
Leaves of Cedrela sinensis Attenuate Chronic Unpredictable Mild Stress-Induced Depression-like Behavior via Regulation of Hormonal and Inflammatory Imbalance
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sample Preparation
2.3. Physiological Compound Analysis
2.4. Evaluation of Neuronal Protective Effect
2.4.1. Cell Culture and Treatment
2.4.2. Cell Viability
2.5. Chronic Unpredictable Mild Stress (CUMS) Procedures
2.6. Animal Design
2.7. Behavioral Tests
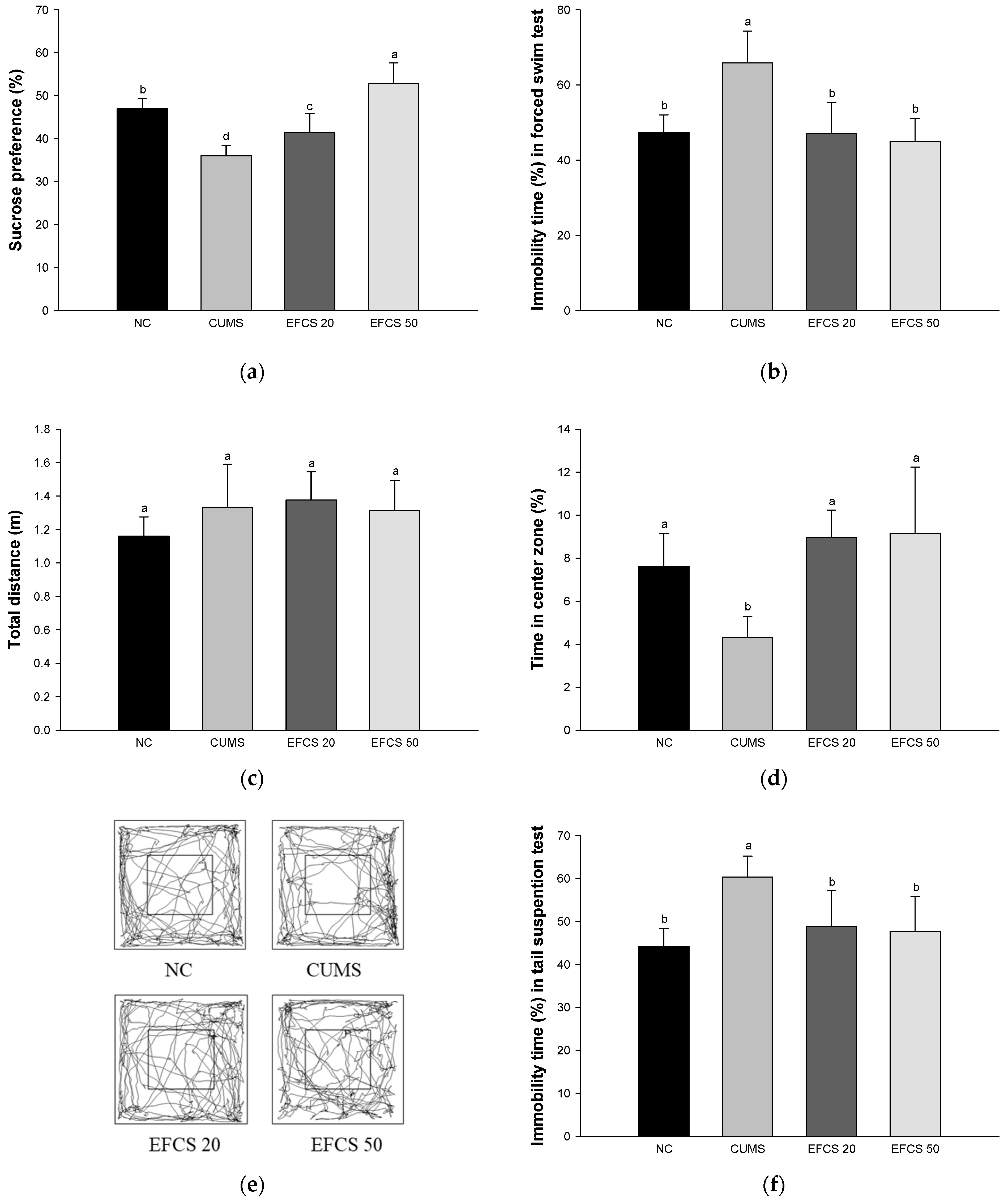
2.7.1. Sucrose Preference Test (SPT)
2.7.2. Forced Swimming Test (FST)
2.7.3. Tail Suspension Test (TST)
2.7.4. Open Field Test (OFT)
2.8. Preparation of Brain Tissues
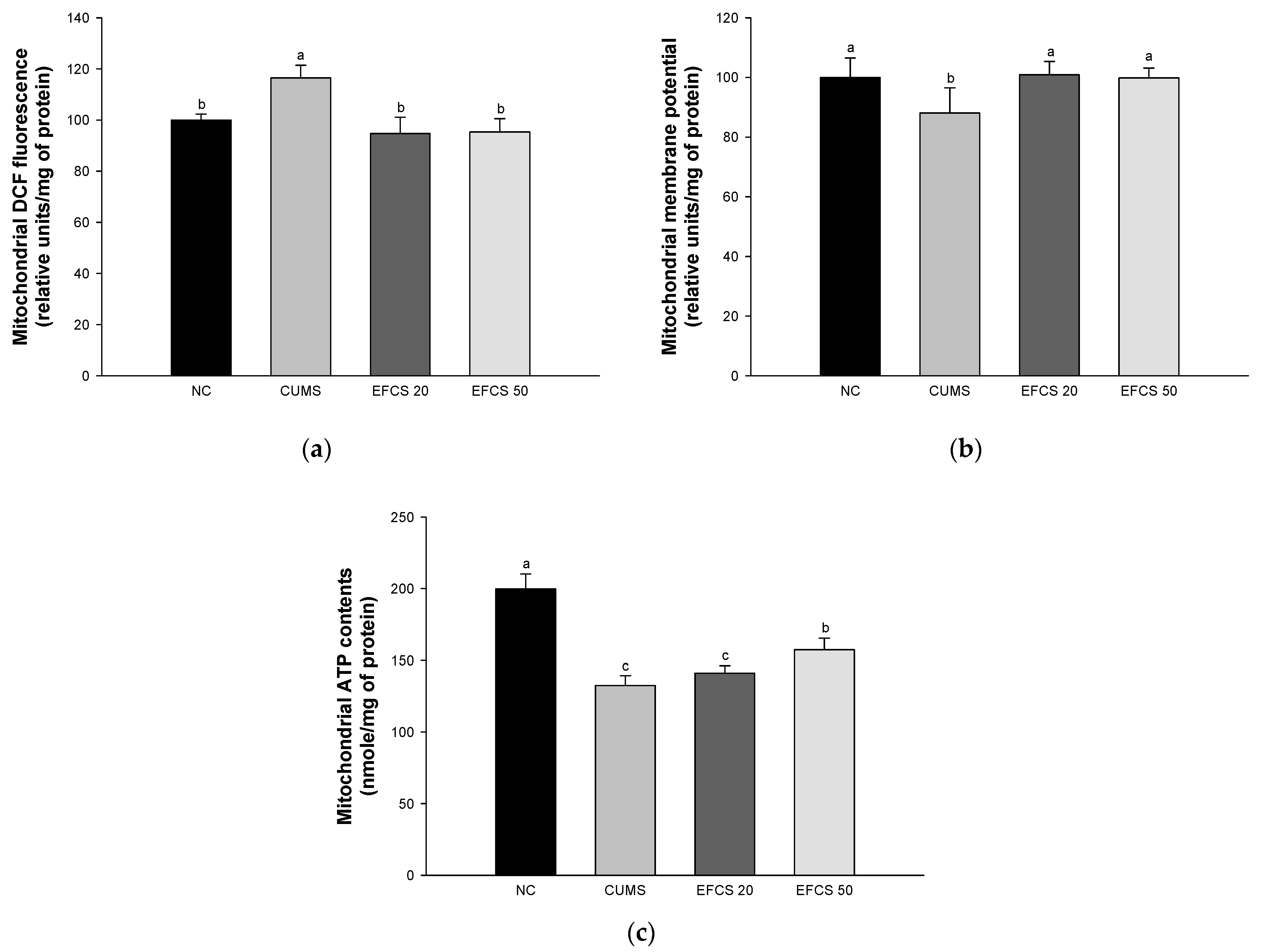
2.9. Mitochondrial Activity
2.9.1. Mitochondrial Isolation
2.9.2. Measurement of Mitochondrial Reactive Oxygen Species (ROS) Contents
2.9.3. Measurement of the Mitochondrial Membrane Potential (MMP)
2.9.4. Measurement of Mitochondrial ATP Contents
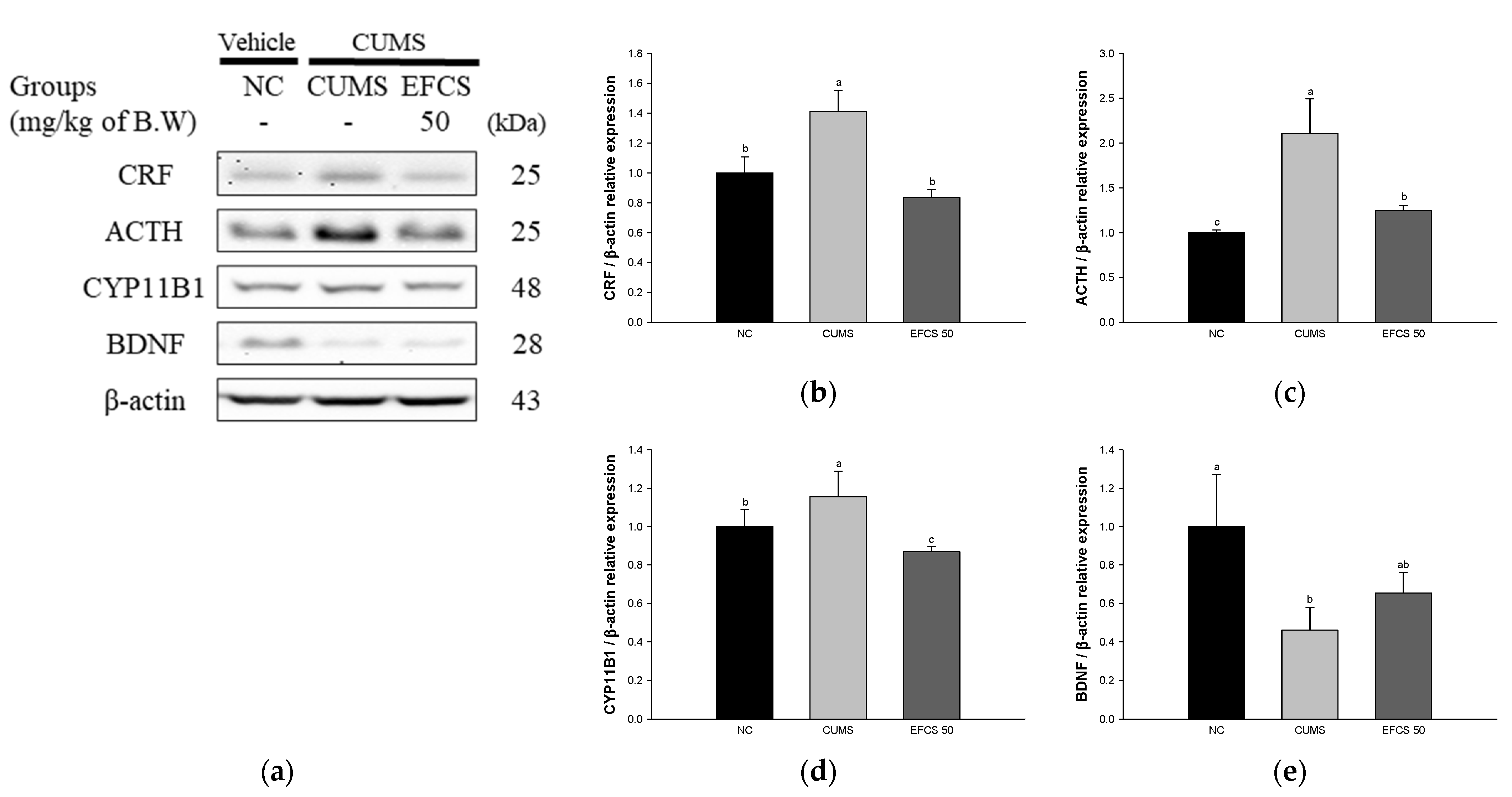
2.10. Hormonal Analysis
2.11. Western Blot Analysis
2.12. Statistical Analysis
3. Results
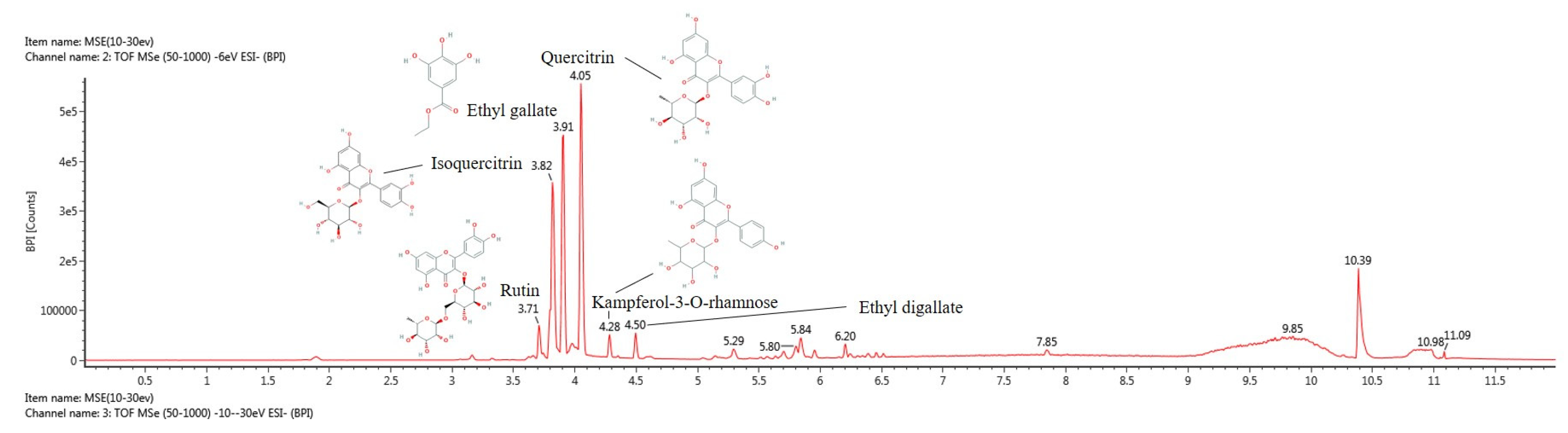
3.1. Physiological Compound Using UPLC Q-TOF/MSE
3.2. Neuroprotective Effect
3.3. Behavioral Test
3.4. Mitochondrial Activity
3.5. Hypothalamic Hormone Change
3.6. Stress-Related Pathway
3.7. Protein Expression of Neuroinflammation
3.8. Protein Expression of Apoptosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bartolomucci, A.; Palanza, P.; Sacerdote, P.; Panerai, A.E.; Sgoifo, A.; Dantzer, R.; Parmigiani, S. Social factors and individual vulnerability to chronic stress exposure. Neurosci. Biobehav. Rev. 2005, 29, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Koolhaas, J.M.; Meerlo, P.; de Boer, S.F.; Strubbe, J.H.; Bohus, B. The temporal dynamics of the stress response. Neurosci. Biobehav. Rev. 1997, 21, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J.; Barrot, M.; DiLeone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M. Neurobiology of depression. Neuron 2002, 34, 13–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathers, C.D.; Loncar, D. Projections of global mortality and burden of disease from 2002 to 2030. PLoS Med. 2006, 3, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauger, R.L.; Risbrough, V.; Oakley, R.H.; Olivares-Reyes, J.A.; Dautzenberg, F.M. Role of CRF receptor signaling in stress vulnerability, anxiety, and depression. Ann. N. Y. Acad. Sci. 2009, 1179, 120–143. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Xie, K.; Yang, X.; Gu, J.; Ge, L.; Wang, X.; Wang, Z. Resveratrol reverses the effects of chronic unpredictable mild stress on behavior, serum corticosterone levels and BDNF expression in rats. Behav. Brain Res. 2014, 264, 9–16. [Google Scholar] [CrossRef]
- Dallman, M.F. Stress update: Adaptation of the hypothalamic-pituitary-adrenal axis to chronic stress. Trends Endocrinol. Metab. 1993, 4, 62–69. [Google Scholar] [CrossRef]
- Marin, M.F.; Lord, C.; Andrews, J.; Juster, R.P.; Sindi, S.; Arsenault-Lapierre, G.; Fiocco, A.J.; Lupien, S.J. Chronic stress, cognitive functioning and mental health. Neurobiol. Learn. Mem. 2011, 96, 583–595. [Google Scholar] [CrossRef]
- Pape, K.; Tamouza, R.; Leboyer, M.; Zipp, F. Immunoneuropsychiatry—Novel perspectives on brain disorders. Nat. Rev. Neurol. 2019, 15, 317–328. [Google Scholar] [CrossRef]
- Eyre, H.; Baune, B.T. Neuroplastic changes in depression: A role for the immune system. Psychoneuroendocrinology 2012, 37, 1397–1416. [Google Scholar] [CrossRef]
- Golovatscka, V.; Ennes, H.; Mayer, E.A.; Bradesi, S. Chronic stress-induced changes in pro-inflammatory cytokines and spinal glia markers in the rat: A time course study. Neuroimmunomodulation 2012, 19, 367–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farhy-Tselnicker, I.; Allen, N.J. Astrocytes, neurons, synapses: A tripartite view on cortical circuit development. Neural Dev. 2018, 13, 7. [Google Scholar] [PubMed] [Green Version]
- Pollak, Y.; Yirmiya, R. Cytokine-induced changes in mood and behaviour: Implications for ‘depression due to a general medical condition’, immunotherapy and antidepressive treatment. Int. J. Neuropsychopharmacol. 2002, 5, 389–399. [Google Scholar] [CrossRef]
- Connor, T.J.; Leonard, B.E. Depression, stress and immunological activation: The role of cytokines in depressive disorders. Life Sci. 1998, 62, 583–606. [Google Scholar] [CrossRef]
- Kobrosly, R.; van Wijngaarden, E. Associations between immunologic, inflammatory, and oxidative stress markers with severity of depressive symptoms: An analysis of the 2005–2006 National Health and Nutrition Examination Survey. Neurotoxicology 2010, 31, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Liu, Y.; Hu, M.; Zhang, M.; Yang, J.; Liang, F.; Huang, Q.; Wu, C. Toona sinensis: A comprehensive review on its traditional usages, phytochemisty, pharmacology and toxicology. Rev. Bras. Farm.-Braz. J. Pharmacogn. 2019, 29, 111–124. [Google Scholar] [CrossRef]
- Cheng, K.W.; Yang, R.Y.; Tsou, S.C.; Lo, C.S.; Ho, C.T.; Lee, T.C.; Wang, M. Analysis of antioxidant activity and antioxidant constituents of Chinese toon. J. Funct. Foods 2009, 1, 253–259. [Google Scholar] [CrossRef]
- Wang, K.J.; Yang, C.R.; Zhang, Y.J. Phenolic antioxidants from Chinese toon (fresh young leaves and shoots of Toona sinensis). Food Chem. 2007, 101, 365–371. [Google Scholar] [CrossRef]
- Kim, S.Y.; Lee, M.H.; Jo, N.R.; Park, S.N. Antibaterial activity and skin moisturizing effect of Cedrela sinensis A. Juss shoots extracts. J. Soc. Cosmet. Sci. Korea 2010, 36, 315–321. [Google Scholar]
- Shin, H.J.; Jeon, Y.J.; Shin, H.J. Physiological activities of extracts of Cedrela sinensis leaves. KSBB J. 2008, 23, 164–168. [Google Scholar]
- Park, J.C.; Yu, Y.B.; Lee, J.H.; Kim, N.J. Studies on the chemical components and biological activities of edible plants in Korea-(6)-anti-inflammatory and analgesic effects of Cedrela sinensis, Oenanthe javanica and Artemisia princeps var. orientalis. Korean J. Food Nutr. 1994, 23, 116–119. [Google Scholar]
- Cao, J.J.; Lv, Q.Q.; Zhang, B.; Chen, H.Q. Structural characterization and hepatoprotective activities of polysaccharides from the leaves of Toona sinensis (A. Juss) Roem. Carbohydr. Polym. 2019, 212, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Jia, K.K.; Zheng, Y.J.; Zhang, Y.X.; Liu, J.H.; Jiao, R.Q.; Pan, Y.; Kong, L.D. Banxia-houpu decoction restores glucose intolerance in CUMS rats through improvement of insulin signaling and suppression of NLRP3 inflammasome activation in liver and brain. J. Ethnopharmacol. 2017, 209, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.F.; Shi, T.Y.; Wang, W.N.; Chen, Y.C.; Tan, Q.R. Long-lasting effects of chronic rTMS to treat chronic rodent model of depression. Behav. Brain Res. 2012, 232, 245–251. [Google Scholar] [CrossRef]
- Shen, J.; Xu, L.; Qu, C.; Sun, H.; Zhang, J. Resveratrol prevents cognitive deficits induced by chronic unpredictable mild stress: Sirt1/miR-134 signalling pathway regulates CREB/BDNF expression in hippocampus in vivo and in vitro. Behav. Brain Res. 2018, 349, 1–7. [Google Scholar] [CrossRef]
- Song, L.; Che, W.; Min-Wei, W.; Murakami, Y.; Matsumoto, K. Impairment of the spatial learning and memory induced by learned helplessness and chronic mild stress. Pharmacol. Biochem. Behav. 2006, 83, 186–193. [Google Scholar] [CrossRef]
- Heo, H.J.; Cho, H.Y.; Hong, B.; Kim, H.K.; Kim, E.K.; Kim, B.G.; Shin, D.H. Protective effect of 4′,5-dihydroxy-3′,6,7-trimethoxyflavone from Artemisia asiatica against Aβ-induced oxidative stress in PC12 cells. Amyloid 2001, 8, 194–201. [Google Scholar] [CrossRef]
- Harro, J.; Häidkind, R.; Harro, M.; Modiri, A.R.; Gillberg, P.G.; Pähkla, R.; Matto, V.; Oreland, L. Chronic mild unpredictable stress after noradrenergic denervation: Attenuation of behavioural and biochemical effects of DSP-4 treatment. Eur. Neuropsychopharmacol. 1999, 10, 5–16. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, X.; Luo, S.; Chu, C.; Wu, D.; Liu, R.; Wang, L.; Wang, J.; Liu, X. Extract of sesame cake and sesamol alleviate chronic unpredictable mild stress-induced depressive-like behaviors and memory deficits. J. Funct. Food 2018, 42, 237–247. [Google Scholar] [CrossRef]
- Can, A.; Dao, D.T.; Arad, M.; Terrillion, C.E.; Piantadosi, S.C.; Gould, T.D. The mouse forced swim test. J. Vis. Exp. 2012, 59, 3638. [Google Scholar] [CrossRef] [Green Version]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P. The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology 1985, 85, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Kilgour, R. The open-field test as an assessment of the temperament of dairy cows. Anim. Behav. 1975, 23, 615–624. [Google Scholar] [CrossRef]
- Kim, J.M.; Kang, J.Y.; Park, S.K.; Moon, J.H.; Kim, M.J.; Lee, H.L.; Jeong, H.R.; Kim, J.C.; Heo, H.J. Powdered green tea (matcha) attenuates the cognitive dysfunction via the regulation of systemic inflammation in chronic PM2.5-exposed BALB/c mice. Antioxidants 2021, 10, 1932. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Yoon, Y.S.; Cho, J.W. Quantitative analysis of flavonoids from Salicomia herbacea L. extracst by LC-MS. Korean J. Med. Crop Sci. 2008, 16, 231–237. [Google Scholar]
- Zhou, H.; Xing, J.; Liu, S.; Song, F.; Cai, Z.; Pi, Z.; Liu, Z.; Liu, S. Screening and determination for potential α-glucosidase inhibitors from leaves of Acanthopanax senticosus harms by using UF-LC/MS and ESI-MSn. Phytochem. Anal. 2012, 23, 315–323. [Google Scholar] [CrossRef]
- Hollecker, L.; Pinna, M.; Filippino, G.; Scrugli, S.; Pinna, B.; Argiolas, F.; Murru, M. Simultaneous determination of polyphenolic compounds in red and white grapes grown in Sardinia by high performance liquid chromatography–electron spray ionisation-mass spectrometry. J. Chromatogr. A 2009, 1216, 3402–3408. [Google Scholar] [CrossRef]
- Sun, J.; Liang, F.; Bin, Y.; Li, P.; Duan, C. Screening non-colored phenolics in red wines using liquid chromatography/ultraviolet and mass spectrometry/mass spectrometry libraries. Molecules 2007, 12, 679–693. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Rabaneda, F.; Jauregui, O.; Lamuela-Raventos, R.M.; Bastida, J.; Viladomat, F.; Codina, C. Identification of phenolic compounds in artichoke waste by high-performance liquid chromatography–tandem mass spectrometry. J. Chromatogr. A 2003, 1008, 57–72. [Google Scholar] [CrossRef]
- Tian, F.; Li, B.; Ji, B.; Zhang, G.; Luo, Y. Identification and structure–activity relationship of gallotannins separated from Galla chinensis. LWT-Food Sci. Technol. 2009, 42, 1289–1295. [Google Scholar] [CrossRef]
- Lucca, G.; Comim, C.M.; Valvassori, S.S.; Réus, G.Z.; Vuolo, F.; Petronilho, F.; Gavioli, E.C.; Dal-Pizzol, F.; Quevedo, J. Increased oxidative stress in submitochondrial particles into the brain of rats submitted to the chronic mild stress paradigm. J. Psychiatr. Res. 2009, 43, 864–869. [Google Scholar] [CrossRef] [PubMed]
- Young, E.A.; Abelson, J.; Lightman, S.L. Cortisol pulsatility and its role in stress regulation and health. Front. Neuroendocr. 2004, 25, 69–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, T.; Kimoto, T.; Tanabe, N.; Hattori, T.A.; Yasumatsu, N.; Kawato, S. Corticosterone acutely prolonged N-methyl-d-aspartate receptor-mediated Ca2+ elevation in cultured rat hippocampal neurons. J. Neurochem. 2022, 83, 1441–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magariños, A.M.; McEwen, B.S.; Flügge, G.; Fuchs, E. Chronic psychosocial stress causes apical dendritic atrophy of hippocampal CA3 pyramidal neurons in subordinate tree shrews. J. Neurosci. 1996, 16, 3534–3540. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.H.; Xie, Y.T.; Guo, J.M.; Wang, X.P.; Jiang, B.; Zhang, W.; Qiang, L.; Kong, L.Y.; Liu, Y.P. Limonoids from the fresh young leaves and buds of Toona sinensis and their potential neuroprotective effects. J. Agric. Food Chem. 2020, 68, 12326–12335. [Google Scholar] [CrossRef]
- Lee, S.B.; Kim, J.Y.; Cho, S.O.; Ban, J.Y.; Ju, H.S.; Bae, K.H.; Seong, Y.H. Extract of Cedrela sinensis leaves protects neuronal cell damage induced by hydrogen peroxide in cultured rat neurons. Korean J. Med. Crop Sci. 2007, 15, 444–450. [Google Scholar]
- Chen, L.; Wu, X.; Shen, T.; Wang, X.; Wang, S.; Wang, J.; Ren, D. Protective effects of ethyl gallate on H2O2-induced mitochondrial dysfunction in PC12 cells. Metab. Brain Dis. 2019, 34, 545–555. [Google Scholar] [CrossRef]
- Guan, L.P.; Liu, B.Y. Antidepressant-like effects and mechanisms of flavonoids and related analogues. Eur. J. Med. Chem. 2016, 121, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Ossola, B.; Kääriäinen, T.M.; Männistö, P.T. The multiple faces of quercetin in neuroprotection. Expert Opin. Drug Saf. 2009, 8, 397–409. [Google Scholar] [CrossRef]
- Azuma, K.; Ippoushi, K.; Ito, H.; Horie, H.; Terao, J. Enhancing effect of lipids and emulsifiers on the accumulation of quercetin metabolites in blood plasma after the short-term ingestion of onion by rats. Biosci. Biotechnol. Biochem. 2003, 67, 2548–2555. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, C.F.; Nemeroff, C.B. Hypercortisolemia and depression. Psychosom. Med. 2005, 67, S26. [Google Scholar] [CrossRef]
- Blackburn-Munro, G.; Blackburn-Munro, R.E. Chronic pain, chronic stress and depression: Coincidence or consequence? J. Neuroendocr. 2001, 13, 1009–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupien, S.J.; McEwen, B.S.; Gunnar, M.R.; Heim, C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat. Rev. Neurosci. 2009, 10, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Belleau, E.L.; Treadway, M.T.; Pizzagalli, D.A. The impact of stress and major depressive disorder on hippocampal and medial prefrontal cortex morphology. Biol. Psychiatry 2019, 85, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Smith, C.J.; Van Eldik, L.J. Importance of MAPK pathways for microglial pro-inflammatory cytokine IL-1β production. Neurobiol. Aging 2004, 25, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Quraishi, M.; Mokale, S.N.; Sakle, N.S. Ameliorative effect of quercetin and rutin via modulation of hypothalamic–pituitary–adrenal axis and regulation of fasting glucose in chronic stress-induced prediabetes. Pharmacogn. Mag. 2018, 14, 65–71. [Google Scholar]
- Park, S.H.; Sim, Y.B.; Han, P.L.; Lee, J.K.; Suh, H.W. Antidepressant-like effect of kaempferol and quercitirin, isolated from Opuntia ficus-indica var. saboten. Exp. Neurobiol. 2010, 19, 30. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zhang, H.; Wu, Z.; Yu, X.; Yin, Y.; Qian, S.; Wang, Z.; Huang, J.; Wang, W.; Liu, T.; et al. Quercitrin rapidly alleviated depression-like behaviors in lipopolysaccharide-treated mice: The involvement of PI3K/AKT/NF-κB signaling suppression and CREB/BDNF signaling restoration in the hippocampus. ACS Chem. Neurosci. 2021, 12, 3387–3396. [Google Scholar] [CrossRef]
- Dunn, J.D.; Alvarez, L.A.; Zhang, X.; Soldati, T. Reactive oxygen species and mitochondria: A nexus of cellular homeostasis. Redox Biol. 2015, 6, 472–485. [Google Scholar] [CrossRef]
- Lenertz, L.Y.; Gavala, M.L.; Hill, L.M.; Bertics, P.J. Cell signaling via the P2X7 nucleotide receptor: Linkage to ROS production, gene transcription, and receptor trafficking. Purinergic Signal. 2009, 5, 175–187. [Google Scholar] [CrossRef] [Green Version]
- Gottlieb, R.A. Role of mitochondria in apoptosis. Crit. Rev.™ Eukaryot. Gene Expr. 2000, 10, 10. [Google Scholar] [CrossRef]
- Liu, W.; Zhou, C. Corticosterone reduces brain mitochondrial function and expression of mitofusin, BDNF in depression-like rodents regardless of exercise preconditioning. Psychoneuroendocrinology 2012, 37, 1057–1070. [Google Scholar] [CrossRef]
- Gong, Y.; Chai, Y.; Ding, J.; Sun, X.; Hu, G. Chronic mild stress damages mitochondrial ultrastructure and function in mouse brain. Neurosci. Lett. 2011, 488, 76–80. [Google Scholar] [CrossRef]
- Manoli, I.; Alesci, S.; Blackman, M.R.; Su, Y.A.; Rennert, O.M.; Chrousos, G.P. Mitochondria as key components of the stress response. Trends Endocrinol. Metab. 2007, 18, 190–198. [Google Scholar] [CrossRef]
- Kokkinopoulou, I.; Moutsatsou, P. Mitochondrial glucocorticoid receptors and their actions. Int. J. Mol. Sci. 2021, 22, 6054. [Google Scholar] [CrossRef]
- Yu, X.L.; Li, Y.N.; Zhang, H.; Su, Y.J.; Zhou, W.W.; Zhang, Z.P.; Wang, S.W.; Xu, P.X.; Wang, Y.J.; Liu, R.T. Rutin inhibits amylin-induced neurocytotoxicity and oxidative stress. Food Funct. 2015, 6, 3296–3306. [Google Scholar] [CrossRef]
- Chang, W.T.; Huang, S.C.; Cheng, H.L.; Chen, S.C.; Hsu, C.L. Rutin and gallic acid regulates mitochondrial functions via the SIRT1 pathway in C2C12 myotubes. Antioxidants 2021, 10, 286. [Google Scholar] [CrossRef]
- Hamon, M.; Blier, P. Monoamine neurocircuitry in depression and strategies for new treatments. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 45, 54–63. [Google Scholar] [CrossRef]
- Cohen, S.; Janicki-Deverts, D.; Doyle, W.J.; Miller, G.E.; Frank, E.; Rabin, B.S.; Turner, R.B. Chronic stress, glucocorticoid receptor resistance, inflammation, and disease risk. Proc. Natl. Acad. Sci. USA 2012, 109, 5995–5999. [Google Scholar] [CrossRef] [Green Version]
- Richard, D.M.; Dawes, M.A.; Mathias, C.W.; Acheson, A.; Hill-Kapturczak, N.; Dougherty, D.M. L-tryptophan: Basic metabolic functions, behavioral research and therapeutic indications. Int. J. Trypotophan Res. 2009, 2, 2129. [Google Scholar] [CrossRef] [Green Version]
- Ewang-Emukowhate, M.; Nair, D.; Caplin, M. The role of 5-hydroxyindoleacetic acid in neuroendocrine tumors: The journey so far. Int. J. Endocr. Oncol. 2019, 6, 17. [Google Scholar] [CrossRef] [Green Version]
- Dunlop, B.W.; Nemeroff, C.B. The role of dopamine in the pathophysiology of depression. Arch. Gen. Psychiatry 2007, 64, 327–337. [Google Scholar] [CrossRef]
- Maurizi, C.P. The therapeutic potential for tryptophan and melatonin: Possible roles in depression, sleep, Alzheimer’s disease and abnormal aging. Med. Hypotheses 1990, 31, 233–242. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.; Wang, S.; Cao, Z.; Gu, D.; Wang, Y.; Tian, J.; Yang, Y. An efficient method based on an inhibitor–enzyme complex to screen an active compound against lipase from Toona sinensis. Food Funct. 2021, 12, 10806–10812. [Google Scholar] [CrossRef]
- Kim, M.H.; Jang, H.L.; Yoon, K.Y. Changes in physicochemical properties of Haetsun vegetables by blanching. J. Korean Soc. Food Sci. Nutr. 2012, 41, 647–654. [Google Scholar] [CrossRef] [Green Version]
- El-Fadl, H.M.I.A.; Morsy, G.M.; Hussein, R.H. Neuro-protective impact of rutin against methionine-induced hyperhomocysteinemia in rat model. Syst. Rev. Pharm. 2020, 11, 1042–1051. [Google Scholar]
- Li, J.; Liu, Q.T.; Chen, Y.; Liu, J.; Shi, J.L.; Liu, Y.; Guo, J.Y. Involvement of 5-HT1A receptors in the anxiolytic-like effects of quercitrin and evidence of the involvement of the monoaminergic system. Evid.-Based Complement. Altern. Med. 2016, 2016, 6530364. [Google Scholar]
- Gravanis, A.; Margioris, A.N. The corticotropin-releasing factor (CRF) family of neuropeptides in inflammation: Potential therapeutic applications. Curr. Med. Chem. 2005, 12, 1503–1512. [Google Scholar] [CrossRef]
- Dedovic, K.; Duchesne, A.; Andrews, J.; Engert, V.; Pruessner, J.C. The brain and the stress axis: The neural correlates of cortisol regulation in response to stress. Neuroimage 2009, 47, 864–871. [Google Scholar] [CrossRef]
- Zhu, W.; Chen, Z.; Li, Q.; Tan, G.; Hu, G. Inhibitors of 11β-hydroxylase (CYP11B1) for treating diseases related to excess cortisol. Curr. Med. Chem. 2016, 23, 623–633. [Google Scholar] [CrossRef]
- Ye, P.; Kenyon, C.J.; MacKenzie, S.M.; Nichol, K.; Seckl, J.R.; Fraser, R.; Connell, J.M.C.; Davies, E. Effects of ACTH, dexamethasone, and adrenalectomy on 11β-hydroxylase (CYP11B1) and aldosterone synthase (CYP11B2) gene expression in the rat central nervous system. J. Endocrinol. 2008, 196, 305–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.X.; Zhang, R.Y.; Rui, W.J.; Wang, Z.Q.; Feng, X. Quercetin alleviates chronic unpredictable mild stress-induced depressive-like behaviors by promoting adult hippocampal neurogenesis via FoxG1/CREB/BDNF signaling pathway. Behav. Brain Res. 2021, 406, 113245. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.T.; Ruan, R.; Tay, F.E.H. Relationship between levels of oxidative DNA damage, lipid peroxidation and mitochondrial membrane potential in young and old F344 rats. Free Radic. Res. 2006, 40, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Yirmyia, R.; Noraberg, J.; Brene, S.; Hibbeln, J.; Perini, G.; Kubera, M.; Bob, P.; Lerer, B.; Maj, M. The inflammatory & neurodegenerative (I&ND) hypothesis of depression: Leads for future research and new drug developments in depression. Metab. Brain Dis. 2009, 24, 27–53. [Google Scholar]
- Yang, J.; Yao, S. JNK-Bcl-2/Bcl-xL-Bax/Bak pathway mediates the crosstalk between matrine-induced autophagy and apoptosis via interplay with beclin 1. Int. J. Mol. Sci. 2015, 16, 25744–25758. [Google Scholar] [CrossRef] [Green Version]
- Kwak, C.S.; Kim, M.J.; Park, S.; Kim, I.G. Antioxidant activity of sprouts extracts is correlated with their anti-obesity and anti-inflammatory effects in high-fat diet-fed mice. Evid.-Based Complement. Altern. Med. 2021, 2021, 8367802. [Google Scholar] [CrossRef]
- Mostafa, D.G.; Khaleel, E.F.; Badi, R.M.; Abdel-Aleem, G.A.; Abdeen, H.M. Rutin hydrate inhibits apoptosis in the brains of cadmium chloride-treated rats via preserving the mitochondrial integrity and inhibiting endoplasmic reticulum stress. Neurol. Res. 2019, 41, 594–608. [Google Scholar] [CrossRef]
- Nkpaa, K.W.; Onyeso, G.I. Rutin attenuates neurobehavioral deficits, oxidative stress, neuro-inflammation and apoptosis in fluoride treated rats. Neurosci. Lett. 2018, 682, 92–99. [Google Scholar] [CrossRef]
- Çelik, H.; Kandemir, F.M.; Caglayan, C.; Özdemir, S.; Çomaklı, S.; Kucukler, S.; Yardım, A. Neuroprotective effect of rutin against colistin-induced oxidative stress, inflammation and apoptosis in rat brain associated with the CREB/BDNF expressions. Mol. Biol. Rep. 2020, 47, 2023–2034. [Google Scholar] [CrossRef]
- Kim, J.M.; Park, S.K.; Guo, T.J.; Kang, J.Y.; Ha, J.S.; Lee, U.; Heo, H.J. Anti-amnesic effect of Dendropanax morbifera via JNK signaling pathway on cognitive dysfunction in high-fat diet-induced diabetic mice. Behav. Brain Res. 2016, 312, 39–54. [Google Scholar] [CrossRef]
- Liu, L.; Huang, S.; Xu, M.; Gong, Y.; Li, D.; Wan, C.; Wu, H.; Tang, Q. Isoquercitrin protects HUVECs against high glucose-induced apoptosis through regulating p53 proteasomal degradation. Int. J. Mol. Med. 2021, 48, 122. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.Q.; Luo, R.Z.; Jiang, H.X.; Liu, C.M. Quercitrin offers protection against brain injury in mice by inhibiting oxidative stress and inflammation. Food Funct. 2016, 7, 549–556. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stressors | Time | Description |
|---|---|---|
| Cage swap | 24 h | Mice were exchanged from their cage to a stranger cage with other mice. |
| Cage tilting | 24 h | Each cage was tilted by leaning at a 45° angle on the cage. |
| Empty cage | 24 h | Mice were housed in empty cages without sawdust. |
| Food or water deprivation | 24 h | Food or water was diverted. |
| Mild restraint | 2 h | Mice were placed in the plastic box (10.5 cm width × 10.5 cm length × 5.5 cm height) with holes in every side except the bottom and top to breathe. |
| Overnight light exposure | 24 h | Mice were exposed to light stress. |
| Wet bedding | 24 h | Mice were housed in cages with added distilled water (200 mL). |
| No. | RT (1) (min) | Parent Iron (2) (m/z) | MSE Fragment (3) (m/z) | Compound |
|---|---|---|---|---|
| 1 | 3.71 | 609.14 | 301.04, 151.00 | Rutin |
| 2 | 3.82 | 463.09 | 300.02, 301.03 | Isoquercitrin |
| 3 | 3.91 | 197.04 | 169.01, 124.01 | Ethyl gallate |
| 4 | 4.05 | 447.09 | 301.03, 271.02, 179.00 | Quercitrin |
| 5 | 4.28 | 431.10 | 285.04, 284.03 | Kaempferol-3-O-rhamnoside |
| 6 | 4.50 | 349.05 | 197.04, 169.01 | Ethyl digallate |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, H.R.; Kim, J.M.; Lee, U.; Kang, J.Y.; Park, S.K.; Lee, H.L.; Moon, J.H.; Kim, M.J.; Go, M.J.; Heo, H.J. Leaves of Cedrela sinensis Attenuate Chronic Unpredictable Mild Stress-Induced Depression-like Behavior via Regulation of Hormonal and Inflammatory Imbalance. Antioxidants 2022, 11, 2448. https://doi.org/10.3390/antiox11122448
Jeong HR, Kim JM, Lee U, Kang JY, Park SK, Lee HL, Moon JH, Kim MJ, Go MJ, Heo HJ. Leaves of Cedrela sinensis Attenuate Chronic Unpredictable Mild Stress-Induced Depression-like Behavior via Regulation of Hormonal and Inflammatory Imbalance. Antioxidants. 2022; 11(12):2448. https://doi.org/10.3390/antiox11122448
Chicago/Turabian StyleJeong, Hye Rin, Jong Min Kim, Uk Lee, Jin Yong Kang, Seon Kyeong Park, Hyo Lim Lee, Jong Hyun Moon, Min Ji Kim, Min Ji Go, and Ho Jin Heo. 2022. "Leaves of Cedrela sinensis Attenuate Chronic Unpredictable Mild Stress-Induced Depression-like Behavior via Regulation of Hormonal and Inflammatory Imbalance" Antioxidants 11, no. 12: 2448. https://doi.org/10.3390/antiox11122448