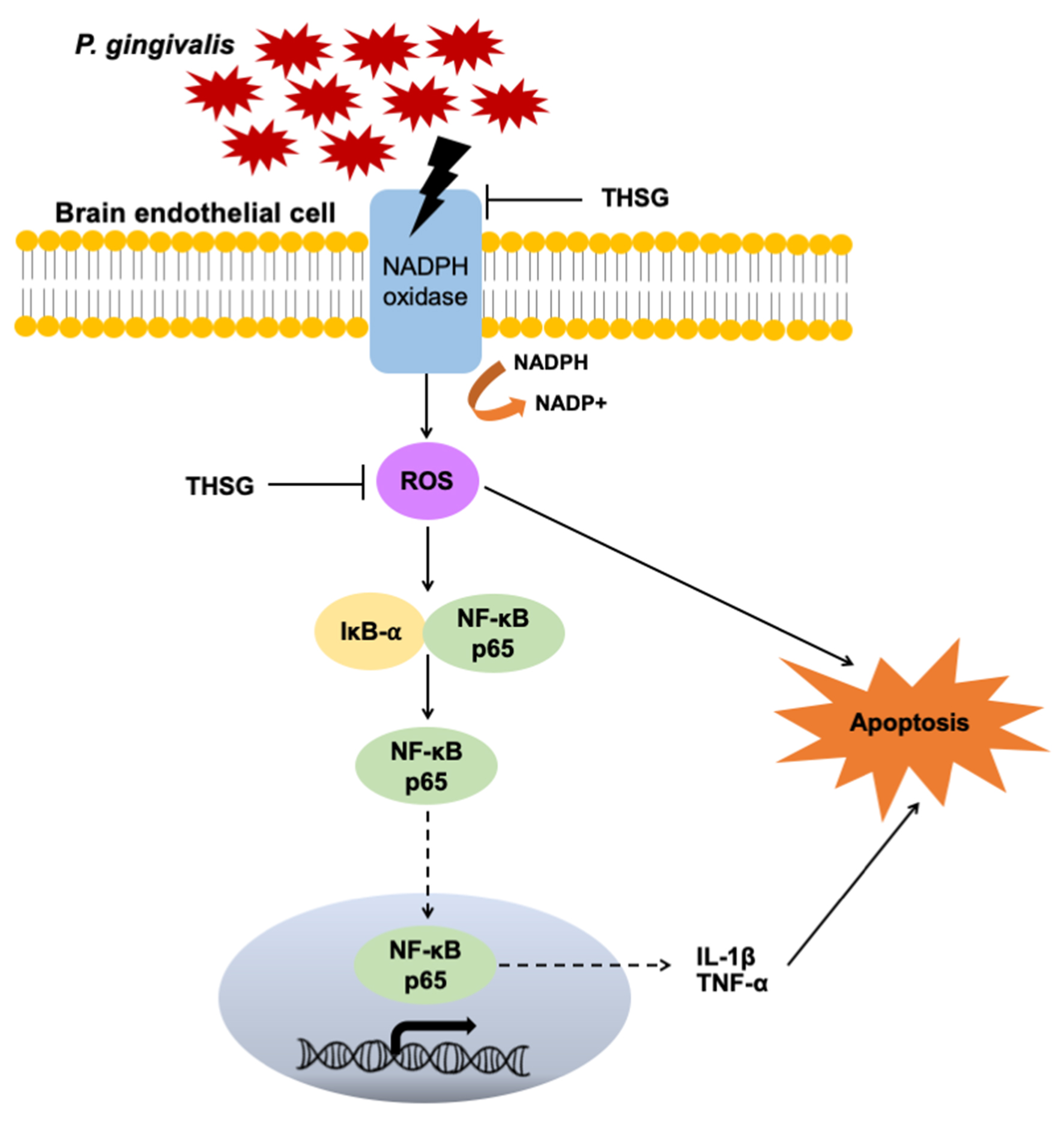
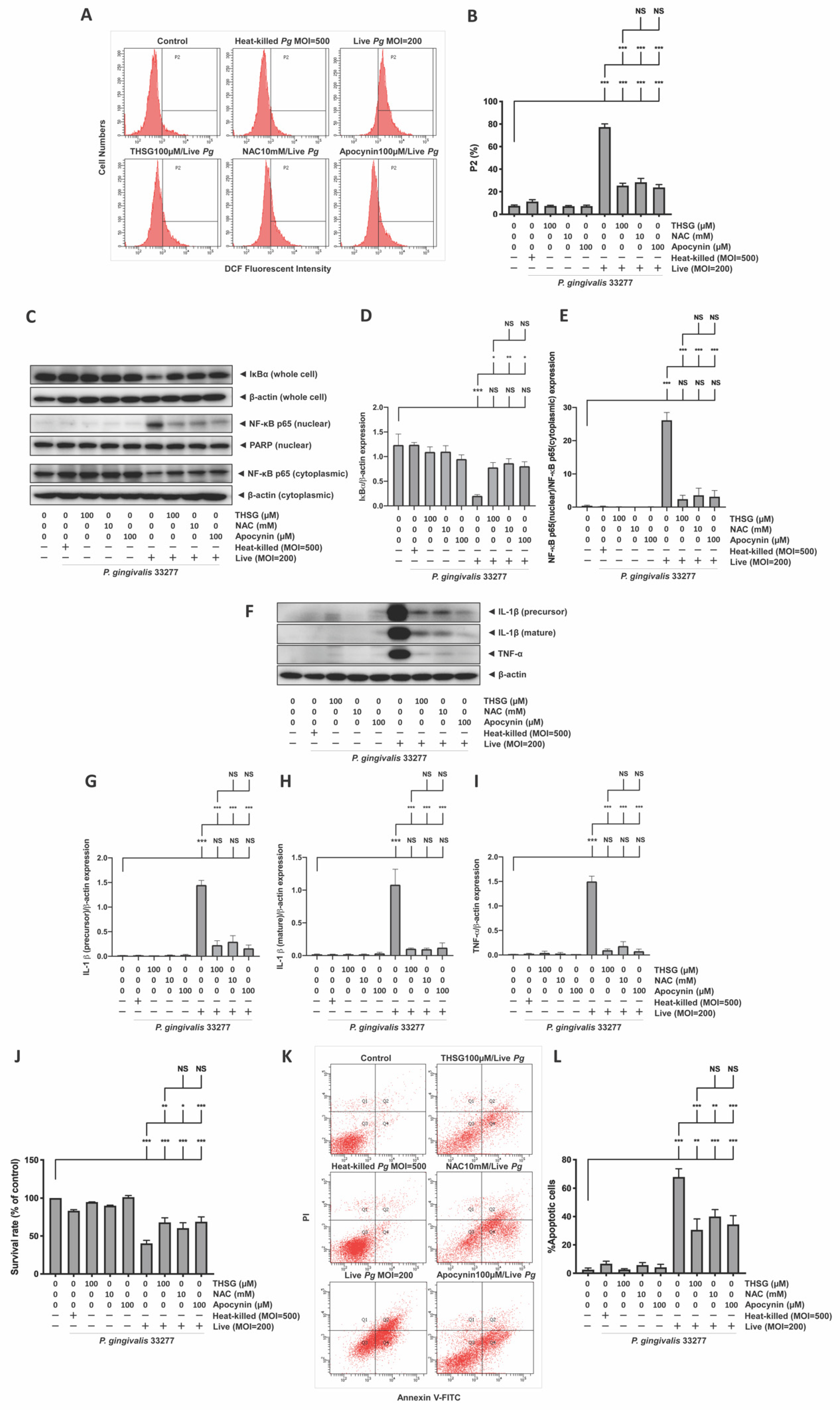
3.4. The Reduction in ROS and NF-κB Activation Is Responsible for the Anti-Inflammatory and Antiapoptotic Properties of THSG in P. gingivalis-Infected Brain Endothelial Cells
3.5. Anti-Inflammatory and Antiapoptotic Properties of THSG in P. gingivalis-Activated Primary Mouse Brain Endothelial Cells
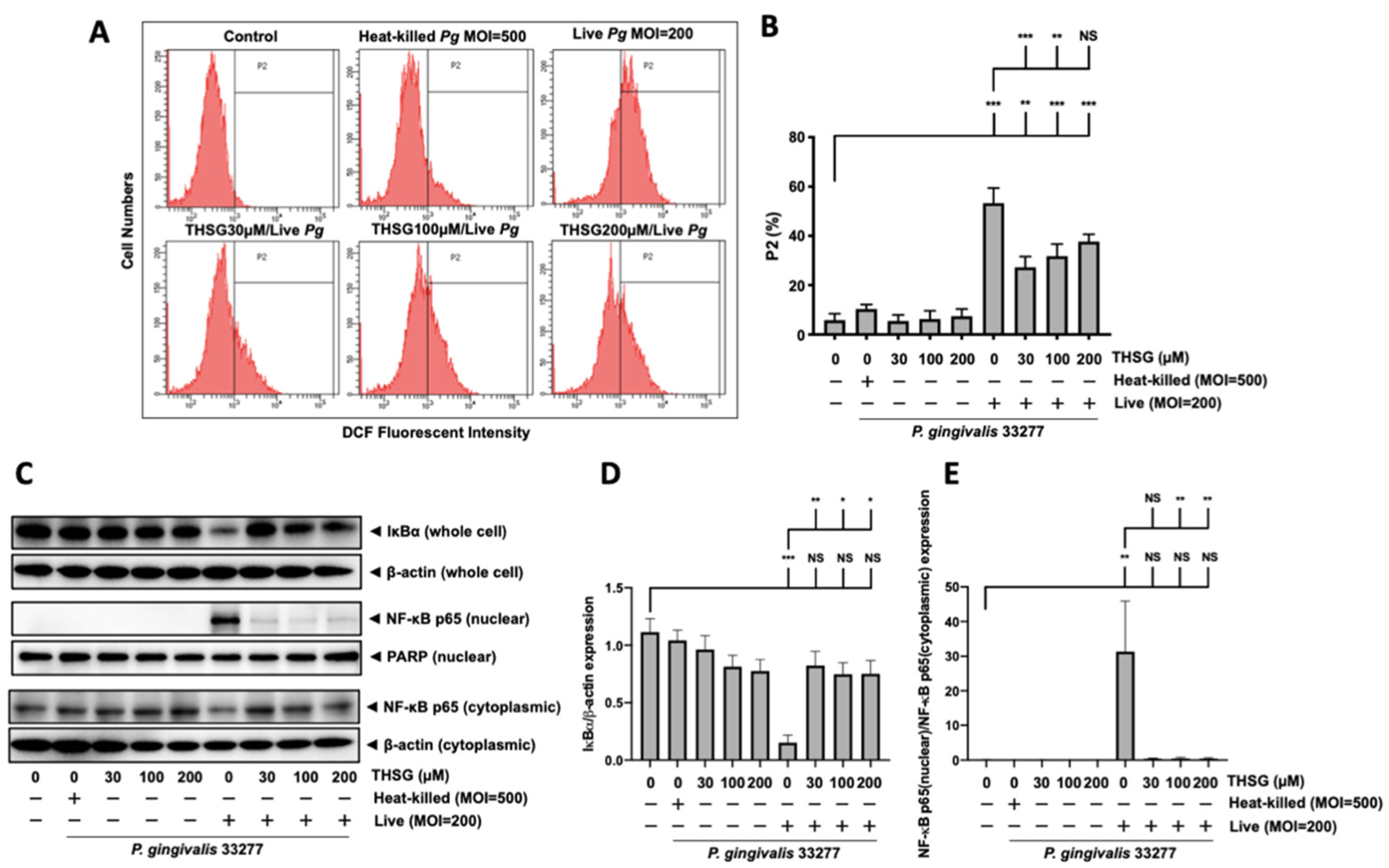
3.6. THSG Protects MBECs from Bacteria-Stimulated Inflammation and Apoptosis through ROS/NF-κB Signaling Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- How, K.Y.; Song, K.P.; Chan, K.G. Porphyromonas gingivalis: An overview of periodontopathic pathogen below the gum line. Front. Microbiol. 2016, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Darveau, R.P.; Curtis, M.A. The keystone-pathogen hypothesis. Nat. Rev. Microbiol. 2012, 10, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G. Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Kolltveit, K.M.; Tronstad, L.; Olsen, I. Systemic diseases caused by oral infection. Clin. Microbiol. Rev. 2000, 13, 547–558. [Google Scholar] [CrossRef]
- Grau, A.J.; Becher, H.; Ziegler, C.M.; Lichy, C.; Buggle, F.; Kaiser, C.; Lutz, R.; Bültmann, S.; Preusch, M.; Dörfer, C.E. Periodontal disease as a risk factor for ischemic stroke. Stroke 2004, 35, 496–501. [Google Scholar] [CrossRef] [Green Version]
- Pussinen, P.J.; Alfthan, G.; Jousilahti, P.; Paju, S.; Tuomilehto, J. Systemic exposure to porphyromonas gingivalis predicts incident stroke. Atherosclerosis 2007, 193, 222–228. [Google Scholar] [CrossRef]
- Ghizoni, J.S.; Taveira, L.A.; Garlet, G.P.; Ghizoni, M.F.; Pereira, J.R.; Dionísio, T.J.; Brozoski, D.T.; Santos, C.F.; Sant’Ana, A.C. Increased levels of porphyromonas gingivalis are associated with ischemic and hemorrhagic cerebrovascular disease in humans: An in vivo study. J. Appl. Oral Sci. 2012, 20, 104–112. [Google Scholar] [CrossRef]
- Bartold, P.M.; Narayanan, A.S. Molecular and cell biology of healthy and diseased periodontal tissues. Periodontol. 2000 2006, 40, 29–49. [Google Scholar] [CrossRef]
- Kleinstein, S.E.; Nelson, K.E.; Freire, M. Inflammatory networks linking oral microbiome with systemic health and disease. J. Dent. Res. 2020, 99, 1131–1139. [Google Scholar] [CrossRef]
- Forner, L.; Larsen, T.; Kilian, M.; Holmstrup, P. Incidence of bacteremia after chewing, tooth brushing and scaling in individuals with periodontal inflammation. J. Clin. Periodontol. 2006, 33, 401–407. [Google Scholar] [CrossRef]
- Charoensaensuk, V.; Chen, Y.C.; Lin, Y.H.; Ou, K.L.; Yang, L.Y.; Lu, D.Y. Porphyromonas gingivalis induces proinflammatory cytokine expression leading to apoptotic death through the oxidative stress/nf-κb pathway in brain endothelial cells. Cells 2021, 10, 3033. [Google Scholar] [CrossRef] [PubMed]
- Bullon, P.; Cordero, M.D.; Quiles, J.L.; Morillo, J.M.; del Carmen Ramirez-Tortosa, M.; Battino, M. Mitochondrial dysfunction promoted by porphyromonas gingivalis lipopolysaccharide as a possible link between cardiovascular disease and periodontitis. Free Radic. Biol. Med. 2011, 50, 1336–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uttara, B.; Singh, A.V.; Zamboni, P.; Mahajan, R.T. Oxidative stress and neurodegenerative diseases: A review of upstream and downstream antioxidant therapeutic options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelova, P.R.; Abramov, A.Y. Role of mitochondrial ros in the brain: From physiology to neurodegeneration. FEBS Lett. 2018, 592, 692–702. [Google Scholar] [CrossRef]
- Zuo, L.; Hemmelgarn, B.T.; Chuang, C.C.; Best, T.M. The role of oxidative stress-induced epigenetic alterations in amyloid-β production in alzheimer’s disease. Oxid Med. Cell Longev. 2015, 2015, 604658. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.F.; Chen, G.W.; Chen, Y.C.; Shen, C.K.; Lu, D.Y.; Yang, L.Y.; Chen, J.H.; Yeh, W.L. Regulatory effects of quercetin on m1/m2 macrophage polarization and oxidative/antioxidative balance. Nutrients 2021, 14, 67. [Google Scholar] [CrossRef]
- Tamaki, N.; Tomofuji, T.; Maruyama, T.; Ekuni, D.; Yamanaka, R.; Takeuchi, N.; Yamamoto, T. Relationship between periodontal condition and plasma reactive oxygen metabolites in patients in the maintenance phase of periodontal treatment. J. Periodontol. 2008, 79, 2136–2142. [Google Scholar] [CrossRef]
- Tamaki, N.; Hayashida, H.; Fukui, M.; Kitamura, M.; Kawasaki, K.; Nakazato, M.; Maeda, T.; Saito, T.; Ito, H.O. Oxidative stress and antibody levels to periodontal bacteria in adults: The nagasaki islands study. Oral Dis. 2014, 20, e49–e56. [Google Scholar] [CrossRef]
- Żukowski, P.; Maciejczyk, M.; Waszkiel, D. Sources of free radicals and oxidative stress in the oral cavity. Arch. Oral Biol. 2018, 92, 8–17. [Google Scholar] [CrossRef]
- Tamaki, N.; Cristina Orihuela-Campos, R.; Inagaki, Y.; Fukui, M.; Nagata, T.; Ito, H.O. Resveratrol improves oxidative stress and prevents the progression of periodontitis via the activation of the sirt1/ampk and the nrf2/antioxidant defense pathways in a rat periodontitis model. Free Radic Biol. Med. 2014, 75, 222–229. [Google Scholar] [CrossRef]
- Shirley, R.; Ord, E.N.; Work, L.M. Oxidative stress and the use of antioxidants in stroke. Antioxidants 2014, 3, 472–501. [Google Scholar] [CrossRef] [Green Version]
- Doyle, K.P.; Simon, R.P.; Stenzel-Poore, M.P. Mechanisms of ischemic brain damage. Neuropharmacology 2008, 55, 310–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamaki, N.; Tomofuji, T.; Ekuni, D.; Yamanaka, R.; Yamamoto, T.; Morita, M. Short-term effects of non-surgical periodontal treatment on plasma level of reactive oxygen metabolites in patients with chronic periodontitis. J. Periodontol. 2009, 80, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, N.; Tomofuji, T.; Ekuni, D.; Yamanaka, R.; Morita, M. Periodontal treatment decreases plasma oxidized ldl level and oxidative stress. Clin. Oral Investig. 2011, 15, 953–958. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.Y.; Tang, C.H.; Chen, Y.H.; Wei, I.H.; Chen, J.H.; Lai, C.H.; Lu, D.Y. Peptidoglycan enhances proinflammatory cytokine expression through the tlr2 receptor, myd88, phosphatidylinositol 3-kinase/akt and nf-kappab pathways in bv-2 microglia. Int. Immunopharmacol. 2010, 10, 883–891. [Google Scholar] [CrossRef]
- Shen, C.K.; Huang, B.R.; Yeh, W.L.; Chen, C.W.; Liu, Y.S.; Lai, S.W.; Tseng, W.P.; Lu, D.Y.; Tsai, C.F. Regulatory effects of il-1β in the interaction of gbm and tumor-associated monocyte through vcam-1 and icam-1. Eur. J. Pharmacol. 2021, 905, 174216. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. Nf-κb signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and nf-κb signaling. Cell Res 2011, 21, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Ott, M.; Gogvadze, V.; Orrenius, S.; Zhivotovsky, B. Mitochondria, oxidative stress and cell death. Apoptosis 2007, 12, 913–922. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, J. Biological effects of tetrahydroxystilbene glucoside: An active component of a rhizome extracted from polygonum multiflorum. Oxid Med. Cell Longev. 2018, 2018, 3641960. [Google Scholar] [CrossRef] [Green Version]
- Chin, Y.T.; Cheng, G.Y.; Shih, Y.J.; Lin, C.Y.; Lin, S.J.; Lai, H.Y.; Whang-Peng, J.; Chiu, H.C.; Lee, S.Y.; Fu, E.; et al. Therapeutic applications of resveratrol and its derivatives on periodontitis. Ann. N. Y. Acad. Sci. 2017, 1403, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Gu, J.; Wu, P.F.; Wang, F.; Xiong, Z.; Yang, Y.J.; Wu, W.N.; Dong, L.D.; Chen, J.G. Protection by tetrahydroxystilbene glucoside against cerebral ischemia: Involvement of jnk, sirt1, and nf-kappab pathways and inhibition of intracellular ros/rns generation. Free Radic. Biol. Med. 2009, 47, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Wang, H.; Zhang, Z.; Liang, K. Stilbene glycoside protects osteoblasts against oxidative damage via nrf2/ho-1 and nf-kappab signaling pathways. Arch. Med. Sci. 2019, 15, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Chin, Y.T.; Hsieh, M.T.; Lin, C.Y.; Kuo, P.J.; Yang, Y.C.; Shih, Y.J.; Lai, H.Y.; Cheng, G.Y.; Tang, H.Y.; Lee, C.C.; et al. 2,3,5,4′-tetrahydroxystilbene-2-o-β-glucoside isolated from polygoni multiflori ameliorates the development of periodontitis. Mediat. Inflamm. 2016, 2016, 6953459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.H.; Wang, W.Q.; Wang, J.L. Protective effect of tetrahydroxystilbene glucoside on cardiotoxicity induced by doxorubicin in vitro and in vivo. Acta Pharmacol. Sin. 2009, 30, 1479–1487. [Google Scholar] [CrossRef] [Green Version]
- Jiao, C.; Gao, F.; Ou, L.; Yu, J.; Li, M.; Wei, P.; Miu, F. Tetrahydroxystilbene glycoside antagonizes β-amyloid-induced inflammatory injury in microglia cells by regulating pu.1 expression. Neuroreport 2018, 29, 787–793. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, L.; Li, X.; Liu, C.; Sun, X.; Wu, L.; Li, T.; Yang, H.; Chen, J. Potential molecular mechanisms mediating the protective effects of tetrahydroxystilbene glucoside on mpp(+)-induced pc12 cell apoptosis. Mol. Cell Biochem. 2017, 436, 203–213. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, S.; Song, F.; Nian, L.; Zhou, X.; Wang, S. 2,3,5,4′-tetrahydroxystilbene-2-o-β-d-glucoside protects human umbilical vein endothelial cells against lysophosphatidylcholine-induced apoptosis by upregulating superoxide dismutase and glutathione peroxidase. IUBMB Life 2014, 66, 711–722. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, C.H.; Li, F.; Zhu, W.Z. 2,3,4′,5-tetrahydroxystilbene-2-o-beta-d-glucoside suppresses matrix metalloproteinase expression and inflammation in atherosclerotic rats. Clin. Exp. Pharmacol. Physiol. 2008, 35, 310–316. [Google Scholar] [CrossRef]
- Tsai, P.W.; Lee, Y.H.; Chen, L.G.; Lee, C.J.; Wang, C.C. In vitro and in vivo anti-osteoarthritis effects of 2,3,5,4′-tetrahydroxystilbene-2-o-β-d-glucoside from polygonum multiflorum. Molecules 2018, 23, 571. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.F.; Guan, S.Y.; Luo, L.; Li, Y.J.; Yang, L.; Zhou, X.X.; Guo, G.D.; Zhao, M.G.; Yang, Q.; Liu, G. Tetrahydroxystilbene glucoside relieves the chronic inflammatory pain by inhibiting neuronal apoptosis, microglia activation, and glun2b overexpression in anterior cingulate cortex. Mol. Pain 2018, 14, 1744806918814367. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Sun, F.; Zhang, L.; Sun, X.; Li, L. Tetrahydroxystilbene glucoside inhibits α-synuclein aggregation and apoptosis in a53t α-synuclein-transfected cells exposed to mpp+. Can. J. Physiol. Pharmacol. 2017, 95, 750–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Jin, M.L.; Wang, Z.; Park, G.; Choi, Y.W. 2,3,4′,5-tetrahydroxystilbene-2-o-beta-d-glucoside exerts anti-inflammatory effects on lipopolysaccharide-stimulated microglia by inhibiting nf-kappab and activating ampk/nrf2 pathways. Food Chem. Toxicol. 2016, 97, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.Y.; Qin, X.Y.; Yuan, M.M.; Lu, G.J.; Cheng, Y. 2,3,5,4′-tetrahydroxystilbene-2-o-beta-d-glucoside reverses stress-induced depression via inflammatory and oxidative stress pathways. Oxid Med. Cell Longev. 2018, 2018, 9501427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, S.; Xu, J.W. Biological activities of 2,3,5,4′-tetrahydroxystilbene-2-o-β-d-glucoside in antiaging and antiaging-related disease treatments. Oxid Med. Cell Longev. 2016, 2016, 4973239. [Google Scholar] [CrossRef] [Green Version]
- Yao, W.; Gu, C.; Shao, H.; Meng, G.; Wang, H.; Jing, X.; Zhang, W. Tetrahydroxystilbene glucoside improves tnf-α-induced endothelial dysfunction: Involvement of tgfβ/smad pathway and inhibition of vimentin expression. Am. J. Chin. Med. 2015, 43, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Shapira, L.; Ayalon, S.; Brenner, T. Effects of porphyromonas gingivalis on the central nervous system: Activation of glial cells and exacerbation of experimental autoimmune encephalomyelitis. J. Periodontol. 2002, 73, 511–516. [Google Scholar] [CrossRef]
- L’Azou, B.; Fernandez, P.; Bareille, R.; Beneteau, M.; Bourget, C.; Cambar, J.; Bordenave, L. In vitro endothelial cell susceptibility to xenobiotics: Comparison of three cell types. Cell Biol. Toxicol. 2005, 21, 127–137. [Google Scholar] [CrossRef]
- Jamalzadeh, L.; Ghafoori, H.; Sariri, R.; Rabuti, H.; Nasirzade, J.; Hasani, H.; Aghamaali, M.R. Cytotoxic effects of some common organic solvents on mcf-7, raw-264.7 and human umbilical vein endothelial cells. Avicenna J. Med. Biochem. 2016, 4, 10-33453. [Google Scholar] [CrossRef] [Green Version]
- Régnier, P.; Bastias, J.; Rodriguez-Ruiz, V.; Caballero-Casero, N.; Caballo, C.; Sicilia, D.; Fuentes, A.; Maire, M.; Crepin, M.; Letourneur, D.; et al. Astaxanthin from haematococcus pluvialis prevents oxidative stress on human endothelial cells without toxicity. Mar. Drugs 2015, 13, 2857–2874. [Google Scholar] [CrossRef] [Green Version]
- Ko, C.H.; Shen, S.C.; Yang, L.Y.; Lin, C.W.; Chen, Y.C. Gossypol reduction of tumor growth through ros-dependent mitochondria pathway in human colorectal carcinoma cells. Int. J. Cancer 2007, 121, 1670–1679. [Google Scholar] [CrossRef] [PubMed]
- Van Engeland, M.; Nieland, L.J.; Ramaekers, F.C.; Schutte, B.; Reutelingsperger, C.P. Annexin v-affinity assay: A review on an apoptosis detection system based on phosphatidylserine exposure. Cytom. J. Int. Soc. Anal. Cytol. 1998, 31, 1–9. [Google Scholar] [CrossRef]
- Yang, L.Y.; Shen, S.C.; Cheng, K.T.; Subbaraju, G.V.; Chien, C.C.; Chen, Y.C. Hispolon inhibition of inflammatory apoptosis through reduction of inos/no production via ho-1 induction in macrophages. J. Ethnopharmacol. 2014, 156, 61–72. [Google Scholar] [CrossRef]
- Dominy, S.S.; Lynch, C.; Ermini, F.; Benedyk, M.; Marczyk, A.; Konradi, A.; Nguyen, M.; Haditsch, U.; Raha, D.; Griffin, C.; et al. Porphyromonas gingivalis in alzheimer’s disease brains: Evidence for disease causation and treatment with small-molecule inhibitors. Sci. Adv. 2019, 5, eaau3333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Zúñiga, J.; More, J.; Melgar-Rodríguez, S.; Jiménez-Unión, M.; Villalobos-Orchard, F.; Muñoz-Manríquez, C.; Monasterio, G.; Valdés, J.L.; Vernal, R.; Paula-Lima, A. Alzheimer’s disease-like pathology triggered by porphyromonas gingivalis in wild type rats is serotype dependent. Front. Immunol. 2020, 11, 588036. [Google Scholar] [CrossRef]
- Tang, Z.; Liang, D.; Cheng, M.; Su, X.; Liu, R.; Zhang, Y.; Wu, H. Effects of porphyromonas gingivalis and its underlying mechanisms on alzheimer-like tau hyperphosphorylation in sprague-dawley rats. J. Mol. Neurosci. MN 2021, 71, 89–100. [Google Scholar] [CrossRef]
- Ding, Y.; Ren, J.; Yu, H.; Yu, W.; Zhou, Y. Porphyromonas gingivalis, a periodontitis causing bacterium, induces memory impairment and age-dependent neuroinflammation in mice. Immun. Ageing I A 2018, 15, 6. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Casiano, C.A.; Fletcher, H.M. Protease-active extracellular protein preparations from porphyromonas gingivalis w83 induce n-cadherin proteolysis, loss of cell adhesion, and apoptosis in human epithelial cells. J. Periodontol. 2001, 72, 641–650. [Google Scholar] [CrossRef]
- Desta, T.; Graves, D.T. Fibroblast apoptosis induced by porphyromonas gingivalis is stimulated by a gingipain and caspase-independent pathway that involves apoptosis-inducing factor. Cell Microbiol. 2007, 9, 2667–2675. [Google Scholar] [CrossRef] [Green Version]
- Roth, G.A.; Ankersmit, H.J.; Brown, V.B.; Papapanou, P.N.; Schmidt, A.M.; Lalla, E. Porphyromonas gingivalis infection and cell death in human aortic endothelial cells. FEMS Microbiol. Lett. 2007, 272, 106–113. [Google Scholar] [CrossRef] [Green Version]
- You, Y.; Cheng, A.-C.; Wang, M.-S.; Jia, R.-Y.; Sun, K.-F.; Yang, Q.; Wu, Y.; Zhu, D.; Chen, S.; Liu, M.-F.; et al. The suppression of apoptosis by α-herpesvirus. Cell Death Dis. 2017, 8, e2749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Z.-F.; Yu, H.-B.; Wang, C.-M.; Qiang, W.-A.; Wang, S.-P.; Zhang, J.-M.; Yu, H.; Cui, L.; Wu, T.; Li, D.-Q.; et al. Furanodiene induces extrinsic and intrinsic apoptosis in doxorubicin-resistant mcf-7 breast cancer cells via nf-κb-independent mechanism. Front. Pharmacol. 2017, 8, 648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, J.Y.; Kim, H.N.; Kim, Y.R.; Choi, Y.W.; Choi, Y.H.; Lee, J.H.; Shin, H.K.; Choi, B.T. Hexane extract from polygonum multiflorum attenuates glutamate-induced apoptosis in primary cultured cortical neurons. J. Ethnopharmacol. 2013, 145, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Wang, J.; Yao, W.; Chen, X.; Zhang, W. Tsg (2,3,4′,5-tetrahydroxystilbene 2-o-β-d-glucoside) suppresses induction of pro-inflammatory factors by attenuating the binding activity of nuclear factor-κb in microglia. J. Neuroinflamm. 2013, 10, 896. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Ahn, S.M.; Wang, Z.; Choi, Y.W.; Shin, H.K.; Choi, B.T. Neuroprotective effects of 2,3,5,4′-tetrahydoxystilbene-2-o-β-d-glucoside from polygonum multiflorum against glutamate-induced oxidative toxicity in ht22 cells. J. Ethnopharmacol. 2017, 195, 64–70. [Google Scholar] [CrossRef]
- Stathopoulou, P.G.; Galicia, J.C.; Benakanakere, M.R.; Garcia, C.A.; Potempa, J.; Kinane, D.F. Porphyromonas gingivalis induce apoptosis in human gingival epithelial cells through a gingipain-dependent mechanism. BMC Microbiol. 2009, 9, 107. [Google Scholar] [CrossRef] [Green Version]
- Pihlstrom, B.L.; Michalowicz, B.S.; Johnson, N.W. Periodontal diseases. Lancet 2005, 366, 1809–1820. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Desta, T.; Fenton, M.; Graves, D.T.; Amar, S. Cytokine profiling of macrophages exposed to porphyromonas gingivalis, its lipopolysaccharide, or its fima protein. Infect. Immun. 2005, 73, 935–943. [Google Scholar] [CrossRef] [Green Version]
- Park, E.; Na, H.S.; Song, Y.R.; Shin, S.Y.; Kim, Y.M.; Chung, J. Activation of nlrp3 and aim2 inflammasomes by porphyromonas gingivalis infection. Infect. Immun. 2014, 82, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Qian, J.; Hou, M.; Wu, X.; Dai, C.; Sun, J.; Dong, L. A review on the extraction, purification, detection, and pharmacological effects of 2,3,5,4′-tetrahydroxystilbene-2-o-β-d-glucoside from polygonum multiflorum. Biomed. Pharmacother. 2020, 124, 109923. [Google Scholar] [CrossRef]
- Choi, B.Y.; Kim, J.H.; Kho, A.R.; Kim, I.Y.; Lee, S.H.; Lee, B.E.; Choi, E.; Sohn, M.; Stevenson, M.; Chung, T.N.; et al. Inhibition of nadph oxidase activation reduces eae-induced white matter damage in mice. J. Neuroinflamm. 2015, 12, 104. [Google Scholar] [CrossRef] [Green Version]
- Boshtam, M.; Kouhpayeh, S.; Amini, F.; Azizi, Y.; Najaflu, M.; Shariati, L.; Khanahmad, H. Anti-inflammatory effects of apocynin: A narrative review of the evidence. All Life 2021, 14, 997–1010. [Google Scholar] [CrossRef]
- Stefanska, J.; Sarniak, A.; Wlodarczyk, A.; Sokolowska, M.; Pniewska, E.; Doniec, Z.; Nowak, D.; Pawliczak, R. Apocynin reduces reactive oxygen species concentrations in exhaled breath condensate in asthmatics. Exp. Lung Res. 2012, 38, 90–99. [Google Scholar] [CrossRef] [PubMed]
- t Hart, B.A.; Copray, S.; Philippens, I. Apocynin, a low molecular oral treatment for neurodegenerative disease. BioMed Res. Int. 2014, 2014, 298020. [Google Scholar]
- Wang, Y.; Shen, Y.; Yu, X.; Gu, J.; Zhang, X.; Zhou, B.; Sun, Y.; Xu, C.; Qian, S. Role of nadph oxidase-induced hypoxia-induced factor-1α increase in blood-brain barrier disruption after 2-hour focal ischemic stroke in rat. Neural Plast. 2021, 2021, 9928232. [Google Scholar] [CrossRef]
- Elahi, S.; Vega-López, M.A.; Herman-Miguel, V.; Ramírez-Estudillo, C.; Mancilla-Ramírez, J.; Motyka, B.; West, L.; Oyegbami, O. Cd71(+) erythroid cells in human neonates exhibit immunosuppressive properties and compromise immune response against systemic infection in neonatal mice. Front. Immunol. 2020, 11, 597433. [Google Scholar] [CrossRef]
- Hou, L.; Sun, F.; Huang, R.; Sun, W.; Zhang, D.; Wang, Q. Inhibition of nadph oxidase by apocynin prevents learning and memory deficits in a mouse parkinson’s disease model. Redox Biol. 2019, 22, 101134. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, M.; Rosen, R.T.; Ho, C.T. 2,2-diphenyl-1-picrylhydrazyl radical-scavenging active components from polygonum multiflorum thunb. J. Agric. Food Chem. 1999, 47, 2226–2228. [Google Scholar] [CrossRef]
- Yao, W.; Huang, C.; Sun, Q.; Jing, X.; Wang, H.; Zhang, W. Tetrahydroxystilbene glucoside protects against oxidized ldl-induced endothelial dysfunction via regulating vimentin cytoskeleton and its colocalization with icam-1 and vcam-1. Cell Physiol. Biochem. 2014, 34, 1442–1454. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charoensaensuk, V.; Chen, Y.-C.; Lin, Y.-H.; Ou, K.-L.; Yang, L.-Y.; Lu, D.-Y. 2,3,5,4′-Tetrahydroxystilbene-2-O-β-glucoside Attenuates Reactive Oxygen Species-Dependent Inflammation and Apoptosis in Porphyromonas gingivalis-Infected Brain Endothelial Cells. Antioxidants 2022, 11, 740. https://doi.org/10.3390/antiox11040740
Charoensaensuk V, Chen Y-C, Lin Y-H, Ou K-L, Yang L-Y, Lu D-Y. 2,3,5,4′-Tetrahydroxystilbene-2-O-β-glucoside Attenuates Reactive Oxygen Species-Dependent Inflammation and Apoptosis in Porphyromonas gingivalis-Infected Brain Endothelial Cells. Antioxidants. 2022; 11(4):740. https://doi.org/10.3390/antiox11040740
Chicago/Turabian StyleCharoensaensuk, Vichuda, Yen-Chou Chen, Yun-Ho Lin, Keng-Liang Ou, Liang-Yo Yang, and Dah-Yuu Lu. 2022. "2,3,5,4′-Tetrahydroxystilbene-2-O-β-glucoside Attenuates Reactive Oxygen Species-Dependent Inflammation and Apoptosis in Porphyromonas gingivalis-Infected Brain Endothelial Cells" Antioxidants 11, no. 4: 740. https://doi.org/10.3390/antiox11040740