
Açai Berry Attenuates Cyclophosphamide-Induced Damage in Genitourinary Axis-Modulating Nrf-2/HO-1 Pathways
 , , , , , , , , and
, , , , , , , , and 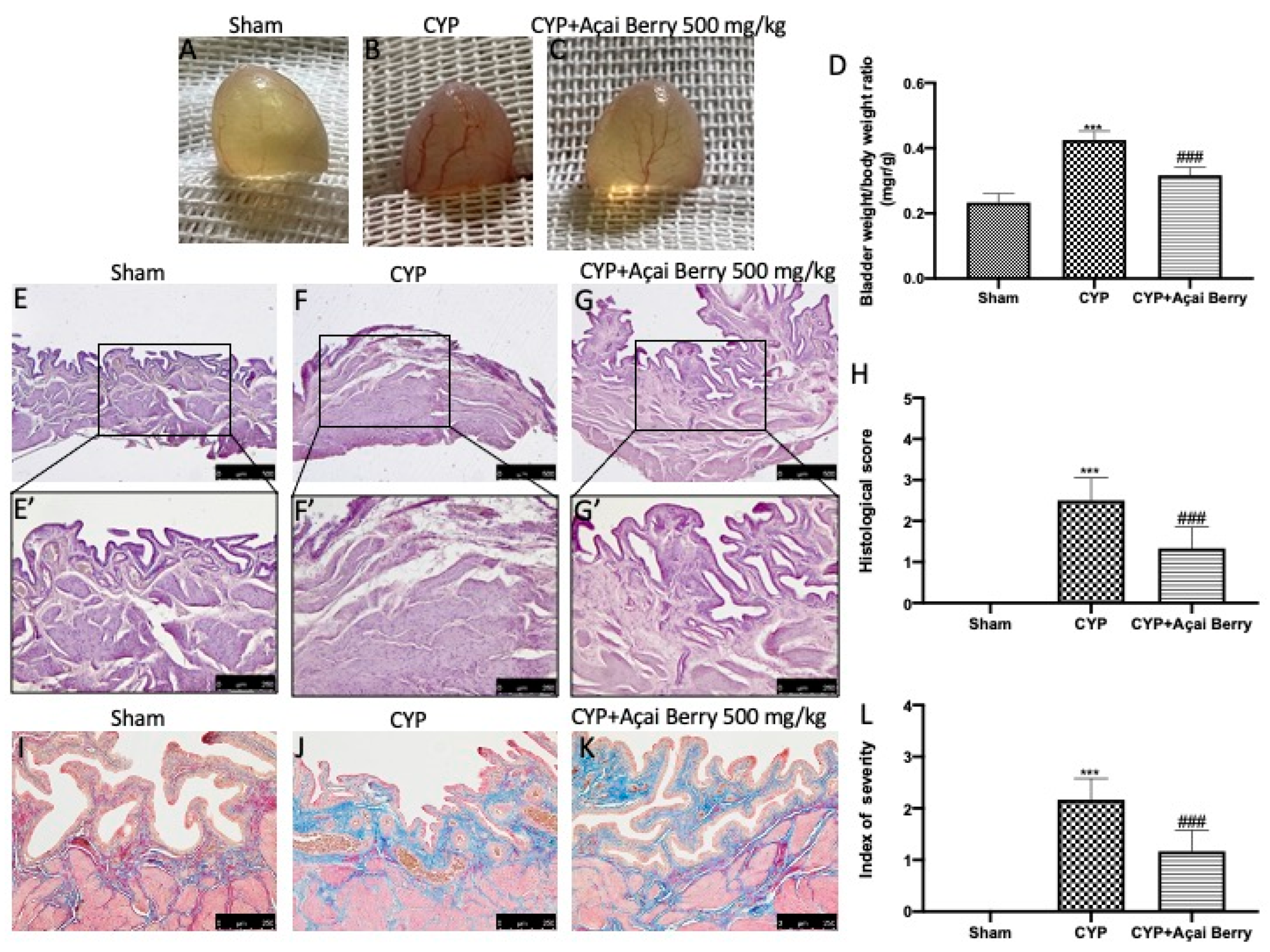{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design and Groups
- (1)
- Sham: animals were administered injections of saline and treated orally with açai berry dissolved in saline;
- (2)
- CYP: animals were administered CYP injections as described above and treated by oral gavage with saline;
- (3)
- CYP+Açai Berry: animals were administered CYP injections as described above and treated with açai berry dissolved in saline (500 mg/kg) by oral gavage 1 h after injection and for the following 5 days.
2.3. Evaluation of Sperm
2.4. Western Blot Analysis of Cytosolic and Nuclear Extracts
2.5. Testosterone Assay
2.6. Histopathological Evaluation
2.7. Evaluation of Tissue Lipid Peroxidation
2.8. Assessment of Tissue Antioxidant Activity
2.9. Terminal Deoxynucleotidyl Nick-End Labeling (TUNEL) Assay
2.10. Cytokine Levels
2.11. Materials
2.12. Statistical Evaluation
3. Results
3.1. Açai Berry Limitis CYP-Induced Macroscopic and Microscopic Alterations
3.2. Effect of Açai Berry Administration on CYP-Induced Bladder Oxidative Stress and Lipid Peroxidation
3.3. Effect of Açai Berry Administration on Apoptosis Pathways after CYP
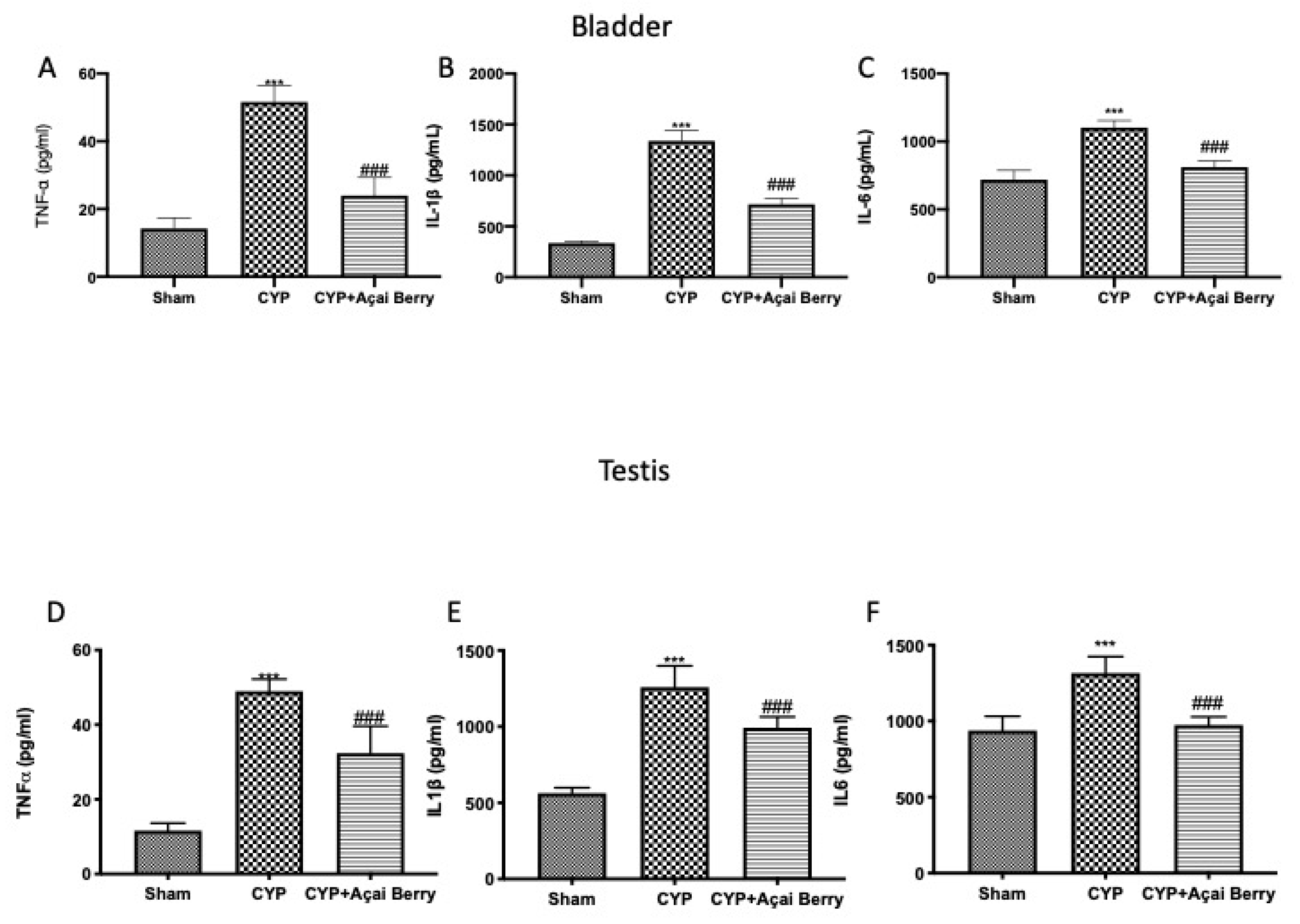
3.4. Effect of Acai Berry Administration on Cytokyne Storm in Bladder and Testes after CYP
3.5. Effect of Açai Berry Administration on Sperm Parameters and Testosterone Levels after CYP
3.6. Effect of Açai Berry on Histological Alteration in Testes after CYP
3.7. Açai Berry Administration Improved Endogenous Oxidative Defense in Testes
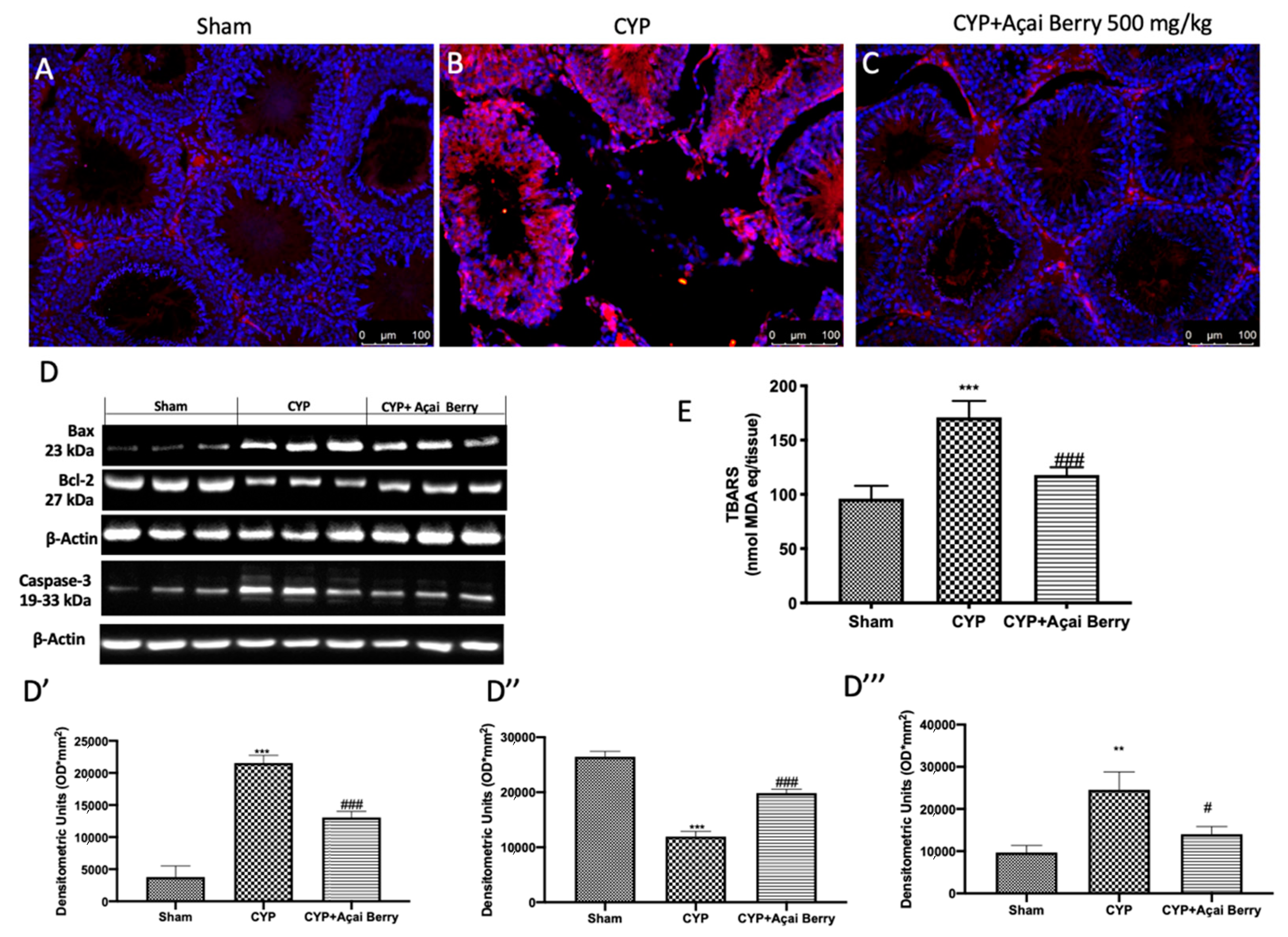
3.8. Açai Berry Administration Limits CYP-Induced Apoptosis and TBARS in Testes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Umar, A.; Dunn, B.K.; Greenwald, P. Future directions in cancer prevention. Nat. Rev. Cancer 2012, 12, 835–848. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, M.A.; Antin, J.H.; Guinan, E.C.; Rappeport, J.M. Cyclophosphamide Cardiotoxicity—An Analysis of Dosing as a Risk Factor. Blood 1986, 68, 1114–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraiser, L.H.; Kanekal, S.; Kehrer, J.P. Cyclophosphamide Toxicity—Characterizing and Avoiding the Problem. Drugs 1991, 42, 781–795. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, X.-L.; Lin, T.; He, D.-W.; Wei, G.-H.; Liu, J.-H.; Li, L.-S. The cyclophosphamide metabolite, acrolein, induces cytoskeletal changes and oxidative stress in Sertoli cells. Mol. Biol. Rep. 2012, 39, 493–500. [Google Scholar] [CrossRef]
- Balow, J.E.; Austin, H.A., 3rd; Tsokos, G.C.; Antonovych, T.T.; Steinberg, A.D.; Klippel, J.H. NIH conference. Lupus nephritis. Ann. Intern. Med. 1987, 106, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Pagnoux, C. Updates in ANCA-associated vasculitis. Eur. J. Rheumatol. 2016, 3, 122–133. [Google Scholar] [CrossRef]
- Saba; Khan, S.; Parvez, S.; Chaudhari, B.; Ahmad, F.; Anjum, S.; Raisuddin, S. Ellagic acid attenuates bleomycin and cyclophosphamide-induced pulmonary toxicity in Wistar rats. Food Chem. Toxicol. 2013, 58, 210–219. [Google Scholar] [CrossRef]
- Zhang, J.; Tian, Q.; Chan, S.Y.; Li, S.C.; Zhou, S.; Duan, W.; Zhu, Y.-Z. Metabolism and Transport of Oxazaphosphorines and the Clinical Implications. Drug Metab. Rev. 2005, 37, 611–703. [Google Scholar] [CrossRef]
- Boddy, A.V.; Yule, S.M. Metabolism and Pharmacokinetics of Oxazaphosphorines. Clin. Pharmacokinet. 2000, 38, 291–304. [Google Scholar] [CrossRef]
- Stork, C.; Schreffler, S. Cyclophosphamide. In Encyclopedia of Toxicology; Elsevier: New York, NY, USA, 2014. [Google Scholar]
- Qin, W.-S.; Deng, Y.-H.; Cui, F.-C. Sulforaphane protects against acrolein-induced oxidative stress and inflammatory responses: Modulation of Nrf-2 and COX-2 expression. Arch. Med. Sci. 2016, 12, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ito, S.; Nishio, N.; Tanaka, Y.; Chen, N.; Liu, L.; Isobe, K.-I. Enhancement of the Acrolein-Induced Production of Reactive Oxygen Species and Lung Injury by GADD. Oxidative Med. Cell. Longev. 2015, 2015, 170309. [Google Scholar] [CrossRef] [Green Version]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Darbandi, M.; Darbandi, S.; Agarwal, A.; Baskaran, S.; Sengupta, P.; Dutta, S.; Mokarram, P.; Saliminejad, K.; Sadeghi, M.R. Oxidative stress-induced alterations in seminal plasma antioxidants: Is there any association with keap1 gene methylation in human spermatozoa? Andrologia 2019, 51, e13159. [Google Scholar] [CrossRef] [PubMed]
- Turner, T.T.; Lysiak, J.J. Oxidative Stress: A Common Factor in Testicular Dysfunction. J. Androl. 2008, 29, 488–498. [Google Scholar] [CrossRef] [Green Version]
- Darbandi, M.; Darbandi, S.; Khorshid, H.R.K.; Akhondi, M.M.; Mokarram, P.; Sadeghi, M.R. A simple, rapid and economic manual method for human sperm DNA extraction in genetic and epigenetic studies. Middle East Fertil. Soc. J. 2018, 23, 216–219. [Google Scholar] [CrossRef]
- Li, Y.; Tollefsbol, T.O. DNA Methylation Detection: Bisulfite Genomic Sequencing Analysis. In Epigenetics Protocols; Springer: Berlin/Heidelberg, Germany, 2011; pp. 11–21. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Yu, S.; Zhang, C.; Kong, A.-N.T. Epigenetic regulation of Keap1-Nrf2 signaling. Free Radic. Biol. Med. 2015, 88, 337–349. [Google Scholar] [CrossRef] [Green Version]
- Chabra, A.; Shokrzadeh, M.; Naghshvar, F.; Salehi, F.; Ahmadi, A. Melatonin ameliorates oxidative stress and reproductive toxicity induced by cyclophosphamide in male mice. Hum. Exp. Toxicol. 2014, 33, 185–195. [Google Scholar] [CrossRef]
- Kim, J.-K.; Jang, H.-D. Nrf2-Mediated HO-1 Induction Coupled with the ERK Signaling Pathway Contributes to Indirect Antioxidant Capacity of Caffeic Acid Phenethyl Ester in HepG2 Cells. Int. J. Mol. Sci. 2014, 15, 12149–12165. [Google Scholar] [CrossRef] [Green Version]
- Ku, B.M.; Joo, Y.; Mun, J.; Roh, G.S.; Kang, S.S.; Cho, G.J.; Choi, W.S.; Kim, H.J. Heme oxygenase protects hippocampal neurons from ethanol-induced neurotoxicity. Neurosci. Lett. 2006, 405, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Radjendirane, V.; Joseph, P.; Lee, Y.-H.; Kimura, S.; Klein-Szanto, A.J.; Gonzalez, F.J.; Jaiswal, A.K. Disruption of the DT Diaphorase (NQO1) Gene in Mice Leads to Increased Menadione Toxicity. J. Biol. Chem. 1998, 273, 7382–7389. [Google Scholar] [CrossRef] [Green Version]
- Salim, S. Oxidative Stress and the Central Nervous System. J. Pharmacol. Exp. Ther. 2017, 360, 201–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, D.; Wu, R.; Guo, Y.; Kong, A.-N.T. Regulation of Keap1–Nrf2 signaling: The role of epigenetics. Curr. Opin. Toxicol. 2016, 1, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Hagan, S.; Khurana, N.; Chandra, S.; Abdel-Mageed, A.B.; Mondal, D.; Hellstrom, W.J.G.; Sikka, S.C. Differential expression of novel biomarkers (TLR-2, TLR-4, COX-2, and Nrf-2) of inflammation and oxidative stress in semen of leukocytospermia patients. Andrology 2015, 3, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Hamada, A.; Sharma, R.; du Plessis, S.S.; Willard, B.; Yadav, S.P.; Sabanegh, E.; Agarwal, A. Two-dimensional differential in-gel electrophoresis–based proteomics of male gametes in relation to oxidative stress. Fertil. Steril. 2013, 99, 1216–1226.e2. [Google Scholar] [CrossRef]
- Yu, B.; Huang, Z. Variations in Antioxidant Genes and Male Infertility. BioMed Res. Int. 2015, 2015, 513196. [Google Scholar] [CrossRef] [Green Version]
- Visalberghi, E.; Barca, V.; Izar, P.; Fragaszy, D.; Truppa, V. Optional tool use: The case of wild bearded capuchins (Sapajus libidinosus) cracking cashew nuts by biting or by using percussors. Am. J. Primatol. 2021, 83, e23221. [Google Scholar] [CrossRef] [PubMed]
- Kilanko, O.; Ojolo, S.J.; Leramo, R.O.; Ilori, T.A.; Oyedepo, S.O.; Babalola, P.O.; Fayomi, O.S.; Onwordi, P.N.; Ufot, E.; Ekwere, A. Dataset on physical properties of raw and roasted cashew nuts. Data Brief 2020, 33, 106514. [Google Scholar] [CrossRef] [PubMed]
- Cordaro, M.; Fusco, R.; D’Amico, R.; Siracusa, R.; Peritore, A.; Gugliandolo, E.; Genovese, T.; Crupi, R.; Mandalari, G.; Cuzzocrea, S.; et al. Cashew (Anacardium occidentale L.) Nuts Modulate the Nrf2 and NLRP3 Pathways in Pancreas and Lung after Induction of Acute Pancreatitis by Cerulein. Antioxidants 2020, 9, 992. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; Cordaro, M.; Siracusa, R.; Peritore, A.F.; Gugliandolo, E.; Genovese, T.; D’Amico, R.; Crupi, R.; Smeriglio, A.; Mandalari, G.; et al. Consumption of Anacardium occidentale L. (Cashew Nuts) Inhibits Oxidative Stress through Modulation of the Nrf2/HO−1 and NF-kB Pathways. Molecules 2020, 25, 4426. [Google Scholar] [CrossRef]
- Cordaro, M.; Siracusa, R.; Fusco, R.; D’Amico, R.; Peritore, A.; Gugliandolo, E.; Genovese, T.; Scuto, M.; Crupi, R.; Mandalari, G.; et al. Cashew (Anacardium occidentale L.) Nuts Counteract Oxidative Stress and Inflammation in an Acute Experimental Model of Carrageenan-Induced Paw Edema. Antioxidants 2020, 9, 660. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; Siracusa, R.; Peritore, A.F.; Gugliandolo, E.; Genovese, T.; D’Amico, R.; Cordaro, M.; Crupi, R.; Mandalari, G.; Impellizzeri, D.; et al. The Role of Cashew (Anacardium occidentale L.) Nuts on an Experimental Model of Painful Degenerative Joint Disease. Antioxidants 2020, 9, 511. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, R.; Fusco, R.; Peritore, A.F.; Cordaro, M.; D’Amico, R.; Genovese, T.; Gugliandolo, E.; Crupi, R.; Smeriglio, A.; Mandalari, G.; et al. The Antioxidant and Anti-Inflammatory Properties of Anacardium occidentale L. Cashew Nuts in a Mouse Model of Colitis. Nutrients 2020, 12, 834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavin, N.F.; Izaguirry, A.P.; Soares, M.B.; Spiazzi, C.C.; Mendez, A.S.L.; Leivas, F.G.; Brum, D.D.S.; Cibin, F.W.S. Tribulus terrestris Protects against Male Reproductive Damage Induced by Cyclophosphamide in Mice. Oxidative Med. Cell. Longev. 2018, 2018, 5758191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Song, W.; Wang, Z.; Wang, Z.; Jin, X.; Xu, J.; Bai, L.; Li, Y.; Cui, J.; Cai, L. Resveratrol attenuates testicular apoptosis in type 1 diabetic mice: Role of Akt-mediated Nrf2 activation and p62-dependent Keap1 degradation. Redox Biol. 2018, 14, 609–617. [Google Scholar] [CrossRef]
- Ramírez-Tortose, M.D.C.; Pulido-Morán, M.; Granados, S.; Gaforio, J.J.; Quiles, J.L. Hydroxytyrosol as a Component of the Mediterranean Diet and Its Role in Disease Prevention. In The Mediterranean Diet; Elsevier: Amsterdam, The Netherlands, 2015; pp. 205–215. [Google Scholar] [CrossRef]
- Maciel-Silva, F.W.; Buller, L.S.; MBB Gonçalves, M.L.; Rostagno, M.A.; Forster-Carneiro, T. Sustainable development in the Legal Amazon: Energy recovery from açaí seeds. Biofuels Bioprod. Biorefining 2021, 15, 1174–1189. [Google Scholar] [CrossRef]
- Melo, P.S.; Massarioli, A.P.; Lazarini, J.G.; Soares, J.C.; Franchin, M.; Rosalen, P.L.; de Alencar, S.M. Simulated gastrointestinal digestion of Brazilian açaí seeds affects the content of flavan-3-ol derivatives, and their antioxidant and anti-inflammatory activities. Heliyon 2020, 6, e05214. [Google Scholar] [CrossRef] [PubMed]
- Melo, P.S.; Selani, M.M.; Gonçalves, R.H.; de Oliveira Paulino, J.; Massarioli, A.P.; de Alencar, S.M. Açaí seeds: An unexplored agro-industrial residue as a potential source of lipids, fibers, and antioxidant phenolic compounds. Ind. Crops Prod. 2021, 161, 113204. [Google Scholar] [CrossRef]
- Rodrigues, R.B.; Lichtenthäler, R.; Zimmermann, B.F.; Papagiannopoulos, M.; Fabricius, H.; Marx, F.; Maia, J.G.S.; Almeida, O. Total Oxidant Scavenging Capacity of Euterpe oleracea Mart. (Açaí) Seeds and Identification of Their Polyphenolic Compounds. J. Agric. Food Chem. 2006, 54, 4162–4167. [Google Scholar] [CrossRef]
- De Moura, R.S.; Pires, K.M.P.; Ferreira, T.S.; Lopes, A.D.A.; Nesi, R.T.; Resende, A.C.; Sousa, P.J.C.; da Silva, A.J.R.; Porto, L.C.; Valenca, S. Addition of açaí (Euterpe oleracea) to cigarettes has a protective effect against emphysema in mice. Food Chem. Toxicol. 2011, 49, 855–863. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kim, N.; Choi, Y.J.; Nam, R.H.; Lee, S.; Ham, M.H.; Suh, J.H.; Choi, Y.J.; Lee, H.S.; Lee, N.H. Anti-inflammatory and Anti-tumorigenic Effects of Açai Berry in Helicobacter felis-infected mice. J. Cancer Prev. 2016, 21, 48–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Moura, R.S.; Ferreira, T.S.; Lopes, A.A.; Pires, K.M.P.; Nesi, R.T.; Resende, A.C.; Souza, P.J.C.; da Silva, A.J.R.; Borges, R.M.; Porto, L.C.; et al. Effects of Euterpe oleracea Mart. (AÇAÍ) extract in acute lung inflammation induced by cigarette smoke in the mouse. Phytomedicine 2012, 19, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Poulose, S.M.; Fisher, D.R.; Larson, J.; Bielinski, D.F.; Rimando, A.M.; Carey, A.N.; Schauss, A.G.; Shukitt-Hale, B. Anthocyanin-rich Açai (Euterpe oleracea Mart.) Fruit Pulp Fractions Attenuate Inflammatory Stress Signaling in Mouse Brain BV-2 Microglial Cells. J. Agric. Food Chem. 2012, 60, 1084–1093. [Google Scholar] [CrossRef] [PubMed]
- Santos, I.B.; de Bem, G.F.; da Costa, C.A.; de Carvalho, L.C.R.M.; de Medeiros, A.F.; Silva, D.L.B.; Romão, M.H.; Soares, R.D.A.; Ognibene, D.T.; de Moura, R.S.; et al. Açaí seed extract prevents the renin-angiotensin system activation, oxidative stress and inflammation in white adipose tissue of high-fat diet–fed mice. Nutr. Res. 2020, 79, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Alnasser, M.N.; Mellor, I.R. Neuroprotective activities of acai berries (Euterpe sp.): A review. J. Herbmed Pharmacol. 2022, 11, 166–181. [Google Scholar] [CrossRef]
- D’Amico, R.; Fusco, R.; Cordaro, M.; Siracusa, R.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Scuto, M.; Cuzzocrea, S.; Di Paola, R.; et al. Modulation of NLRP3 Inflammasome through Formyl Peptide Receptor 1 (Fpr-1) Pathway as a New Therapeutic Target in Bronchiolitis Obliterans Syndrome. Int. J. Mol. Sci. 2020, 21, 2144. [Google Scholar] [CrossRef] [Green Version]
- Impellizzeri, D.; Siracusa, R.; Cordaro, M.; Crupi, R.; Peritore, A.F.; Gugliandolo, E.; D’Amico, R.; Petrosino, S.; Evangelista, M.; Di Paola, R.; et al. N-Palmitoylethanolamine-oxazoline (PEA-OXA): A new therapeutic strategy to reduce neuroinflammation, oxidative stress associated to vascular dementia in an experimental model of repeated bilateral common carotid arteries occlusion. Neurobiol. Dis. 2019, 125, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, R.; Paterniti, I.; Cordaro, M.; Crupi, R.; Bruschetta, G.; Campolo, M.; Cuzzocrea, S.; Esposito, E. Neuroprotective Effects of Temsirolimus in Animal Models of Parkinson’s Disease. Mol. Neurobiol. 2018, 55, 2403–2419. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E.; Impellizzeri, D.; Bruschetta, G.; Cordaro, M.; Siracusa, R.; Gugliandolo, E.; Crupi, R.; Cuzzocrea, S. A new co-micronized composite containing palmitoylethanolamide and polydatin shows superior oral efficacy compared to their association in a rat paw model of carrageenan-induced inflammation. Eur. J. Pharmacol. 2016, 782, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Siracusa, R.; Cordaro, M.; Peritore, A.F.; Gugliandolo, E.; Mancuso, G.; Midiri, A.; Di Paola, R.; Cuzzocrea, S. Therapeutic potential of dinitrobenzene sulfonic acid (DNBS)-induced colitis in mice by targeting IL-1β and IL-18. Biochem. Pharmacol. 2018, 155, 150–161. [Google Scholar] [CrossRef]
- Cordaro, M.; Cuzzocrea, S.; Crupi, R. An Update of Palmitoylethanolamide and Luteolin Effects in Preclinical and Clinical Studies of Neuroinflammatory Events. Antioxidants 2020, 9, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.; Li, Z.; Wu, T.; Jensen, G.S.; Schauss, A.G.; Wu, X. Anti-oxidant capacities of flavonoid compounds isolated from acai pulp (Euterpe oleracea Mart.). Food Chem. 2010, 122, 610–617. [Google Scholar] [CrossRef]
- Arnoso, B.J.D.M.; Magliaccio, F.M.; de Araújo, C.A.; Soares, R.D.A.; Santos, I.B.; de Bem, G.F.; Fernandes-Santos, C.; Ognibene, D.T.; de Moura, R.S.; Resende, A.C.; et al. Açaí seed extract (ASE) rich in proanthocyanidins improves cardiovascular remodeling by increasing antioxidant response in obese high-fat diet-fed mice. Chem. Interact. 2022, 351, 109721. [Google Scholar] [CrossRef]
- Bellucci, E.R.B.; dos Santos, J.M.; Carvalho, L.T.; Borgonovi, T.F.; Lorenzo, J.M.; da Silva-Barretto, A.C. Açaí extract powder as natural antioxidant on pork patties during the refrigerated storage. Meat Sci. 2022, 184, 108667. [Google Scholar] [CrossRef]
- da Silva, T.V.N.; Torres, M.F.; Sampaio, L.A.; Hamoy, M.; Monserrat, J.M.; Barbas, L.A.L. Dietary Euterpe oleracea Mart. attenuates seizures and damage to lipids in the brain of Colossoma macropomum. Fish Physiol. Biochem. 2021, 47, 1851–1864. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.J.; Kim, Y.; Jin, S.G.; Kim, J.Y. Acai berry extract as a regulator of intestinal inflammation pathways in a Caco-2 and RAW 264.7 co-culture model. J. Food Biochem. 2021, 45, e13848. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Costa, J.; Pacheco-Fill, T.; Ruiz, A.; Vidal, F.; Borges, K.; Guimarães, S.; Azevedo-Santos, A.; Buglio, K.; Foglio, M.; et al. Açai (Euterpe oleracea Mart.) Seed Extract Induces ROS Production and Cell Death in MCF-7 Breast Cancer Cell Line. Molecules 2021, 26, 3546. [Google Scholar] [CrossRef]
- Martins, G.R.; Guedes, D.; de Paula, U.L.M.; de Oliveira, M.D.S.P.; Lutterbach, M.T.S.; Reznik, L.Y.; Sérvulo, E.F.C.; Alviano, C.S.; da Silva, A.J.R.; Alviano, D.S. Açaí (Euterpe oleracea Mart.) Seed Extracts from Different Varieties: A Source of Proanthocyanidins and Eco-Friendly Corrosion Inhibition Activity. Molecules 2021, 26, 3433. [Google Scholar] [CrossRef] [PubMed]
- De Bem, M.G.F.; Okinga, A.; Ognibene, D.T.; da Costa, C.A.; Santos, M.I.B.; Soares, R.A.; Silva, M.D.L.B.; da Rocha, M.A.P.M.; Fernandes, J.I.; Fraga, M.C.; et al. Anxiolytic and antioxidant effects of Euterpe oleracea Mart. (açaí) seed extract in adult rat offspring submitted to periodic maternal separation. Appl. Physiol. Nutr. Metab. 2020, 45, 1277–1286. [Google Scholar] [CrossRef]
- Remigante, A.; Spinelli, S.; Straface, E.; Gambardella, L.; Caruso, D.; Falliti, G.; Dossena, S.; Marino, A.; Morabito, R. Açaì (Euterpe oleracea) Extract Protects Human Erythrocytes from Age-Related Oxidative Stress. Cells 2022, 11, 2391. [Google Scholar] [CrossRef] [PubMed]
- Amiri, F.T.; Hamzeh, M.; Hosseinimehr, S.J.; Karimpour, A.; Mohammadi, H.R.; Khalatbary, A.R. Cerium oxide nanoparticles protect cyclophosphamide-induced testicular toxicity in mice. Int. J. Prev. Med. 2019, 10, 5. [Google Scholar] [CrossRef]
- Fusco, R.; Salinaro, A.; Siracusa, R.; D’Amico, R.; Impellizzeri, D.; Scuto, M.; Ontario, M.; Crea, R.; Cordaro, M.; Cuzzocrea, S.; et al. Hidrox® Counteracts Cyclophosphamide-Induced Male Infertility through NRF2 Pathways in a Mouse Model. Antioxidants 2021, 10, 778. [Google Scholar] [CrossRef]
- Impellizzeri, D.; D’Amico, R.; Fusco, R.; Genovese, T.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Interdonato, L.; Di Paola, D.; Di Paola, R.; et al. Açai Berry Mitigates Vascular Dementia-Induced Neuropathological Alterations Modulating Nrf-2/Beclin1 Pathways. Cells 2022, 11, 2616. [Google Scholar] [CrossRef]
- D’Amico, R.; Impellizzeri, D.; Genovese, T.; Fusco, R.; Peritore, A.F.; Crupi, R.; Interdonato, L.; Franco, G.; Marino, Y.; Arangia, A.; et al. Açai Berry Mitigates Parkinson’s Disease Progression Showing Dopaminergic Neuroprotection via Nrf2-HO1 Pathways. Mol. Neurobiol. 2022, 59, 6519–6533. [Google Scholar] [CrossRef] [PubMed]
- Genovese, T.; D’Amico, R.; Fusco, R.; Impellizzeri, D.; Peritore, A.F.; Crupi, R.; Interdonato, L.; Gugliandolo, E.; Cuzzocrea, S.; Di Paola, R.; et al. Açaí (Euterpe oleraceae Mart.) Seeds Regulate NF-κB and Nrf2/ARE Pathways Protecting Lung against Acute and Chronic Inflammation. Cell. Physiol. Biochem. 2022, 56, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Aksu, E.H.; Kandemir, F.M.; Özkaraca, M.; Ömür, A.D.; Küçükler, S.; Çomaklı, S. Rutin ameliorates cisplatin-induced reproductive damage via suppression of oxidative stress and apoptosis in adult male rats. Andrologia 2017, 49, e12593. [Google Scholar] [CrossRef] [PubMed]
- Cordaro, M.; Paterniti, I.; Siracusa, R.; Impellizzeri, D.; Esposito, E.; Cuzzocrea, S. KU0063794, a Dual mTORC1 and mTORC2 Inhibitor, Reduces Neural Tissue Damage and Locomotor Impairment After Spinal Cord Injury in Mice. Mol. Neurobiol. 2017, 54, 2415–2427. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, R.; Impellizzeri, D.; Cordaro, M.; Crupi, R.; Esposito, E.; Petrosino, S.; Cuzzocrea, S. Anti-Inflammatory and Neuroprotective Effects of Co-UltraPEALut in a Mouse Model of Vascular Dementia. Front. Neurol. 2017, 8, 233. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, D.; Iaria, C.; Capparucci, F.; Cordaro, M.; Crupi, R.; Siracusa, R.; D’Amico, R.; Fusco, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Aflatoxin B1 Toxicity in Zebrafish Larva (Danio rerio): Protective Role of Hericium erinaceus. Toxins 2021, 13, 710. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, D.; Natale, S.; Gugliandolo, E.; Cordaro, M.; Crupi, R.; Siracusa, R.; D’Amico, R.; Fusco, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Assessment of 2-Pentadecyl-2-oxazoline Role on Lipopolysaccharide-Induced Inflammation on Early Stage Development of Zebrafish (Danio rerio). Life 2022, 12, 128. [Google Scholar] [CrossRef]
- Di Paola, D.; Capparucci, F.; Natale, S.; Crupi, R.; Cuzzocrea, S.; Spanò, N.; Gugliandolo, E.; Peritore, A.F. Combined Effects of Potassium Perchlorate and a Neonicotinoid on Zebrafish Larvae (Danio rerio). Toxics 2022, 10, 203. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, D.; Capparucci, F.; Lanteri, G.; Cordaro, M.; Crupi, R.; Siracusa, R.; D’Amico, R.; Fusco, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Combined Toxicity of Xenobiotics Bisphenol A and Heavy Metals on Zebrafish Embryos (Danio rerio). Toxics 2021, 9, 344. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, D.; Natale, S.; Iaria, C.; Crupi, R.; Cuzzocrea, S.; Spanò, N.; Gugliandolo, E.; Peritore, A.F. Environmental Co-Exposure to Potassium Perchlorate and Cd Caused Toxicity and Thyroid Endocrine Disruption in Zebrafish Embryos and Larvae (Danio rerio). Toxics 2022, 10, 198. [Google Scholar] [CrossRef] [PubMed]
- Salimnejad, R.; Rad, J.S.; Nejad, D.M. Protective Effect of Ghrelin on Oxidative Stress and Tissue Damages of Mice Testes Followed by Chemotherapy with Cyclophosphamide. Crescent J. Med. Biol. Sci. 2018, 5, 138–143. [Google Scholar]
- D’Amico, R.; Salinaro, A.T.; Cordaro, M.; Fusco, R.; Impellizzeri, D.; Interdonato, L.; Scuto, M.; Ontario, M.; Crea, R.; Siracusa, R.; et al. Hidrox® and Chronic Cystitis: Biochemical Evaluation of Inflammation, Oxidative Stress, and Pain. Antioxidants 2021, 10, 1046. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, D.; Abbate, J.M.; Iaria, C.; Cordaro, M.; Crupi, R.; Siracusa, R.; D’Amico, R.; Fusco, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Environmental Risk Assessment of Dexamethasone Sodium Phosphate and Tocilizumab Mixture in Zebrafish Early Life Stage (Danio rerio). Toxics 2022, 10, 279. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Kamil, J.; Jeon, Y.-J.; Kim, S.-K. Antioxidant Role of Chitosan in a Cooked Cod (Gadus morhua) Model System. J. Food Lipids 2002, 9, 57–64. [Google Scholar] [CrossRef]
- Misra, H.P.; Fridovich, I. The Role of Superoxide Anion in the Autoxidation of Epinephrine and a Simple Assay for Superoxide Dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Bonaventura, J.; Schroeder, W.; Fang, S. Human erythrocyte catalase: An improved method of isolation and a reevaluation of reported properties. Arch. Biochem. Biophys. 1972, 150, 606–617. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Di Paola, D.; Capparucci, F.; Abbate, J.M.; Cordaro, M.; Crupi, R.; Siracusa, R.; D’Amico, R.; Fusco, R.; Genovese, T.; Impellizzeri, D.; et al. Environmental Risk Assessment of Oxaliplatin Exposure on Early Life Stages of Zebrafish (Danio rerio). Toxics 2022, 10, 81. [Google Scholar] [CrossRef]
- Di Paola, D.; Capparucci, F.; Lanteri, G.; Crupi, R.; Marino, Y.; Franco, G.A.; Cuzzocrea, S.; Spanò, N.; Gugliandolo, E.; Peritore, A.F. Environmental Toxicity Assessment of Sodium Fluoride and Platinum-Derived Drugs Co-Exposure on Aquatic Organisms. Toxics 2022, 10, 272. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, D.; Natale, S.; Iaria, C.; Cordaro, M.; Crupi, R.; Siracusa, R.; D’Amico, R.; Fusco, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Intestinal Disorder in Zebrafish Larvae (Danio rerio): The Protective Action of N-Palmitoylethanolamide-oxazoline. Life 2022, 12, 125. [Google Scholar] [CrossRef] [PubMed]
- Genovese, T.; Impellizzeri, D.; D’Amico, R.; Fusco, R.; Peritore, A.F.; Di Paola, D.; Interdonato, L.; Gugliandolo, E.; Crupi, R.; Di Paola, R.; et al. Role of Bevacizumab on Vascular Endothelial Growth Factor in Apolipoprotein E Deficient Mice after Traumatic Brain Injury. Int. J. Mol. Sci. 2022, 23, 4162. [Google Scholar] [CrossRef] [PubMed]
- Cordaro, M.; Siracusa, R.; D’Amico, R.; Genovese, T.; Franco, G.; Marino, Y.; Di Paola, D.; Cuzzocrea, S.; Impellizzeri, D.; Di Paola, R.; et al. Role of Etanercept and Infliximab on Nociceptive Changes Induced by the Experimental Model of Fibromyalgia. Int. J. Mol. Sci. 2022, 23, 6139. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, R.; Gugliandolo, E.; Cordaro, M.; Fusco, R.; Genovese, T.; Peritore, A.F.; Crupi, R.; Interdonato, L.; Di Paola, D.; Cuzzocrea, S.; et al. Toxic Effects of Endocrine Disruptor Exposure on Collagen-Induced Arthritis. Biomolecules 2022, 12, 564. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, R.; Gugliandolo, E.; Siracusa, R.; Cordaro, M.; Genovese, T.; Peritore, A.F.; Crupi, R.; Interdonato, L.; Di Paola, D.; Cuzzocrea, S.; et al. Toxic Exposure to Endocrine Disruptors Worsens Parkinson’s Disease Progression through NRF2/HO-1 Alteration. Biomedicines 2022, 10, 1073. [Google Scholar] [CrossRef]
- Cordaro, M.; Siracusa, R.; Crupi, R.; Impellizzeri, D.; Peritore, A.F.; D’Amico, R.; Gugliandolo, E.; Di Paola, R.; Cuzzocrea, S. 2-Pentadecyl-2-Oxazoline Reduces Neuroinflammatory Environment in the MPTP Model of Parkinson Disease. Mol. Neurobiol. 2018, 55, 9251–9266. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Cordaro, M.; Bruschetta, G.; Crupi, R.; Pascali, J.P.; Alfonsi, D.; Marcolongo, G.; Cuzzocrea, S. 2-pentadecyl-2-oxazoline: Identification in coffee, synthesis and activity in a rat model of carrageenan-induced hindpaw inflammation. Pharmacol. Res. 2016, 108, 23–30. [Google Scholar] [CrossRef]
- Di Paola, R.; Fusco, R.; Impellizzeri, D.; Cordaro, M.; Britti, D.; Morittu, V.M.; Evangelista, M.; Cuzzocrea, S. Adelmidrol, in combination with hyaluronic acid, displays increased anti-inflammatory and analgesic effects against monosodium iodoacetate-induced osteoarthritis in rats. Arthritis Res. Ther. 2016, 18, 291. [Google Scholar] [CrossRef]
- Fusco, R.; Cordaro, M.; Genovese, T.; Impellizzeri, D.; Siracusa, R.; Gugliandolo, E.; Peritore, A.; D’Amico, R.; Crupi, R.; Cuzzocrea, S.; et al. Adelmidrol: A New Promising Antioxidant and Anti-Inflammatory Therapeutic Tool in Pulmonary Fibrosis. Antioxidants 2020, 9, 601. [Google Scholar] [CrossRef]
- Fusco, R.; Cordaro, M.; Siracusa, R.; D’Amico, R.; Genovese, T.; Gugliandolo, E.; Peritore, A.F.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Biochemical Evaluation of the Antioxidant Effects of Hydroxytyrosol on Pancreatitis-Associated Gut Injury. Antioxidants 2020, 9, 781. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; Gugliandolo, E.; Siracusa, R.; Scuto, M.; Cordaro, M.; D’Amico, R.; Evangelista, M.; Peli, A.; Peritore, A.; Impellizzeri, D.; et al. Formyl Peptide Receptor 1 Signaling in Acute Inflammation and Neural Differentiation Induced by Traumatic Brain Injury. Biology 2020, 9, 238. [Google Scholar] [CrossRef] [PubMed]
- Brock, N.; Wilmanns, H. Effect of a cyclic nitrogen mustard-phosphamidester on experimentally induced tumors in rats; chemotherapeutic effect and pharmacological properties of B 518 ASTA. DMW Dtsch. Med. Wochenschr. 1958, 83, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.; Jena, G. Astaxanthin inhibits cytotoxic and genotoxic effects of cyclophosphamide in mice germ cells. Toxicology 2008, 248, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Fiorucci, E.; Lucantoni, G.; Paone, G.; Zotti, M.; Li, B.E.; Serpilli, M.; Regimenti, P.; Cammarella, I.; Puglisi, G.; Schmid, G. Colchicine, cyclophosphamide and prednisone in the treatment of mild-moderate idiopathic pulmonary fibrosis: Comparison of three currently available therapeutic regimens. Eur. Rev. Med Pharmacol. Sci. 2008, 12, 105. [Google Scholar]
- Tripathi, D.N.; Jena, G.B. Intervention of astaxanthin against cyclophosphamide-induced oxidative stress and DNA damage: A study in mice. Chem. Biol. Interact. 2009, 180, 398–406. [Google Scholar] [CrossRef]
- Forni, A.M.; Koss, L.G.; Geller, W. Cytological study of the effect of cyclophosphamide on the epithelium of the urinary bladder in man. Cancer 1964, 17, 1348–1355. [Google Scholar] [CrossRef]
- Fairchild, W.V.; Spence, C.R.; Solomon, H.D.; Gangai, M.P. The Incidence of Bladder Cancer after Cyclophosphamide Therapy. J. Urol. 1979, 122, 163–164. [Google Scholar] [CrossRef]
- Christophidis, N. Carcinoma of the urinary bladder in patients receiving cyclophosphamide. Aust. N. Z. J. Med. 1985, 15, 87. [Google Scholar] [CrossRef]
- Rezvanfar, M.A.; Sadrkhanlou, R.; Ahmadi, A.; Shojaei-Sadee, H.; Mohammadirad, A.; Salehnia, A.; Abdollahi, M. Protection of cyclophosphamide-induced toxicity in reproductive tract histology, sperm characteristics, and DNA damage by an herbal source; evidence for role of free-radical toxic stress. Hum. Exp. Toxicol. 2008, 27, 901–910. [Google Scholar] [CrossRef]
- Abarikwu, S.; Ekor, M.; Osobu, D.; Otuechere, C.; Monwuba, K. Rutin ameliorates cyclophosphamide-induced reproductive toxicity in male rats. Toxicol. Int. 2012, 19, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.-P.; Mei, X.-T.; Wang, Y.; Zheng, Y.-P.; Xue, Y.-F.; Xu, D.-H. Zn(II)–curcumin protects against oxidative stress, deleterious changes in sperm parameters and histological alterations in a male mouse model of cyclophosphamide-induced reproductive damage. Environ. Toxicol. Pharmacol. 2015, 39, 515–524. [Google Scholar] [CrossRef] [PubMed]
- McDonald, G.B.; Slattery, J.T.; Bouvier, M.E.; Ren, S.; Batchelder, A.L.; Kalhorn, T.F.; Schoch, H.G.; Anasetti, C.; Gooley, T. Cyclophosphamide metabolism, liver toxicity, and mortality following hematopoietic stem cell transplantation. Blood 2003, 101, 2043–2048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shokrzadeh, M.; Ahmadi, A.; Naghshvar, F.; Chabra, A.; Jafarinejhad, M. Prophylactic Efficacy of Melatonin on Cyclophosphamide-Induced Liver Toxicity in Mice. BioMed Res. Int. 2014, 2014, 470425. [Google Scholar] [CrossRef] [Green Version]
- Roy, P.; Yu, L.J.; Crespi, C.L.; Waxman, D.J. Development of a substrate-activity based approach to identify the major human liver P-450 catalysts of cyclophosphamide and ifosfamide activation based on cDNA-expressed activities and liver microsomal P-450 profiles. Drug. Metab. Dispos. 1999, 27, 655–666. [Google Scholar]
- Korkmaz, A.; Topal, T.; Oter, S. Pathophysiological aspects of cyclophosphamide and ifosfamide induced hemorrhagic cystitis; implication of reactive oxygen and nitrogen species as well as PARP activation. Cell Biol. Toxicol. 2007, 23, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.R.; Pearson, A.E. The clinical use of mutagenic anticancer drugs. Mutat. Res. Mol. Mech. Mutagen. 1996, 355, 1–12. [Google Scholar] [CrossRef]
- Silva, A.F.R.; Resende, D.; Monteiro, M.; Coimbra, M.A.; Silva, A.M.S.; Cardoso, S.M. Application of Hydroxytyrosol in the Functional Foods Field: From Ingredient to Dietary Supplements. Antioxidants 2020, 9, 1246. [Google Scholar] [CrossRef]
- Chen, W.-W.; Zhang, X.; Huang, W.-J. Role of neuroinflammation in neurodegenerative diseases (Review). Mol. Med. Rep. 2016, 13, 3391–3396. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, T.S.S.D.A.; Macedo, P.C.D.O.; Converti, A.; De Lima, A.N. The Use of Euterpe oleracea Mart. As a New Perspective for Disease Treatment and Prevention. Biomolecules 2020, 10, 813. [Google Scholar] [CrossRef]
- Denzer, I.; Münch, G.; Friedland, K. Modulation of mitochondrial dysfunction in neurodegenerative diseases via activation of nuclear factor erythroid-2-related factor 2 by food-derived compounds. Pharmacol. Res. 2016, 103, 80–94. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, G.G. Molecular Pathological Classification of Neurodegenerative Diseases: Turning towards Precision Medicine. Int. J. Mol. Sci. 2016, 17, 189. [Google Scholar] [CrossRef] [Green Version]
- Lewerenz, J.; Maher, P. Chronic Glutamate Toxicity in Neurodegenerative Diseases—What is the Evidence? Front. Neurosci. 2015, 9, 469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, A.K.; Andreazza, A.C.; da Silva, T.M.; Boligon, A.A.; Nascimento, V.D.; Scola, G.; Duong, A.; Cadoná, F.C.; Ribeiro, E.E.; da Cruz, I.B.M. Neuroprotective Effects of Açaí (Euterpe oleracea Mart.) against Rotenone In Vitro Exposure. Oxidative Med. Cell. Longev. 2016, 2016, 8940850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torma, P.D.C.M.R.; Brasil, A.V.S.; Carvalho, A.V.; Jablonski, A.; Rabelo, T.K.; Moreira, J.C.F.; Gelain, D.P.; Flôres, S.H.; Augusti, P.R.; Rios, A.D.O. Hydroethanolic extracts from different genotypes of açaí (Euterpe oleracea) presented antioxidant potential and protected human neuron-like cells (SH-SY5Y). Food Chem. 2017, 222, 94–104. [Google Scholar] [CrossRef]
- Xie, C.; Kang, J.; Li, Z.; Schauss, A.; Badger, T.M.; Nagarajan, S.; Wu, T.; Wu, X. The açaí flavonoid velutin is a potent anti-inflammatory agent: Blockade of LPS-mediated TNF-α and IL-6 production through inhibiting NF-κB activation and MAPK pathway. J. Nutr. Biochem. 2012, 23, 1184–1191. [Google Scholar] [CrossRef] [PubMed]
- Ostardo, E.; Impellizzeri, D.; Cervigni, M.; Porru, D.; Sommariva, M.; Cordaro, M.; Siracusa, R.; Fusco, R.; Gugliandolo, E.; Crupi, R.; et al. Adelmidrol + sodium hyaluronate in IC/BPS or conditions associated to chronic urothelial inflammation. A translational study. Pharmacol. Res. 2018, 134, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Elangovan, N.; Chiou, T.-J.; Tzeng, W.-F.; Chu, S.-T. Cyclophosphamide treatment causes impairment of sperm and its fertilizing ability in mice. Toxicology 2006, 222, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Ilbey, Y.O.; Ozbek, E.; Simsek, A.; Otunctemur, A.; Cekmen, M.; Somay, A. Potential chemoprotective effect of melatonin in cyclophosphamide- and cisplatin-induced testicular damage in rats. Fertil. Steril. 2009, 92, 1124–1132. [Google Scholar] [CrossRef]
- Ghosh, D.; Das, U.B.; Misro, M. Protective Role of α-tocopherol-succinate (Provitamin-E) in Cyclophosphamide Induced Testicular Gametogenic and Steroidogenic Disorders: A Correlative Approach to Oxidative Stress. Free Radic. Res. 2002, 36, 1209–1218. [Google Scholar] [CrossRef] [PubMed]
- Matés, J. Effects of antioxidant enzymes in the molecular control of reactive oxygen species toxicology. Toxicology 2000, 153, 83–104. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Trocchia, S.; Abdel-Gawad, F.; Ciarcia, G. Roles of Reactive Oxygen Species in the Spermatogenesis Regulation. Front. Endocrinol. 2014, 5, 56. [Google Scholar] [CrossRef] [Green Version]
- Leri, M.; Scuto, M.; Ontario, M.L.; Calabrese, V.; Calabrese, E.J.; Bucciantini, M.; Stefani, M. Healthy Effects of Plant Polyphenols: Molecular Mechanisms. Int. J. Mol. Sci. 2020, 21, 1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montoya, T.; Soto, M.A.; Castejón, M.L.; Rosillo, M.Á.; Sánchez-Hidalgo, M.; Begines, P.; Fernández-Bolaños, J.G.; Alarcón-De-La-Lastra, C. Peracetylated hydroxytyrosol, a new hydroxytyrosol derivate, attenuates LPS-induced inflammatory response in murine peritoneal macrophages via regulation of non-canonical inflammasome, Nrf2/HO1 and JAK/STAT signaling pathways. J. Nutr. Biochem. 2018, 57, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Maremanda, K.; Khan, S.; Jena, G. Zinc protects cyclophosphamide-induced testicular damage in rat: Involvement of metallothionein, tesmin and Nrf. Biochem. Biophys. Res. Commun. 2014, 445, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Türk, G.; Ateşşahin, A.; Sönmez, M.; Yüce, A.; Ceribasi, A.O. Lycopene protects against cyclosporine A-induced testicular toxicity in rats. Theriogenology 2007, 67, 778–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceribasi, A.O.; Türk, G.; Sönmez, M.; Sakin, F.; Ateşşahin, A. Toxic Effect of Cyclophosphamide on Sperm Morphology, Testicular Histology and Blood Oxidant-Antioxidant Balance, and Protective Roles of Lycopene and Ellagic Acid. Basic Clin. Pharmacol. Toxicol. 2010, 107, 730–736. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siracusa, R.; D’Amico, R.; Fusco, R.; Impellizzeri, D.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Interdonato, L.; Cordaro, M.; Cuzzocrea, S.; et al. Açai Berry Attenuates Cyclophosphamide-Induced Damage in Genitourinary Axis-Modulating Nrf-2/HO-1 Pathways. Antioxidants 2022, 11, 2355. https://doi.org/10.3390/antiox11122355
Siracusa R, D’Amico R, Fusco R, Impellizzeri D, Peritore AF, Gugliandolo E, Crupi R, Interdonato L, Cordaro M, Cuzzocrea S, et al. Açai Berry Attenuates Cyclophosphamide-Induced Damage in Genitourinary Axis-Modulating Nrf-2/HO-1 Pathways. Antioxidants. 2022; 11(12):2355. https://doi.org/10.3390/antiox11122355
Chicago/Turabian StyleSiracusa, Rosalba, Ramona D’Amico, Roberta Fusco, Daniela Impellizzeri, Alessio Filippo Peritore, Enrico Gugliandolo, Rosalia Crupi, Livia Interdonato, Marika Cordaro, Salvatore Cuzzocrea, and et al. 2022. "Açai Berry Attenuates Cyclophosphamide-Induced Damage in Genitourinary Axis-Modulating Nrf-2/HO-1 Pathways" Antioxidants 11, no. 12: 2355. https://doi.org/10.3390/antiox11122355