
Protective Actions of α-Tocopherol on Cell Membrane Lipids of Paraquat-Stressed Human Astrocytes Using Microarray Technology, MALDI-MS and Lipidomic Analysis
 , , , and
, , , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Drugs and Reagents
2.2. Cell Culture and Treatments
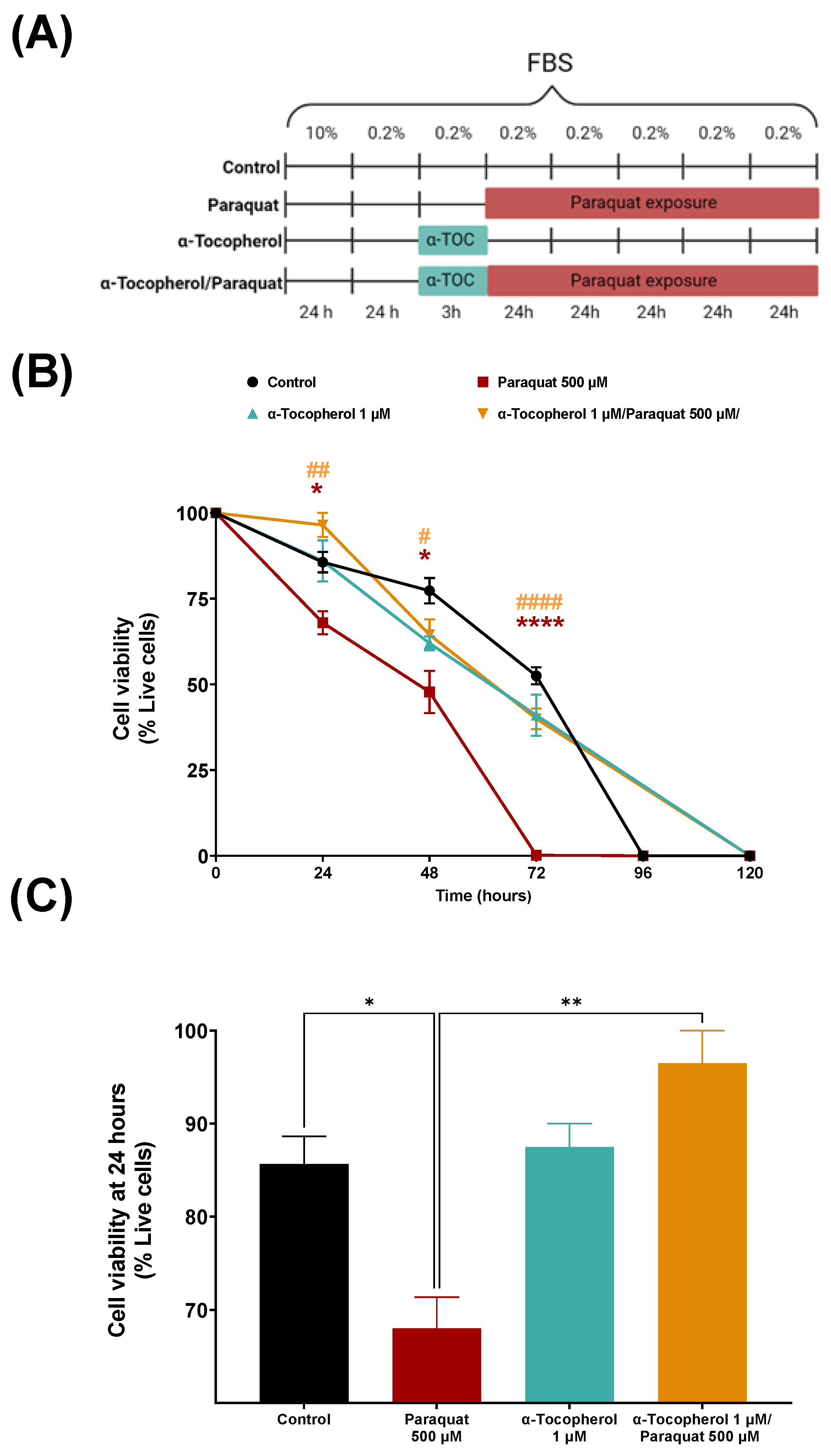
2.3. Viability Assay
2.4. Cell Membrane Extraction and CMMA Fabrication
2.5. MALDI-MS Lipidomic Analysis
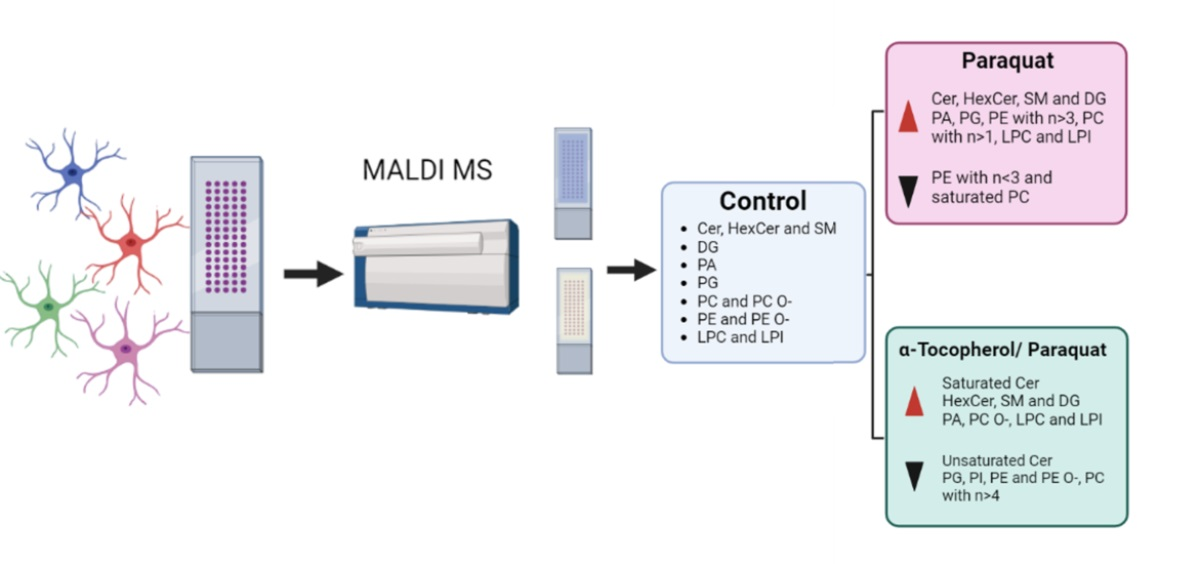
3. Results
3.1. Viability of Human Astrocytic Cells upon Different Treatments
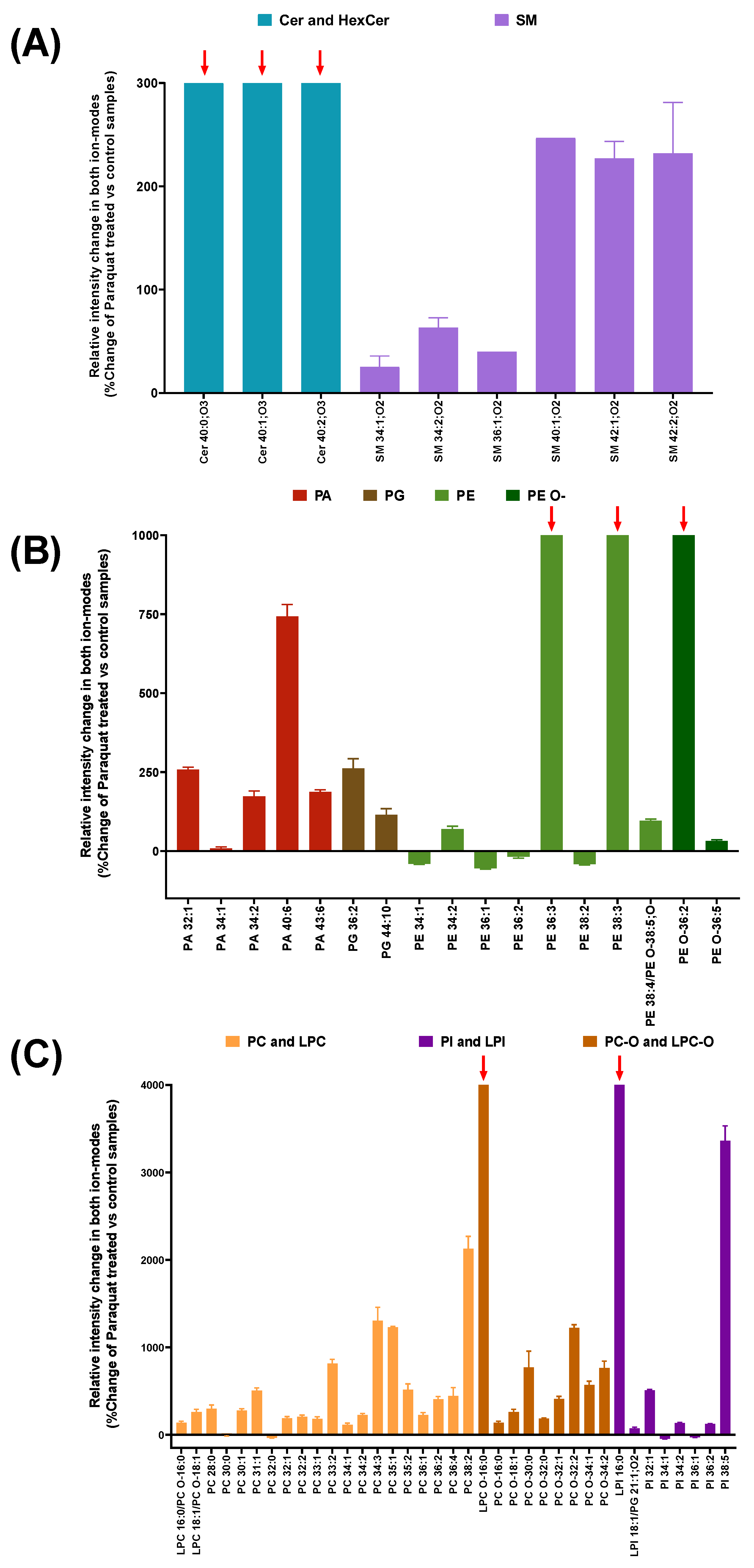
3.2. Lipidomic Analysis in Cell Membrane Microarrys Is Able to Reveal Paraquat-Triggered Changes in Human Astrocytic Membranes
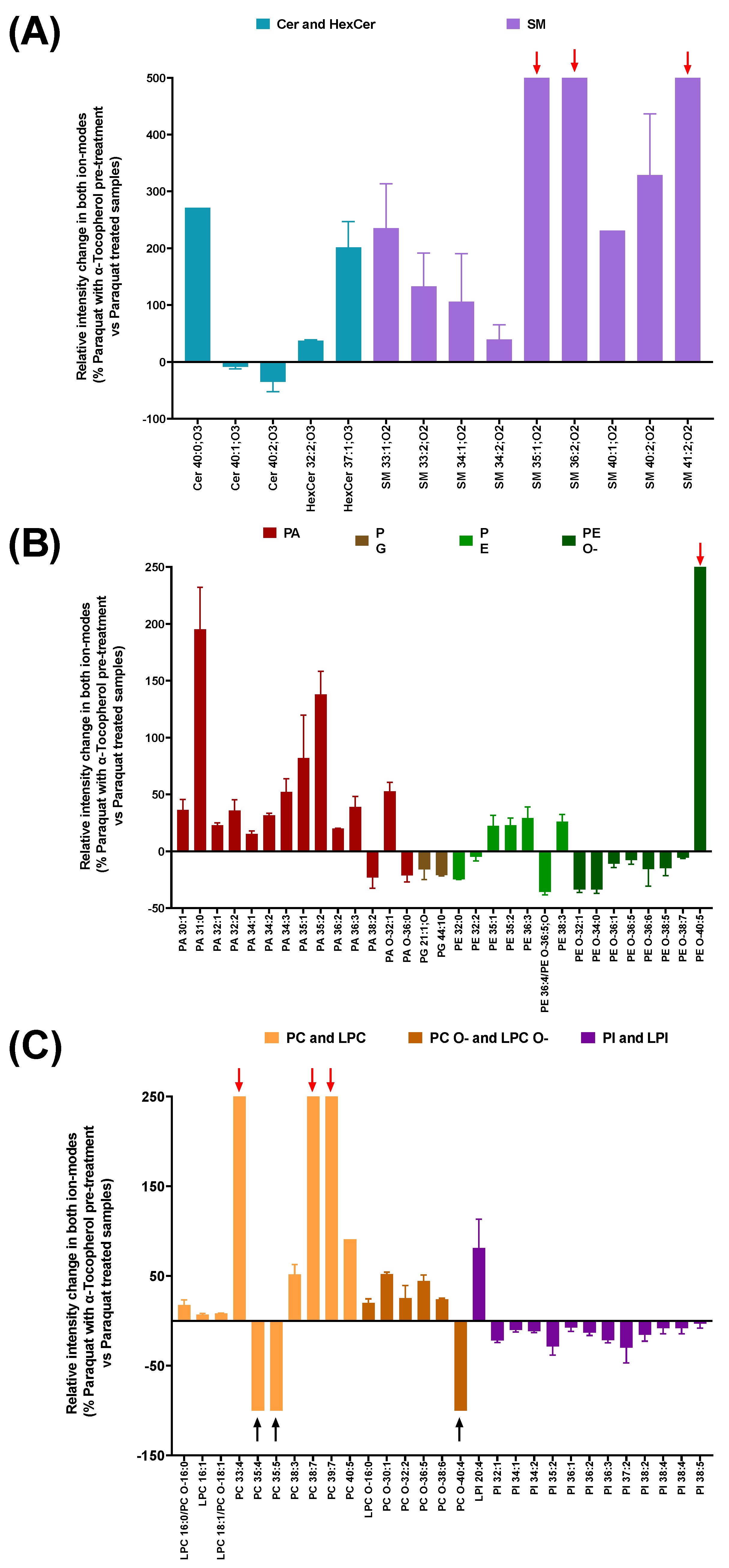
3.3. Lipidomic Analysis in Cell Membrane Microarrys Is Able to Reveal the Effect of α-Tocopherol Pre-Treatment on the Paraquat-Triggered Signature
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yin, H.; Xu, L.; Porter, N.A. Free Radical Lipid Peroxidation: Mechanisms and Analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef] [PubMed]
- Girotti, A.W. Nitric Oxide-Elicited Resistance to Antitumor Photodynamic Therapy via Inhibition of Membrane Free Radical-Mediated Lipid Peroxidation. Photochem. Photobiol. 2021, 97, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Niki, E.; Yoshida, Y.; Saito, Y.; Noguchi, N. Lipid Peroxidation: Mechanisms, Inhibition, and Biological Effects. Biochem. Biophys. Res. Commun. 2005, 338, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Niki, E. Lipid Peroxidation Products as Oxidative Stress Biomarkers. Biofactors 2008, 34, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Gaschler, M.M.; Stockwell, B.R. Lipid Peroxidation in Cell Death. Biochem. Biophys. Res. Commun. 2017, 482, 419–425. [Google Scholar] [CrossRef]
- Senoner, T.; Dichtl, W. Oxidative Stress in Cardiovascular Diseases: Still a Therapeutic Target? Nutrients 2019, 11, 2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramana, K.V.; Srivastava, S.; Singhal, S.S. Lipid Peroxidation Products in Human Health and Disease 2014. Oxid. Med. Cell Longev. 2014, 2014, 162414. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Chirico, S. Lipid Peroxidation: Its Mechanism, Measurement, and Significance. Am. J. Clin. Nutr. 1993, 57, 715S–724S. [Google Scholar] [CrossRef] [Green Version]
- Jaganjac, M.; Cindrić, M.; Jakovčević, A.; Žarković, K.; Žarković, N. Lipid Peroxidation in Brain Tumors. Neurochem. Int. 2021, 149, 105118. [Google Scholar] [CrossRef]
- Catalá, A. Lipid Peroxidation of Membrane Phospholipids Generates Hydroxy-Alkenals and Oxidized Phospholipids Active in Physiological and/or Pathological Conditions. Chem. Phys. Lipids 2009, 157, 1–11. [Google Scholar] [CrossRef]
- Venkataraman, S.; Schafer, F.Q.; Buettner, G.R. Detection of Lipid Radicals Using EPR. Antioxid. Redox Signal. 2004, 6, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Greene, L.E.; Lincoln, R.; Cosa, G. Spatio-Temporal Monitoring of Lipid Peroxyl Radicals in Live Cell Studies Combining Fluorogenic Antioxidants and Fluorescence Microscopy Methods. Free Radic. Biol. Med. 2018, 128, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Guéraud, F.; Atalay, M.; Bresgen, N.; Cipak, A.; Eckl, P.M.; Huc, L.; Jouanin, I.; Siems, W.; Uchida, K. Chemistry and Biochemistry of Lipid Peroxidation Products. Free Radic. Res. 2010, 44, 1098–1124. [Google Scholar] [CrossRef] [PubMed]
- del Rio, D.; Stewart, A.J.; Pellegrini, N. A Review of Recent Studies on Malondialdehyde as Toxic Molecule and Biological Marker of Oxidative Stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Mills, K.; le Cessie, S.; Noordam, R.; van Heemst, D. Ageing, Age-Related Diseases and Oxidative Stress: What to Do Next? Ageing Res. Rev. 2020, 57, 100982. [Google Scholar] [CrossRef]
- Chen, Z.; Zhong, C. Oxidative Stress in Alzheimer’s Disease. Neurosci. Bull. 2014, 30, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Lian, G. ROS and Diseases: Role in Metabolism and Energy Supply. Mol. Cell Biochem. 2020, 467, 1–12. [Google Scholar] [CrossRef]
- Elexpe, A.; Sánchez-Sánchez, L.; Tolentino-Cortez, T.; Astigarraga, E.; Torrecilla, M.; Barreda-Gómez, G. Analysis of Mitochondrial Function in Cell Membranes as Indicator of Tissue Vulnerability to Drugs in Humans. Biomedicines 2022, 10, 980. [Google Scholar] [CrossRef]
- Youdim, M.B.H.; Edmondson, D.; Tipton, K.F. The Therapeutic Potential of Monoamine Oxidase Inhibitors. Nat. Rev. Neurosci. 2006, 7, 295–309. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.H. The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Brieger, K.; Schiavone, S.; Miller, F.J.; Krause, K.H. Reactive Oxygen Species: From Health to Disease. Swiss Med. Wkly. 2012, 142, w13659. [Google Scholar] [CrossRef] [PubMed]
- Holley, A.K.; Dhar, S.K.; Xu, Y.; Clair, D.K.S. Manganese Superoxide Dismutase: Beyond Life and Death. Amino Acids 2012, 42, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Tiffany-Castiglioni, E. Paraquat-Induced Apoptosis in Human Neuroblastoma SH-SY5Y Cells: Involvement of P53 and Mitochondria. J. Toxicol. Environ. Health A 2008, 71, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Ayala, T.; Andérica-Romero, A.C.; Pedraza-Chaverri, J. New Insights into Antioxidant Strategies against Paraquat Toxicity. Free Radic. Res. 2014, 48, 623–640. [Google Scholar] [CrossRef]
- Han, J.; Zhang, Z.; Yang, S.; Wang, J.; Yang, X.; Tan, D. Betanin Attenuates Paraquat-Induced Liver Toxicity through a Mitochondrial Pathway. Food Chem Toxicol. 2014, 70, 100–106. [Google Scholar] [CrossRef]
- Vicente, J.A.; Peixoto, F.; Lopes, M.L.; Madeira, V.M. Differential Sensitivities of Plant and Animal Mitochondria to the Herbicide Paraquat. J. Biochem. Mol. Toxicol. 2001, 15, 322–330. [Google Scholar] [CrossRef] [Green Version]
- Reczek, C.R.; Birsoy, K.; Kong, H.; Martínez-Reyes, I.; Wang, T.; Gao, P.; Sabatini, D.M.; Chandel, N.S. A CRISPR Screen Identifies a Pathway Required for Paraquat-Induced Cell Death. Nat. Chem. Biol. 2017, 13, 1274. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.-Y.; Xu, X.; Li, X.-C.; Zhang, P. Cardiovascular Diseases: Oxidative Damage and Antioxidant Protection. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 3091–3096. [Google Scholar]
- Jiang, Q. Natural Forms of Vitamin E: Metabolism, Antioxidant, and Anti-Inflammatory Activities and Their Role in Disease Prevention and Therapy. Free Radic. Biol. Med. 2014, 72, 76–90. [Google Scholar] [CrossRef] [Green Version]
- Niki, E.; Noguchi, N. Antioxidant Action of Vitamin E in Vivo as Assessed from Its Reaction Products with Multiple Biological Oxidants. Free Radic. Res. 2021, 55, 352–363. [Google Scholar] [CrossRef]
- Zingg, J.M. Vitamin E: Regulatory Role on Signal Transduction. IUBMB Life 2019, 71, 456–478. [Google Scholar] [CrossRef]
- Miyazawa, T.; Burdeos, G.C.; Itaya, M.; Nakagawa, K.; Miyazawa, T. Vitamin E: Regulatory Redox Interactions. IUBMB Life 2019, 71, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Peña-Bautista, C.; Baquero, M.; Vento, M.; Cháfer-Pericás, C. Free Radicals in Alzheimer’s Disease: Lipid Peroxidation Biomarkers. Clin. Chim. Acta 2019, 491, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Devaraj, S.; Tang, R.; Adams-Huet, B.; Harris, A.; Seenivasan, T.; de Lemos, J.A.; Jialal, I. Effect of High-Dose α-Tocopherol Supplementation on Biomarkers of Oxidative Stress and Inflammation and Carotid Atherosclerosis in Patients with Coronary Artery Disease. Am. J. Clin. Nutr. 2007, 86, 1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, R.; Garate, J.; Tolentino-Cortez, T.; Herraiz, A.; Lombardero, L.; Ducrocq, F.; Rodríguez-Puertas, R.; Trifilieff, P.; Astigarraga, E.; Barreda-Gómez, G.; et al. Microarray and Mass Spectrometry-Based Methodology for Lipid Profiling of Tissues and Cell Cultures. Anal. Chem. 2019, 91, 15967–15973. [Google Scholar] [CrossRef]
- Yamamoto, N.; Yamashita, Y.; Yoshioka, Y.; Nishiumi, S.; Ashida, H. Rapid Preparation of a Plasma Membrane Fraction: Western Blot Detection of Translocated Glucose Transporter 4 from Plasma Membrane of Muscle and Adipose Cells and Tissues. Curr. Protoc. Protein Sci. 2016, 85, 29.18.1–29.18.12. [Google Scholar] [CrossRef]
- Nishiumi, S.; Ashida, H. Rapid Preparation of a Plasma Membrane Fraction from Adipocytes and Muscle Cells: Application to Detection of Translocated Glucose Transporter 4 on the Plasma Membrane. Biosci. Biotechnol. Biochem. 2007, 71, 2343–2346. [Google Scholar] [CrossRef] [Green Version]
- Rienda, B.; Elexpe, A.; Tolentino-Cortez, T.; Gulak, M.; Bruzos-Cidón, C.; Torrecilla, M.; Astigarraga, E.; Barreda-Gómez, G. Analysis of Acetylcholinesterase Activity in Cell Membrane Microarrays of Brain Areas as a Screening Tool to Identify Tissue Specific Inhibitors. Analytica 2021, 2, 25–36. [Google Scholar] [CrossRef]
- Elexpe, A.; Nieto, N.; Fernández-Cuétara, C.; Domínguez-Fernández, C.; Morera-Herreras, T.; Torrecilla, M.; Miguélez, C.; Laso, A.; Ochoa, E.; Bailen, M.; et al. Study of Tissue-Specific Reactive Oxygen Species Formation by Cell Membrane Microarrays for the Characterization of Bioactive Compounds. Membranes 2021, 11, 943. [Google Scholar] [CrossRef]
- Manuel, I.; Barreda-Gomez, G.; González De San Román, E.; Veloso, A.; Fernández, J.A.; Giralt, M.T.; Rodriguez-Puertas, R. Neurotransmitter Receptor Localization: From Autoradiography to Imaging Mass Spectrometry. ACS Chem. Neurosci. 2015, 6, 362–373. [Google Scholar] [CrossRef]
- Hebert-Chatelain, E.; Desprez, T.; Serrat, R.; Bellocchio, L.; Soria-Gomez, E.; Busquets-Garcia, A.; Pagano Zottola, A.C.; Delamarre, A.; Cannich, A.; Vincent, P.; et al. A Cannabinoid Link between Mitochondria and Memory. Nature 2016, 539, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Astigarraga, E.; Barreda-Gómez, G.; Lombardero, L.; Fresnedo, O.; Castaño, F.; Giralt, M.T.; Ochoa, B.; Rodríguez-Puertas, R.; Fernández, J.A. Profiling and Imaging of Lipids on Brain and Liver Tissue by Matrix-Assisted Laser Desorption/ Ionization Mass Spectrometry Using 2-Mercaptobenzothiazole as a Matrix. Anal. Chem. 2008, 80, 9105–9114. [Google Scholar] [CrossRef] [PubMed]
- Suntres, Z.E.; Hepworth, S.R.; Shek, P.N. Protective Effect of Liposome-Associated α-Tocopherol against Paraquat-Induced Acute Lung Toxicity. Biochem. Pharmacol. 1992, 44, 1811–1818. [Google Scholar] [CrossRef] [PubMed]
- Azzi, A.; Gysin, R.; Kempná, P.; Ricciarelli, R.; Villacorta, L.; Visarius, T.; Zingg, J.M. The Role of α-Tocopherol in Preventing Disease: From Epidemiology to Molecular Events. Mol. Asp. Med. 2003, 24, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Zakharova, I.O.; Bayunova, L.V.; Zorina, I.I.; Sokolova, T.V.; Shpakov, A.O.; Avrova, N.F. Insulin and α-Tocopherol Enhance the Protective Effect of Each Other on Brain Cortical Neurons under Oxidative Stress Conditions and in Rat Two-Vessel Forebrain Ischemia/Reperfusion Injury. Int. J. Mol. Sci. 2021, 22, 11768. [Google Scholar] [CrossRef] [PubMed]
- Duncan, K.R.; Suzuki, Y.J. Vitamin E Nicotinate. Antioxidants 2017, 6, 20. [Google Scholar] [CrossRef] [Green Version]
- Shayman, J.A.; Tesmer, J.J.G. Lysosomal Phospholipase A2. Biochim Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 932. [Google Scholar] [CrossRef]
- Nikolova-Karakashian, M.N.; Rozenova, K.A. Ceramide in Stress Response. Adv. Exp. Med. Biol. 2010, 688, 86–108. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. Principles of Bioactive Lipid Signalling: Lessons from Sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef]
- Ho, Q.W.C.; Zheng, X.; Ali, Y. Ceramide Acyl Chain Length and Its Relevance to Intracellular Lipid Regulation. Int. J. Mol. Sci. 2022, 23, 9697. [Google Scholar] [CrossRef]
- Thomas, J.M.; Sudhadevi, T.; Basa, P.; Ha, A.W.; Natarajan, V.; Harijith, A. The Role of Sphingolipid Signaling in Oxidative Lung Injury and Pathogenesis of Bronchopulmonary Dysplasia. Int. J. Mol. Sci. 2022, 23, 1254. [Google Scholar] [CrossRef] [PubMed]
- Jazvinšćak Jembrek, M.; Hof, P.R.; Šimić, G. Ceramides in Alzheimer’s Disease: Key Mediators of Neuronal Apoptosis Induced by Oxidative Stress and Aβ Accumulation. Oxid. Med. Cell Longev. 2015, 2015, 346783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, B.N.; Oyeniran, C.; Sturgill, J.L.; Newton, J.; Martin, R.K.; Bieberich, E.; Weigel, C.; Maczis, M.A.; Palladino, E.N.D.; Lownik, J.C.; et al. Ceramide in apoptosis and oxidative stress in allergic inflammation and asthma. J. Allergy Clin. Immunol 2021, 147, 1936. [Google Scholar] [CrossRef] [PubMed]
- Zwara, A.; Wertheim-Tysarowska, K.; Mika, A. Alterations of Ultra Long-Chain Fatty Acids in Hereditary Skin Diseases—Review Article. Front. Med. 2021, 8, 730855. [Google Scholar] [CrossRef] [PubMed]
- Panchal, M.; Gaudin, M.; Lazar, A.N.; Salvati, E.; Rivals, I.; Ayciriex, S.; Dauphinot, L.; Dargère, D.; Auzeil, N.; Masserini, M.; et al. Ceramides and Sphingomyelinases in Senile Plaques. Neurobiol. Dis. 2014, 65, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Poleschuk, T.S.; Sultanov, R.M.; Ermolenko, E.V.; Shulgina, L.V.; Kasyanov, S.P. Protective Action of Alkylglycerols under Stress. Stress 2020, 23, 213–220. [Google Scholar] [CrossRef]
- Sindelar, P.J.; Guan, Z.; Dallner, G.; Ernster, L. The Protective Role of Plasmalogens in Iron-Induced Lipid Peroxidation. Free Radic. Biol. Med. 1999, 26, 318–324. [Google Scholar] [CrossRef]
- Brites, P.; Waterham, H.R.; Wanders, R.J.A. Functions and Biosynthesis of Plasmalogens in Health and Disease. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2004, 1636, 219–231. [Google Scholar] [CrossRef]
- Drechsler, R.; Chen, S.-W.; Dancy, B.C.R.; Mehrabkhani, L.; Olsen, C.P. HPLC-Based Mass Spectrometry Characterizes the Phospholipid Alterations in Ether-Linked Lipid Deficiency Models Following Oxidative Stress. PLoS ONE 2016, 11, e0167229. [Google Scholar] [CrossRef] [Green Version]
- Messias, M.C.F.; Mecatti, G.C.; Priolli, D.G.; de Oliveira Carvalho, P. Plasmalogen Lipids: Functional Mechanism and Their Involvement in Gastrointestinal Cancer. Lipids Health Dis. 2018, 17, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Janssen, C.I.F.; Kiliaan, A.J. Long-Chain Polyunsaturated Fatty Acids (LCPUFA) from Genesis to Senescence: The Influence of LCPUFA on Neural Development, Aging, and Neurodegeneration. Prog. Lipid Res. 2014, 53, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mîinea, C.; Kuruvilla, R.; Merrikh, H.; Eichberg, J. Altered Arachidonic Acid Biosynthesis and Antioxidant Protection Mechanisms in Schwann Cells Grown in Elevated Glucose. J. Neurochem. 2002, 81, 1253–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, A.; Oka, S.; Tanikawa, T.; Hayashi, Y.; Nemoto-Sasaki, Y.; Sugiura, T. The Actions and Metabolism of Lysophosphatidylinositol, an Endogenous Agonist for GPR55. Prostaglandins Lipid Mediat. 2013, 107, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Okuda, S.; Saito, H.; Katsuki, H. Arachidonic Acid: Toxic and Trophic Effects on Cultured Hippocampal Neurons. Neuroscience 1994, 63, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Lee, J.J.; Chen, Y.J.; Lin, S.I.; Lin, L.D.; Liou, E.J.W.; Huang, W.L.; Chan, C.P.; Huang, C.C.; Jeng, J.H. Lysophosphatidylcholine Induces Cytotoxicity/Apoptosis and IL-8 Production of Human Endothelial Cells: Related Mechanisms. Oncotarget 2017, 8, 106177. [Google Scholar] [CrossRef] [Green Version]
- Charollais, J.; van der Goot, F.G. Palmitoylation of Membrane Proteins (Review). Mol. Membr. Biol. 2009, 26, 55–66. [Google Scholar] [CrossRef]
- Koca, T.; Canatan, D.; Örmeci, A.R.; Koca, Y.S.; Duman, H.; Baykal, A.; Akçam, M. Amino Acids and Fatty Acids in Patients with Beta Thalassemia Major. Acta Bio. Med. Atenei Parm. 2020, 91, 79. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Sánchez, L.; Fernández, R.; Ganfornina, M.D.; Astigarraga, E.; Barreda-Gómez, G. Protective Actions of α-Tocopherol on Cell Membrane Lipids of Paraquat-Stressed Human Astrocytes Using Microarray Technology, MALDI-MS and Lipidomic Analysis. Antioxidants 2022, 11, 2440. https://doi.org/10.3390/antiox11122440
Sánchez-Sánchez L, Fernández R, Ganfornina MD, Astigarraga E, Barreda-Gómez G. Protective Actions of α-Tocopherol on Cell Membrane Lipids of Paraquat-Stressed Human Astrocytes Using Microarray Technology, MALDI-MS and Lipidomic Analysis. Antioxidants. 2022; 11(12):2440. https://doi.org/10.3390/antiox11122440
Chicago/Turabian StyleSánchez-Sánchez, Laura, Roberto Fernández, Maria Dolores Ganfornina, Egoitz Astigarraga, and Gabriel Barreda-Gómez. 2022. "Protective Actions of α-Tocopherol on Cell Membrane Lipids of Paraquat-Stressed Human Astrocytes Using Microarray Technology, MALDI-MS and Lipidomic Analysis" Antioxidants 11, no. 12: 2440. https://doi.org/10.3390/antiox11122440