
Neonatal Selenium Deficiency Decreases Selenoproteins in the Lung and Impairs Pulmonary Alveolar Development

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Model of Neonatal Se Deficiency
2.2. Collection of Blood and Organs
2.3. Morphometric Analysis
2.4. Glutathione Peroxidase Activity
2.5. Thioredoxin Reductase (Txnrd) Activity Level
2.6. Immunoblot Analysis
2.7. Analysis of Relative mRNA Levels by RT-qPCR
2.8. Statistical Analysis
3. Results
3.1. Neonatal Se Deficiency Impairs Postnatal Growth at Day of Life 7 through Adulthood
3.2. Neonatal Se Deficiency Impairs Alveolar Development
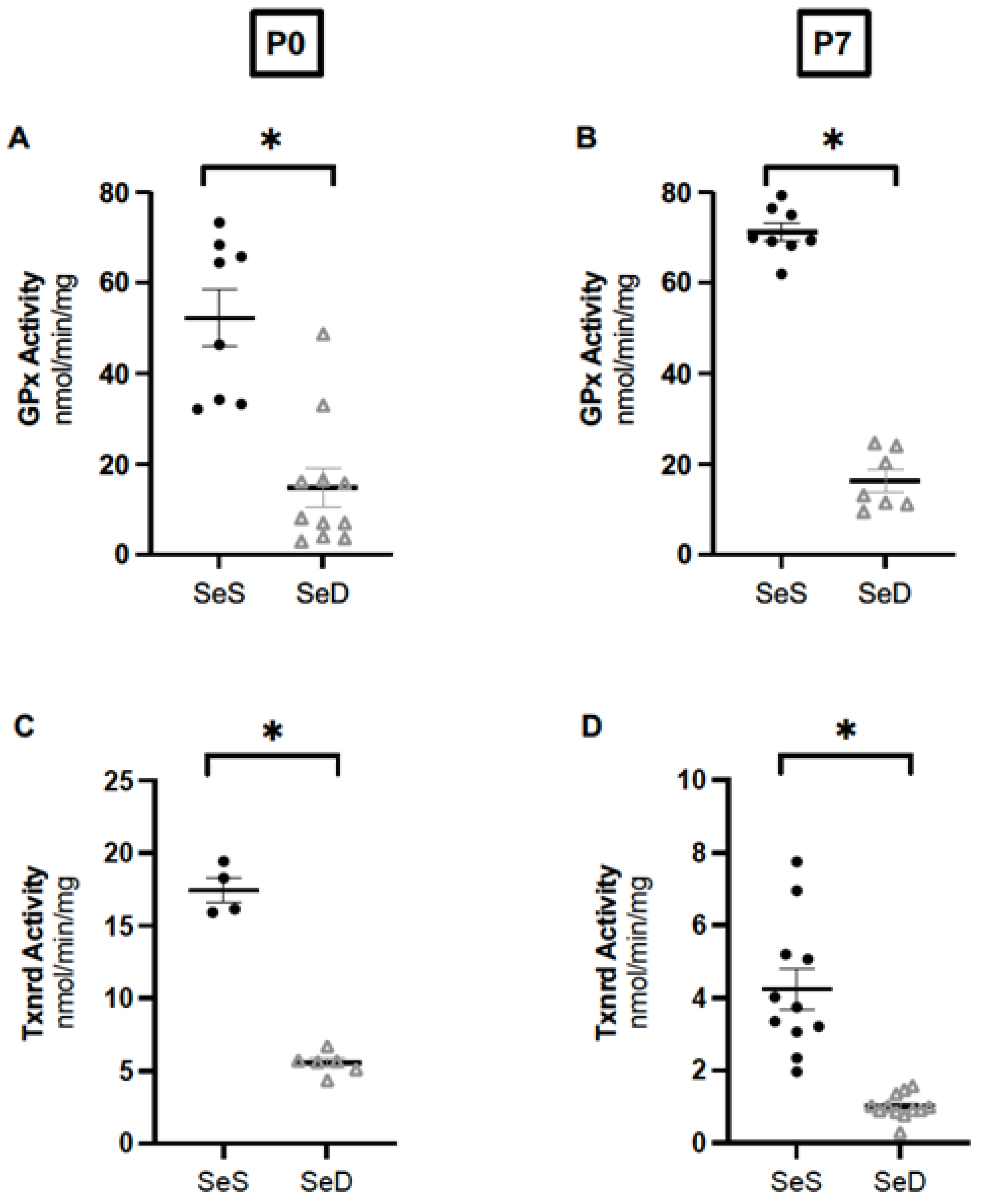
3.3. Neonatal Se Deficiency Decreases Glutathione Peroxidase and Thioredoxin Reductase Activity in the Lung
3.4. Neonatal Se Deficiency Decreases Pulmonary GPx1, GPx3 and Txnrd1 Content; Txnrd2 Content Is Preserved
3.5. Neonatal Se Deficiency Decreases Transcription of Pulmonary Glutathione Peroxidase 1 and 3
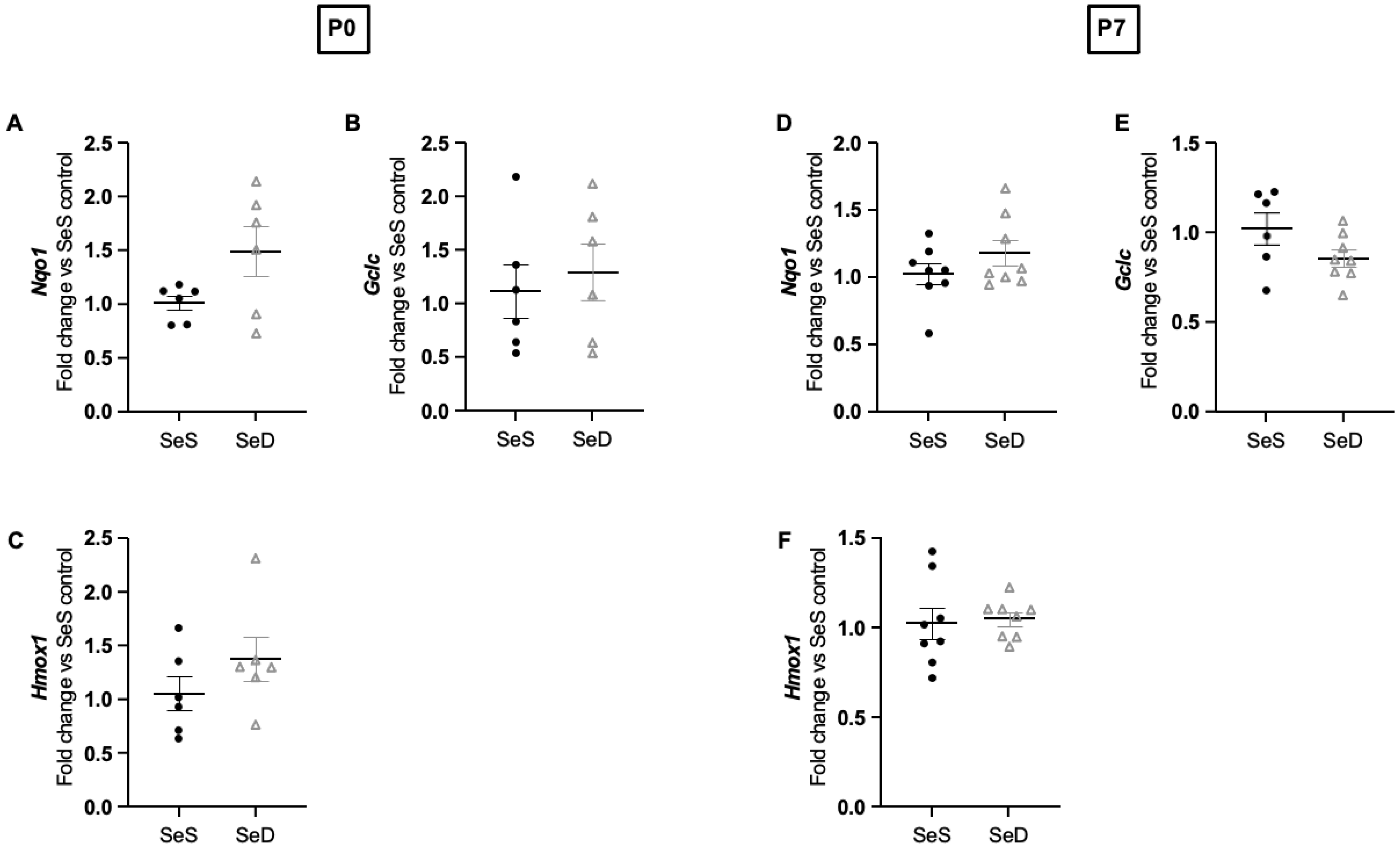
3.6. Neonatal Se Deficiency Does Not Alter Transcription of NRF-2 Targets in the Lung
3.7. Neonatal Se Deficiency Does Not Increase Pulmonary Content of Non-Se Containing Antioxidant Enzymes at P0 or P7
3.8. Neonatal Se Deficiency Decreases Selenoprotein N, an Endoplasmic Reticulum Selenoprotein Implicated in Normal Alveolar Development, at Both P0 and P7
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klevebro, S.; Westin, V.; Stoltz Sjöström, E.; Norman, M.; Domellöf, M.; Edstedt Bonamy, A.K.; Hallberg, B. Early energy and protein intakes and associations with growth, BPD, and ROP in extremely preterm infants. Clin. Nutr. 2019, 38, 1289–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dani, C.; Poggi, C. Nutrition and bronchopulmonary dysplasia. J. Matern. Fetal. Neonatal Med. 2012, 25 (Suppl. 3), 37–40. [Google Scholar] [CrossRef] [PubMed]
- Arigliani, M.; Stocco, C.; Valentini, E.; De Pieri, C.; Castriotta, L.; Ferrari, M.E.; Canciani, C.; Driul, L.; Orsaria, M.; Cattarossi, L.; et al. Lung function between 8 and 15 years of age in very preterm infants with fetal growth restriction. Pediatr. Res. 2021, 90, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.; Lunt, A.; Bisquera, A.; Peacock, J.; Greenough, A. Intrauterine growth retardation and lung function of very prematurely born young people. Pediatr. Pulmonol. 2021, 56, 2284–2291. [Google Scholar] [CrossRef] [PubMed]
- Regev, R.H.; Reichman, B. Prematurity and intrauterine growth retardation–double jeopardy? Clin. Perinatol. 2004, 31, 453–473. [Google Scholar] [CrossRef]
- Darlow, B.A.; Winterbourn, C.C.; Inder, T.E.; Graham, P.J.; Harding, J.E.; Weston, P.J.; Austin, N.C.; Elder, D.E.; Mogridge, N.; Buss, I.H.; et al. The effect of selenium supplementation on outcome in very low birth weight infants: A randomized controlled trial. J. Pediatr. 2000, 136, 473–480. [Google Scholar] [CrossRef]
- Darlow, B.A.; Austin, N.C. Selenium supplementation to prevent short-term morbidity in preterm neonates. Cochrane Database Syst. Rev. 2003, 2011, CD003312. [Google Scholar] [CrossRef]
- Ramel, S.E.; Brown, L.D.; Georgieff, M.K. The Impact of Neonatal Illness on Nutritional Requirements-One Size Does Not Fit All. Curr. Pediatr. Rep. 2014, 2, 248–254. [Google Scholar] [CrossRef] [Green Version]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Schomburg, L. Dietary Selenium and Human Health. Nutrients 2016, 9, 22. [Google Scholar] [CrossRef]
- Jones, G.D.; Droz, B.; Greve, P.; Gottschalk, P.; Poffet, D.; McGrath, S.P.; Seneviratne, S.I.; Smith, P.; Winkel, L.H. Selenium deficiency risk predicted to increase under future climate change. Proc. Natl. Acad. Sci. USA 2017, 114, 2848–2853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baïz, N.; Chastang, J.; Ibanez, G.; Annesi-Maesano, I. Prenatal exposure to selenium may protect against wheezing in children by the age of 3. Immun. Inflamm. Dis. 2017, 5, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carneiro, M.F.; Rhoden, C.R.; Amantéa, S.L.; Barbosa, F. Low concentrations of selenium and zinc in nails are associated with childhood asthma. Biol. Trace Elem. Res. 2011, 144, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Kuti, B.P.; Kuti, D.K.; Smith, O.S. Serum Zinc, Selenium and Total Antioxidant Contents of Nigerian Children with Asthma: Association with Disease Severity and Symptoms Control. J. Trop. Pediatr. 2020, 66, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Zhang, X.; Hui, Y.; Xiang, H.; Wang, Q.; Xu, S.; Li, L. Sex Difference Between Trace Elements and Pulmonary Functions in Children. Biol. Trace Elem. Res. 2020, 197, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Leite, H.P.; Nogueira, P.C.; Iglesias, S.B.; de Oliveira, S.V.; Sarni, R.O. Increased plasma selenium is associated with better outcomes in children with systemic inflammation. Nutrition 2015, 31, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Picciano, M.F.; Wallig, M.A.; Milner, J.A. The role of selenium nutrition in the development of neonatal rat lung. Pediatr. Res. 1991, 29, 440–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burk, R.F.; Olson, G.E.; Hill, K.E.; Winfrey, V.P.; Motley, A.K.; Kurokawa, S. Maternal-fetal transfer of selenium in the mouse. FASEB J. 2013, 27, 3249–3256. [Google Scholar] [CrossRef] [Green Version]
- Papp, L.V.; Lu, J.; Holmgren, A.; Khanna, K.K. From selenium to selenoproteins: Synthesis, identity, and their role in human health. Antioxid. Redox Signal. 2007, 9, 775–806. [Google Scholar] [CrossRef] [Green Version]
- Schamberger, A.C.; Schiller, H.B.; Fernandez, I.E.; Sterclova, M.; Heinzelmann, K.; Hennen, E.; Hatz, R.; Behr, J.; Vašáková, M.; Mann, M.; et al. Glutathione peroxidase 3 localizes to the epithelial lining fluid and the extracellular matrix in interstitial lung disease. Sci. Rep. 2016, 6, 29952. [Google Scholar] [CrossRef]
- Yamada, Y.; Limmon, G.V.; Zheng, D.; Li, N.; Li, L.; Yin, L.; Chow, V.T.; Chen, J.; Engelward, B.P. Major shifts in the spatio-temporal distribution of lung antioxidant enzymes during influenza pneumonia. PLoS ONE 2012, 7, e31494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akahoshi, N.; Anan, Y.; Hashimoto, Y.; Tokoro, N.; Mizuno, R.; Hayashi, S.; Yamamoto, S.; Shimada, K.I.; Kamata, S.; Ishii, I. Dietary selenium deficiency or selenomethionine excess drastically alters organ selenium contents without altering the expression of most selenoproteins in mice. J. Nutr. Biochem. 2019, 69, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Berggren, M.M.; Mangin, J.F.; Gasdaka, J.R.; Powis, G. Effect of selenium on rat thioredoxin reductase activity: Increase by supranutritional selenium and decrease by selenium deficiency. Biochem. Pharmacol. 1999, 57, 187–193. [Google Scholar] [CrossRef]
- Bukalis, K.; Alber, D.; Bukalis, G.; Behne, D.; Kyriakopoulos, A. Effects of selenium diet on expression of selenoproteins in the lung of the rat. Ann. N. Y. Acad. Sci. 2007, 1095, 467–472. [Google Scholar] [CrossRef]
- Deneke, S.M.; Gershoff, S.N.; Fanburg, B.L. Changes in O2 toxicity and glutathione peroxidase levels in selenium deficient rats. Chest 1983, 83, 39S–40S. [Google Scholar] [CrossRef] [Green Version]
- Hawker, F.H.; Ward, H.E.; Stewart, P.M.; Wynne, L.A.; Snitch, P.J. Selenium deficiency augments the pulmonary toxic effects of oxygen exposure in the rat. Eur. Respir. J. 1993, 6, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Beck, M.A.; Nelson, H.K.; Shi, Q.; Van Dael, P.; Schiffrin, E.J.; Blum, S.; Barclay, D.; Levander, O.A. Selenium deficiency increases the pathology of an influenza virus infection. FASEB J. 2001, 15, 1481–1483. [Google Scholar] [CrossRef] [Green Version]
- Beck, M.A.; Handy, J.; Levander, O.A. Host nutritional status: The neglected virulence factor. Trends Microbiol. 2004, 12, 417–423. [Google Scholar] [CrossRef]
- Jaspers, I.; Zhang, W.; Brighton, L.E.; Carson, J.L.; Styblo, M.; Beck, M.A. Selenium deficiency alters epithelial cell morphology and responses to influenza. Free Radic. Biol. Med. 2007, 42, 1826–1837. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Sun, L.; Nan, Y.; Zhu, L.Y. Protection from H1N1 influenza virus infections in mice by supplementation with selenium: A comparison with selenium-deficient mice. Biol. Trace Elem. Res. 2011, 141, 254–261. [Google Scholar] [CrossRef]
- Glass, M.; Sutherland, M.W.; Forman, H.J.; Fisher, A.B. Selenium deficiency potentiates paraquat-induced lipid peroxidation in isolated perfused rat lung. J. Appl. Physiol. 1985, 59, 619–622. [Google Scholar] [CrossRef] [PubMed]
- Omaye, S.T.; Reddy, K.A.; Cross, C.E. Enhanced lung toxicity of paraquat in selenium-deficient rats. Toxicol. Appl. Pharmacol. 1978, 43, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Schomburg, L.; Schweizer, U. Hierarchical regulation of selenoprotein expression and sex-specific effects of selenium. Biochim. Biophys. Acta 2009, 1790, 1453–1462. [Google Scholar] [CrossRef] [PubMed]
- Coursin, D.B.; Cihla, H.P. Pulmonary effects of short term selenium deficiency. Thorax 1996, 51, 479–483. [Google Scholar] [CrossRef] [Green Version]
- Styblo, M.; Walton, F.S.; Harmon, A.W.; Sheridan, P.A.; Beck, M.A. Activation of superoxide dismutase in selenium-deficient mice infected with influenza virus. J. Trace Elem. Med. Biol. 2007, 21, 52–62. [Google Scholar] [CrossRef]
- Sherlock, L.G.; Balasubramaniyan, D.; Zheng, L.; Zarate, M.; Sizemore, T.; Delaney, C.; Tipple, T.E.; Wright, C.J.; Nozik-Grayck, E. Neonatal Selenoenzyme Expression Is Variably Susceptible to Duration of Maternal Selenium Deficiency. Antioxidants 2021, 10, 288. [Google Scholar] [CrossRef]
- Wolterink-Donselaar, I.G.; Meerding, J.M.; Fernandes, C. A method for gender determination in newborn dark pigmented mice. Lab Anim. 2009, 38, 35–38. [Google Scholar] [CrossRef]
- Delaney, C.; Wright, R.H.; Tang, J.R.; Woods, C.; Villegas, L.; Sherlock, L.; Savani, R.C.; Abman, S.H.; Nozik-Grayck, E. Lack of EC-SOD worsens alveolar and vascular development in a neonatal mouse model of bleomycin-induced bronchopulmonary dysplasia and pulmonary hypertension. Pediatr. Res. 2015, 78, 634–640. [Google Scholar] [CrossRef] [Green Version]
- Koeberle, S.C.; Gollowitzer, A.; Laoukili, J.; Kranenburg, O.; Werz, O.; Koeberle, A.; Kipp, A.P. Distinct and overlapping functions of glutathione peroxidases 1 and 2 in limiting NF-kappaB-driven inflammation through redox-active mechanisms. Redox Biol. 2020, 28, 101388. [Google Scholar] [CrossRef]
- Britt, R.D.; Velten, M.; Locy, M.L.; Rogers, L.K.; Tipple, T.E. The thioredoxin reductase-1 inhibitor aurothioglucose attenuates lung injury and improves survival in a murine model of acute respiratory distress syndrome. Antioxid. Redox Signal. 2014, 20, 2681–2691. [Google Scholar] [CrossRef]
- Dunigan, K.; Li, Q.; Li, R.; Locy, M.L.; Wall, S.; Tipple, T.E. The thioredoxin reductase inhibitor auranofin induces heme oxygenase-1 in lung epithelial cells via Nrf2-dependent mechanisms. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 315, L545–L552. [Google Scholar] [CrossRef] [PubMed]
- Dunigan-Russell, K.; Lin, V.; Silverberg, M.; Wall, S.B.; Li, R.; Gotham, J.; Nicola, T.; Sridharan, A.; Snowball, J.; Delaney, C.; et al. Aurothioglucose enhances proangiogenic pathway activation in lungs from room air and hyperoxia-exposed newborn mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 318, L1165–L1171. [Google Scholar] [CrossRef] [PubMed]
- Masumoto, K.; Nagata, K.; Higashi, M.; Nakatsuji, T.; Uesugi, T.; Takahashi, Y.; Nishimoto, Y.; Kitajima, J.; Hikino, S.; Hara, T.; et al. Clinical features of selenium deficiency in infants receiving long-term nutritional support. Nutrition 2007, 23, 782–787. [Google Scholar] [CrossRef]
- Crowley, G.; Kwon, S.; Caraher, E.J.; Haider, S.H.; Lam, R.; Batra, P.; Melles, D.; Liu, M.; Nolan, A. Quantitative lung morphology: Semi-automated measurement of mean linear intercept. BMC Pulm. Med. 2019, 19, 206. [Google Scholar] [CrossRef]
- Nguyen, L.; Castro, O.; De Dios, R.; Sandoval, J.; McKenna, S.; Wright, C.J. Sex-differences in LPS-induced neonatal lung injury. Sci. Rep. 2019, 9, 8514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooney, T.P.; Thurlbeck, W.M. The radial alveolar count method of Emery and Mithal: A reappraisal 2–intrauterine and early postnatal lung growth. Thorax 1982, 37, 580–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EMERY, J.L.; MITHAL, A. The number of alveoli in the terminal respiratory unit of man during late intrauterine life and childhood. Arch. Dis. Child. 1960, 35, 544–547. [Google Scholar] [CrossRef] [Green Version]
- Lingappan, K.; Jiang, W.; Wang, L.; Moorthy, B. Sex-specific differences in neonatal hyperoxic lung injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 311, L481–L493. [Google Scholar] [CrossRef] [Green Version]
- Hill, K.E.; McCollum, G.W.; Boeglin, M.E.; Burk, R.F. Thioredoxin reductase activity is decreased by selenium deficiency. Biochem. Biophys. Res. Commun. 1997, 234, 293–295. [Google Scholar] [CrossRef]
- Burk, R.F.; Olson, G.E.; Winfrey, V.P.; Hill, K.E.; Yin, D. Glutathione peroxidase-3 produced by the kidney binds to a population of basement membranes in the gastrointestinal tract and in other tissues. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G32–G38. [Google Scholar] [CrossRef]
- Conrad, M.; Jakupoglu, C.; Moreno, S.G.; Lippl, S.; Banjac, A.; Schneider, M.; Beck, H.; Hatzopoulos, A.K.; Just, U.; Sinowatz, F.; et al. Essential role for mitochondrial thioredoxin reductase in hematopoiesis, heart development, and heart function. Mol. Cell. Biol. 2004, 24, 9414–9423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakupoglu, C.; Przemeck, G.K.; Schneider, M.; Moreno, S.G.; Mayr, N.; Hatzopoulos, A.K.; de Angelis, M.H.; Wurst, W.; Bornkamm, G.W.; Brielmeier, M.; et al. Cytoplasmic thioredoxin reductase is essential for embryogenesis but dispensable for cardiac development. Mol. Cell. Biol. 2005, 25, 1980–1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostert, V.; Hill, K.E.; Burk, R.F. Loss of activity of the selenoenzyme thioredoxin reductase causes induction of hepatic heme oxygenase-1. FEBS Lett. 2003, 541, 85–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Wall, S.B.; Ren, C.; Velten, M.; Hill, C.L.; Locy, M.L.; Rogers, L.K.; Tipple, T.E. Thioredoxin Reductase Inhibition Attenuates Neonatal Hyperoxic Lung Injury and Enhances Nuclear Factor E2-Related Factor 2 Activation. Am. J. Respir. Cell Mol. Biol. 2016, 55, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Burk, R.F.; Hill, K.E.; Nakayama, A.; Mostert, V.; Levander, X.A.; Motley, A.K.; Johnson, D.A.; Johnson, J.A.; Freeman, M.L.; Austin, L.M. Selenium deficiency activates mouse liver Nrf2-ARE but vitamin E deficiency does not. Free Radic. Biol. Med. 2008, 44, 1617–1623. [Google Scholar] [CrossRef] [Green Version]
- Mostert, V.; Hill, K.E.; Ferris, C.D.; Burk, R.F. Selective induction of liver parenchymal cell heme oxygenase-1 in selenium-deficient rats. Biol. Chem. 2003, 384, 681–687. [Google Scholar] [CrossRef]
- Addinsall, A.B.; Wright, C.R.; Andrikopoulos, S.; van der Poel, C.; Stupka, N. Emerging roles of endoplasmic reticulum-resident selenoproteins in the regulation of cellular stress responses and the implications for metabolic disease. Biochem. J. 2018, 475, 1037–1057. [Google Scholar] [CrossRef]
- Moghadaszadeh, B.; Rider, B.E.; Lawlor, M.W.; Childers, M.K.; Grange, R.W.; Gupta, K.; Boukedes, S.S.; Owen, C.A.; Beggs, A.H. Selenoprotein N deficiency in mice is associated with abnormal lung development. FASEB J. 2013, 27, 1585–1599. [Google Scholar] [CrossRef] [Green Version]
- Darlow, B.A.; Inder, T.E.; Graham, P.J.; Sluis, K.B.; Malpas, T.J.; Taylor, B.J.; Winterbourn, C.C. The relationship of selenium status to respiratory outcome in the very low birth weight infant. Pediatrics 1995, 96, 314–319. [Google Scholar] [CrossRef]
- Acker, S.N.; Seedorf, G.J.; Abman, S.H.; Nozik-Grayck, E.; Kuhn, K.; Partrick, D.A.; Gien, J. Altered pulmonary artery endothelial-smooth muscle cell interactions in experimental congenital diaphragmatic hernia. Pediatr. Res. 2015, 77, 511–519. [Google Scholar] [CrossRef]
- Ewan, R.C. Effect of selenium on rat growth, growth hormone and diet utilization. J. Nutr. 1976, 106, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Hofstee, P.; Bartho, L.A.; McKeating, D.R.; Radenkovic, F.; McEnroe, G.; Fisher, J.J.; Holland, O.J.; Vanderlelie, J.J.; Perkins, A.V.; Cuffe, J.S.M. Maternal selenium deficiency during pregnancy in mice increases thyroid hormone concentrations, alters placental function and reduces fetal growth. J. Physiol. 2019, 597, 5597–5617. [Google Scholar] [CrossRef] [PubMed]
- Mistry, H.D.; Kurlak, L.O.; Young, S.D.; Briley, A.L.; Pipkin, F.B.; Baker, P.N.; Poston, L. Maternal selenium, copper and zinc concentrations in pregnancy associated with small-for-gestational-age infants. Matern. Child Nutr. 2014, 10, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Nogales, F.; Ojeda, M.L.; Fenutría, M.; Murillo, M.L.; Carreras, O. Role of selenium and glutathione peroxidase on development, growth, and oxidative balance in rat offspring. Reproduction 2013, 146, 659–667. [Google Scholar] [CrossRef] [Green Version]
- Biniwale, M.A.; Ehrenkranz, R.A. The role of nutrition in the prevention and management of bronchopulmonary dysplasia. Semin. Perinatol. 2006, 30, 200–208. [Google Scholar] [CrossRef]
- Arigliani, M.; Spinelli, A.M.; Liguoro, I.; Cogo, P. Nutrition and Lung Growth. Nutrients 2018, 10, 919. [Google Scholar] [CrossRef] [Green Version]
- Aydin, K.; Bideci, A.; Kendirci, M.; Cinaz, P.; Kurtoglu, S. Insulin-like growth factor-I and insulin-like growth factor binding protein-3 levels of children living in an iodine- and selenium-deficient endemic goiter area. Biol. Trace Elem. Res. 2002, 90, 25–30. [Google Scholar] [CrossRef]
- Narayan, V.; Ravindra, K.C.; Liao, C.; Kaushal, N.; Carlson, B.A.; Prabhu, K.S. Epigenetic regulation of inflammatory gene expression in macrophages by selenium. J. Nutr. Biochem. 2015, 26, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Weitzel, F.; Ursini, F.; Wendel, A. Phospholipid hydroperoxide glutathione peroxidase in various mouse organs during selenium deficiency and repletion. Biochim. Biophys. Acta 1990, 1036, 88–94. [Google Scholar] [CrossRef]
- Tindell, R.; Wall, S.B.; Li, Q.; Li, R.; Dunigan, K.; Wood, R.; Tipple, T.E. Selenium supplementation of lung epithelial cells enhances nuclear factor E2-related factor 2 (Nrf2) activation following thioredoxin reductase inhibition. Redox Biol. 2018, 19, 331–338. [Google Scholar] [CrossRef]
- Staples, S.; Wall, S.B.; Li, R.; Tipple, T.E. Selenium-independent antioxidant and anti-inflammatory effects of thioredoxin reductase inhibition in alveolar macrophages. Life Sci. 2020, 259, 118285. [Google Scholar] [CrossRef] [PubMed]
- Wall, S.B.; Li, R.; Butler, B.; Burg, A.R.; Tse, H.M.; Larson-Casey, J.L.; Carter, A.B.; Wright, C.J.; Rogers, L.K.; Tipple, T.E. Auranofin-Mediated NRF2 Induction Attenuates Interleukin 1 Beta Expression in Alveolar Macrophages. Antioxidants 2021, 10, 632. [Google Scholar] [CrossRef] [PubMed]
- Wall, S.B.; Wood, R.; Dunigan, K.; Li, Q.; Li, R.; Rogers, L.K.; Tipple, T.E. Thioredoxin Reductase-1 Inhibition Augments Endogenous Glutathione-Dependent Antioxidant Responses in Experimental Bronchopulmonary Dysplasia. Oxid. Med. Cell. Longev. 2019, 2019, 7945983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suvorova, E.S.; Lucas, O.; Weisend, C.M.; Rollins, M.F.; Merrill, G.F.; Capecchi, M.R.; Schmidt, E.E. Cytoprotective Nrf2 pathway is induced in chronically txnrd 1-deficient hepatocytes. PLoS ONE 2009, 4, e6158. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Li, R.; Wall, S.B.; Dunigan, K.; Ren, C.; Jilling, T.; Rogers, L.K.; Tipple, T.E. Aurothioglucose does not improve alveolarization or elicit sustained Nrf2 activation in C57BL/6 models of bronchopulmonary dysplasia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 314, L736–L742. [Google Scholar] [CrossRef] [Green Version]
- Petit, N.; Lescure, A.; Rederstorff, M.; Krol, A.; Moghadaszadeh, B.; Wewer, U.M.; Guicheney, P. Selenoprotein N: An endoplasmic reticulum glycoprotein with an early developmental expression pattern. Hum. Mol. Genet. 2003, 12, 1045–1053. [Google Scholar] [CrossRef] [Green Version]
- LungMap. Available online: www.lungmap.net (accessed on 19 September 2022).
- Tindell, R.; Tipple, T. Selenium: Implications for outcomes in extremely preterm infants. J. Perinatol. 2018, 38, 197–202. [Google Scholar] [CrossRef]
- Lockitch, G.; Jacobson, B.; Quigley, G.; Dison, P.; Pendray, M. Selenium deficiency in low birth weight neonates: An unrecognized problem. J. Pediatr. 1989, 114, 865–870. [Google Scholar] [CrossRef]
- Atli, M.; Erikoglu, M.; Kaynak, A.; Esen, H.H.; Kurban, S. The effects of selenium and vitamin E on lung tissue in rats with sepsis. Clin. Investig. Med. 2012, 35, E48–E54. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Yang, G.; Guan, G.; Zhang, Y.; Ren, W.; Yin, J.; Aguilar, Y.M.; Luo, W.; Fang, J.; Yu, X.; et al. Effect of dietary selenium yeast supplementation on porcine circovirus type 2 (PCV2) infections in mice. PLoS ONE 2015, 10, e0115833. [Google Scholar] [CrossRef]
- Amini, P.; Kolivand, S.; Saffar, H.; Rezapoor, S.; Motevaseli, E.; Najafi, M.; Nouruzi, F.; Shabeeb, D.; Musa, A.E. Protective Effect of Selenium-L-methionine on Radiation-induced Acute Pneumonitis and Lung Fibrosis in Rat. Curr. Clin. Pharmacol. 2019, 14, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Fawaz, Y.B.; Matta, J.M.; Moustafa, M.E. Effects of selenium supplementation on lung oxidative stress after exposure to exhaust emissions from pyrolysis oil, biodiesel and diesel. Toxicol. Mech. Methods 2019, 29, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.A.; Eltahawy, N.F.; Abdalla, A.M.; Khalaf, H.M. Protective effects of selenium in tacrolimus-induced lung toxicity: Potential role of heme oxygenase 1. Can. J. Physiol. Pharmacol. 2021, 99, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Suh, G.J.; Kwon, W.Y.; Kwak, Y.H.; Lee, K.; Lee, H.J.; Jeong, K.Y.; Lee, M.W. Antioxidant effects of selenium on lung injury in paraquat intoxicated rats. Clin. Toxicol. 2012, 50, 749–753. [Google Scholar] [CrossRef] [PubMed]
- Ghorbel, I.; Elwej, A.; Chaabane, M.; Jamoussi, K.; Mnif, H.; Boudawara, T.; Zeghal, N. Selenium Alleviates Oxidative Stress and Lung Damage Induced by Aluminum Chloride in Adult Rats: Biochemical and Histological Approach. Biol. Trace Elem. Res. 2017, 176, 181–191. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sherlock, L.G.; McCarthy, W.C.; Grayck, M.R.; Solar, M.; Hernandez, A.; Zheng, L.; Delaney, C.; Tipple, T.E.; Wright, C.J.; Nozik, E.S. Neonatal Selenium Deficiency Decreases Selenoproteins in the Lung and Impairs Pulmonary Alveolar Development. Antioxidants 2022, 11, 2417. https://doi.org/10.3390/antiox11122417
Sherlock LG, McCarthy WC, Grayck MR, Solar M, Hernandez A, Zheng L, Delaney C, Tipple TE, Wright CJ, Nozik ES. Neonatal Selenium Deficiency Decreases Selenoproteins in the Lung and Impairs Pulmonary Alveolar Development. Antioxidants. 2022; 11(12):2417. https://doi.org/10.3390/antiox11122417
Chicago/Turabian StyleSherlock, Laura G., William C. McCarthy, Maya R. Grayck, Mack Solar, Andres Hernandez, Lijun Zheng, Cassidy Delaney, Trent E. Tipple, Clyde J. Wright, and Eva S. Nozik. 2022. "Neonatal Selenium Deficiency Decreases Selenoproteins in the Lung and Impairs Pulmonary Alveolar Development" Antioxidants 11, no. 12: 2417. https://doi.org/10.3390/antiox11122417