Being climacteric, mango fruits from tropical countries undergoing long-distance transportation (e.g., to the EU markets) are usually harvested firm at the mature, green stage and complete ripening off-vine [
33]. Thus, large retail distribution typically offers a selection of mango fruits heterogeneous for ripening, with some at the ready-to-eat stage and others still unripe. Generally, at harvest, the total soluble solids of mango fruit have values between 7 and 9 °Brix, which increase during ripening up to 13–15 °Brix. Simultaneously, a reduction in pulp firmness and an intensification of the yellow color of the pulp occurs [
34]. In the present research, mango fruits uniform for ripening were carefully selected based on the instrumental determination of pulp firmness and total soluble solids, and on the visual comparison of the pulp color with cultivar specific reference chromatic scales (
www.mango.org; accessed on 15 April 2020). In particular, the Tommy Atkins fruits selected for this research had values of firmness and total soluble solids of 3.9 kg/cm
2 and 12.8 °Brix, respectively, while in the cultivar Keitt the corresponding average values were 3.3 kg/cm
2 and 11.5 °Brix.
3.1. Polyphenolic Composition
Polyphenols (including flavonoids, xanthones, and phenolic acids) are the most abundant dietary antioxidants of mango fruits. Well known for their ability to scavenge free radicals through hydrogen atom transfer, single electron transfer, and/or chelation of metal cations, polyphenols are thought to account for a large part of the biological and pharmacological activities attributed to mango fruits [
35]. Plant polyphenols occur in soluble or insoluble forms, both with important nutritional value as they have a different metabolic fate and biological significance in humans [
36]. Soluble phenolics generally segregate within the vacuole, free or conjugated to oligosaccharides and peptides through ester or ether bonds, and play a pivotal role in plant defense against ultraviolet radiation, aggression by pathogens, parasites, and predators, as well as in contributing to plant organ color. Instead, insoluble phenolics are covalently bound to the cell wall polymers and exert a mainly structural role [
37].
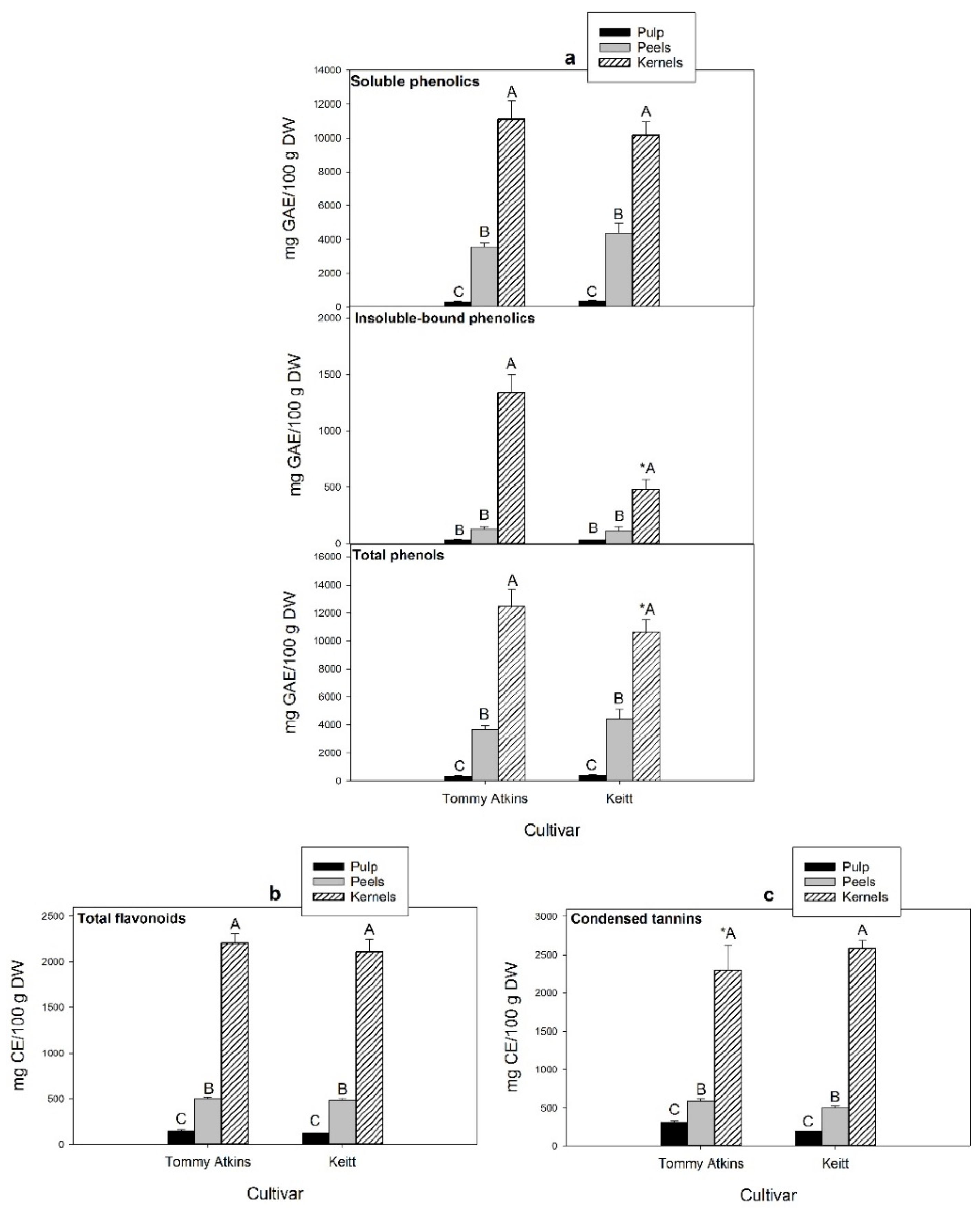
The average contents of phenolic compounds (soluble, insoluble bound, and total), flavonoids, and condensed tannins in the pulp, peels, and kernels obtained from ripe mango fruits of the cultivars Tommy Atkins and Keitt are reported in
Figure 3. Regardless of cultivar and fruit fractions, soluble phenolics largely exceeded the insoluble bound, representing up to 97% of the total. Significant differences were detected among fruit fractions (
Figure 3a), with kernels showing the highest contents of both soluble and insoluble-bound phenolics, followed by peels and pulp. No statistically significant variation was detected between the two cultivars, except for the concentration of kernel insoluble bound phenolics, much higher in Tommy Atkins (1340 mg GAE/100 g dw) than in Kent (480 mg GAE/100 g dw). The present findings are consistent with those of other authors, who reported total phenolic concentrations in the range of 19,749–20,034, 2032–9200, and 253–699 mg GAE/100 g dw, respectively, in the kernels, peels and pulp from ripe fruits of the same cultivars [
38,
39,
40,
41].
Interestingly, large differences among pulp, peels, and kernels of both cultivars were found for the conjugated phenolic content, which contributed to 95–97%, 83–94%, and 11–29% of the total soluble phenolics, respectively (
Table 1), possibly in relation to the metabolic state of the tissues. Free phenolics are usually associated to dead or dying tissues. Thus, the high content found in the kernels may relate to seed development, culminating in programmed cell death and hardening of tissues enclosing the embryo (e.g., seed coat). Besides, conjugation is of considerable biological significance. Indeed, the degree of glycosylation was reported to significantly affect the antioxidant activity of phenolics. Aglycones of quercetin and myricetin, for example, demonstrated higher antioxidant activity than their corresponding glycosides, though conjugated forms are better absorbed in humans [
42].
Flavonoids represent a significant portion of soluble phenolics in ripe mangoes. Thus, their total content followed a similar distribution trend in the different fruit fractions (
Figure 3b), being the highest in kernels (2110–2204 mg CE/100 g dw), followed by peels (480–504 mg CE/100 g dw) and pulp (119–147 mg CE/100 g dw), with no statistically significant differences between cultivars. A marked genotype-associated variability in the content of total phenolics and flavonoids (within the range 2930–6624 mg GAE/100 g dw and 502–795 mg CE/100 g dw, respectively) was instead reported by Marcillo-Parra et al. [
43] in the peels of three mango cultivars (Tommy Atkins, Kent, and Haden) from the Ecuadorian region. Besides, compared to our results, lower and higher flavonoid levels were reported for the kernels (1300 mg CE/100 g dw) and peels (700 mg CE/100 g dw) of Keitt mangoes by Dorta et al. [
44], indicating the existence of a certain intravarietal variability. This was further supported by Peng et al. [
41], who found only 186 mg CE/100 g dw in the peels of the same cultivar.
Condensed tannins (proanthocyanidins) are oligo/polymeric flavonoids consisting of flavan-3-ol structural units (i.e., catechin, epicatechin, and/or epigallocatechin) naturally present in many vegetables, seeds, and fruits. Mango kernels also ranked first for this class of compounds (2299–2580 CE mg/100 g dw), followed by peels (500–584 CE mg/100 g dw) and pulp (190–313 mg CE/100 g dw) with no significant differences among cultivars, except for kernels (
Figure 3c). High levels of condensed tannins (1323 mg CE/100 g dw) were previously reported by Makkar et al. [
45] in mango kernels, which have proven to be among the agro industrial byproducts with the greatest potential as sources for the large scale extraction of these compounds. Given their natural antioxidant and antimicrobial activity, condensed tannins are commercially used as a preservative to stabilize food colors, prevent rancidity, and avoid microbial growth [
46]. Levels of up to 7.0 and to 0.58 mg leucoanthocyanidin equivalents (LE)/100 g dw were reported, respectively, in mango kernels and peels of the cultivar Keitt by Hung et al. [
38], who highlighted a large variability associated with the used solvent and extraction temperature and the need for optimizing the extraction parameters to obtain extracts rich in antioxidants from mango byproducts.
With regard to the profiles of the major phenolics (free and conjugated forms), overall, seven individual compounds were identified in the tree fruit fractions (
Table 1).
Kernels of both cultivars were characterized by high levels of free methyl gallate (up to 2126.5 mg/100 g dw) and propyl gallate (about 1400 mg/100 g dw), also detected in the peels, though in considerably lower concentrations. Methyl gallate was the only free polyphenol identified in the pulp of both varieties, where it was present also in the conjugated form together with large amounts of gallic acid. Free gallic acid was detected only in the kernels with a significant difference in concentration between Tommy Atkins (14.6 mg/100 g dw) and Keitt (104.2 mg/100 g dw) cultivars. On the contrary, Kim et al. [
47] reported free gallic acid as the major phenolic acid in mango pulp (cultivar Tommy Atkins).
Gallic acid and its methyl- and propyl-derivatives are known to exert strong antioxidative and antiviral effects in vitro and in vivo [
48,
49]. Besides, gallic acid and methyl gallate extracted from seeds of white catamaran tree (
Givotia rottleriformis Griff. ex Wight) reduced the growth of human epidermoid carcinoma cells and exhibited inhibitory activity against hepatitis C virus [
50,
51,
52].
Though identified in a number of other plants, mangiferin is a polyphenol of mango distinctive for its high levels characterizing the fruits from which it is mainly produced [
53]. Mangiferin has been reported to possess a broad range of therapeutic effects, including anti-inflammation, anti-diabetic, immunomodulatory, anti-tumor, and antioxidant activities [
54,
55]. It has been used in many food supplements and is considered one of the main active constituents in more than 40 polyherbal formulations in traditional Chinese medicine [
53]. Mangiferin was differentially distributed in the fruit of mango. It was the predominant soluble conjugated phenolic compound in Tommy Atkins peels (411 mg/100 g dw) but was not observed in Keitt peel samples. Mangiferin was also present, at lower and significantly different levels, in the kernels of the two cultivars. In agreement with our findings, Luo et al. [
56] found the highest mangiferin level in the peels (4–749 mg/100 g dw), followed by kernels (14–243 mg/100 g dw) and pulp (0–20 mg/100 g dw) with a large intervarietal variability among the eleven Chinese mango cultivars assayed. Differently, López-Cobo et al. [
57] found higher levels of mangiferin in the kernels (22–73 mg/100 g dw) than in peels (4–30 mg/100 g dw) and husk (2–17 mg/100 g dw) of three mango cultivars (Keitt, Osteen and Sensación) from Spain, while it was not detected in the pulp. A genotype-dependent variation of mangiferin levels was also reported by Berardini et al. [
58] who found a high amount of mangiferin in Tommy Atkins peels (126.3 mg/100 g dw), but much lower levels in samples from Haden (1.1 mg/100 g dw) and Kent (1.4 mg/100 g dw) cultivars. Rutin was also detected in the peels (30.2–65.2 mg/100 g dw) and kernels (25.5–40.3 mg/100 g dw), while luteolin-7-
O-glucoside was exclusive to the peels (5.3–13.1 mg/100 g dw) of both cultivars.
3.2. Ascorbic (AsA) and Dehydroascorbic (DHA) Acid Contents
Ascorbic acid (AsA) in plants plays a central role in several physiological processes, including cell redox potential buffering, regulation of photosynthesis and production of phytohormones, control of cell division and growth, and signal transduction. It is also a chief component of the efficient antioxidant machinery evolved by plants to counteract harmful reactive species [
59]. With regard to human nutrition, AsA represents the dominant biologically active form of vitamin C present in most edible plants including mango, although the pulp of the ripe fruits is not considered a major source of this essential nutrient. Besides, its reversibly oxidized form, dehydroascorbic acid (DHA), has also been detected in conventional and organic mango fruits at much lower levels and reported to contribute up to 18% of the total Vitamin C, though large variations were observed depending on cultivar, fruit tissue, post-harvest manipulations/treatments, and storage [
60,
61]. An inverse correlation was found between mango peel browning resulting from chilling injuries and ascorbic acid concentrations [
62]. AsA levels between 2.8- and 19.0-fold those of DHA, depending on fraction and cultivar, were observed in the present study (
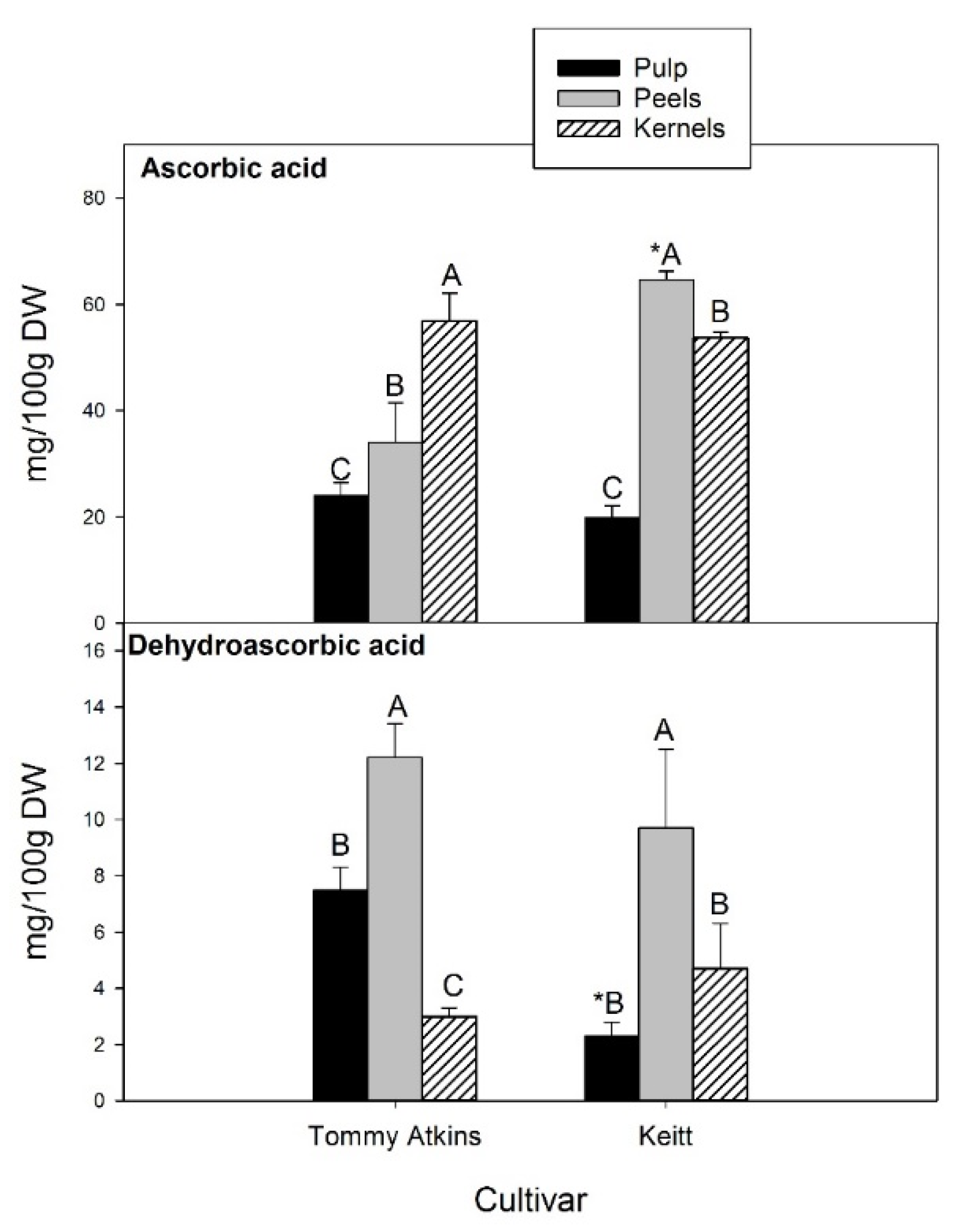
Figure 4).
In kernels, AsA and DHA contents were about 55.0 and 3.8 mg/100 g dw, respectively, with no significant differences between Tommy Atkins and Keitt, while showing a different ranking in the different fruit fractions. In Tommy Atkins, kernels ranked first for AsA content, followed by peels (34.0 mg/100 g dw) and pulp (24.0 mg/100 g dw). In the cultivar Keitt, the highest concentration was registered in the peels (64.6 mg/100 g dw), which also turned out to be the fruit fraction with the highest DHA content (on average 11.0 mg/100 g dw). There were no statistically significant differences between cultivars. With regard to pulp, Tommy Atkins showed a DHA level 3.3-fold higher than Keitt. Our values are consistent with those reported by Sogi et al. [
40] and Carvalho et al. [
63], who found ascorbic acid contents from 68.5 to 84.74 mg/100 g in dried peel powder and 61.2–74.5 mg/100 g dw in the kernels of market-ripe mangoes (cv Tommy Atkins).
3.3. Lipophilic Bioactives
A qualitative–quantitative determination of the main lipophilic phytochemicals of mango fruits (i.e., phytosterols, pentacyclic triterpenes, tocopherols, carotenoids, and chlorophylls) is reported in
Table 2.
Phytosterols are the predominant lipophilic class of bioactives in all mango fruit tissues. They are well known for their cholesterol-lowering effects, anti-inflammatory and antioxidant properties, and the benefits they offer to the immune system. Previous analyses of three mango cultivars from Spain indicated peels as the fruit fraction richest in phytosterols with a total amount in the range 56.6–69.3 mg/100 g dw, followed by kernels (31.8–54.4 mg/100 g dw) and pulp (24.5–40.1 mg/100 g dw) [
64]. However, our findings support a different ranking with pulp first (up to 151.5 mg/100 g dw), followed by kernels (106.9–135.5 mg/100 g dw) and peels (55.9–83.0 mg/100 g dw). Values still lower but much closer to those we found in the pulp were reported by Vilela et al. [
65] comparing 12 cultivars from Madeira Island, though a large variability in the level of total phytosterols within a range of 34.3–103.0 mg/100 g dw was reported among genotypes. β-sitosterol was the only sterol identified in the kernels of both cultivars and contributed to over 80% and 70–80% of the total sterols in the pulp and kernels, respectively. Stigmasterol and campesterol were also detected, though with different relative percentages, in agreement with the reports of other authors [
64,
65,
66].
Pentacyclic triterpenes exhibit a large range of biological activities, including anti-inflammatory, anti-cancer, and gastroprotective properties [
67,
68]. In this work, two pentacyclic triterpenes, α-amyrin and lupeol, were identified in the pulp and peel fractions of both cultivars, with peels showing the highest total concentration (17.7 and 14.2 mg/100 g dw in Tommy Atkins and Keitt, respectively) (
Table 2). In all fruit fractions, α-amyrin and lupeol were almost equivalent in concentration, with the exception of the Tommy Atkins pulp, in which the α-amyrin level was about twice as much. Recently, Mannowetz et al. [
69] reported that lupeol may find application as a non-hormonal contraceptive, inhibiting sperm hyperactivation critical to egg cell penetration of the zona pellucida. In accordance with the findings of Ruiz-Montañez et al. [
70] and Jyotshna et al. [
71], lupeol concentration was 1.7–4.0 times higher in peels than in pulp, indicating a potential use of this massive by-product of mango processing as a source for the extraction of this pharmacologically active triterpenoid.
Tocopherols are known as potent antioxidants with vitamin E activity and essential micronutrients in the human diet. In mango fruit, their accumulation is highly correlated with
p-hydroxyphenyl pyruvate dioxygenase gene expression during ripening, which is in turn ethylene induced and differentially expressed depending on cultivars [
72]. The highest tocopherol total content was registered in the peels of both cultivars, though with statistically significant differences between Tommy Atkins and Keitt (10.6 and 12.3 mg/100 g dw, respectively), while in pulp it was 3.3–4.6-fold lower. Mango kernels showed a very low amount of total tocopherols (0.5–0.2 mg/100 g dw) with no significant differences between cultivars. The α-isoform prevailed over the β in kernels (70% of the total tocopherols), and it was the only isoform identified in the pulp, in agreement with the data reported by López-Cobo [
64]. In the kernels, however, the two isoforms were in almost equivalent concentrations. Differently, Jin et al. [
66] reported that mango kernel fat was mostly dominated by α-tocopherol (>40%) and presented high percentages (15–45%) of the δ-isoform, albeit with large differences in total tocopherol contents and isoform profiles depending on cultivar.
Several pigments, including carotenoids and chlorophylls, are responsible for the cultivar-specific mango fruit pulp and peel color changes occurring during ripening. Most of them also show biological activities, acting as antioxidants or provitamins. Our results revealed differences in the carotenoid profile among the assayed mango fruit fractions. Peel samples had the highest total carotenoid content (1.75 mg/100 g dw in Tommy Atkins and 3.76 mg/100 g dw in Keitt), followed by pulp (0.71 and 1.22 mg/100 g dw, respectively) (
Table 2). The cultivar Keitt exhibited significantly higher total carotenoid contents than Tommy Atkins, while kernels recorded similar contents. Seven carotenoids (four carotenes and three xanthophylls) were comprehensively identified. Violaxanthin was characteristic of the peel of the cultivar Keitt, lutein was not detected in the pulp of Tommy Atkins, and α-carotene characterized the peel of both cultivars. β-Carotene was detected in all fruit fractions and contributed about 80% and 86% to the total carotenoids of Tommy Atkins and Keitt pulp, respectively. β-carotene and violaxanthin were previously reported to prevail over the other carotenoids in Keitt and Tommy Atkins mangoes, accounting for 28–38% and 27–33% of the total, respectively, according to Mercandante et al. [
73,
74] and Ruales et al. [
46]. However, the levels of β-carotene found by Mercandante et al. [
73,
74] in the pulp of Tommy Atkins and Keitt cultivars (0.58 and 1.5 mg/100 g dw, respectively) were similar to those reported in this study. Our findings are in agreement with the results obtained by Fratianni et al. [
75], who reported the presence of 9-cis and 13-cis-isomers of β-carotene in the pulp of Keitt mangoes, although the total amount (about 12 mg/100 g dw) was much higher. Total carotenoids were 2.5–3-fold more concentrated in peels than in pulp. In the peels of Tommy Atkins, β-carotene contributed about 46% of the total carotenoids, followed by lutein (39%). Similar results were reported by Marcillo-Parra [
43] for peels isolated from Tommy Atkins, Haden, and Kent cultivars. On the contrary, Keitt had 68% lutein and 23% β-carotene with respect to the total identified carotenoids. Some authors found lutein as the main carotenoid of peels, followed by β-carotene and low amounts of other carotenoids [
46,
76]. The high concentration of carotenoids in the peel is probably related to the high exposure to sunlight that induces an increase in carotengenesis [
77]. Albeit in low quantity, kernel contained exclusively lutein and β-carotene.
Chlorophylls (a and b) were found in the peels of Tommy Atkins and Keitt cultivars (
Table 2) in ratios of 1:3 and 1:4, respectively. Similar results were obtained by Dorta et al. [
41] in freeze-dried peels of the cultivar Keitt.
3.4. Antioxidant Properties
Table 3 reports the HAA, LAA, and TAA of the pulp, peels and kernels obtained from the ripe fruits of both mango cultivars. HAA largely exceeded LAA in all fruit fractions, with both varying significantly among cultivars. Kernels showed the highest HAA (approximately 120 mol TE/100 g dw in both cultivars) and LAA (28.6 and 19.7 mol TE/100 g dw in Tommy Atkins and Keitt, respectively). After kernels, peels exhibited greater HAA and LAA than pulp. In particular, HAA and LAA of Keitt peel were greater than those evaluated in the peel of Tommy Atkins. Finally, the antioxidant activities determined in the pulp of both cultivars did not show statistically significant differences.
Several authors have previously investigated and demonstrated the positive correlation between TPC and antioxidant activity [
78,
79]. Our results (
Table 4) indicated that TPC has a statistically significant (
p < 0.001) positive Pearson correlation with HAA (
r = 0.990). The strong correlation suggests that phenolic compounds present in the different mango fruit fractions likely contribute to radical scavenging activity, as also reported by Ma et al. [
80]. Similarly, Puravankara et al. [
81] found phenolic compounds as the main contributors to the HAA of mango seed kernel extracts, which, in line with the high phenolic content, exerted the highest antioxidant activity among several fruit seeds, including jackfruit, longan, avocado, and tamarind [
82]. A strong correlation was also observed between HAA and TFC (
r = 0.994). In previous studies, Rumainum et al. [
78] reported a strong correlation between TFC and total antioxidant activity in Thai mangoes. Furthermore, HAA and LAA were positively correlated with TCT (
r = 0.984,
r = 0.982, respectively). Overall, our findings suggest that total phenolics, flavonoids, and condensed tannins are the major compounds contributing to the antioxidant activity in our samples. None of the lipophilic molecules correlates significantly with LAA, indicating an idiosyncratic rather than synergistic interaction of the individual contributors.
3.5. Principal Component Analysis
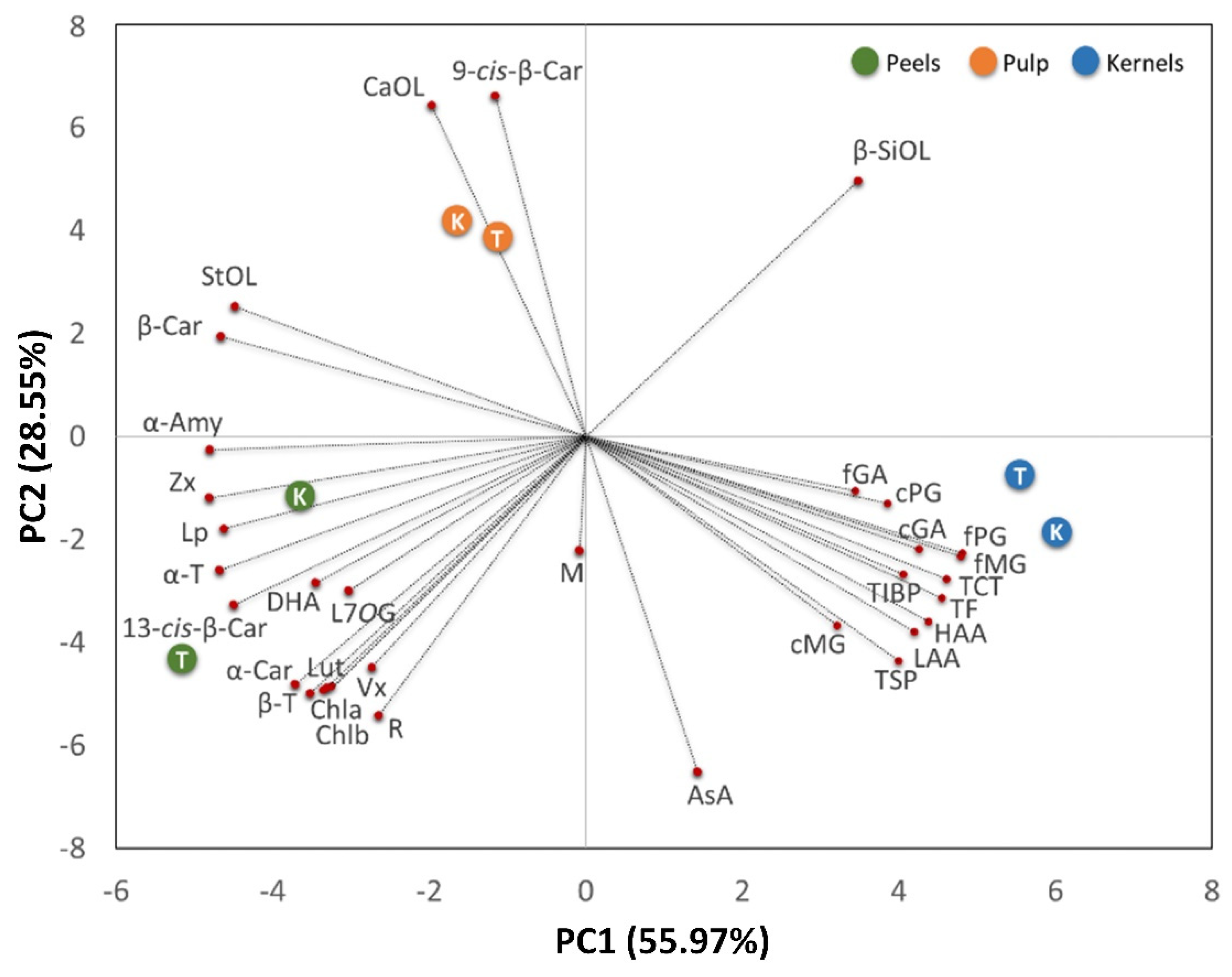
To highlight the biochemical differences among fruit fractions and cultivars and any clustering of the observations, a multivariate analysis (PCA) was carried out (
Figure 5).
Two relevant principal components (PCs) that explained 84.52% of the total variance of the collected data were extracted, with principal component 1 (PC1) and principal component 2 (PC2) accounting for 55.97% and 28.55%, respectively. The contribution of each quality parameter (variables) is reported in
Table 5. The PC1 vs. PC2 plot shows a clear clustering of fruit fractions.
Independently of cultivars, pulp fractions grouped on the upper-left side of the chart show a high concentration of phytosterols (mainly campesterol and β-sitosterol) and β- and 9-cis-β-carotene-isoforms. Phytosterols are lipophilic membrane components that are not only essential for diverse cellular functions, but are also biosynthetic precursors of the brassinosteroids [
83]. Thus, their abundance in the mango pulp may be related to the extensive membrane system of the metabolically active parenchymal cells of the mesocarp tissue and/or to the physiological role of brassinosteroid phytohormones in the regulation of fleshy fruit ripening.
Peels grouped on the lower-left side panel, characterized by the presence of chlorophylls, responsible for the green colour, carotenes (α- and 13-cis-β-isoforms), and xanthophylls, are likely involved in photosynthesis as accessory pigments. Peels also correlated with lupeol, α-amyrin, and α- and β-tocopherols, presumably located in the cuticular waxes coating the surface of fleshy fruits with the role of limiting water loss, providing mechanical support, preventing fruit softening, and acting as a barrier to pathogens [
84,
85], as well as with dehydroascorbic acid, luteolin-7-
O-glucoside, and rutin, which reflect the protective role that peel flavonoid compounds exert against the adverse biotic (e.g., pathogens, insects and herbivores attack) and abiotic (e.g., UV radiation and temperature) factors [
86].
Kernels were distributed on the bottom right panel showing a high content phenolic (both soluble and insoluble), total flavonoid, total condensed tannins, as well as high hydrophilic and lipophilic antioxidant activities. As reported by Corso et al. [
87] phenolics favor seed survival and dispersion and provide chemical defense against pathogens, seed defense against biotic attacks, as well as predators, such as insect pests and herbivores.
3.6. Fatty Acids Composition
The fatty acid profiles of the pulp, peels, and kernels obtained from the ripe fruits of Tommy Atkins and Keitt mango cultivars are reported in
Table 6. Myristic, palmitic, stearic, arachidic, oleic, and linolenic acids were detected in all samples, though with significant quantitative differences. Palmitic was the most abundant fatty acid in the pulp (contributing to 18.5% and 20.1% of the total identified fatty acids in Tommy Atkins and Keitt, respectively), followed by the polyunsaturated fatty acids (PUFA) linolenic (15.9–19.8%) and linoleic (14.2–17.2%). The latter two, together with palmitic acid, prevailed in the peels of both cultivars, constituting up to 71.9% of the total. Myristic, heptadecanoic, stearic, arachidic, behenic, lignoceric, palmitoleic, and oleic acids were also detected as minor components in peels. Kernels were a rich source of oleic (38.4–41.6%), stearic (32.8–36.3%) and palmitic acids (13.8–11.1%). Our results fall within the ranges reported for the pulp, peels, and kernels of ripe mango fruits by other authors [
65,
88,
89,
90]. It is worth mentioning that some PUFA, including linoleic and linolenic acids, are essential nutrients for humans and must be introduced with the diet [
89]. The role of n-3 and n-6 fatty acids for health is related with the prevention, delay, or treatment of chronic and acute diseases, including cancer, cardiovascular diseases, osteoporosis, and immune disorders [
91,
92,
93,
94]. Our analysis revealed an n6/n3 ratio <1, indicating mango fruits as a perfect source of essential fatty acids [
88].









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}