
Hexavalent-Chromium-Induced Oxidative Stress and the Protective Role of Antioxidants against Cellular Toxicity
 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
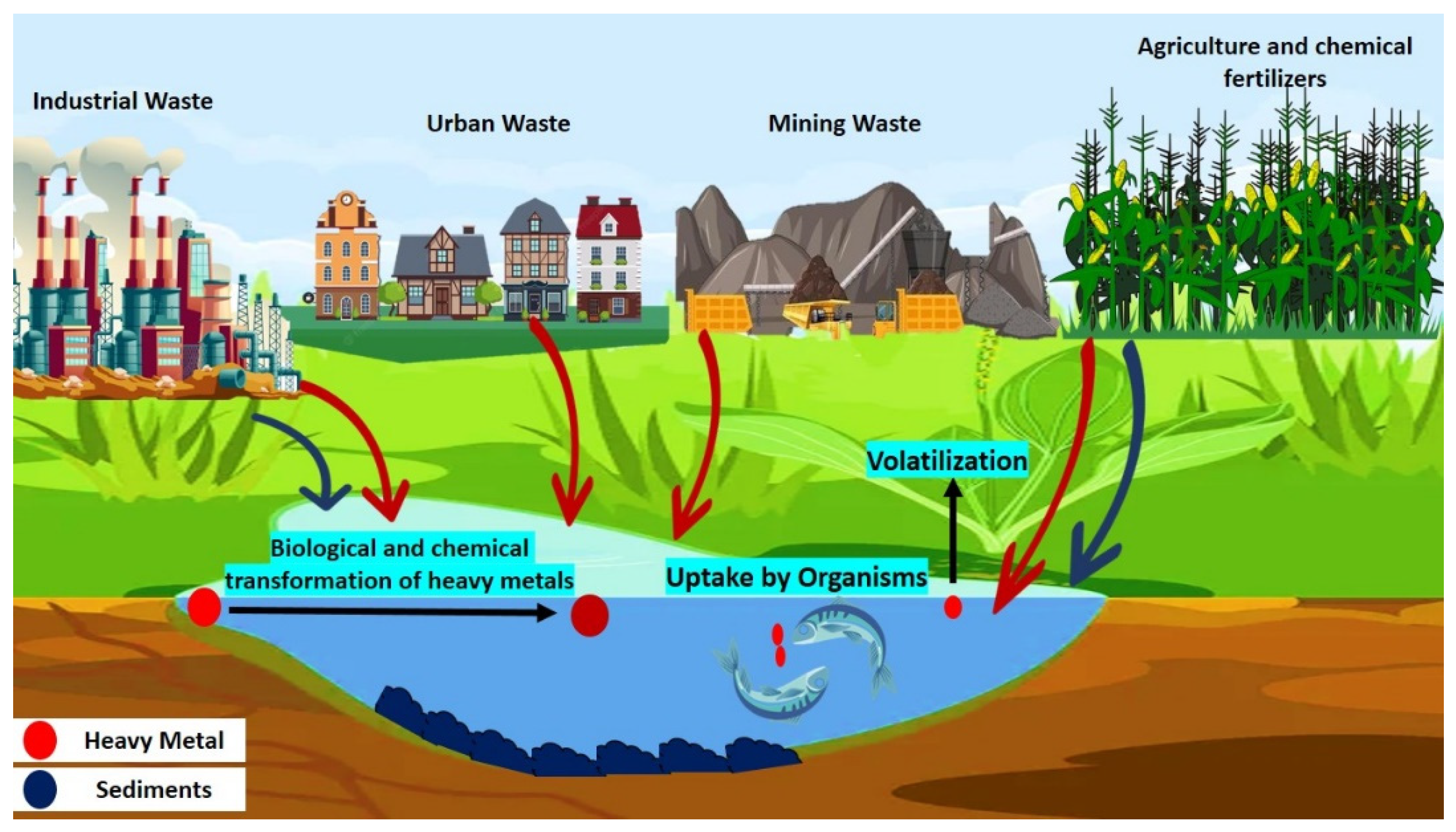
2. Source of Cr (VI) Contamination
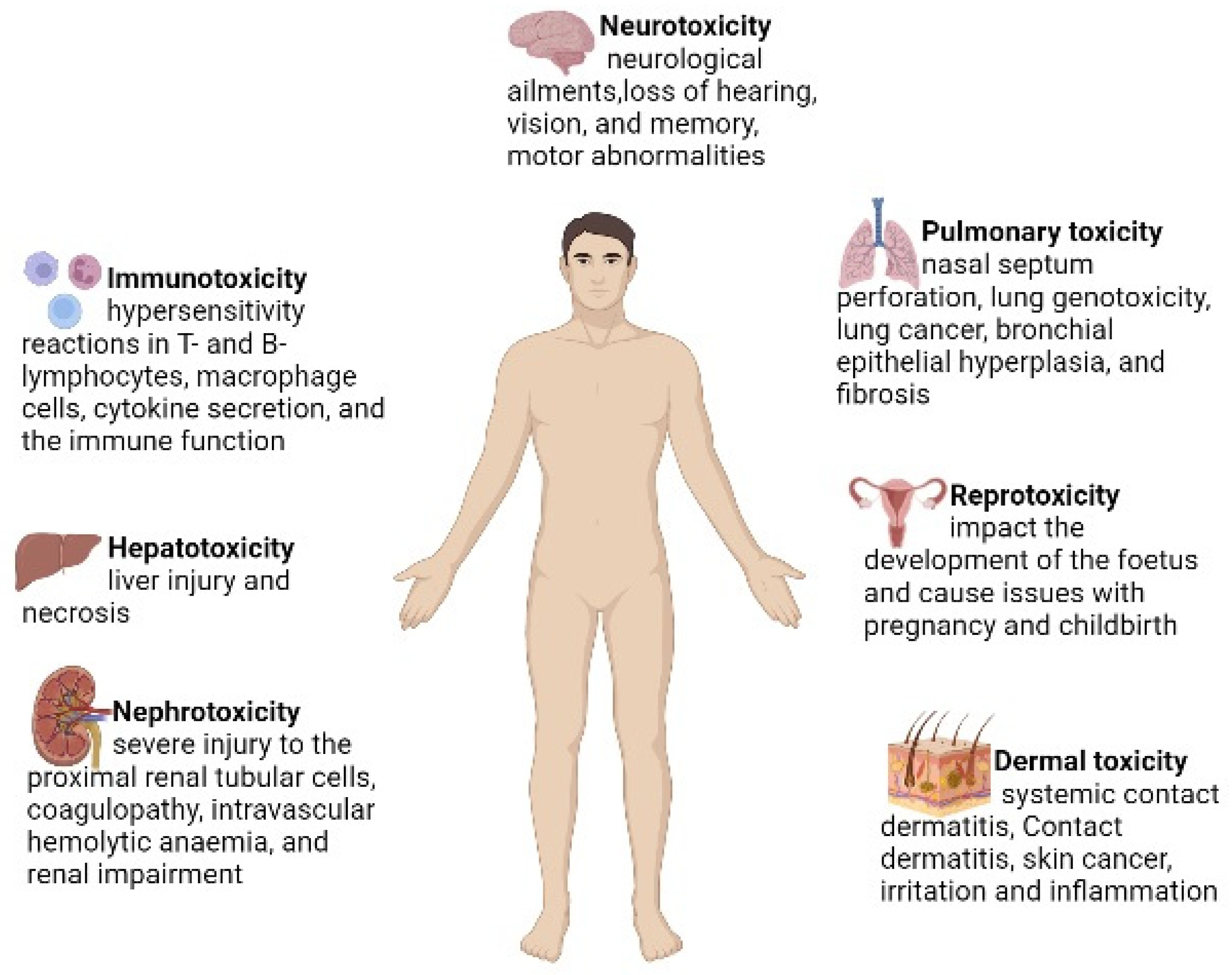
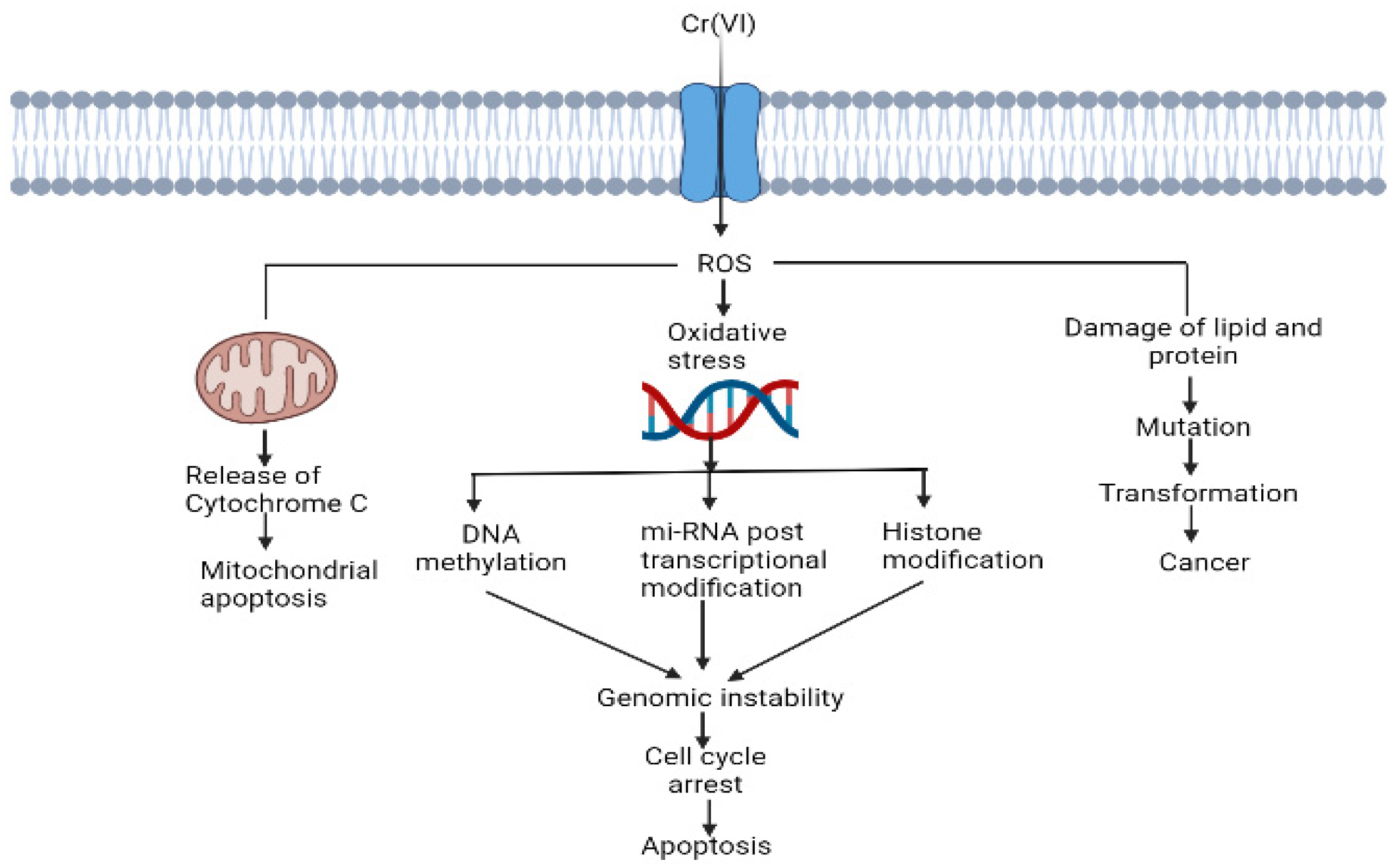
3. Cr (VI)-Induced Oxidative Stress and Disruption of Cell Components
4. Antioxidants and Their Protective Role against Cr (VI) Toxicity
4.1. Enzymatic Antioxidants
4.1.1. Superoxide Dismutase (EC 1.15.1.1)
4.1.2. Catalase (EC 1.11.1.6)
4.1.3. Glutathione Peroxidase (EC 1.11.1.19)
4.2. Endogenous and Exogenous Antioxidants
4.3. Polyphenolic Antioxidants
5. Mechanism and Mode of Action of Antioxidants
- (1)
- React with ROS and decrease their level in the cell. SOD, catalase and peroxidase are major antioxidants considered in the category.
- (2)
- Glutathione reductase (GR) is one of the well-known antioxidants of the group. This type of antioxidants restores the reduced forms of non-enzymatic antioxidants (GSH) [87].
6. Antioxidant Measurement Methods
6.1. HAT-Based Assays
6.2. ET-Based Assays
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wong, M.H. Environmental Contamination: Health Risks and Ecological Restoration; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar] [CrossRef]
- Fergusson, J.E. The Heavy Elements: Chemistry, Environmental Impact and Health Effects; Pergamon Press: Oxford, UK, 1990; pp. 85–547. [Google Scholar]
- Wu, M.; Xu, Y.; Ding, W.; Li, Y.; Xu, H. Mycoremediation of manganese and phenanthrene by Pleurotus eryngii mycelium enhanced by Tween 80 and saponin. Appl. Microbiol. Biotechnol. 2016, 100, 7249–7261. [Google Scholar] [CrossRef] [PubMed]
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy metal pollution in the environment and their toxicological effects on humans. Heliyon 2020, 6, e04691. [Google Scholar] [CrossRef] [PubMed]
- Flora, S.J.; Pachauri, V. Chelation in metal intoxication. Int. J. Environ. Res. Public Health 2010, 7, 2745–2788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobhanardakani, S.; Zandi Pak, R.; Mohammadi, M.J. Removal of Ni (II) and Zn (II) from aqueous solutions using chitosan. Arch. Hyg. Sci. 2016, 5, 47–55. [Google Scholar]
- Barakat, M.A. New trends in removing heavy metals from industrial wastewater. Arab. J. Chem. 2011, 4, 361–377. [Google Scholar] [CrossRef] [Green Version]
- Rai, P.K. Heavy metal pollution in aquatic ecosystems and its phytoremediation using wetland plants: An ecosustainable approach. Int. J. Phytorem. 2008, 10, 131–158. [Google Scholar] [CrossRef] [PubMed]
- Ghose, M.K. Design of cost-effective coal washery effluent treatment plant for clean environment. J. Sci. Ind. Res. 2001, 60, 40–47. [Google Scholar]
- ATSDR; Public Health Service. Toxicological Profile for Lead; ATSDR: Atlanta, GA, USA, 2020. Available online: https://www.atsdr.cdc.gov/toxprofiles/tp13.pdf (accessed on 1 March 2020).
- Owlad, M.; Aroua, M.K.; Daud, W.A.W.; Baroutian, S. Removal of hexavalent chromium-contaminated water and wastewater: A review. Water Air Soil Pollut. 2009, 200, 59–70. [Google Scholar] [CrossRef]
- Central Pollution Control Board (CPCB). India (2000) Environmental Standards for Ambient Air, Automobiles, Fuels, Industries and Noise. Available online: http://www.cpcbenvis.nic.in/scanned%20reports/PCL%204%20Environmental%20Standards.pdf (accessed on 15 March 2018).
- Peterson-Roth, E.; Reynolds, M.; Quievryn, G.; Zhitkovich, A. Mismatch repair proteins are activators of toxic responses to chromium-DNA damage. Mol. Cell. Biol. 2005, 25, 3596–3607. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Singh, M.P.; Mishra, V. Bioremediation of toxic metal ions from coal washery effluent. Desalin. Water Treat. 2020, 197, 300–318. [Google Scholar] [CrossRef]
- Singh, V.; Singh, J.; Mishra, V. Development of a cost-effective, recyclable and viable metal ion doped adsorbent for simultaneous adsorption and reduction of toxic Cr (VI) ions. J. Environ. Chem. Eng. 2021, 9, 105124–105137. [Google Scholar] [CrossRef]
- Ahemad, M. Enhancing phytoremediation of chromium-stressed soils through plant-growth-promoting bacteria. J. Genet. Eng. Biotechnol. 2015, 13, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubrak, O.I.; Lushchak, V.; Lushchak, J.V.; Torous, I.M.; Storey, J.M.; Storey, K.B.; Lushchak, V.I. Chromium effects on free radical processes in goldfish tissues: Comparison of Cr (III) and Cr (VI) exposures on oxidative stress markers, glutathione status and antioxidant enzymes. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 152, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.S.; Praveenkumar, R.; Ilavarasi, A.; Rajeshwari, K.; Thajuddin, N. Biochemical changes of fresh water cyanobacteria Dolichospermum flos-aquae NTMS07 to chromium-induced stress with special reference to antioxidant enzymes and cellular fatty acids. Bull. Environ. Contam. Toxicol. 2013, 90, 730–735. [Google Scholar] [CrossRef]
- Zafar, S.; Aqil, F.; Ahmad, I. Metal tolerance and biosorption potential of filamentous fungi isolated from metal contaminated agricultural soil. Bioresour. Technol. 2007, 98, 2557–2561. [Google Scholar] [CrossRef]
- Joutey, N.T.; Sayel, H.; Bahafid, W.; Ghachtouli, N. Mechanisms of hexavalent chromium resistance and removal by microorganisms. Rev. Environ. Contam. Toxicol. 2015, 233, 45–69. [Google Scholar]
- Qian, J.; Zhou, J.; Wang, L.; Wei, L.; Li, Q.; Wang, D.; Wang, Q. Direct Cr (VI) Bio-Reduction with Organics as Electron Donor by Anaerobic Sludge. Chem. Eng. J. 2017, 309, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Tian, Y. Hexavalent Chromium Reducing Bacteria: Mechanism of Reduction and Characteristics. Environ. Sci. Pollut. Res. 2021, 28, 20981–20997. [Google Scholar] [CrossRef]
- Xia, S.; Song, Z.; Jeyakumar, P.; Shaheen, S.M.; Rinklebe, J.; Ok, Y.S.; Bolan, N.; Wang, H. A Critical Review on Bioremediation Technologies for Cr(VI)-Contaminated Soils and Wastewater. Crit. Rev. Environ. Sci. Technol. 2019, 49, 1027–1078. [Google Scholar] [CrossRef]
- Kapahi, M.; Sachdeva, S. Mycoremediation Potential of Pleurotus Species for Heavy Metals: A Review. Bioresour. Bioprocess. 2017, 4, 32. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Yu, X. Hexavalent Chromium in Drinking Water: Chemistry, Challenges and Future Outlook on Sn(II)- and Photocatalyst-Based Treatment. Front. Environ. Sci. Eng. 2020, 14, 88. [Google Scholar] [CrossRef] [PubMed]
- Bokare, A.D.; Choi, W. Advanced oxidation process based on the Cr (III)/Cr (VI) redox cycle. Environ. Sci. Technol. 2011, 45, 9332–9338. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Bharagava, R.N. Toxic and Genotoxic Effects of Hexavalent Chromium in Environment and Its Bioremediation Strategies. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2016, 34, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Abdel-Moneim, A.-M.E.; Yang, B. Signaling Pathways and Genes Associated with Hexavalent Chromium-Induced Hepatotoxicity. Biol. Trace Elem. Res. 2011, 225, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, R.; Renu, K.; Eladl, M.A.; El-Sherbiny, M.; Elsherbini, D.M.A.; Mirza, A.K.; Vellingiri, B.; Iyer, M.; Dey, A.; Gopalakrishnan, A.V. Mechanism of Chromium-Induced Toxicity in Lungs, Liver, and Kidney and Their Ameliorative Agents. Biomed. Pharmacother. 2022, 151, 113119. [Google Scholar] [CrossRef]
- Kim, H.G.; Yoon, D.H.; Lee, W.H.; Han, S.K.; Shrestha, B.; Kim, C.H.; Lim, M.H.; Chang, W.; Lim, S.; Choi, S.; et al. Phellinus linteus inhibits inflammatory mediators by suppressing redox-based NF-κB and MAPKs activation in lipopolysaccharide-induced RAW 264.7 macrophage. J. Ethnopharmacol. 2007, 114, 307–315. [Google Scholar] [CrossRef]
- Bagchi, D.; Vuchetich, P.J.; Bagchi, M.; Hassoun, E.A.; Tran, M.X.; Tang, L.; Stohs, S.J. Induction of oxidative stress by chronic administration of sodium dichromate [chromium VI] and cadmium chloride [cadmium II] to rats. Free Radical Biol. Med. 1997, 22, 471–478. [Google Scholar] [CrossRef]
- Wise, S.S.; Holmes, A.L.; Ketterer, M.E.; Hartsock, W.J.; Fomchenko, E.; Katsifis, S.; Thompson, W.D.; Wise, J.P., Sr. Chromium is the proximate clastogenic species for lead chromate-induced clastogenicity in human bronchial cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2004, 560, 79–89. [Google Scholar] [CrossRef]
- Wakeel, A.; Xu, M.; Gan, Y. Chromium-induced reactive oxygen species accumulation by altering the enzymatic antioxidant system and associated cytotoxic, genotoxic, ultrastructural, and photosynthetic changes in plants. Int. J. Mol. Sci. 2020, 21, 728. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.D.; Kargacin, B.; Klein, C.B.; Costa, M. Mechanisms of chromium carcinogenicity and toxicity. Crit. Rev. Toxicol. 1993, 23, 255–281. [Google Scholar] [CrossRef]
- Balali-Mood, M.; Naseri, K.; Tahergorabi, Z.; Khazdair, M.R.; Sadeghi, M. Toxic Mechanisms of Five Heavy Metals: Mercury, Lead, Chromium, Cadmium, and Arsenic. Front. Pharmacol. 2021, 12, 643972. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.A.; El-Houseiny, W.; El-Murr, A.E.; Ebraheim, L.L.M.; Ahmed, A.I.; El-Hakim, Y.M.A. Effect of Hexavalent Chromium Exposure on the Liver and Kidney Tissues Related to the Expression of CYP450 and GST Genes of Oreochromis Niloticus Fish: Role of Curcumin Supplemented Diet. Ecotoxicol. Environ. Saf. 2020, 188, 109890. [Google Scholar] [CrossRef]
- Li, S.; Baiyun, R.; Lv, Z.; Li, J.; Han, D.; Zhao, W.; Yu, L.; Deng, N.; Liu, Z.; Zhang, Z. Exploring the Kidney Hazard of Exposure to Mercuric Chloride in Mice: Disorder of Mitochondrial Dynamics Induces Oxidative Stress and Results in Apoptosis. Chemosphere 2019, 234, 822–829. [Google Scholar] [CrossRef]
- Li, H.; Shi, J.; Gao, H.; Yang, X.; Fu, Y.; Peng, Y.; Xia, Y.; Zhou, D. Hexavalent Chromium Causes Apoptosis and Autophagy by Inducing Mitochondrial Dysfunction and Oxidative Stress in Broiler Cardiomyocytes. Biol. Trace Elem. Res. 2022, 200, 2866–2875. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.; Mondal, P.; Dey Bhowmik, A.; Bandyopadhyay, A.; Sudarshan, M.; Chakraborty, A.; Chattopadhyay, A. Environmentally Relevant Hexavalent Chromium Disrupts Elemental Homeostasis and Induces Apoptosis in Zebrafish Liver. Bull. Environ. Contam. Toxicol. 2022, 108, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Patlolla, A.K.; Barnes, C.; Yedjou, C.; Velma, V.R.; Tchounwou, P.B. Oxidative Stress, DNA Damage, and Antioxidant Enzyme Activity Induced by Hexavalent Chromium in Sprague-Dawley Rats. Environ Toxicol. 2009, 24, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Das, J.; Sarkar, A.; Sil, P.C. Hexavalent Chromium Induces Apoptosis in Human Liver (HepG2) Cells via Redox Imbalance. Toxicol. Rep. 2015, 2, 600–608. [Google Scholar] [CrossRef] [Green Version]
- Lei, T.; He, Q.Y.; Cai, Z.; Zhou, Y.; Wang, Y.L.; Si, L.S.; Cai, Z.; Chiu, J.F. Proteomic Analysis of Chromium Cytotoxicity in Cultured Rat Lung Epithelial Cells. Proteomics 2008, 8, 2420–2429. [Google Scholar] [CrossRef]
- Velma, V.; Tchounwou, P.B. Chromium-Induced Biochemical, Genotoxic and Histopathologic Effects in Liver and Kidney of Goldfish, Carassius Auratus. Mutat. Res. 2010, 698, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Son, Y.O.; Chang, Q.; Sun, L.; Hitron, J.A.; Budhraja, A.; Zhang, Z.; Ke, Z.; Chen, F.; Luo, J.; et al. NADPH Oxidase Activation Is Required in Reactive Oxygen Species Generation and Cell Transformation Induced by Hexavalent Chromium. Toxicol. Sci. 2011, 123, 399–410. [Google Scholar] [CrossRef]
- Bagchi, D.; Stohs, S.J.; Downs, B.W.; Bagchi, M.; Preuss, H.G. Cytotoxicity and Oxidative Mechanisms of Different Forms of Chromium. Toxicology 2002, 180, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Maria, V.L.; Oliveira, M.; Pacheco, M.; Santos, M.A. Oxidative stress and genotoxic effects in gill and kidney of Anguilla anguilla L. exposed to chromium with or without pre-exposure to beta-naphthoflavone. Mutat. Res. 2006, 608, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Li, S.; Li, J.; Lv, Y.; Wang, X.; Wu, P.; Yang, Q.; Tang, Y.; Liu, Y.; Zhang, Z. Hexavalent Chromium Induces Renal Apoptosis and Autophagy via Disordering the Balance of Mitochondrial Dynamics in Rats. Ecotoxicol. Environ. Saf. 2020, 204, 111061. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, M.; Pei, L.; Zhang, R.; Liu, X.; Wei, L.; Yang, M.; Xu, Q. Oxidative stress and DNA damage in a long-term hexavalent chromium-exposed population in North China: A cross-sectional study. BMJ Open 2018, 8, e021470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavesi, T.; Moreira, J.C. Mechanisms and Individuality in Chromium Toxicity in Humans. J. Appl. Toxicol. 2020, 40, 1183–1197. [Google Scholar] [CrossRef]
- Bagchi, D.; Bagchi, M.; Stohs, S.J. Chromium (VI)-induced oxidative stress, apoptotic cell death and modulation of p53 tumor suppressor gene. Mol. Cell. Biochem. 2001, 222, 149–158. [Google Scholar] [CrossRef]
- Holmes, A.L.; Wise, S.S.; Wise, J.P., Sr. Carcinogenicity of hexavalent chromium. Indian J. Med. Res. 2008, 128, 353–372. [Google Scholar]
- Son, Y.O.; Hitron, J.A.; Wang, X.; Chang, Q.; Pan, J.; Zhang, Z.; Liu, J.; Wang, S.; Lee, J.C.; Shi, X. Cr(VI) induces mitochondrial-mediated and caspase-dependent apoptosis through reactive oxygen species-mediated p53 activation in JB6 Cl41 cells. Toxicol. Appl. Pharmacol. 2010, 245, 226–235. [Google Scholar] [CrossRef] [Green Version]
- Monga, A.; Fulke, A.B.; Dasgupta, D. Recent Developments in Essentiality of Trivalent Chromium and Toxicity of Hexavalent Chromium: Implications on Human Health and Remediation Strategies. J. Hazard. Mater. Adv. 2022, 7, 100113. [Google Scholar] [CrossRef]
- Islam, M.N.; Rauf, A.; Fahad, F.I.; Emran, T.B.; Mitra, S.; Olatunde, A.; Shariati, M.A.; Rebezov, M.; Rengasamy, K.R.R.; Mubarak, M.S. Superoxide dismutase: An updated review on its health benefits and industrial applications. Crit. Rev. Food Sci. Nutr. 2022, 62, 7282–7300. [Google Scholar] [CrossRef]
- Aziz, M.A.; Diab, A.S.; Mohammed, A.A. Antioxidant Categories and Mode of Action; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Matés, J.M. Effects of antioxidant enzymes in the molecular control of reactive oxygen species toxicology. Toxicology 2000, 153, 83–104. [Google Scholar] [CrossRef] [PubMed]
- Dworzański, J.; Strycharz-Dudziak, M.; Kliszczewska, E.; Kiełczykowska, M.; Dworzańska, A.; Drop, B.; Polz-Dacewicz, M. Glutathione peroxidase (GPx) and superoxide dismutase (SOD) activity in patients with diabetes mellitus type 2 infected with Epstein-Barr virus. PLoS ONE 2020, 15, e0230374. [Google Scholar] [CrossRef] [PubMed]
- Azadmanesh, J.; Borgstahl, G.E.O. A review of the catalytic mechanism of human manganese superoxide dismutase. Antioxidants 2018, 7, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewandowski, L.; Kepinska, M.; Milnerowicz, H. The copper-zinc superoxide dismutase activity in selected diseases. Eur. J. Clin. Investig. 2019, 49, e13036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yulyana, Y.; Tovmasyan, A.; Ho, I.A.; Sia, K.C.; Newman, J.P.; Ng, W.H.; Guo, C.M.; Hui, K.M.; Batinic-Haberle, I.; Lam, P.Y. Redox-Active Mn Porphyrin-based Potent SOD Mimic, MnTnBuOE-2-PyP5+, Enhances Carbenoxolone-Mediated TRAIL-Induced Apoptosis in Glioblastoma Multiforme. Stem Cell Rev. Rep. 2016, 12, 140–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soh, J.W.; Hotic, S.; Arking, R. Dietary restriction in Drosophila is dependent on mitochondrial efficiency and constrained by pre-existing extended longevity. Mech. Ageing Dev. 2007, 128, 581–593. [Google Scholar] [CrossRef]
- Rottner, M.; Tual-Chalot, S.; Mostefai, H.A.; Andriantsitohaina, R.; Freyssinet, J.M.; Martinez, M.C. Increased oxidative stress induces apoptosis in human cystic fibrosis cells. PLoS ONE 2011, 6, e24880. [Google Scholar] [CrossRef] [Green Version]
- Di Naso, F.C.; Dias, A.S.; Porawski, M.; Marroni, N.A.P. Exogenous superoxide dismutase: Action on liver oxidative stress in animals with streptozotocin-induced diabetes. Exp. Diabetes Res. 2011, 2011, 754132. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.W.; Shen, C.J.; Tung, Y.T.; Chen, H.L.; Chen, Y.H.; Chang, W.H.; Cheng, K.C.; Yang, S.H.; Chen, C.M. Extracellular superoxide dismutase ameliorates streptozotocin-induced rat diabetic nephropathy via inhibiting the ROS/ERK1/2 signaling. Life Sci. 2015, 135, 77–86. [Google Scholar] [CrossRef]
- Masih, S.A.M.A.; Devasahayam, M. Developments in Microbial Fuel Cell System for Electricity Generation. Trends. Biosci. 2014, 6, 701–704. [Google Scholar]
- Stančić, A.; Otašević, V.; Janković, A.; Vučetić, M.; Ivanović-Burmazović, I.; Filipović, M.R.; Korać, A.; Markelić, M.; Veličković, K.; Golić, I. Molecular basis of hippocampal energy metabolism in diabetic rats: The effects of SOD mimic. Brain Res. Bull. 2013, 99, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Balendra, V.; Singh, S.K. Therapeutic potential of astaxanthin and superoxide dismutase in Alzheimer’s disease. Open Biol. 2021, 11, 210013. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Rees, H.D.; Weintraub, S.T.; Levey, A.I.; Chin, L.S.; Li, L. Oxidative modifications and aggregation of Cu, Zn-superoxide dismutase associated with Alzheimer and Parkinson diseases. J. Biol. Chem. 2005, 280, 11648–11655. [Google Scholar] [CrossRef] [PubMed]
- Premkumar, B. Antioxidant defense and disease activity in rheumatoid arthritis. Res. J. Pharm. Technol. 2018, 11, 1810–1814. [Google Scholar] [CrossRef]
- Tehrani, S.H.; Moosavi-Movahedi, A.A. Catalase and its mysteries. Prog. Biophys. Mol. Biol. 2018, 140, 5–12. [Google Scholar] [CrossRef]
- Hunt, C.R.; Sim, J.E.; Sullivan, S.J.; Featherstone, T.; Golden, W.; Von Kapp-Herr, C.; Hock, R.A.; Gomez, R.A.; Parsian, A.J.; Spitz, D.R. Genomic instability and catalase gene amplification induced by chronic exposure to oxidative stress. Cancer Res. 1998, 58, 3986–3992. [Google Scholar]
- Flohé, L.; Toppo, S.; Orian, L. The glutathione peroxidase family: Discoveries and mechanism. Free Radic. Biol. Med. 2022, 187, 113–122. [Google Scholar] [CrossRef]
- Cubas-Gaona, L.L.; de Francisco, P.; Martín-González, A.; Gutiérrez, J.C. Tetrahymena glutathione peroxidase family: A comparative analysis of these antioxidant enzymes and differential gene expression to metals and oxidizing agents. Microorganisms 2020, 8, 1008. [Google Scholar] [CrossRef]
- Mirończuk-Chodakowska, I.; Witkowska, A.M.; Zujko, M.E. Endogenous Non-Enzymatic Antioxidants in the Human Body. Adv. Med. Sci. 2018, 63, 68–78. [Google Scholar] [CrossRef]
- Bouayed, J.; Bohn, T. Exogenous Antioxidants—Double-Edged Swords in Cellular Redox State: Health Beneficial Effects at Physiologic Doses versus Deleterious Effects at High Doses. Oxid. Med. Cell. Longev. 2010, 3, 228–237. [Google Scholar] [CrossRef] [Green Version]
- Husain, N.; Mahmood, R. 3,4-Dihydroxybenzaldehyde Quenches ROS and RNS and Protects Human Blood Cells from Cr(VI)-Induced Cytotoxicity and Genotoxicity. Toxicol. Vitr. 2018, 50, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.B.; Chul Hong, S.; Jin Jeong, H. 3,4-Dihydroxybenzaldehyde Purified from the Barley Seeds (Hordeum vulgare) Inhibits Oxidative DNA Damage and Apoptosis via Its Antioxidant Activity. Phytomedicine 2009, 16, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.W.; Yamane, T.; Maita, H.; Ishikawa, S.; Iguchi-Ariga, S.M.M.; Pu, X.P.; Ariga, H. DJ-1-Mediated Protective Effect of Protocatechuic Aldehyde against Oxidative Stress in SH-SY5Y Cells. J. Pharmacol. Sci. 2011, 115, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuevas-Magaña, M.Y.; Vega-García, C.C.; León-Contreras, J.C.; Hernández-Pando, R.; Zazueta, C.; García-Niño, W.R. Ellagic Acid Ameliorates Hexavalent Chromium-Induced Renal Toxicity by Attenuating Oxidative Stress, Suppressing TNF-α and Protecting Mitochondria. Toxicol. Appl. Pharmacol. 2022, 454, 116242. [Google Scholar] [CrossRef] [PubMed]
- Losso, J.N.; Bansode, R.R.; Trappey, A.; Bawadi, H.A.; Truax, R. In Vitro Anti-Proliferative Activities of Ellagic Acid. J. Nutr. Biochem. 2004, 15, 672–678. [Google Scholar] [CrossRef]
- Li, R.; Fan, X.; Zhang, T.; Song, H.; Bian, X.; Nai, R.; Li, J.; Zhang, J. Expression of selenium-independent glutathione peroxidase 5 (GPx5) in the epididymis of Small Tail Han sheep. Asian-Australas. J. Anim. Sci. 2018, 31, 1591–1597. [Google Scholar] [CrossRef]
- Gupta, D. Methods for determination of antioxidant capacity: A review. Int. J. Pharm. Sci. Res. 2015, 6, 546–566. [Google Scholar]
- Soto-Vaca, A.; Gutierrez, A.; Losso, J.N.; Xu, Z.; Finley, J.W.; Finley, J.W. Evolution of phenolic compounds from color and flavor problems to health benefits. J. Agric. Food Chem. 2012, 60, 6658–6677. [Google Scholar] [CrossRef]
- Prenzler, P.D.; Ryan, D.; Robards, K. Chapter 1. Introduction to Basic Principles of Antioxidant Activity. In Food Chemistry, Function and Analysis; Prenzler, P.D., Ryan, D., Robards, K., Eds.; Royal Society of Chemistry: Cambridge, UK, 2021; pp. 1–62. [Google Scholar] [CrossRef]
- Mamta, M.K.; Dhillon, G.S.; Brar, S.K.; Verma, M. Antioxidants. In Biotransformation of Waste Biomass into High Value Biochemicals; Brar, S.K., Dhillon, G.S., Soccol, C.R., Eds.; Springer: New York, NY, USA, 2014; pp. 117–138. [Google Scholar] [CrossRef]
- Sun, Y.; Deng, Z.; Liu, R.; Zhang, H.; Zhu, H.; Jiang, L.; Tsao, R. A comprehensive profiling of free, conjugated and bound phenolics and lipophilic antioxidants in red and green lentil processing by-products. Food Chem. 2020, 325, 126925. [Google Scholar] [CrossRef]
- Asatiani, N.; Abuladze, M.; Kartvelishvili, T.; Kulikova, N.; Asanishvili, L.; Holman, H.-Y.; Sapojnikova, N. Response of antioxidant defense system to chromium (VI)-induced cytotoxicity in human diploid cells. Biometals 2010, 23, 161–172. [Google Scholar] [CrossRef]
- Thorpe, G.W.; Fong, C.S.; Alic, N.; Higgins, V.J.; Dawes, I.W. Cells have distinct mechanisms to maintain protection against different reactive oxygen species: Oxidative-stress-response genes. Proc. Natl. Acad. Sci. USA 2004, 101, 6564–6569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamran, M.; Wang, D.; Alhaithloul, H.A.S.; Alghanem, S.M.; Aftab, T.; Xie, K.; Lu, Y.; Shi, C.; Sun, J.; Gu, W.; et al. Jasmonic acid-mediated enhanced regulation of oxidative, glyoxalase defense system and reduced chromium uptake contributes to alleviation of chromium (VI) toxicity in choysum (Brassica parachinensis L.). Ecotoxicol. Environ. Saf. 2021, 208, 111758. [Google Scholar] [CrossRef] [PubMed]
- Ordoudi, S.A.; Tsimidou, M.Z. Crocin bleaching assay step by step: Observations and suggestions for an alternative validated protocol. J. Agric. Food Chem. 2006, 54, 1663–1671. [Google Scholar] [CrossRef] [PubMed]
- Prieto, M.A.; Vázquez, J.A.; Murado, M.A. Crocin bleaching antioxidant assay revisited: Application to microplate to analyse antioxidant and pro-oxidant activities. Food Chem. 2015, 167, 299–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robak, J.; Gryglewski, R.J. Flavonoids are scavengers of superoxide anions. Biochem. Pharmacol. 1998, 37, 837–841. [Google Scholar] [CrossRef]
- Apak, R.; Güçlü, K.; Ozyürek, M.; Karademir, S.E.; Altun, M. Total antioxidant capacity assay of human serum using copper(II)neocuproine as chromogenic oxidant: The CUPRAC method. Free Radic. Res. 2005, 39, 949–996. [Google Scholar] [CrossRef]
- Kumar, C.S.; Loh, W.S.; Ooi, C.W.; Quah, C.K.; Fun, H.K. Structural correlation of some heterocyclic chalcone analogues and evaluation of their antioxidant potential. Molecules 2013, 18, 11996–12011. [Google Scholar] [CrossRef] [Green Version]
- Friel, J.K.; Diehl-Jones, W.L.; Suh, M.; Tsopmo, A.; Shirwadkar, V.P. Impact of iron and vitamin C-containing supplements on preterm human milk: In vitro. Free Radic. Biol. Med. 2007, 42, 1591–1598. [Google Scholar] [CrossRef]
- Asghar, M.N.; Khan, I.U.; Arshad, M.N.; Sherin, L. Evaluation of Antioxidant Activity Using an Improved DMPD Radical Cation Decolorization Assay. Acta Chim. Slov. 2007, 54, 295–300. [Google Scholar]
- Ndhlala, A.R.; Moyo, M.; Van Staden, J. Natural antioxidants: Fascinating or mythical biomolecules? Molecules 2010, 15, 6905–6930. [Google Scholar] [CrossRef]
- Tonguc Yayintas, O.; Sogut, O.; Konyalioglu, S.; Yilmaz, S.; Tepelİ, B. Antioxidant activities and chemical compositon of different extracts of mosses gathered from Turkey. AgroLife Sci. J. 2017, 6, 205–213. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Cell Type | Impact | Citation |
|---|---|---|
| Cardiomyocytes of broiler | Apoptosis-related genes Bax and p53 expression, mitochondrial malfunction and oxidative stress, myocardial apoptosis and autophagy | [38] |
| Liver cell of zebrafish | Downregulation of the Bcl2 gene and transcriptional activation of apoptosis-related p53, Bax, caspase 9 and caspase 3 genes, which changes the elemental composition of the liver | [39] |
| Liver and kidney cells of Sprague-Dawley rats | Dose- and time-dependent effects induced DNA damage due to increase in ROS levels | [40] |
| Hepatocytes (HepG2) of humans | Mitochondrial damage, apoptosis, oxidative stress | [41] |
| Lung epithelial cells of rats | ROS-induced cell death and activation of the p53-related pathway | [42] |
| Liver and kidney cells of carassius auratus | Oxidative stress, genotoxicity and histopathology | [43] |
| Bronchial epithelial cells (Beas-2B) of humans | Cell transformation | [44] |
| Liver and brain tissues of mice | Oxidative stress and tissue damage | [45] |
| Gill and kidney cells of Anguilla anguilla L. | Genotoxicity at a higher concentration in gills and at all concentrations in kidneys | [46] |
| Kidney cells of Wistar rats | Apoptosis and autophagy, oxidative stress in kidneys, mitochondrial dynamics disorder via inhibiting the sirt/pgc-1a pathway | [47] |
| Disease | Role of Superoxide Dismutase | References |
|---|---|---|
| Cancer | SOD mimetics, such as MnTnBuOE-2-PyP5+, play a role in stimulation or work as tumour necrosis factor, which is associated with apoptosis. | [60] |
| Skin | SOD mimetics could encourage the recovery of wounds, reduce scars and reduce pigmentation of the skin caused by ultraviolet rays. | [55] |
| Aging attenuation | SOD restores cognitive impairments caused by aging. | [61] |
| Cystic fibrosis | SOD reduces the pro-inflammatory stimulus. | [62] |
| Diabetes | SOD treatment reduces oxidative stress in the liver. An SOD mimetic Mn-II (pyane) C12 has been found effective in diabetes. Chemically modified SODs, such as carboxymethyl cellulose-SOD, have been found effective in treating diabetes. | [63,64,65,66] |
| Neurodegenerative diseases | Cu/Zn-SOD manages oxidative stress in Alzheimer’s and Parkinson’s diseases. | [67,68] |
| Rheumatoid arthritis | SOD activity reduces rheumatoid arthritis by lipid peroxidation in mitochondria, and SOD processing via liposomes has been found effective against arthritis development. | [54,69] |
| Assay | Mechanism | Absorbance (nm) | References |
|---|---|---|---|
| Oxygen radical absorbance capacity (ORAC) | HAT | 443 nm, pH = 7.0–7.5 | [90,91] |
| Ferric-reducing antioxidant potential (FRAP) | ET | 593 nm, pH = 3.6 | [92] |
| Cupric-reducing antioxidant capacity (CUPRAC) | ET | 450/490 nm, pH = 7 | [93] |
| Potassium-ferricyanide-reducing power assay (PFRAP) | ET | 700 nm, pH = 6.6 | [94] |
| Ferrous oxidation-xylenol orange (FOX) assay | ET | 550 nm | [95] |
| N,N-dimethyl-p-phenyl-diamine (DMPD) | HAT | 517 nm, pH = 5.25 | [96] |
| Thiobarbituric-acid-reactive substances (TBARS) assay | ET | 532 nm, pH = 4 | [97,98] |
| Total peroxyl-radical-trapping antioxidant parameter (TRAP) | HAT | Abs. = lambda excitation = 485, lambda emission = 538, pH = 7.4 | [90] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, V.; Singh, N.; Verma, M.; Kamal, R.; Tiwari, R.; Sanjay Chivate, M.; Rai, S.N.; Kumar, A.; Singh, A.; Singh, M.P.; et al. Hexavalent-Chromium-Induced Oxidative Stress and the Protective Role of Antioxidants against Cellular Toxicity. Antioxidants 2022, 11, 2375. https://doi.org/10.3390/antiox11122375
Singh V, Singh N, Verma M, Kamal R, Tiwari R, Sanjay Chivate M, Rai SN, Kumar A, Singh A, Singh MP, et al. Hexavalent-Chromium-Induced Oxidative Stress and the Protective Role of Antioxidants against Cellular Toxicity. Antioxidants. 2022; 11(12):2375. https://doi.org/10.3390/antiox11122375
Chicago/Turabian StyleSingh, Veer, Nidhi Singh, Manisha Verma, Rashmi Kamal, Ritesh Tiwari, Mahesh Sanjay Chivate, Sachchida Nand Rai, Ashish Kumar, Anupama Singh, Mohan P. Singh, and et al. 2022. "Hexavalent-Chromium-Induced Oxidative Stress and the Protective Role of Antioxidants against Cellular Toxicity" Antioxidants 11, no. 12: 2375. https://doi.org/10.3390/antiox11122375