
Genome-Wide Identification of the Bcl-2 Associated Athanogene (BAG) Gene Family in Solanum lycopersicum and the Functional Role of SlBAG9 in Response to Osmotic Stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome-Wide Identification and Analysis of the Tomato BAG Gene Family
2.2. Phylogenetic Analysis
2.3. Motif Analysis of SlBAG Proteins
2.4. Gene Structure and Cis-Element Analysis of SlBAG Genes
2.5. Expression Analysis of SlBAG Genes
2.6. Phenotypic Analysis of SlBAG9 Overexpression Plants under Osmotic Stress
2.7. H2O2 Content, Lipid Peroxidation, and Antioxidant Enzymes
2.8. RNA Isolation and qRT-PCR
3. Results
3.1. Genome-Wide Identification of BAG Gene Family in Tomato
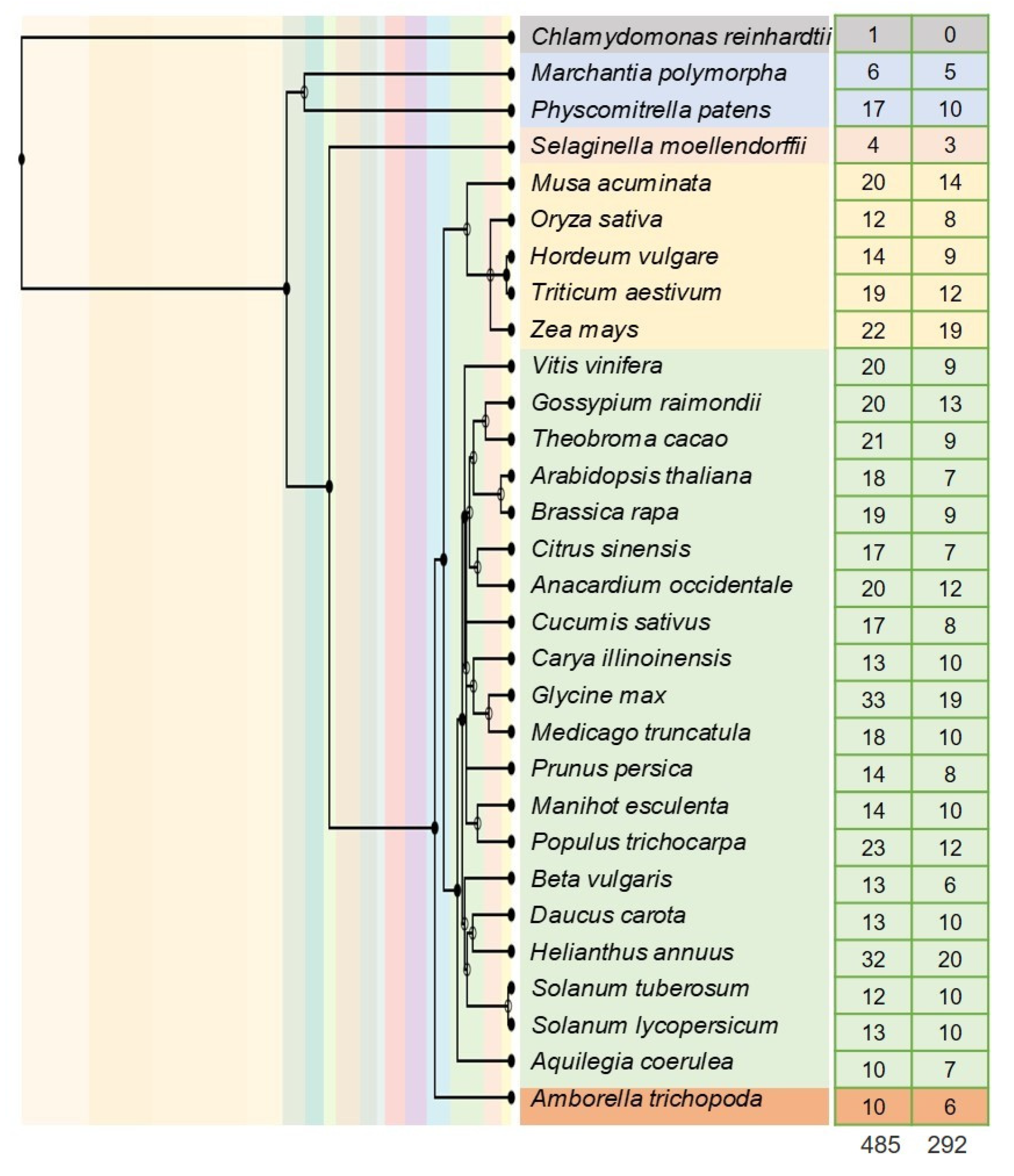
3.2. Phylogenetic Characterization of BAG Family Genes in Tomato
3.3. Exon–Intron Arrangement and Conserved Motifs Analysis
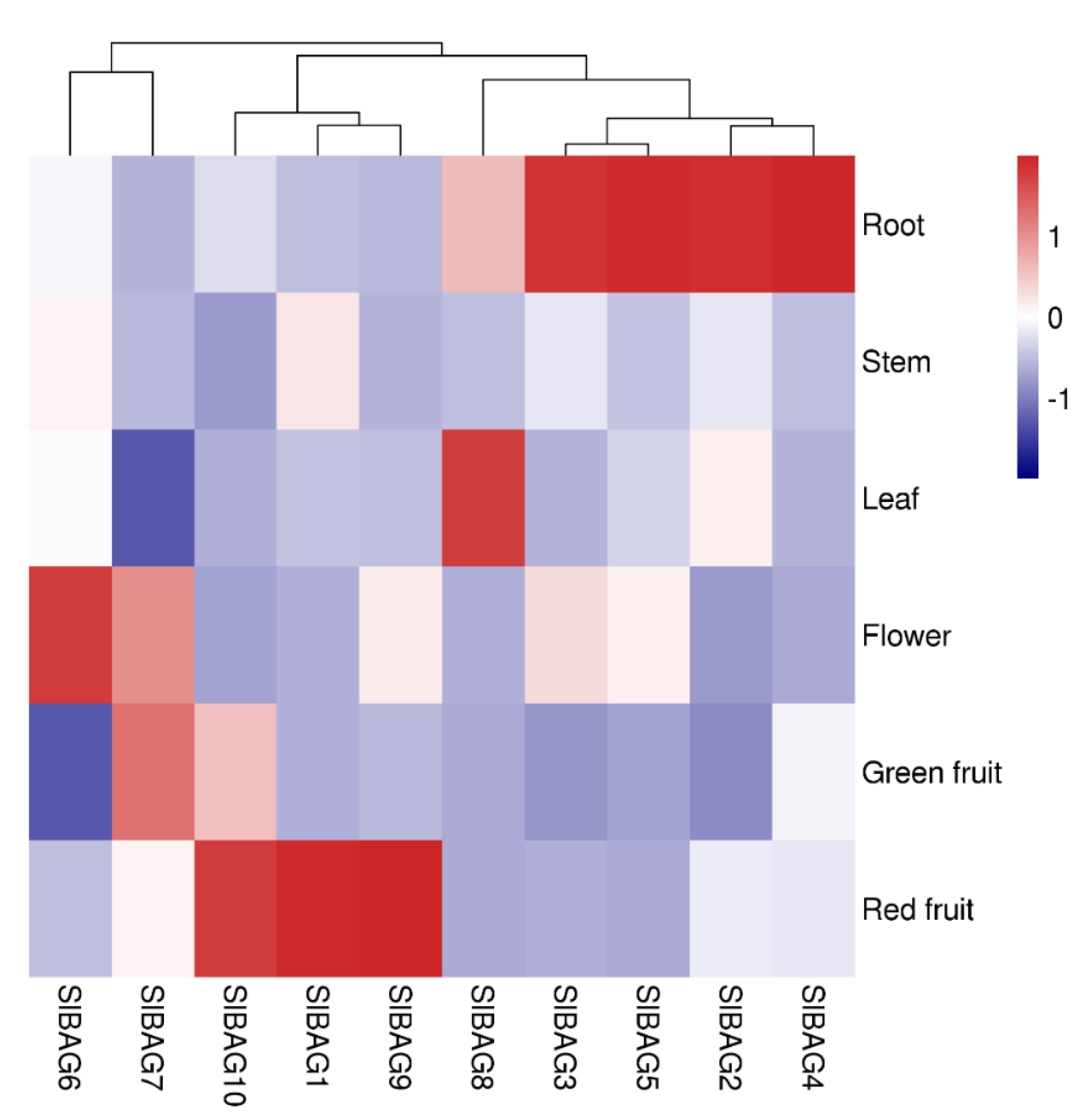
3.4. Organ-Specific Expression Profiles of SlBAG Genes
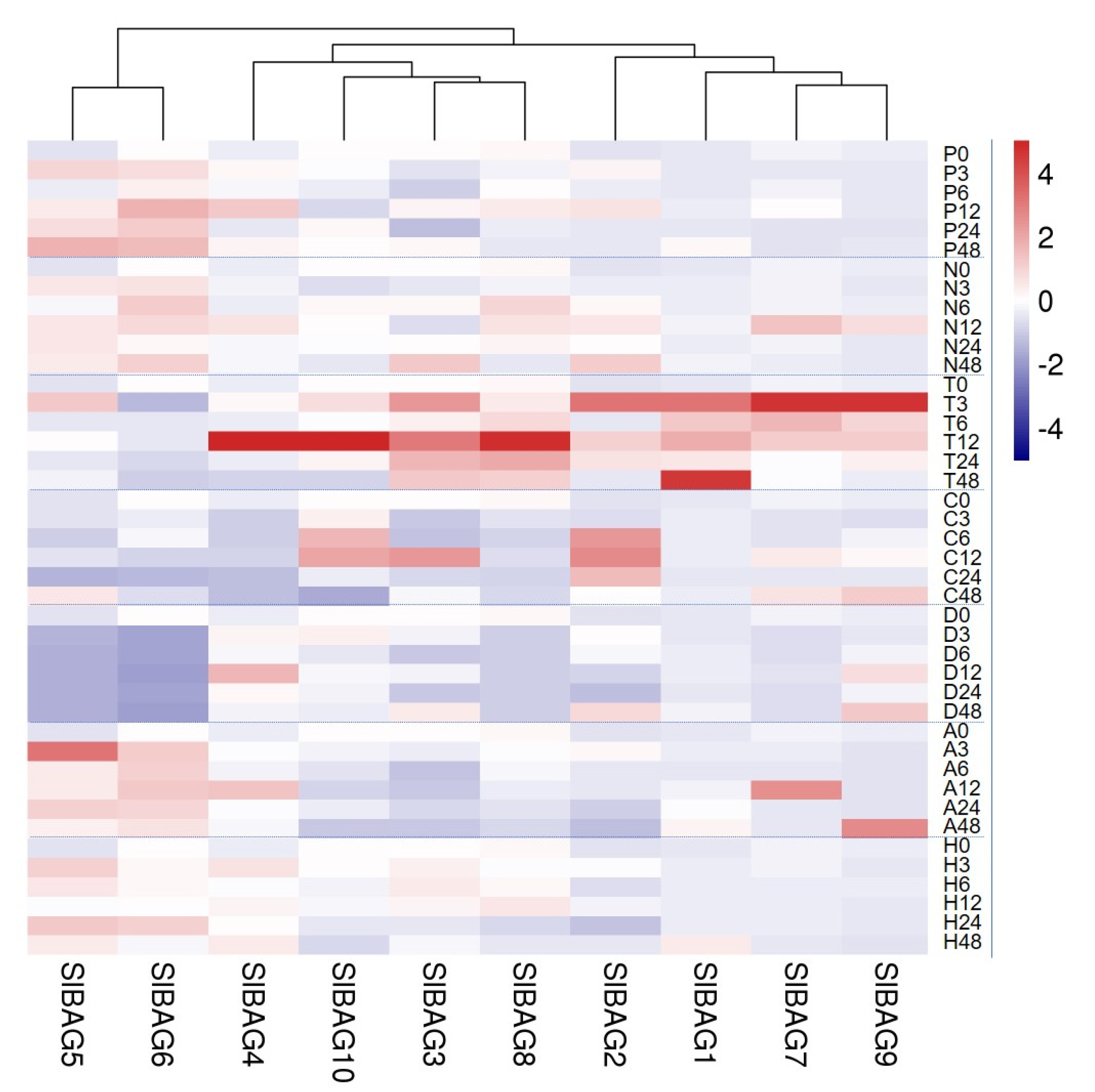
3.5. Expression Pattern of SlABG Genes under Abiotic Stress
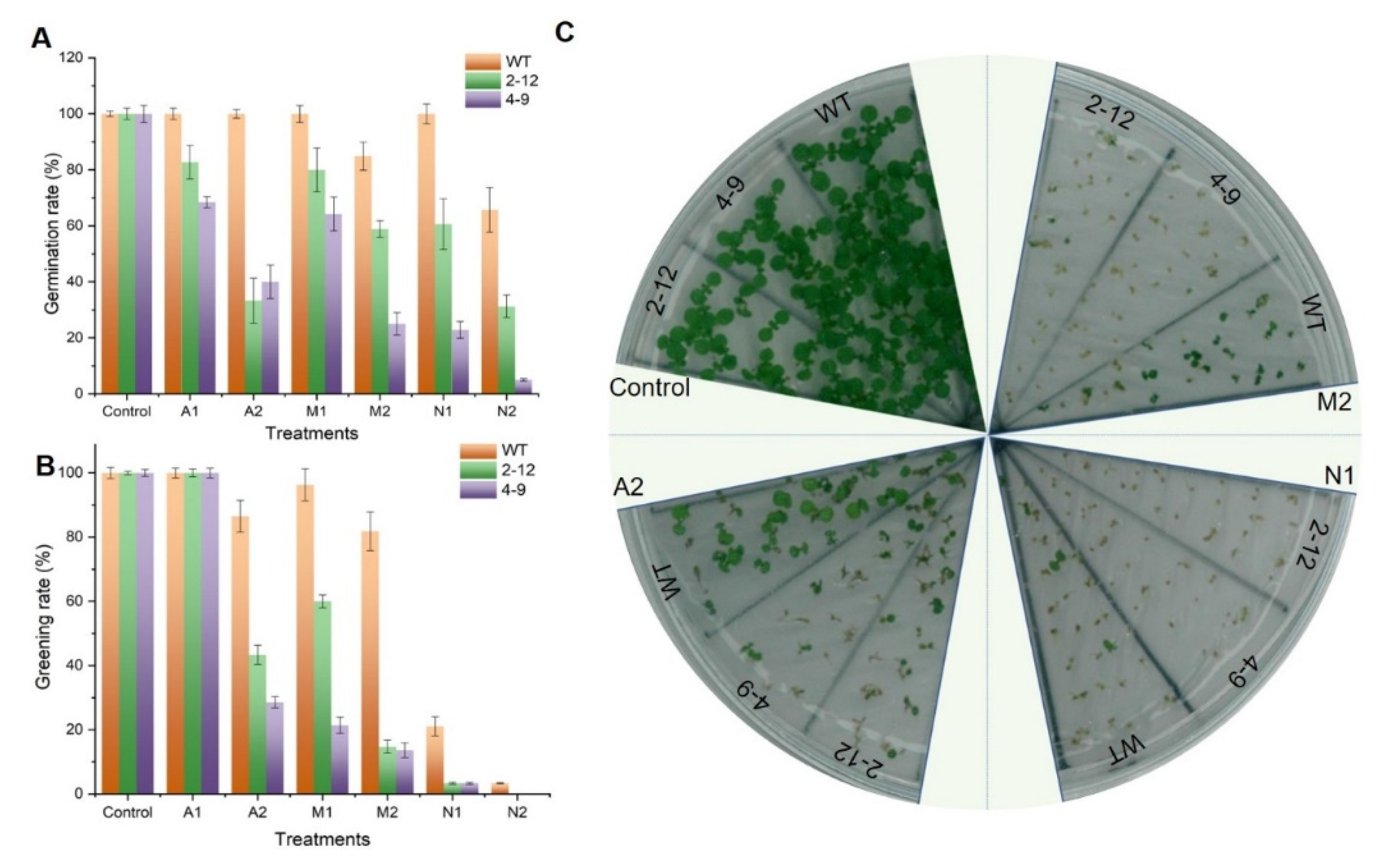
3.6. Heterologous Expression of SlBAG9 Conferred Sensitivity to Drought, Salt, and ABA
3.7. SlBAG9 Downregulated Stress/ABA-Responsive Genes
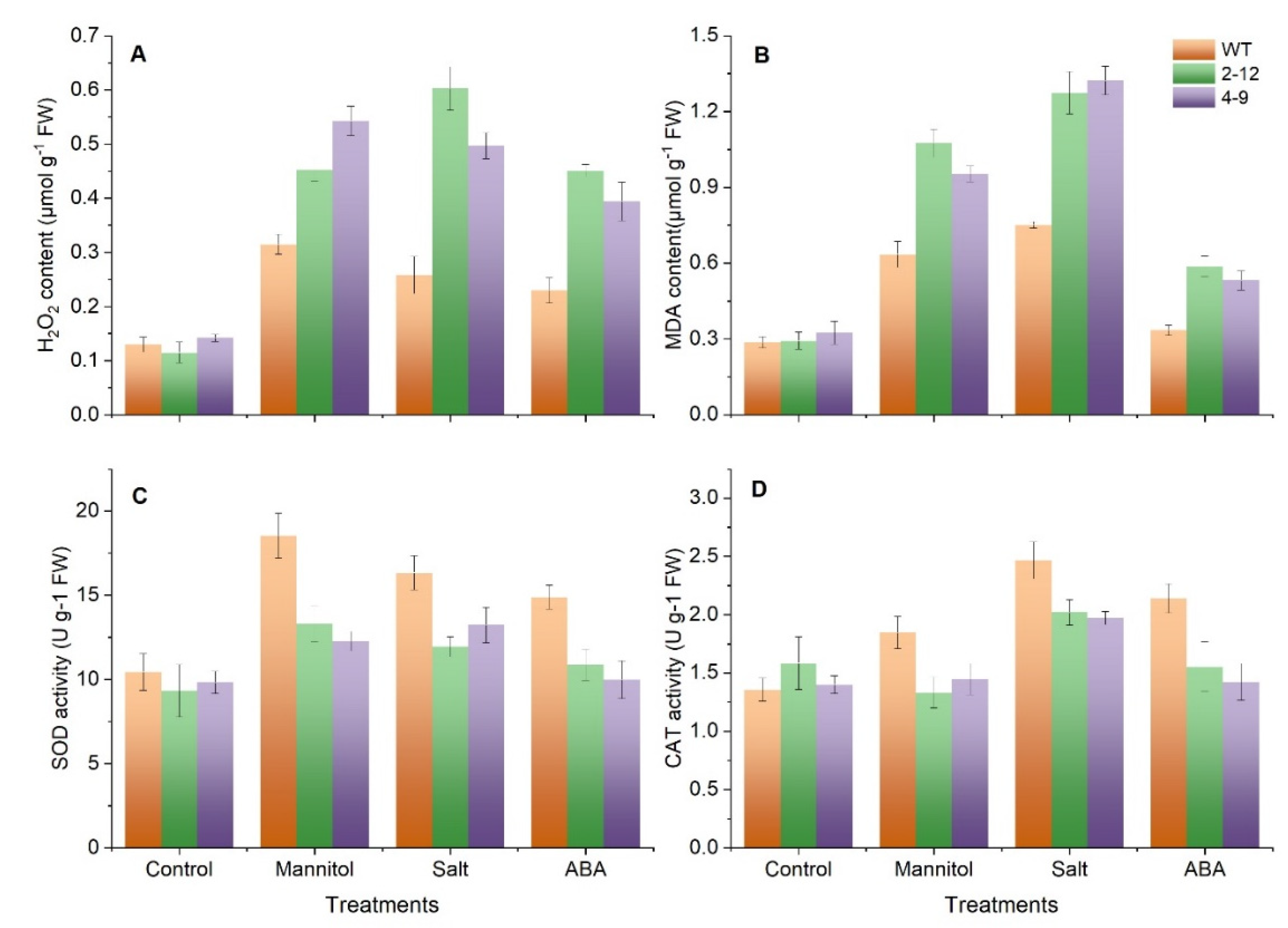
3.8. SlBAG9 Overexpression Aggravated Oxidative Damage under Drought, Salt, and ABA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Takayama, S.; Sato, T.; Krajewski, S.; Kochel, K.; Irie, S.; Millan, J.A.; Reed, J.C. Cloning and functional analysis of BAG-1: A novel Bcl-2-binding protein with anti-cell death activity. Cell 1995, 80, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Kabbage, M.; Dickman, M.B. The BAG proteins: A ubiquitous family of chaperone regulators. Cell Mol. Life Sci. 2008, 65, 1390–1402. [Google Scholar] [CrossRef] [PubMed]
- Doukhanina, E.V.; Chen, S.; van der Zalm, E.; Godzik, A.; Reed, J.; Dickman, M.B. Identification and functional characterization of the BAG protein family in Arabidopsis thaliana. J. Biol. Chem. 2006, 281, 18793–18801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawkar, G.M.; Maibam, P.; Park, J.H.; Woo, S.G.; Kim, C.Y.; Lee, S.Y.; Kang, C.H. In silico study on Arabidopsis BAG gene expression in response to environmental stresses. Protoplasma 2017, 254, 409–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rana, R.M. Identification and characterization of the Bcl−2-associated athanogene (BAG) protein family in rice. Afr. J. Biotechnol. 2012, 11, 88–98. [Google Scholar]
- Ge, S.; Kang, Z.; Li, Y.; Zhang, F.; Shen, Y.; Ge, R.; Huang, Z. Cloning and function analysis of BAG family genes in wheat. Funct. Plant Biol. 2016, 43, 393–402. [Google Scholar] [CrossRef]
- Thanthrige, N.; Jain, S.; Bhowmik, S.D.; Ferguson, B.J.; Kabbage, M.; Mundree, S.; Williams, B. Centrality of BAGs in plant PCD, stress responses, and host defense. Trends Plant Sci. 2020, 25, 1131–1140. [Google Scholar] [CrossRef]
- He, M.; Wang, Y.; Jahan, M.S.; Liu, W.; Raziq, A.; Sun, J.; Su, S.; Guo, S. Characterization of SlBAG genes from Solanum lycopersicum and its function in response to dark-induced leaf senescence. Plants 2021, 10, 947. [Google Scholar] [CrossRef]
- Yan, J.; He, C.; Zhang, H. The BAG-family proteins in Arabidopsis thaliana. Plant Sci. 2003, 165, 1–7. [Google Scholar] [CrossRef]
- Lee, D.W.; Kim, S.J.; Oh, Y.J.; Choi, B.; Lee, J.; Hwang, I. Arabidopsis BAG1 functions as a cofactor in Hsc70-mediated proteasomal degradation of unimported plastid proteins. Mol. Plant 2016, 9, 1428–1431. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Xing, Y.; Chang, D.; Fang, S.; Cui, B.; Li, Q.; Wang, X.; Guo, S.; Yang, X.; Men, S.; et al. CaM/BAG5/Hsc70 signaling complex dynamically regulates leaf senescence. Sci. Rep. 2016, 6, 31889. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Li, L.; Kang, H.; Yang, X.; Men, S.; Shen, Y. Chronic mitochondrial calcium elevation suppresses leaf senescence. Biochem. Biophys. Res. Commun. 2017, 487, 672–677. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kabbage, M.; Liu, W.; Dickman, M.B. Aspartyl protease-mediated cleavage of BAG6 is necessary for autophagy and fungal resistance in plants. Plant Cell 2016, 28, 233–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, B.; Kabbage, M.; Britt, R.; Dickman, M.B. AtBAG7, an Arabidopsis Bcl-2–associated athanogene, resides in the endoplasmic reticulum and is involved in the unfolded protein response. Proc. Natl. Acad. Sci. USA 2010, 107, 6088–6093. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Yang, K.; Cheng, M.; Cheng, Y.; Li, Y.; Ai, G.; Bai, T.; Xu, R.; Duan, W.; Peng, H.; et al. Double-faced role of Bcl-2-associated athanogene 7 in plant–Phytophthora interaction. J. Exp. Bot. 2021, 72, 5751–5765. [Google Scholar] [CrossRef]
- Eckardt, N.A. Programmed cell death in plants: A role for mitochondrial associated hexokinases. Plant Cell 2006, 18, 2097–2099. [Google Scholar] [CrossRef] [Green Version]
- You, Q.; Zhai, K.; Yang, D.; Yang, W.; Wu, J.; Liu, J.; Pan, W.; Wang, J.; Zhu, X.; Jian, Y.; et al. An E3 ubiquitin ligase-BAG protein module controls plant innate immunity and broad-spectrum disease resistance. Cell Host Microbe 2016, 20, 758–769. [Google Scholar] [CrossRef] [Green Version]
- Yeckel, G.J. Characterization of a Soybean BAG Gene and Its Potential Role in Nematode Resistance; University of Missouri: Columbia, MO, USA, 2012. [Google Scholar]
- Kobayashi, M.; Takato, H.; Fujita, K.; Suzuki, S. HSG1, a grape Bcl-2-associated athanogene, promotes floral transition by activating CONSTANS expression in transgenic Arabidopsis plant. Mol. Biol. Rep. 2012, 39, 4367–4374. [Google Scholar] [CrossRef]
- Bansal, R.; Kumawat, S.; Dhiman, P.; Sudhakaran, S.; Rana, N.; Jaswal, R.; Singh, A.; Sonah, H.; Sharma, T.R.; Deshmukh, R. Evolution of Bcl-2 Anthogenes (BAG) as the regulators of cell death in wild and cultivated Oryza Species. J. Plant Growth Regul. 2022. [Google Scholar] [CrossRef]
- Castro, A.; Saavedra, L.L.; Ruibal, C.; Lascano, R.; Vidal, S. Genome-wide identification, characterization and expression analysis of the Bcl-2 associated athanogene (BAG) gene family in Physcomitrium Patens. bioRxiv 2020. [Google Scholar] [CrossRef]
- Ding, H.; Mo, S.; Qian, Y.; Yuan, G.; Wu, X.; Ge, C. Integrated proteome and transcriptome analyses revealed key factors involved in tomato (Solanum lycopersicum) under high-temperature stress. Food Energy Secur. 2020, 9, e239. [Google Scholar] [CrossRef]
- Ding, H.; He, J.; Wu, Y.; Wu, X.X.; Ge, C.; Wang, Y.; Xu, W. The tomato mitogen-activated protein kinase SlMPK1 is as a negative regulator of the high temperature stress response. Plant Physiol. 2018, 177, 633–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irfan, M.; Kumar, P.; Ahmad, I.; Datta, A. Unraveling the role of tomato Bcl-2-associated athanogene (BAG) proteins during abiotic stress response and fruit ripening. Sci. Rep. 2021, 11, 21734. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Nan, N.; Li, N.; Liu, Y.; Wang, T.J.; Hwang, I.; Liu, B.; Xu, Z.Y. A DNA methylation reader-chaperone regulator transcription factor complex activates OsHKT1;5 expression during salinity stress. Plant Cell 2020, 32, 3535–3558. [Google Scholar] [CrossRef]
- Gao, S.; Gao, J.; Zhu, X.; Song, Y.; Li, Z.; Ren, G.; Zhou, X.; Kuai, B. ABF2, ABF3, and ABF4 promote ABA-mediated chlorophyll degradation and leaf senescence by transcriptional activation of chlorophyll catabolic genes and senescence-associated genes in Arabidopsis. Mol. Plant 2016, 9, 1272–1285. [Google Scholar] [CrossRef] [Green Version]
- Verslues, E.; Agarwal, M.; Katiyar-Agarwal, J.; Zhu, S.; Zhu, J.K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 2006, 45, 523–539. [Google Scholar] [CrossRef]
- Jambunathan, N. Determination and detection of reactive oxygen species (ROS), lipid peroxidation, and electrolyte leakage in plants. Methods Mol. Biol. 2010, 639, 292–298. [Google Scholar] [PubMed]
- Ding, H.; Qian, L.; Jiang, H.; Ji, Y.; Fang, Y.; Sheng, J.; Xu, X.; Ge, C. Overexpression of a Bcl-2-associated athanogene SlBAG9 negatively regulates high-temperature response in tomato. Int. J. Biol. Macromol. 2022, 194, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Alberti, S.; Demand, J.; Esser, C.; Emmerich, N.; Schild, H.; Hohfeld, J. Ubiquitylation of BAG-1 suggests a novel regulatory mechanism during the sorting of chaperone substrates to the proteasome. J. Biol. Chem. 2002, 277, 45920–45927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabbage, M.; Kessens, R.; Dickman, M.B. A plant Bcl-2- associated athanogene is proteolytically activated to confer fungal resistance. Microb. Cell 2016, 3, 224–226. [Google Scholar] [CrossRef] [Green Version]
- Kang, C.H.; Jung, W.Y.; Kang, Y.H.; Kim, J.Y.; Kim, D.G.; Jeong, J.C.; Baek, D.W.; Jin, J.B.; Lee, J.Y.; Kim, M.O.; et al. AtBAG6, a novel calmodulin-binding protein, induces programmed cell death in yeast and plants. Cell Death Differ. 2006, 13, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Arif, M.; Li, Z.; Luo, Q.; Li, L.; Shen, Y.; Men, S. The BAG2 and BAG6 genes are involved in multiple abiotic stress tolerances in Arabidopsis thaliana. Int. J. Mol. Sci. 2021, 22, 5856. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Li, L.; Cui, B.; Men, S.; Shen, Y.; Yang, X. Structural insight into plant programmed cell death mediated by BAG proteins in Arabidopsis thaliana. Acta Crystallogr. D Biol. Crystallogr. 2013, 69, 934–945. [Google Scholar] [CrossRef]
- Danino, Y.M.; Even, D.; Ideses, D.; Juven-Gershon, T. The core promoter: At the heart of gene expression. Biochim. Biophys. Acta 2015, 1849, 1116–1131. [Google Scholar] [CrossRef]
- Li, Y.; Williams, B.; Dickman, M. Arabidopsis B-cell lymphoma2 (Bcl-2)-associated athanogene 7 (BAG 7)-mediated heat tolerance requires translocation, sumoylation and binding to WRKY 29. New Phytol. 2017, 214, 695–705. [Google Scholar] [CrossRef]
- Hoang, T.M.L.; Lalehvash, M.; Brett, W.; Harjeet, K.; James, D.; Mundree, S.G. Development of salinity tolerance in rice by constitutive overexpression of genes involved in the regulation of programmed cell death. Front. Plant Sci. 2015, 6, 175. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.J.; Liu, L.; Lin, Y.C.; Zu, Y.G.; Li, L.P.; Tang, Z.H. Ethylene antagonizes saltinduced growth retardation and cell death process via transcriptional controlling of ethylene-, BAG- and senescence- associated genes in Arabidopsis. Front. Plant Sci. 2016, 7, 696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locascio, A.; Marqués, M.C.; García-Martínez, G.; Corratgé-Faillie, C.; Andrés-Colás, N.; Rubio, L.; Fernández, J.A.; Véry, A.A.; Mulet, J.M.; Yenush, L. Bcl-2-associated athanogene regulates the KAT1 potassium channel and controls stomatal movement. Plant Physiol. 2019, 181, 1277–1294. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Hu, X.; Song, J.; Zong, X.; Li, D.; Li, D. Over-expression of a Zea mays L. protein phosphatase 2C gene (ZmPP2C) in Arabidopsis thaliana decreases tolerance to salt and drought. J. Plant Physiol. 2009, 166, 531–542. [Google Scholar] [CrossRef]
- Bedi, S.; Sengupta, S.; Ray, A.; Chaudhuri, R.N. ABI3 mediates dehydration stress recovery response in Arabidopsis thaliana by regulating expression of downstream genes. Plant Sci. 2016, 250, 125–140. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, W.; Gao, X.; Xu, M.; Guo, Y. DEAR4, a member of DREB/CBF family, positively regulates leaf senescence and response to multiple stressors in Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 367. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, Y. Unraveling salt stress signaling in plants. J. Integ. Plant Biol. 2018, 60, 796–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dvořák, P.; Krasylenko, Y.; Ovečka, M.; Basheer, J.; Zapletalová, V.; Šamaj, J.; Takáč, T. In vivo light-sheet microscopy resolves localisation patterns of FSD1, a superoxide dismutase with function in root development and osmoprotection. Plant Cell Environ. 2021, 44, 68–87. [Google Scholar] [CrossRef] [PubMed]
- Piacentini, D.; Rovere, F.D.; Bertoldi, I.; Massimi, L.; Sofo, A.; Altamura, M.M.; Falasca, G. Peroxisomal PEX7 receptor affects cadmium-induced ROS and auxin homeostasis in Arabidopsis root system. Antioxidants 2021, 10, 1494. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhang, X.; Lv, Q.; Zhu, D.; Qiu, T.; Xu, Y.; Bao, F.; He, Y.; Hu, Y. Physcomitrella patens dehydrins (PpDHNA and PpDHNC) confer salinity and drought tolerance to transgenic Arabidopsis plants. Front. Plant Sci. 2017, 8, 1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.X.; Zhang, H.; Su, H.X.; Xia, K.F.; Jian, S.G.; Zhang, M. Ipomoea pes-caprae IpASR improves salinity and drought tolerance in transgenic Escherichia coli and Arabidopsis. Int. J. Mol. Sci. 2018, 19, 2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name a | Gene Identifier b | Gene Symbol c | Gene d | Protein e | ||||
|---|---|---|---|---|---|---|---|---|
| Chr | Intron | CDS (bp) | AA | MW (kDa) | pI | |||
| SlBAG1 | Solyc01g095320 | LOC101246665 | 1 | 2 | 3708 | 1235 | 137.35 | 5.30 |
| SlBAG2 | Solyc03g026220 | LOC101264896 | 3 | 3 | 1026 | 341 | 38.25 | 9.52 |
| SlBAG3 | Solyc03g083970 | LOC101246514 | 3 | 2 | 1188 | 395 | 45.28 | 9.43 |
| SlBAG4 | Solyc04g014740 | LOC109119998 | 4 | 0 | 651 | 216 | 24.44 | 5.83 |
| SlBAG5 | Solyc06g007240 | LOC101243790 | 6 | 3 | 837 | 278 | 31.53 | 6.28 |
| SlBAG6 | Solyc06g035720 | LOC101267811 | 6 | 3 | 1002 | 333 | 37.56 | 9.45 |
| SlBAG7 | Solyc06g072430 | LOC104648101 | 6 | 0 | 1122 | 373 | 42.50 | 5.66 |
| SlBAG8 | Solyc08g080320 | LOC101246459 | 6 | 3 | 855 | 284 | 32.33 | 9.73 |
| SlBAG9 | Solyc10g084170 | LOC101250069 | 10 | 0 | 513 | 170 | 19.43 | 10.26 |
| SlBAG10 | Solyc10g085290 | LOC101258018 | 10 | 4 | 1185 | 394 | 41.91 | 5.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, H.; Ji, Y.; Sheng, J.; Wang, Y.; Liu, X.; Xiao, P.; Ding, H. Genome-Wide Identification of the Bcl-2 Associated Athanogene (BAG) Gene Family in Solanum lycopersicum and the Functional Role of SlBAG9 in Response to Osmotic Stress. Antioxidants 2022, 11, 598. https://doi.org/10.3390/antiox11030598
Jiang H, Ji Y, Sheng J, Wang Y, Liu X, Xiao P, Ding H. Genome-Wide Identification of the Bcl-2 Associated Athanogene (BAG) Gene Family in Solanum lycopersicum and the Functional Role of SlBAG9 in Response to Osmotic Stress. Antioxidants. 2022; 11(3):598. https://doi.org/10.3390/antiox11030598
Chicago/Turabian StyleJiang, Hailong, Yurong Ji, Jiarong Sheng, Yan Wang, Xiaoya Liu, Peixiang Xiao, and Haidong Ding. 2022. "Genome-Wide Identification of the Bcl-2 Associated Athanogene (BAG) Gene Family in Solanum lycopersicum and the Functional Role of SlBAG9 in Response to Osmotic Stress" Antioxidants 11, no. 3: 598. https://doi.org/10.3390/antiox11030598