
Effect of Water Supplementation on Oxidant/Antioxidant Activities and Total Phenol Content in Growing Olives of the Morisca and Manzanilla Varieties


Abstract
:1. Introduction
2. Materials and Methods
2.1. Olive Sampling and Sample Preparation
2.2. Determination of the Dry Weight/Fresh Weight Ratio, and the Soluble Amino Acid and Protein Contents
2.3. Total Phenolics, Flavonoids and Phenylpropanoid Glycosides (PPGs)
2.4. Oxidant/Antioxidant Enzyme Activities
2.5. Total Antioxidant Activity Assay (FRAP)
2.6. Data Analysis
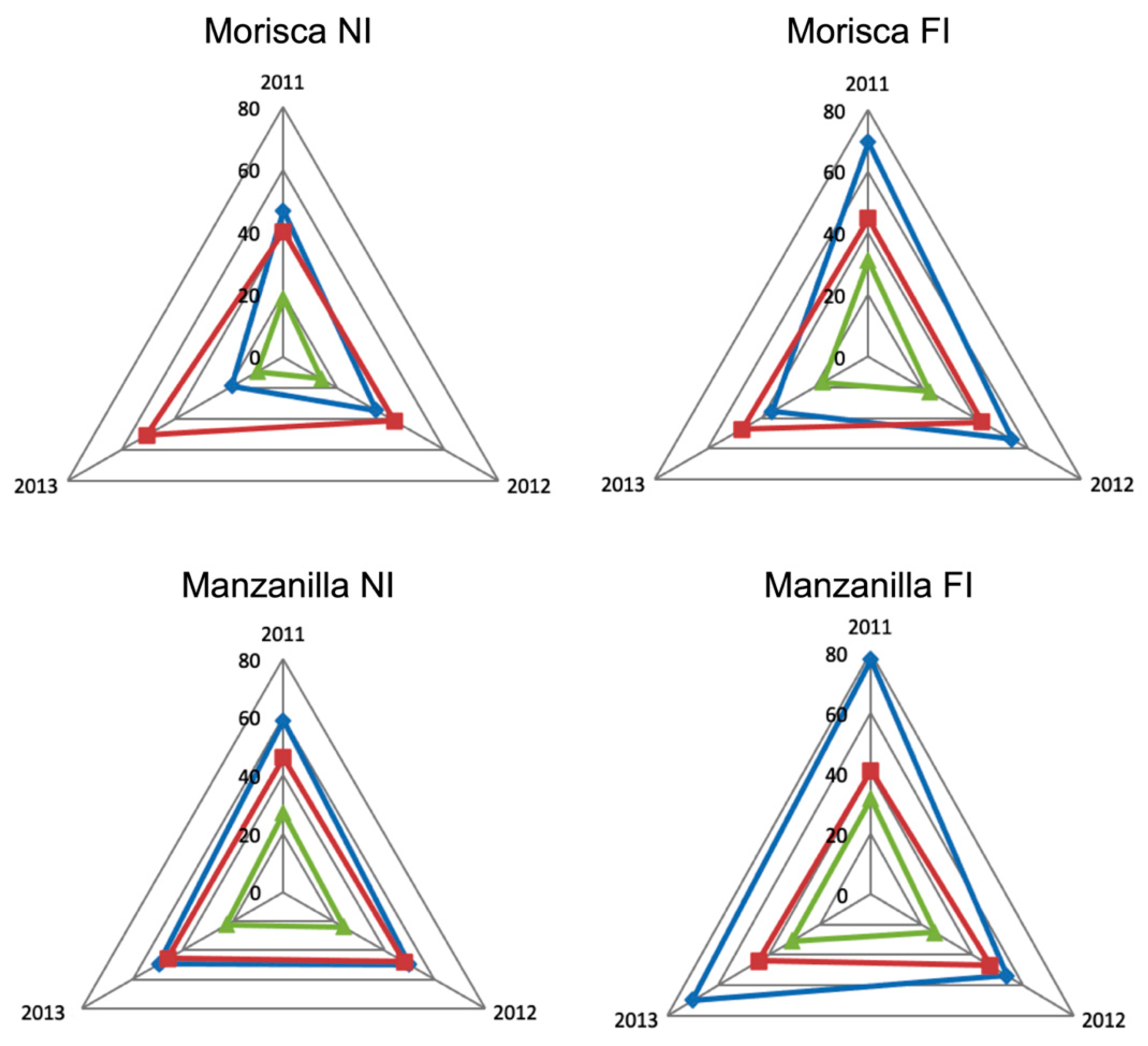
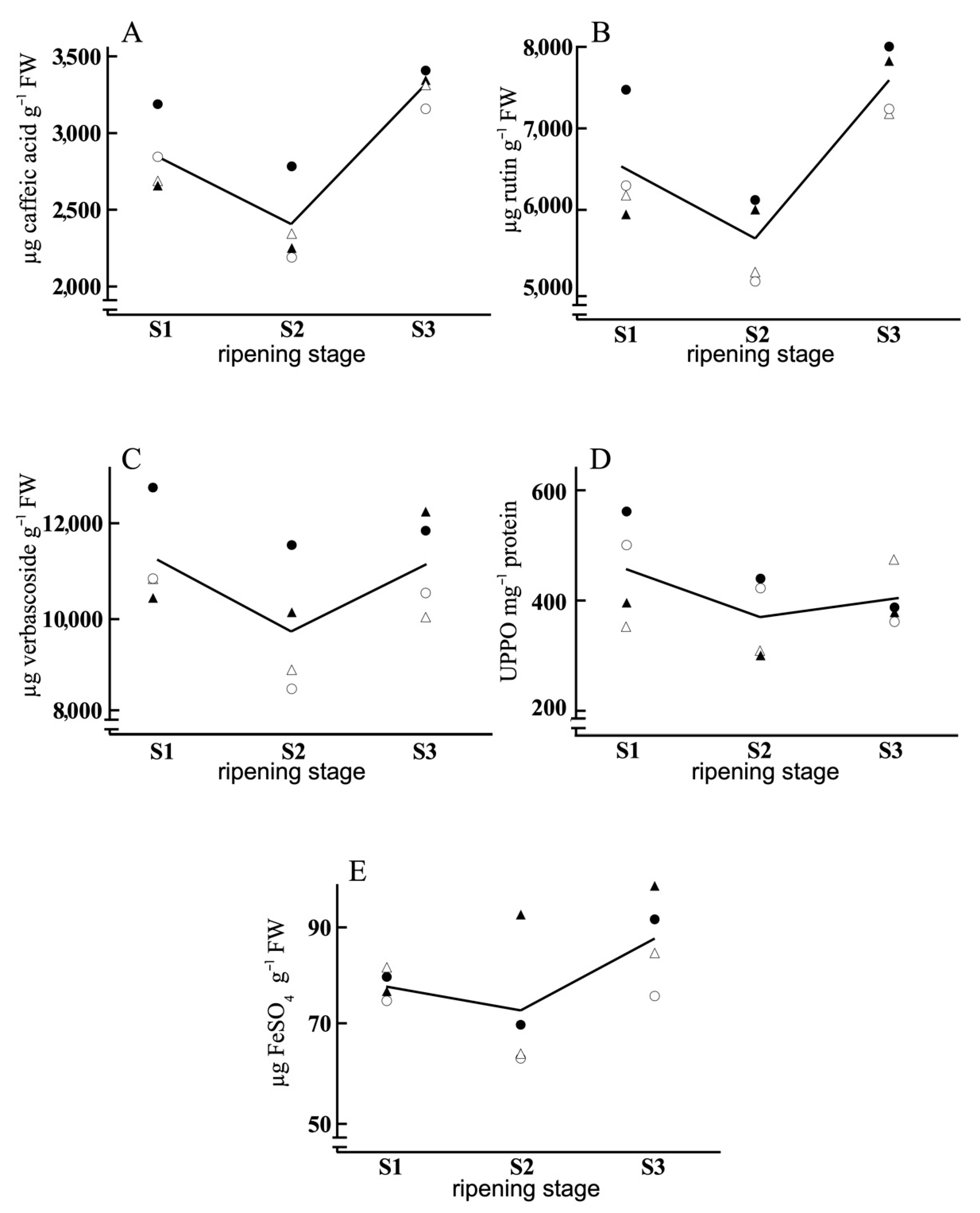
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lucas, L.; Russell, A.; Keast, R. Molecular Mechanisms of Inflammation. Anti-Inflammatory Benefits of Virgin Olive Oil and the Phenolic Compound Oleocanthal. Curr. Pharm. Des. 2011, 17, 754–768. [Google Scholar] [CrossRef] [PubMed]
- Fernández del Río, L.; Gutiérrez-Casado, E.; Varela-López, A.; Villalba, J.M. Olive Oil and the Hallmarks of Aging. Molecules 2016, 21, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rallo, L.; Díez, C.M.; Morales-Sillero, A.; Miho, H.; Priego-Capote, F.; Rallo, P. Quality of olives: A focus on agricultural preharvest factors. Sci. Hort. 2018, 233, 491–509. [Google Scholar] [CrossRef]
- Bendini, A.; Cerretani, L.; Carrasco-Pancorbo, A.; Gómez-Caravaca, A.M.; Segura-Carretero, A.; Fernández-Gutiérrez, A.; Lercker, G. Phenolic Molecules in Virgin Olive Oils: A Survey of Their Sensory Properties, Health Effects, Antioxidant Activity and Analytical Methods. An Overview of the Last Decade. Molecules 2007, 12, 1679–1719. [Google Scholar] [CrossRef]
- Martín-Peláez, S.; Covas, M.I.; Fit, M.; Kušar, A.; Pravst, I. Health Effects of Olive Oil Polyphenols: Recent Advances and Possibilities for the Use of Health Claims. Mol. Nutr. Food Res. 2013, 57, 760–771. [Google Scholar] [CrossRef]
- Servili, M. The phenolic compounds: A comercial argument in the economic war to come on the quality of olive oil? Oilseed Fats Crops Lipids 2014, 21, D509. [Google Scholar] [CrossRef] [Green Version]
- Baccouri, O.; Guerfel, M.; Baccouri, B.; Cerretani, L.; Bendini, A.; Lercker, G.; Zarrouk, M.; Daoud Ben Miled, D. Chemical Composition and Oxidative Stability of Tunisian Monovarietal Virgin Olive Oils with Regard to Fruit Ripening. Food Chem. 2008, 109, 743–754. [Google Scholar] [CrossRef]
- Montaño, A.; Hernández, M.; Garrido, I.; Llerena, J.L.; Espinosa, F. Fatty Acid and Phenolic Compound Concentrations in Eight Different Monovarietal Virgin Olive Oils from Extremadura and the Relationship with Oxidative Stability. Int. J. Mol. Sci. 2016, 17, 1960. [Google Scholar] [CrossRef] [Green Version]
- Guissous, M.; Le Dréau, Y.; Boulkhroune, H.; Madani, T.; Artaud, J. Chemometric Characterization of Eight Monovarietal Algerian Virgin Olive Oils. J. Am. Oil Chem. Soc. 2018, 95, 267–281. [Google Scholar] [CrossRef]
- Špika, M.J.; Perica, S.; Žanetić, M.; Škevin, D. Virgin Olive Oil Phenols, Fatty Acid Composition and Sensory Profile: Can Cultivar Overpower Environmental and Ripening Effect? Antioxidants 2021, 10, 689. [Google Scholar] [CrossRef]
- Rallo, L.; Barranco, D.; Castro-García, S.; Connor, D.J.; Gómez del Campo, M.; Rallo, P. High-density olive plantations. Hort. Rev. 2013, 41, 303–384. [Google Scholar] [CrossRef]
- Rosati, A.; Paoletti, A.; Caporali, S.; Perri, E. The role of tree architecture in super high density olive orchards. Sci. Hortic. 2013, 161, 24–29. [Google Scholar] [CrossRef]
- Díez, C.M.; Moral, J.; Cabello, D.; Morello, P.; Rallo, L.; Barranco, D. Cultivar and Tree Density as Key Factors in the Long-Term Performance of Super High-Density Olive Orchards. Front. Plant Sci. 2016, 7, 1226. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, D.; Sharma, R.; Handa, N.; Kaur, H.; Rattan, A.; Yadav, P.; Gautam, V.; Kaur, R.; Bhardwaj, R. Redox homeostasis in plants under abiotic stress: Role of electron carrieers, energy metabolism mediators and proteinaceous thiols. Front. Environ. Sci. 2015, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Moral, J.; Xaviér, C.; Roca, L.F.; Romero, J.; Moreda, W.; Trapero, A. La Antracnosis del olivo y su efecto en la calidad del aceite. Grasas Y Aceites 2014, 65, e028. [Google Scholar] [CrossRef]
- Romero, A.; Díaz, I.; Tous, J. Optimal harvesting period for ’Arbequina’ olive cultivar in Catalonia (Spain). Acta Hortic. 2002, 586, 393–396. [Google Scholar] [CrossRef]
- Vinha, A.F.; Ferreres, F.; Silva, B.M.; Valentão, P.; Gonçalves, A.; Pereira, J.A.; Oliveira, M.B.; Seabra, R.M.; Andrade, P.B. Phenolic profiles of Portuguese olive fruits (Olea europaea L.): Influences of cultivar and geographical origin. Food Chem. 2005, 89, 561–568. [Google Scholar] [CrossRef]
- Uylaşer, V. Changes in phenolic compounds during ripening in Gemlik variety olive fruits obtained from different locations. CyTA J. Food 2005, 13, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Conde, C.; Delrot, S.; Geroś, H. Physiological, biochemical and molecular changes occurring during olive development and ripening. J. Plant Physiol. 2008, 165, 1545–1562. [Google Scholar] [CrossRef]
- Kumar, V.; Irfan, M.; Ghosh, S.; Chakraborty, N.; Chakraborty, S.; Datta, A. Fruit ripening mutants reveal cell metabolism and redox state during ripening. Protoplasma 2016, 253, 581–594. [Google Scholar] [CrossRef]
- Sáinz, J.A.; Garrido, I.; Hernández, M.; Montaño, A.; Llerena, J.L.; Espinosa, F. Influence of cultivar, irrigation, ripening stage, and annual variability on the oxidant/antioxidant systems of olives as determined by MDS-PTA. PLoS ONE 2019, 14, e0215540. [Google Scholar] [CrossRef] [PubMed]
- López-Huertas, E.; Palma, J.M. Changes in Glutathione, Ascorbate, and Antioxidant Enzymes during Olive Fruit Ripening. J. Agric. Food Chem. 2020, 68, 12221–12228. [Google Scholar] [CrossRef] [PubMed]
- Uceda, M.; Frias, L. Trend of the quality and quantitative composition of olive fruit oil during ripening. In Proceedings of the International Meeting on Olive Oil, Cordoba, Spain, 6–7 October 1975; pp. 25–46. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Yemm, E.W.; Cooking, E.C. The determination of amino acids with ninhidrin. Analyst 1955, 80, 209–213. [Google Scholar] [CrossRef]
- Singleton, V.L.; Salgues, M.; Zaya, J.; Troudsale, E. Caftaric acid disappearance and conversion to products of enzymatic oxidation in grape must and wine. Am. J. Enol. Viticult. 1985, 36, 50–56. [Google Scholar]
- Kim, D.; Jeong, S.W.; Leo, C.Y. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem. 2003, 81, 321–326. [Google Scholar] [CrossRef]
- Gálvez, M.; Martín-Cordero, C.; Houghton, P.J.; Ayuso, M.J. Antioxidant activity of methanol extracts obtained from Plantago species. J. Agric. Food Chem. 2008, 53, 1927–1933. [Google Scholar] [CrossRef]
- Rubinstein, B.; Stern, A.I.; Stout, R. Redox Activity at the Surface of Oat Root Cells. Plant Physiol. 1984, 76, 386–391. [Google Scholar] [CrossRef] [Green Version]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Garrido, I.; García-Sánchez, M.; Casimiro, I.; Casero, P.J.; García-Romera, I.; Ocampo, J.A.; Espinosa, F. Oxidative stress induced in sunflower seedling roots by aqueous dry olive-mill residues. PLoS ONE 2012, 7, e46137. [Google Scholar] [CrossRef] [Green Version]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Ngo, T.T.; Lenhoff, H.M. A sensitive and versatile chromogenic assay for peroxidase and peroxidase-coupled reactions. Anal. Biochem. 1980, 195, 389–397. [Google Scholar] [CrossRef]
- Thipyapong, P.; Hunt, M.D.; Steffens, J.C. Systemic wound induction of potato (Solanum tuberosum) polyphenol oxidase. Phytochemistry 1995, 40, 673–676. [Google Scholar] [CrossRef]
- Rios, J.; Rosales, M.; Blasco, M.B.; Cervilla, L.M.; Romero, L.; Ruiz, J.M. Biofortification of Se and induction of the antioxidant capacity in lettuce plants. Sci. Hort. 2008, 116, 248–255. [Google Scholar] [CrossRef]
- Brockwell, P.J.; Davis, R.A. Introduction to Time Series and Forecasting, 3rd ed.; Springer Texts in Statistics; Springer: New York, NY, USA, 2016. [Google Scholar] [CrossRef]
- Fernandes-Silva, A.A.; Ferreira, T.C.; Correia, C.M.; Malheiro, A.C.; Villalobos, F.J. Influence of different irrigation regimes on crop yield and water use efficiency of olive. Plant Soil 2010, 333, 35–47. [Google Scholar] [CrossRef] [Green Version]
- Cirilli, M.; Caruso, G.; Gennai, C.; Urbani, S.; Frioni, E.; Ruzzi, M.; Servili, M.; Gucci, R.; Poerio, E.; Muleo, R. The role of polyphenoloxidase, peroxidase, and ß-glucosidase in phenolics accumulation in Olea europea L. Fruits under different water regimes. Front. Plant Sci. 2017, 8, 717. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Rico, A.; Salvador, M.D.; Moriana, A.; Pérez, D.; Olmedilla, N.; Ribas, F.; Fregapane, G. Influence of different irrigation strategies in a traditional Cornicabra cv. olive orchard on virgin olive oil composition and quality. Food Chem. 2007, 100, 568–578. [Google Scholar] [CrossRef]
- Tura, D.; Gigliotti, C.; Pedò, S.; Failla, O.; Bassi, D.; Serraiocco, A. Influence of cultivar and site of cultivation on levels of lipophilic and hydrophilic antioxidants in virgin olive oils (Olea europea L.) and correlations with oxidative stability. Sci. Hortic. 2007, 112, 108–119. [Google Scholar] [CrossRef]
- Pandey, V.P.; Singh, S.; Jaiswal, N.; Awasthi, M.; Pandey, B.; Dwivedi, U.N. Papaya fruit ripening: ROS metabolism, gene cloning, characterization and molecular docking of peroxidase. J. Mol. Catal. B Enzym. 2013, 98, 98–105. [Google Scholar] [CrossRef]
- Hachicha Hbaieb, R.; Kotti, F.; García-Rodríguez, R.; Gargouri, M.; Sanz, C.; Pérez, A.G. Monitoring endogenous enzymes during olive fruit ripening and storage: Correlation with virgin olive oil phenolic profiles. Food Chem. 2015, 174, 240–247. [Google Scholar] [CrossRef] [Green Version]
- Huan, C.; Jiang, L.; An, X.; Yu, M.; Xu, Y.; Ma, R.; Yu, Z. Potential role of reactive oxygen species and antioxidant genes in the regulation of peach fruit development and ripening. Plant Physiol. Biochem. 2016, 104, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Xi, F.-F.; Guo, L.-L.; Yu, Y.-H.; Wang, Y.; Li, O.; Zhao, H.-L.; Zhang, G.-H.; Guo, D.-L. Comparison of reactive oxygen species metabolism during grape berry development between ‘Kyoho’ and its early ripening bud mutant ‘Fengzao’. Plant Physiol. Biochem. 2017, 118, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjum, N.A.; Gill, S.S.; Gill, R.; Hasanuzzaman, M.; Duarte, A.C.; Pereira, E.; Ahmad, I.; Tuteja, R.; Tuteja, N. Metal/metalloid stress tolerance in plants: Role of ascorbate, its redox couple, and associated enzymes. Protoplasma 2014, 251, 1265–1283. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Trchounian, A.; Petrosyan, M.; Sahakyan, N. Redox State as a Central Regulator of Plant Cell Stress Response; Gupta, D.K., Palma, J.M., Corpas, F.J., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 25–50. [Google Scholar]
- Waliszewski, K.N.; Márquez, O.; Pardio, V.T. Quantification and characterization of polyphenol oxidase from vanilla bean. Food Chem. 2009, 117, 196–203. [Google Scholar] [CrossRef]
- Takahama, U.; Oniki, T. Flavonoids and some other phenolics as substrates of peroxidase: Physiological significance of the redox reactions. J. Plant Res. 2000, 113, 301–309. [Google Scholar] [CrossRef]
- García-Rodríguez, R.; Romero-Segura, C.; Sanz, C.; Sánchez-Ortiz, A.; Pérez, A.G. Role of polyphenol oxi- dase and peroxidase in shaping the phenolic profile of virgin olive oil. Food Res. Int. 2011, 44, 629–635. [Google Scholar] [CrossRef]
- Luaces, P.; Romero, C.; Gutiérrez, F.; Sanz, C.; Pérez, A.G. Contribution of olive seed to the phenolic profile and related quality parameters of virgin olive oil. J. Sci. Food Agric. 2007, 87, 2721–2727. [Google Scholar] [CrossRef]
- Zamora, R.; Alaiz, M.; Hidalgo, F.J. Influence of cultivar and fruit ripening on olive (Olea europaea) fruit protein content, composition, and antioxidant activity. J. Agric. Food Chem. 2001, 49, 4267–4270. [Google Scholar] [CrossRef]
- Ebrahimzdeh, H.; Motamed, N.; Rastgar-Jazii, F.; Montasser-Kouhsari, S.; Shokraii, E.H. Oxidative enzyme activities and soluble protein content in leaves and fruits of olives during ripening. J. Food Biochem. 2003, 27, 181–196. [Google Scholar] [CrossRef]
- Ortega-García, F.; Blanco, S.; Peinado, M.A.; Peragón, J. Polyphenol oxidase and its relationship with oleur- opein concentration in fruits and leaves of olive (Olea europaea) cv. ’Picual’ trees during fruit ripening. Tree Physiol. 2008, 28, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Kan, J.; Wang, H.M.; Jin, C.-H.; Xie, H.-Y. Changes of Reactive Oxygen Species and Related Enzymes in Mitochondria Respiratory Metabolism During the Ripening of Peach Fruit. Agric. Sci. China 2010, 9, 138–146. [Google Scholar] [CrossRef]
- Kumar, N.; Ebel, R.C.; Roberts, P.D. H2O2 degradation is suppressed in kumquat leaves infected with Xanthomonas axonopodis pv. Citri. Sci. Hortic. 2011, 130, 241–247. [Google Scholar] [CrossRef]
- Sgherri, C.; Cosi, E.; Navari-Izzo, F. Phenols and antioxidative status of Raphanus sativus grown in copper excess. Physiol. Plant. 2003, 118, 21–28. [Google Scholar] [CrossRef]
- Kocsy, G.; Tari, I.; Vanková, R.; Zechmann, B.; Gulyás, Z.; Poór, P.; Galiba, G. Redox control of plant growth and development. Plant Sci. 2013, 211, 77–91. [Google Scholar] [CrossRef]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornero-Méndez, D.; Gallardo-Guerrero, L.; Jarén-Galán, M.; Mínguez-Mosquera, M.I. Differences in the Activity of Superoxide Dismutase, Polyphenol Oxidase and Cu-Zn Content in the Fruits of Gordal and Manzanilla Olive varieties. Z. Nat. C 2002, 57, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Hachicha Hbaieb, R.; Kotti, F.; Gargouri, M.; Msallem, M.; Vichi, S. Ripening and storage conditions of Chétoui and Arbequina olives: Part, I. Effect on olive oils volatiles profile. Food Chem. 2016, 203, 548–558. [Google Scholar] [CrossRef]
- Foyer, C.; Noctor, G. Redox Homeostasis and Antioxidant Signaling: A Metabolic Interface Between Stress Perception and Physiological Responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [Green Version]
- Petridis, A.; Therios, I.; Samouris, G.; Koundouras, S.; Giannakoula, A. Effect of water deficit on leaf phenolic composition, gas exchange, oxidative damage and antioxidant activity of four Greek olive (Olea europaea L.) cultivars. Plant Physiol. Biochem. 2012, 60, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ripa, V.; Rose, F.; Caravita, M.A.; Parise, M.R.; Perri, E.; Rosati, A.; Pandolfi, S.; Paoletti, A.; Pannelli, G.; Padula, G.; et al. Qualitative evaluation of olive oils from new olive selections and effects of genotype and environment on oil quality. Adv. Hortic. Sci. 2008, 22, 95–103. [Google Scholar]
- Cerretani, L.; Bendini, A.; Rotondi, A.; Mari, M.; Lercker, G.; Toschi, T. Evaluation of the oxidative stability and organoleptic properties of extra-virgin olive oils in relation to olive ripening degree. Prog. Nutr. 2004, 6, 50–56. [Google Scholar]
- Ortega-García, F.; Peragón, J. Phenylalanine Ammonia-Lyase, Polyphenol Oxidase, and Phenol Concentration in Fruits of Olea europaea L. cv. Picual, Verdial, Arbequina, and Frantoio during Ripening. J. Agric. Food Chem. 2009, 57, 10331–10340. [Google Scholar] [CrossRef] [PubMed]
- Morelló, J.R.; Vuorela, S.; Romero, M.P.; Motilva, M.J.; Heinonen, M. Antioxidant activity of olive pulp and olive oil phenolic compounds of the arbequina cultivar. J. Agric. Food Chem. 2005, 53, 2002–2008. [Google Scholar] [CrossRef]
- Sobhy El-Sohaimy, A.A.; Mohamed El-Sheikh, H.; Taha Refaay, M.; Mohamed Zaytoun, A.M. Effect of Harvesting in Different Ripening Stages on Olive (Olea europea) Oil Quality. Am. J. Food Technol. 2016, 11, 1–11. [Google Scholar] [CrossRef]
- Arslan, D.; Ozcan, M.M. Phenolic profile and antioxidant activity of olive fruits of the Turkish variety “Sanulak” from different locations. Grasas Aceites 2011, 62, 453–461. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Mean | Cultivar | Irrigation | Ripening |
|---|---|---|---|---|
| DW/FW | 0.30 ± 0.04 | 0.004 | ||
| Total amino acids (mg g−1 FW) | 2866.35 ± 1637.10 | 8.28 × 10−5 | 1.46 × 10−5 | |
| Total protein (mg g−1 FW) | 1.73 ± 0.83 | 4.64 × 10−6 | 1.64 × 10−9 | |
| NADH oxidation (nmol min−1 mg−1 protein) | 730.74 ± 395.62 | 0.039 | ||
| O2− production (nmol min−1 mg−1 protein) | 324.46 ± 174.50 | |||
| SOD activity (USOD mg−1 protein) | 112.93 ± 54.87 | 3.06 × 10−5 | ||
| POX activity (nmol min−1 mg−1 protein) | 113.27 ± 57.17 | 0.002 | ||
| Total phenols (µg caffeic acid g−1 FW) | 2856.15 ± 847.18 | 3.06 × 10−6 | ||
| Total flavonoids (µg rutin g−1 FW) | 6609.48 ± 1571.89 | 0.001 | ||
| Total PPGs (µg verbascoside g−1 FW) | 10,706.68 ± 2572.43 | 0.007 | 0.032 | |
| PPO activity (UPPO mg−1 protein) | 403.37 ± 137.06 | 0.016 | ||
| FRAP (µg FeSO4 g−1 FW) | 79.53 ± 42.16 | 0.031 | 0.002 |
| Parameter | Average | Seasonal Variation Index | ||
|---|---|---|---|---|
| 2011 | 2012 | 2013 | ||
| DW/FW | 0.30 ± 0.02 | 0.01 | 0.00 | −0.02 |
| Total amino acids (mg g−1 FW) | 2866.35 ± 1094.85 | 807.90 | 156.87 | −964.78 |
| Total protein (mg g−1 FW) | 1.73 ± 0.77 | 0.13 | 0.02 | −0.15 |
| NADH oxidation (nmol min−1 mg−1 protein) | 730.74 ± 222.74 | −164.79 | 207.70 | −42.90 |
| O2− production (nmol min−1 mg−1 protein) | 324.46 ± 67.76 | −97.23 | 142.93 | −45.70 |
| SOD activity (USOD mg−1 protein) | 112.93 ± 20.04 | −33.64 | 8.68 | 24.96 |
| POX activity (nmol min−1 mg−1 protein) | 113.27 ± 37.83 | −7.12 | 28.50 | −21.38 |
| Total phenols (µg caffeic acid g−1 FW) | 2856.15 ± 452.13 | −901.98 | 483.20 | 418.78 |
| Total flavonoids (µg rutin g−1 FW) | 6609.48 ± 973.57 | −1132.08 | 965.16 | 166.92 |
| Total PPGs (µg verbascoside g−1 FW) | 10,706.68 ± 884.33 | −2603.04 | 1435.53 | 1167.51 |
| PPO activity (UPPO mg−1 protein) | 403.37 ± 43.72 | 118.41 | −44.77 | −73.64 |
| FRAP (µg FeSO4 g−1 FW) | 79.53 ± 7.56 | −54.16 | 37.31 | 16.86 |
| Flavonoids | PPGs | FRAP | Total Amino Acids | POX | NADH Oxidation | SOD | PPO | DW/FW | Tmin | Tmax | Irrigation | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total phenols | 0.894 (0.001) | 0.889 (0.001) | 0.866 (0.003) | −0.687 (0.041) | −0.869 (0.002) | |||||||
| Total flavonoids | 0.863 (0.003) | 0.729 (0.026) | −0.823 (0.006) | |||||||||
| Total PPGs | 0.872 (0.002) | −0.689 (0.040) | −0.801 (0.009) | |||||||||
| FRAP | −0.829 (0.006) | |||||||||||
| Total protein | 0.838 (0.005) | |||||||||||
| Total amino acids | 0.807 (0.009) | |||||||||||
| O2− production | 0.793 (0.011) | 0.882 (0.002) | ||||||||||
| POX | 0.876 (0.002) | |||||||||||
| NADH oxidation | 0.691 (0.039) | |||||||||||
| DW/FW | −0.483 (0.007) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrido, I.; Hernández, M.; Llerena, J.L.; Espinosa, F. Effect of Water Supplementation on Oxidant/Antioxidant Activities and Total Phenol Content in Growing Olives of the Morisca and Manzanilla Varieties. Antioxidants 2022, 11, 729. https://doi.org/10.3390/antiox11040729
Garrido I, Hernández M, Llerena JL, Espinosa F. Effect of Water Supplementation on Oxidant/Antioxidant Activities and Total Phenol Content in Growing Olives of the Morisca and Manzanilla Varieties. Antioxidants. 2022; 11(4):729. https://doi.org/10.3390/antiox11040729
Chicago/Turabian StyleGarrido, Inmaculada, Marcos Hernández, José Luis Llerena, and Francisco Espinosa. 2022. "Effect of Water Supplementation on Oxidant/Antioxidant Activities and Total Phenol Content in Growing Olives of the Morisca and Manzanilla Varieties" Antioxidants 11, no. 4: 729. https://doi.org/10.3390/antiox11040729