
Blue Light Supplemented at Intervals in Long-Day Conditions Intervenes in Photoperiodic Flowering, Photosynthesis, and Antioxidant Properties in Chrysanthemums



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
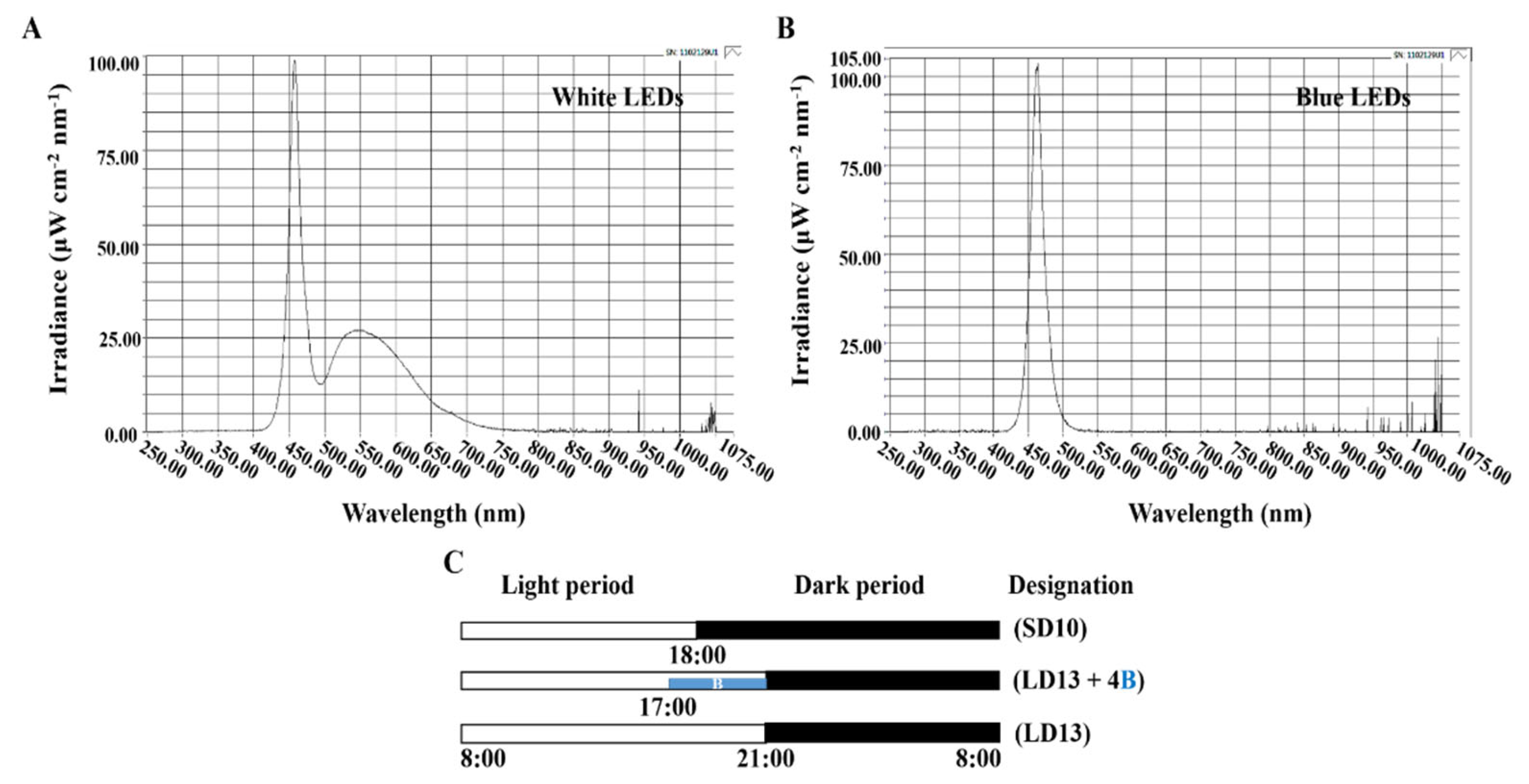
2.2. Photoperiodic Light Treatments
2.3. Measurements of Growth Parameters
2.4. Chlorophyll Content
2.5. Measurements of Photosynthesis and Chlorophyll Fluorescence
2.6. Accumulation of Carbohydrates and Soluble Proteins
2.7. Enzyme Activities
2.8. Extraction and Colorimetric Assays of Non-Enzymatic Antioxidants
2.9. Real-Time Quantitative PCR Verification
2.10. Principal Component Analysis
2.11. Statistical Analysis
3. Results
3.1. Flowering and Growth Parameters
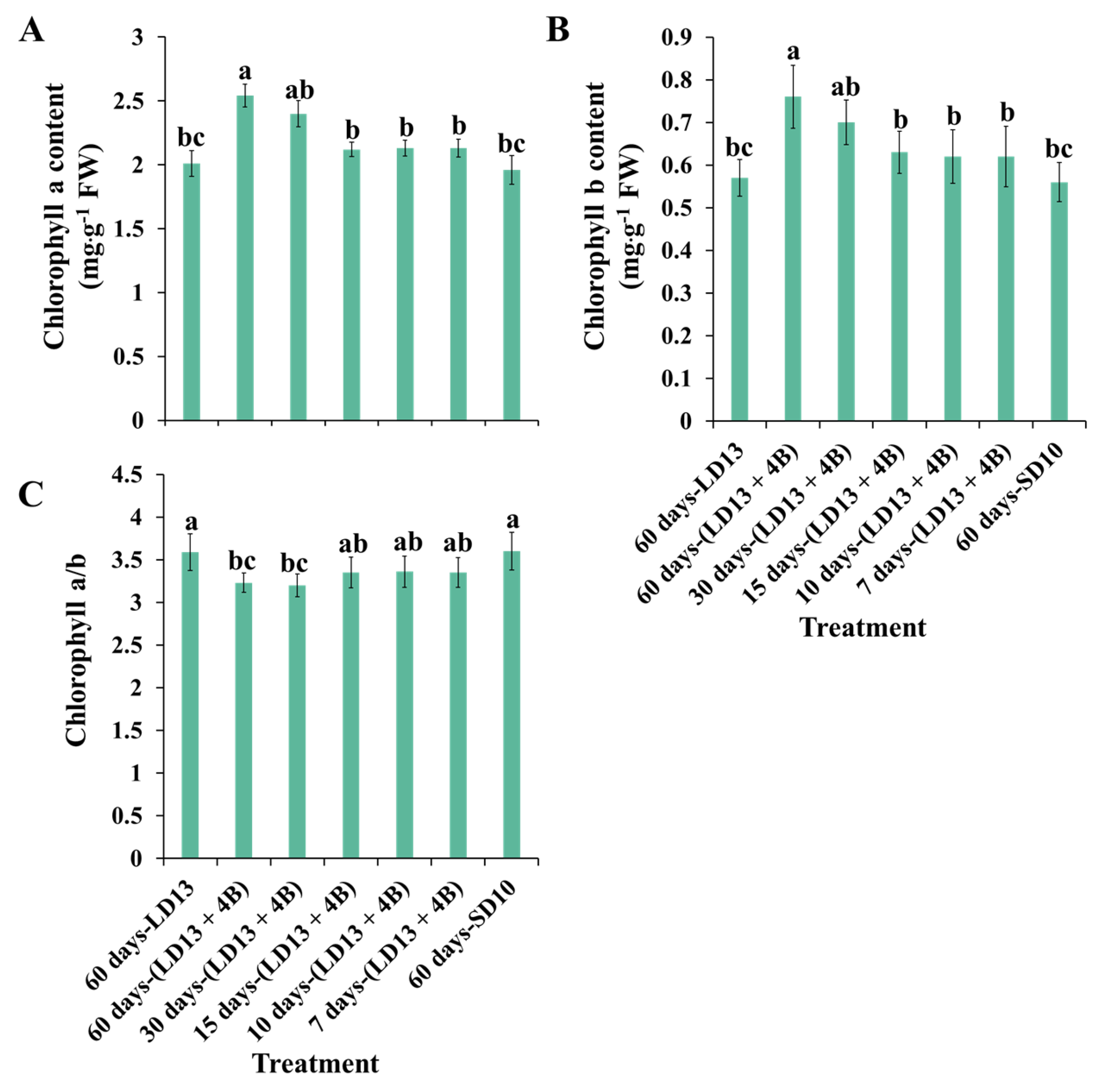
3.2. Chlorophyll Content
3.3. Photosynthetic Index and Chlorophyll Fluorescence
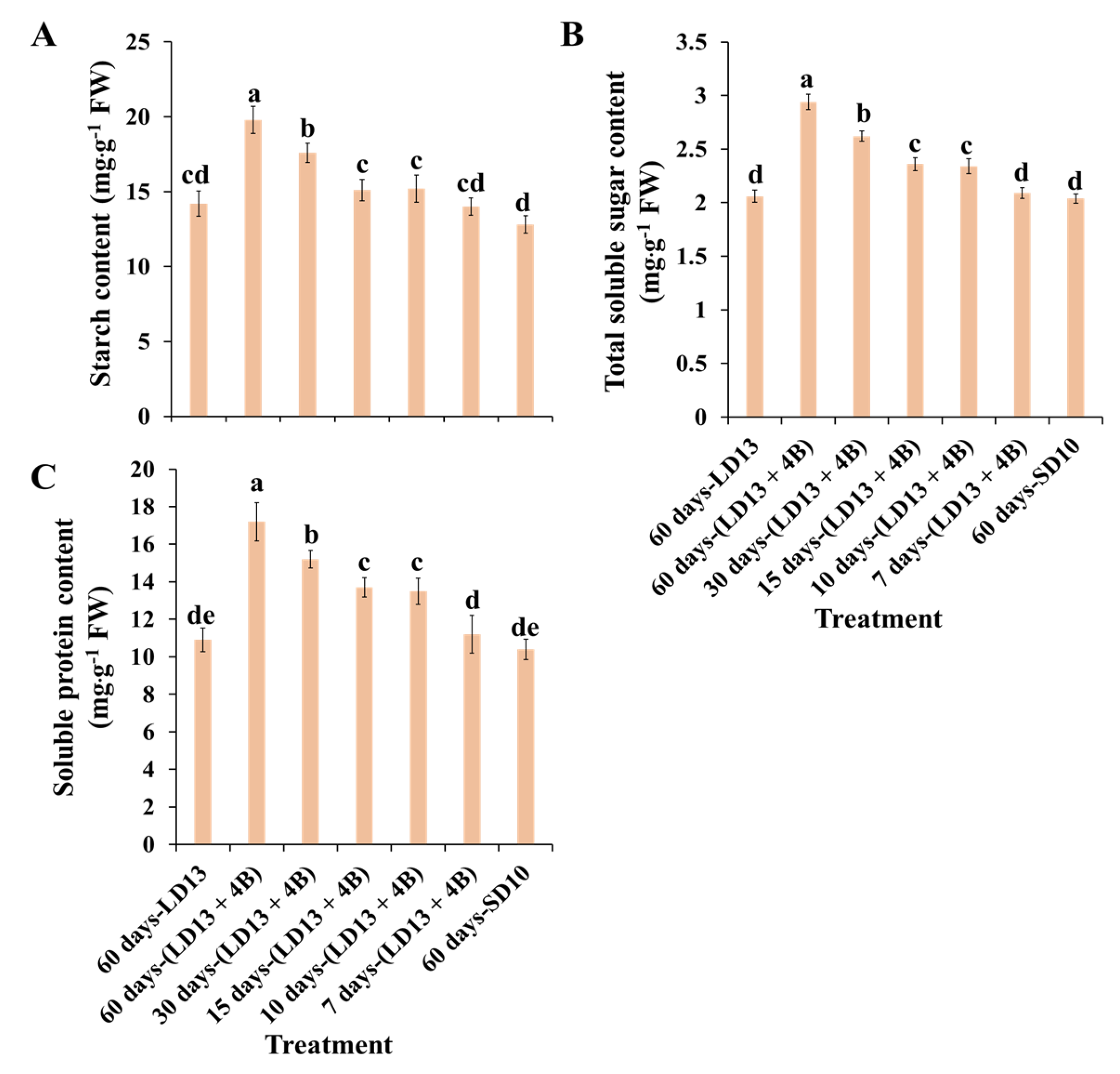
3.4. Contents of Carbohydrates and Soluble Proteins
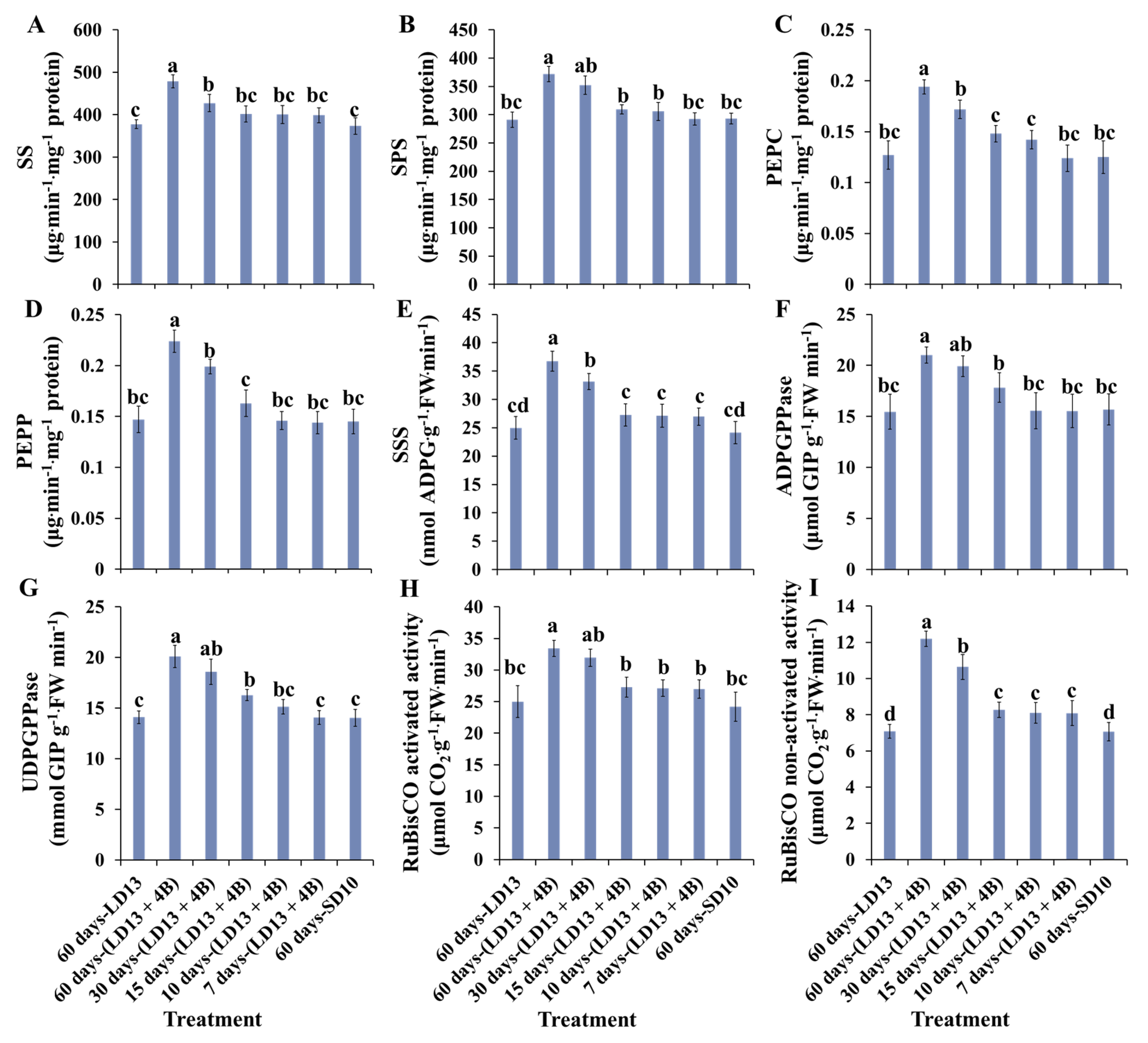
3.5. Enzymatic Activities of Carbohydrate Synthesis- or Photosynthesis-Related Enzymes
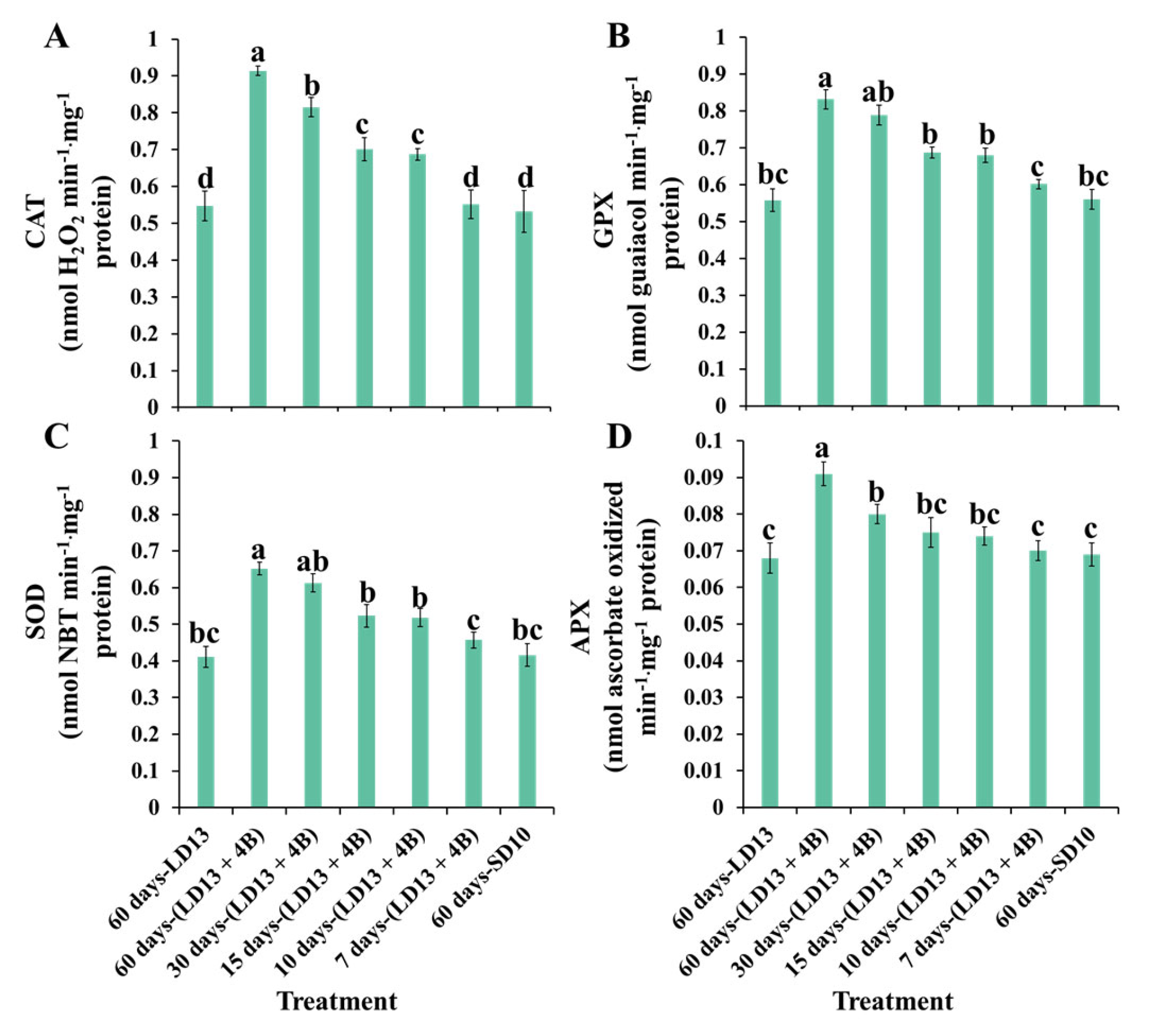
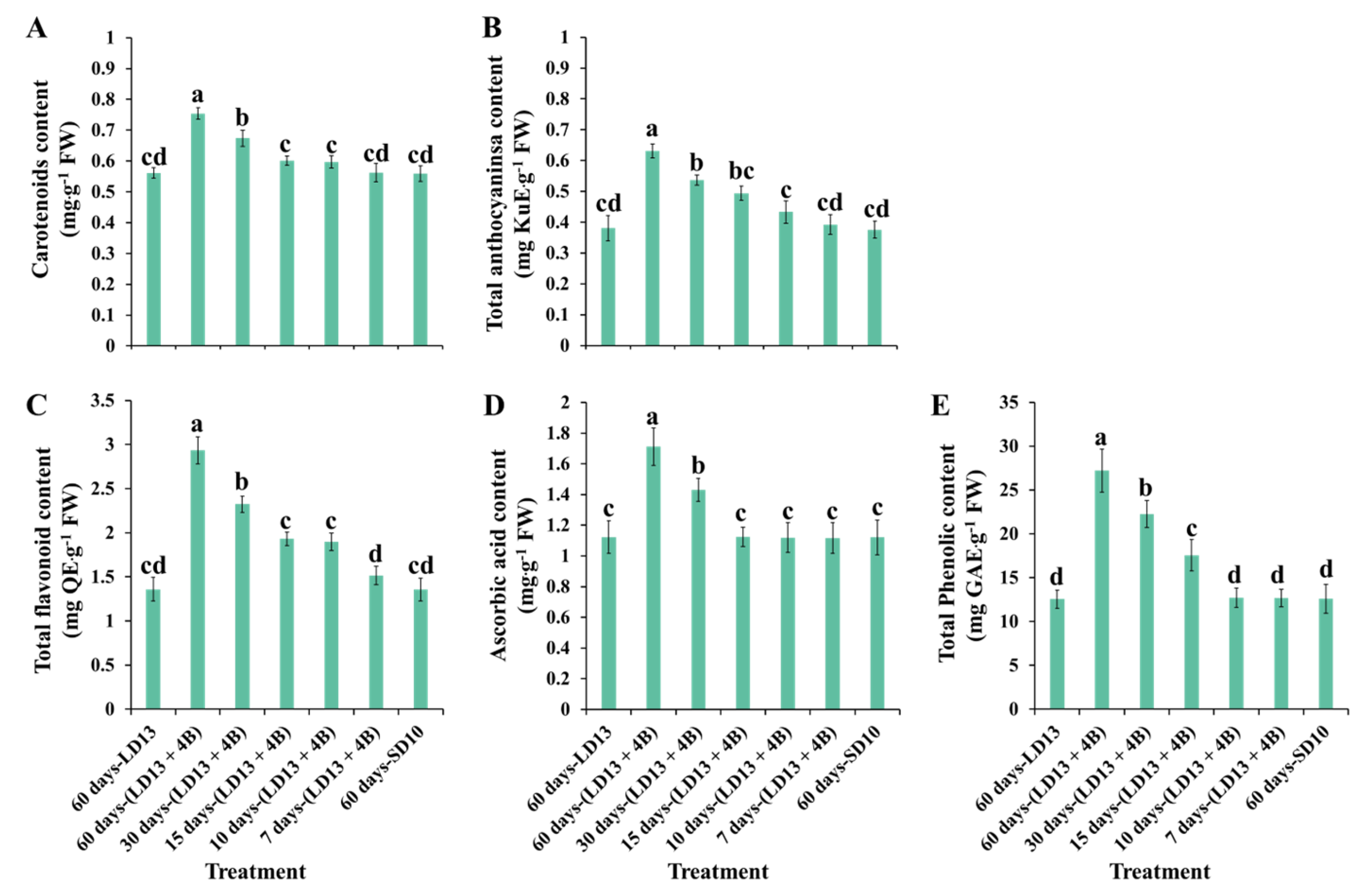
3.6. Antioxidant Ability
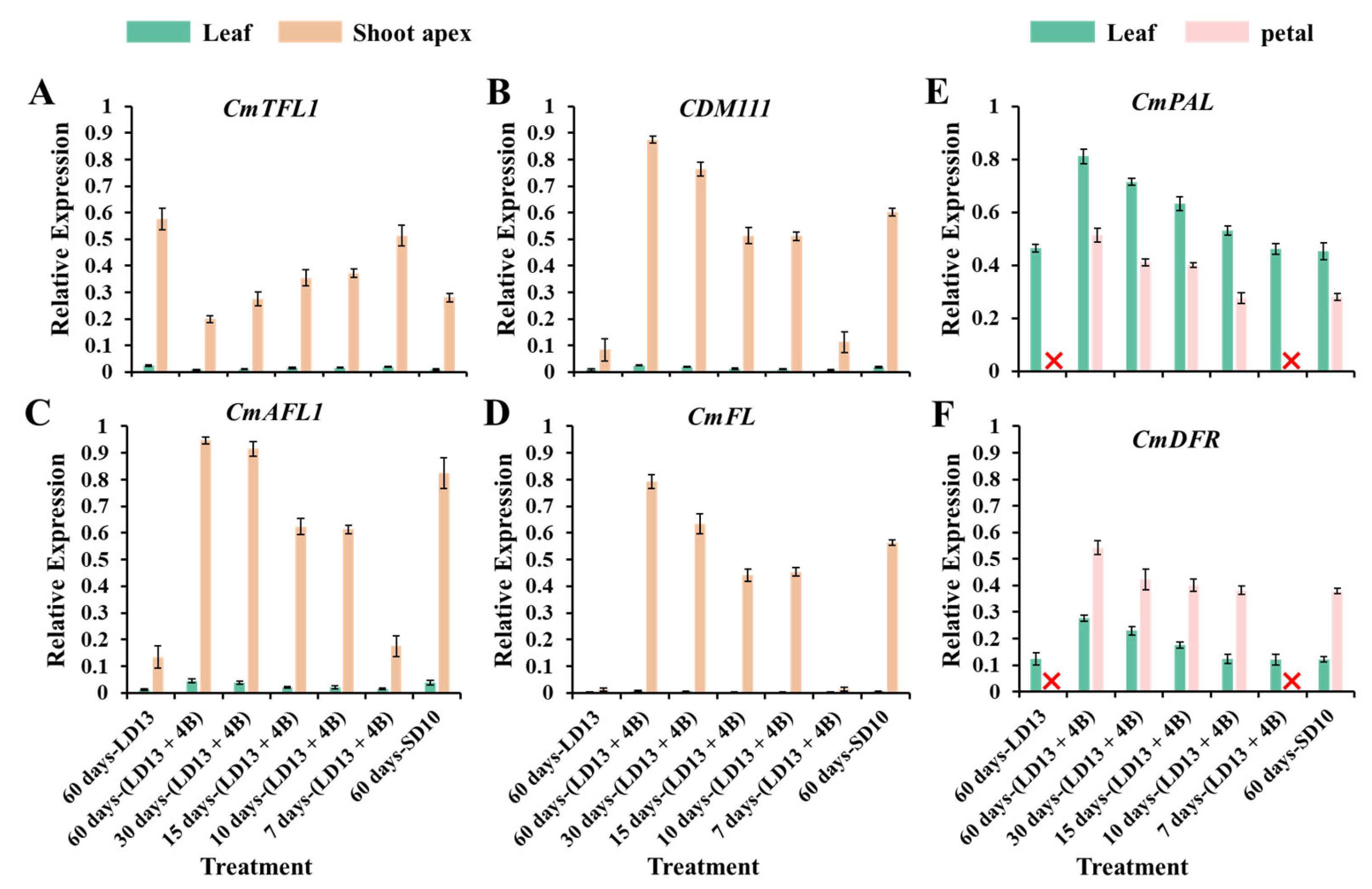
3.7. Gene Expression
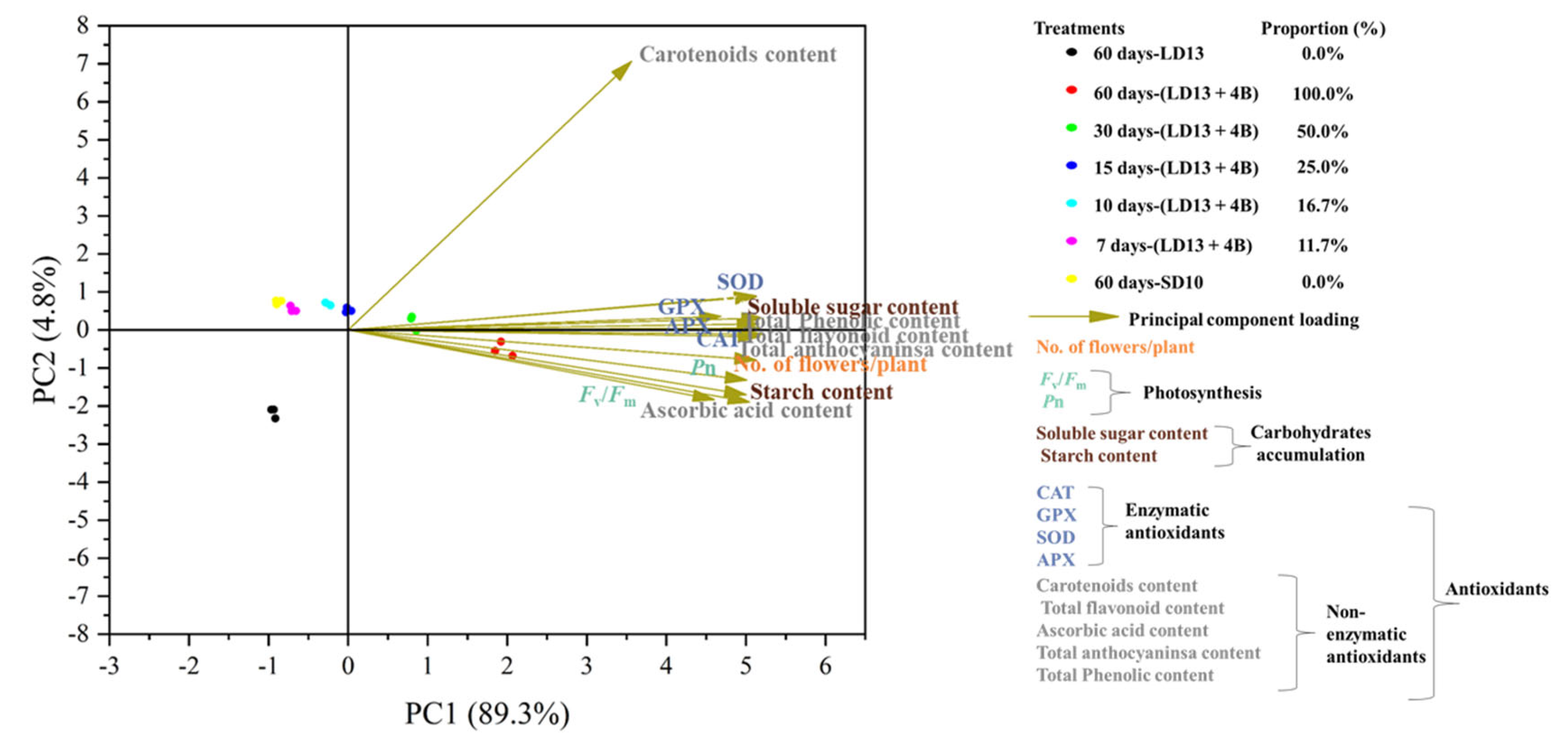
3.8. Principal Component Analysis
4. Discussion
4.1. The Chlorophyll Content, Photosynthetic Efficiency, Related Enzymatic Activities, and Accumulation of Nutrients in Response to Different Proportions of S-BL Days
4.2. The Branching and Flowering in Response to Different Proportions of S-BL Days
4.3. The Antioxidant Property and the Expression of Antioxidant Synthesis-Related Genes in Response to Different Proportions of S-BL Days
4.4. The Positive Interactive Relations among the Blue Light-Mediated-Photosynthetic Carbon Assimilation, -Antioxidant Ability, and -Flowering
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thomas, B.; Vince-Prue, D. Photoperiodic control of flower initiation: Some general principles. In Photoperiodism in Plants, 2nd ed.; Academic Press: San Diego, CA, USA, 1997; pp. 3–28. [Google Scholar]
- Kozai, T. Current status of plant factories with artificial lighting (PFALs) and smart PFALs. In Smart Plant Factory; Springer: Berlin/Heidelberg, Germany, 2018; pp. 3–13. [Google Scholar]
- Bantis, F.; Smirnakou, S.; Ouzounis, T.; Koukounaras, A.; Ntagkas, N.; Radoglou, K. Current status and recent achievements in the field of horticulture with the use of light-emitting diodes (LEDs). Sci. Hortic. 2018, 235, 437–451. [Google Scholar]
- Kigel, J.; Cosgrove, D.J. Photoinhibition of stem elongation by blue and red light: Effects on hydraulic and cell wall properties. Plant Physiol. 1991, 95, 1049–1056. [Google Scholar] [CrossRef] [Green Version]
- CASAL, J.J.; SMITH, H. Effects of blue light pretreatments on internode extension growth in mustard seedlings after the transition to darkness: Analysis of the interaction with phytochrome. J. Exp. Bot. 1989, 40, 893–899. [Google Scholar] [CrossRef]
- Laskowski, M.J.; Briggs, W.R. Regulation of pea epicotyl elongation by blue light: Fluence-response relationships and growth distribution. Plant Physiol. 1989, 89, 293–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warpeha, K.M.; Kaufman, L.S. Blue-light regulation of epicotyl elongation in Pisum sativum. Plant Physiol. 1989, 89, 544–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, M.; Borthwick, H.; Hendricks, S. Effects of radiation on flowering of Hyoscyamus niger. Am. J. Bot. 1967, 54, 1241–1249. [Google Scholar] [CrossRef]
- Bagnall, D.J.; King, R.W.; Hangarter, R.P. Blue-light promotion of flowering is absent in hy4 mutants of Arabidopsis. Planta 1996, 200, 278–280. [Google Scholar] [CrossRef]
- Park, Y.; Runkle, E.S. Spectral effects of light-emitting diodes on plant growth, visual color quality, and photosynthetic photon efficacy: White versus blue plus red radiation. PLoS ONE 2018, 13, e0202386. [Google Scholar] [CrossRef] [Green Version]
- Runkle, E.S.; Heins, R.D. Specific functions of red, far red, and blue light in flowering and stem extension of long-day plants. J. Am. Soc. Hortic. Sci. 2001, 126, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.G.; Jeong, B.R. How supplementary or night-interrupting low-intensity blue light affects the flower induction in chrysanthemum, a qualitative short-day plant. Plants 2020, 9, 1694. [Google Scholar] [CrossRef]
- Yang, J.; Song, J.; Jeong, B.R. The flowering of SDP chrysanthemum in response to intensity of supplemental or night-interruptional blue light is modulated by both photosynthetic carbon assimilation and photoreceptor-mediated regulation. Front. Plant Sci. 2022, 13, 981143. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Song, J.; Jeong, B.R. Low-intensity blue light supplemented during photoperiod in controlled environment induces flowering and antioxidant production in kalanchoe. Antioxidants 2022, 11, 811. [Google Scholar] [CrossRef] [PubMed]
- Koch, K. Sucrose metabolism: Regulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr. Opin. Plant Biol. 2004, 7, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Gangadhar, B.H.; Mishra, R.K.; Pandian, G.; Park, S.W. Comparative study of color, pungency, and biochemical composition in chili pepper (Capsicum annuum) under different light-emitting diode treatments. HortScience 2012, 47, 1729–1735. [Google Scholar] [CrossRef]
- Urbonavičiūtė, A.; Pinho, P.; Samuolienė, G.; Duchovskis, P.; Vitta, P.; Stonkus, A.; Tamulaitis, G.; Žukauskas, A.; Halonen, L. Effect of short-wavelength light on lettuce growth and nutritional quality. Sodininkystė Daržininkystė 2007, 26, 157–165. [Google Scholar]
- Samuolienė, G.; Brazaitytė, A.; Urbonavičiūtė, A.; Šabajevienė, G.; Duchovskis, P. The effect of red and blue light component on the growth and development of frigo strawberries. Zemdirbyste 2010, 97, 99–104. [Google Scholar]
- Avercheva, O.; Berkovich, Y.A.; Smolyanina, S.; Bassarskaya, E.; Pogosyan, S.; Ptushenko, V.; Erokhin, A.; Zhigalova, T. Biochemical, photosynthetic and productive parameters of chinese cabbage grown under blue–red LED assembly designed for space agriculture. Adv. Space Res. 2014, 53, 1574–1581. [Google Scholar] [CrossRef]
- Chen, X.L.; Guo, W.Z.; Xue, X.Z.; Wang, L.C.; Qiao, X.J. Growth and quality responses of ‘green oak leaf’ lettuce as affected by monochromic or mixed radiation provided by fluorescent lamp (FL) and light-emitting diode (LED). Sci. Hortic. 2014, 172, 168–175. [Google Scholar] [CrossRef]
- Torres, M.A.; Jones, J.D.; Dangl, J.L. Reactive oxygen species signaling in response to pathogens. Plant Physiol. 2006, 141, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Ashry, N.A.; Mohamed, H.I. Impact of secondary metabolites and related enzymes in flax resistance and or susceptibility to powdery mildew. World J. Agric. Sci 2011, 7, 78–85. [Google Scholar]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Kook, H.; Jang, Y.; Lee, W.; Kamala-Kannan, S.; Chae, J.; Lee, K. The effect of blue-light-emitting diodes on antioxidant properties and resistance to Botrytis cinerea in tomato. J. Plant Pathol. Microbiol. 2013, 4, 203. [Google Scholar]
- Samuolienė, G.; Brazaitytė, A.; Jankauskienė, J.; Viršilė, A.; Sirtautas, R.; Novičkovas, A.; Sakalauskienė, S.; Sakalauskaitė, J.; Duchovskis, P. Led irradiance level affects growth and nutritional quality of Brassica microgreens. Cent. Eur. J. Biol. 2013, 8, 1241–1249. [Google Scholar] [CrossRef]
- Son, K.H.; Oh, M.M. Leaf shape, growth, and antioxidant phenolic compounds of two lettuce cultivars grown under various combinations of blue and red light-emitting diodes. HortScience 2013, 48, 988–995. [Google Scholar] [CrossRef]
- Ren, J.; Guo, S.; Xu, C.; Yang, C.; Ai, W.; Tang, Y.; Qin, L. Effects of different carbon dioxide and LED lighting levels on the anti-oxidative capabilities of Gynura bicolor DC. Adv. Space Res. 2014, 53, 353–361. [Google Scholar] [CrossRef]
- Ouzounis, T.; Fretté, X.; Rosenqvist, E.; Ottosen, C.-O. Spectral effects of supplementary lighting on the secondary metabolites in roses, chrysanthemums, and campanulas. J. Plant Physiol. 2014, 171, 1491–1499. [Google Scholar] [CrossRef]
- Kim, E.Y.; Park, S.; Park, B.J.; Lee, Y.; Oh, M.M. Growth and antioxidant phenolic compounds in cherry tomato seedlings grown under monochromatic light-emitting diodes. Hortic. Environ. Biotechnol. 2014, 55, 506–513. [Google Scholar] [CrossRef]
- Wu, M.C.; Hou, C.Y.; Jiang, C.M.; Wang, Y.T.; Wang, C.Y.; Chen, H.H.; Chang, H.M. A novel approach of LED light radiation improves the antioxidant activity of pea seedlings. Food Chem. 2007, 101, 1753–1758. [Google Scholar] [CrossRef]
- Walters, R.G.; Ibrahim, D.G.; Horton, P.; Kruger, N.J. A mutant of Arabidopsis lacking the triose-phosphate/phosphate translocator reveals metabolic regulation of starch breakdown in the light. Plant Physiol. 2004, 135, 891–906. [Google Scholar] [CrossRef] [Green Version]
- Eckardt, N.A.; Snyder, G.W.; Portis Jr, A.R.; Ogren, W.L. Growth and photosynthesis under high and low irradiance of Arabidopsis thaliana antisense mutants with reduced Ribulose-1, 5-Bisphosphate Carboxylase/Oxygenase Activase content. Plant Physiol. 1997, 113, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Sami, F.; Yusuf, M.; Faizan, M.; Faraz, A.; Hayat, S. Role of sugars under abiotic stress. Plant Physiol. Biochem. 2016, 109, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Van den Ende, W.; Peshev, D. Sugars as antioxidants in plants. In Crop Improvement under Adverse Conditions; Springer: Berlin/Heidelberg, Germany, 2013; pp. 285–307. [Google Scholar]
- Keunen, E.; Peshev, D.; Vangronsveld, J.; Van Den Ende, W.; Cuypers, A. Plant sugars are crucial players in the oxidative challenge during abiotic stress: Extending the traditional concept. Plant Cell Environ. 2013, 36, 1242–1255. [Google Scholar] [CrossRef] [PubMed]
- Barth, C.; De Tullio, M.; Conklin, P.L. The role of ascorbic acid in the control of flowering time and the onset of senescence. J. Exp. Bot. 2006, 57, 1657–1665. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4. 3.1–F4. 3.8. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. BBA-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Roháček, K. Chlorophyll fluorescence parameters: The definitions, photosynthetic meaning, and mutual relationships. Photosynthetica 2002, 40, 13–29. [Google Scholar] [CrossRef]
- Loewus, F.A. Improvement in anthrone method for determination of carbohydrates. Anal. Chem. 1952, 24, 219. [Google Scholar] [CrossRef]
- Yemm, E.; Willis, A. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Doehlert, D.C.; Kuo, T.M.; Felker, F.C. Enzymes of sucrose and hexose metabolism in developing kernels of two inbreds of maize. Plant Physiol. 1988, 86, 1013–1019. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.S.; Cao, X.; Xu, S.; Zhu, Q.; Song, P. Studies on the relationship between the grain sink strength and its starch accumulation in rice (O. sativa). Acta Agron. Sin 1994, 20, 685–691. [Google Scholar]
- Yang, L.T.; Chen, L.S.; Peng, H.Y.; Guo, P.; Wang, P.; Ma, C.L. Organic acid metabolism in Citrus grandis leaves and roots is differently affected by nitric oxide and aluminum interactions. Sci. Hortic. 2012, 133, 40–46. [Google Scholar] [CrossRef]
- Feng, L.; Raza, M.A.; Li, Z.; Chen, Y.; Khalid, M.H.B.; Du, J.; Liu, W.; Wu, X.; Song, C.; Yu, L. The influence of light intensity and leaf movement on photosynthesis characteristics and carbon balance of soybean. Front. Plant Sci. 2019, 9, 1952. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase. In Methods of Enzymatic Analysis; Elsevier: Amsterdam, The Netherlands, 1974; pp. 673–684. [Google Scholar]
- Castillo, F.J.; Penel, C.; Greppin, H. Peroxidase release induced by ozone in sedum album leaves: Involvement of Ca2+. Plant Physiol. 1984, 74, 846–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becana, M.; Aparicio-Tejo, P.; Irigoyen, J.J.; Sanchez-Diaz, M. Some enzymes of hydrogen peroxide metabolism in leaves and root nodules of Medicago sativa. Plant Physiol. 1986, 82, 1169–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Andre, C.M.; Ghislain, M.; Bertin, P.; Oufir, M.; del Rosario Herrera, M.; Hoffmann, L.; Hausman, J.-F.; Larondelle, Y.; Evers, D. Andean potato cultivars (Solanum tuberosum L.) as a source of antioxidant and mineral micronutrients. J. Agr. Food Chem. 2007, 55, 366–378. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B. A Guide to Carotenoid Analysis in Foods; ILSI Press: Washington, DC, USA, 2001; Volume 71. [Google Scholar]
- Awika, J.M.; Rooney, L.W.; Waniska, R.D. Anthocyanins from black sorghum and their antioxidant properties. Food Chem. 2005, 90, 293–301. [Google Scholar] [CrossRef]
- Nielsen, I.L.F.; Haren, G.R.; Magnussen, E.L.; Dragsted, L.O.; Rasmussen, S.E. Quantification of anthocyanins in commercial black currant juices by simple high-performance liquid chromatography. Investigation of their pH stability and antioxidative potency. J. Agric. Food Chem. 2003, 51, 5861–5866. [Google Scholar] [CrossRef]
- Schultze, M. Methods of vitamin assay. Prepared and edited by the association of vitamin chemists. J. Phys. Chem. 1947, 51, 1452–1453. [Google Scholar] [CrossRef]
- Perva-Uzunalić, A.; Škerget, M.; Knez, Ž.; Weinreich, B.; Otto, F.; Grüner, S. Extraction of active ingredients from green tea (Camellia sinensis): Extraction efficiency of major catechins and caffeine. Food Chem. 2006, 96, 597–605. [Google Scholar] [CrossRef]
- Proestos, C.; Boziaris, I.; Nychas, G.-J.; Komaitis, M. Analysis of flavonoids and phenolic acids in Greek aromatic plants: Investigation of their antioxidant capacity and antimicrobial activity. Food Chem. 2006, 95, 664–671. [Google Scholar] [CrossRef]
- Siddhuraju, P.; Becker, K. Antioxidant properties of various solvent extracts of total phenolic constituents from three different agroclimatic origins of drumstick tree (Moringa oleifera Lam.) leaves. J. Agric. Food Chem. 2003, 51, 2144–2155. [Google Scholar] [CrossRef]
- Kumaran, A. Antioxidant and free radical scavenging activity of an aqueous extract of Coleus aromaticus. Food Chem. 2006, 97, 109–114. [Google Scholar] [CrossRef]
- Atanassova, M.; Georgieva, S.; Ivancheva, K. Total phenolic and total flavonoid contents, antioxidant capacity and biological contaminants in medicinal herbs. J. Univ. Chem. Technol. Metall. 2011, 46, 81–88. [Google Scholar]
- Shahidi, F.; Naczk, M.; Griffiths, W. Food phenolics: Sources, chemistry, effects, applications. Trends Food Sci. Technol. 1996, 7, 243. [Google Scholar]
- Waterhouse, A.L. Wine phenolics. Ann. N. Y. Acad. Sci. 2002, 957, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−△△ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Higuchi, Y.; Sumitomo, K.; Oda, A.; Shimizu, H.; Hisamatsu, T. Day light quality affects the night-break response in the short-day plant chrysanthemum, suggesting differential phytochrome-mediated regulation of flowering. J. Plant Physiol. 2012, 169, 1789–1796. [Google Scholar] [CrossRef]
- Gu, C.; Chen, S.; Liu, Z.; Shan, H.; Luo, H.; Guan, Z.; Chen, F. Reference gene selection for quantitative real-time PCR in chrysanthemum subjected to biotic and abiotic stress. Mol. Biotechnol. 2011, 49, 192–197. [Google Scholar] [CrossRef]
- Higuchi, Y.; Narumi, T.; Oda, A.; Nakano, Y.; Sumitomo, K.; Fukai, S.; Hisamatsu, T. The gated induction system of a systemic floral inhibitor, antiflorigen, determines obligate short-day flowering in chrysanthemums. Proc. Natl. Acad. Sci. USA 2013, 110, 17137–17142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shchennikova, A.V.; Shulga, O.A.; Immink, R.; Skryabin, K.G.; Angenent, G.C. Identification and characterization of four chrysanthemum MADS-box genes, belonging to the APETALA1/FRUITFULL and SEPALLATA3 subfamilies. Plant Physiol. 2004, 134, 1632–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Niki, T.; Nishijima, T.; Douzono, M.; Koshioka, M.; Hisamatsu, T. Roles of CmFL, CmAFL1, and CmSOC1 in the transition from vegetative to reproductive growth in Chrysanthemum morifolium Ramat. J. Hortic. Sci. Biotechnol. 2009, 84, 447–453. [Google Scholar] [CrossRef]
- An, C.; Zhang, Y.; Zhang, W.W.; Zhang, T.; Jiang, J.; Chen, F. Cloning and expression analysis of CmPAL gene in Chrysanthemum morifolium. J. Nanjing Agric. Univ. 2019, 42, 73–80. [Google Scholar]
- Chen, L.; Wang, T.; Guo, Q.S.; Zhang, X.M.; Song, L.S. Cloning of DFR gene and its expression characteristics in flowers of Chrysanthemum morifolium. Zhong Cao Yao (Chin. Tradit. Herb. Drugs) 2016, 47, 1187–1192. [Google Scholar]
- Ciarmiello, L.F.; Piccirillo, P.; Carillo, P.; De Luca, A.; Woodrow, P. Determination of the genetic relatedness of fig (Ficus carica L.) accessions using rapid fingerprint and their agro-morphological characterization. S. Afr. J. Bot. 2015, 97, 40–47. [Google Scholar] [CrossRef]
- Ferchichi, S.; Hessini, K.; Dell’Aversana, E.; D’Amelia, L.; Woodrow, P.; Ciarmiello, L.F.; Fuggi, A.; Carillo, P. Hordeum vulgare and Hordeum maritimum respond to extended salinity stress displaying different temporal accumulation pattern of metabolites. Funct. Plant Biol. 2018, 45, 1096–1109. [Google Scholar] [CrossRef]
- Snowden, M.C.; Cope, K.R.; Bugbee, B. Sensitivity of seven diverse species to blue and green light: Interactions with photon flux. PLoS ONE 2016, 11, e0163121. [Google Scholar] [CrossRef]
- Klem, K.; Gargallo-Garriga, A.; Rattanapichai, W.; Oravec, M.; Holub, P.; Veselá, B.; Sardans, J.; Peñuelas, J.; Urban, O. Distinct morphological, physiological, and biochemical responses to light quality in barley leaves and roots. Front. Plant Sci. 2019, 10, 1026. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, Y.; Zhong, S. Interplay between light and plant hormones in the control of Arabidopsis seedling chlorophyll biosynthesis. Front. Plant Sci. 2017, 8, 1433. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Tanaka, A.; Tsuji, H. Effects of 5-Aminolevulinic Acid on the accumulation of chlorophyll b and apoproteins of the light-harvesting chlorophyll a/b-protein complex of photosystem II. Plant Cell Physiol. 1993, 34, 465–472. [Google Scholar]
- Melis, A.; Harvey, G. Regulation of photosystem stoichiometry, chlorophyll a and chlorophyll b content and relation to chloroplast ultrastructure. Biochim. Biophys. Acta (BBA)-Bioenerg. 1981, 637, 138–145. [Google Scholar] [CrossRef]
- Tanaka, A.; Ito, H.; Tanaka, R.; Tanaka, N.K.; Yoshida, K.; Okada, K. Chlorophyll a oxygenase (CAO) is involved in chlorophyll b formation from chlorophyll a. Proc. Natl. Acad. Sci. USA 1998, 95, 12719–12723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimazaki, K.I.; Doi, M.; Assmann, S.M.; Kinoshita, T. Light regulation of stomatal movement. Annu. Rev. Plant Biol. 2007, 58, 219–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collatz, G.J.; Ball, J.T.; Grivet, C.; Berry, J.A. Physiological and environmental regulation of stomatal conductance, photosynthesis and transpiration: A model that includes a laminar boundary layer. Agric. For. Meteorol. 1991, 54, 107–136. [Google Scholar] [CrossRef]
- Dai, Y.; Shen, Z.; Liu, Y.; Wang, L.; Hannaway, D.; Lu, H. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg. Environ. Exp. Bot. 2009, 65, 177–182. [Google Scholar] [CrossRef]
- Yao, X.; Li, C.; Li, S.; Zhu, Q.; Zhang, H.; Wang, H.; Yu, C.; St Martin, S.K.; Xie, F. Effect of shade on leaf photosynthetic capacity, light-intercepting, electron transfer and energy distribution of soybeans. Plant Growth Regul. 2017, 83, 409–416. [Google Scholar] [CrossRef]
- Richter, G.; Wessel, K. Red light inhibits blue light-induced chloroplast development in cultured plant cells at the mRNA level. Plant Mol. Biol. 1985, 5, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Wu, L.; Chen, J.; Dong, L. Morphological plasticity, photosynthesis and chlorophyll fluorescence of Athyrium pachyphlebium at different shade levels. Photosynthetica 2011, 49, 611–618. [Google Scholar] [CrossRef]
- Wang, H.; Gu, M.; Cui, J.; Shi, K.; Zhou, Y.; Yu, J. Effects of light quality on CO2 assimilation, chlorophyll-fluorescence quenching, expression of calvin cycle genes and carbohydrate accumulation in Cucumis sativus. J. Photochem. Photobiol. B Biol. 2009, 96, 30–37. [Google Scholar] [CrossRef]
- Xu, D.; Gao, W.; Ruan, J. Effects of light quality on plant growth and development. Plant Physiol. J. 2015, 51, 1217–1234. [Google Scholar]
- Hundrieser, J.; Richter, G. Blue light-induced synthesis of ribulosebisphosphate carboxylase in cultured plant cells. Plant Cell Rep. 1982, 1, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Roscher, E.; Zetsche, K. The effects of light quality and intensity on the synthesis of ribulose-1, 5-bisphosphate carboxylase and its mrnas in the green alga Chlorogonium elongatum. Planta 1986, 167, 582–586. [Google Scholar] [CrossRef]
- Kamiya, A.; Miyachi, S. Blue light-induced formation of phosphoenolpyruvate carboxylase in colorless Chlorella mutant cells. Plant Cell Physiol. 1975, 16, 729–736. [Google Scholar]
- Conradt, W.; Ruyters, G. Blue light-effects on enzymes of the carbohydrate metabolism in Chlorella 2. Glyceraldehyde 3-phosphate dehydrogenase (NADP-dependent). In The Blue Light Syndrome; Springer: Berlin/Heidelberg, Germany, 1980; pp. 368–371. [Google Scholar]
- Gao, Y.; Gao, Y.; Wu, Z.; Bu, X.; Fan, M.; Zhang, Q. Characterization of TERMINAL FLOWER 1 homologs CmTFL1c gene from Chrysanthemum morifolium. Plant Mol. Biol. 2019, 99, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.S.; Salchert, K.; Nielsen, K.K. A Terminal Flower1-Like gene from perennial ryegrass involved in floral transition and axillary meristem identity. Plant Physiol. 2001, 125, 1517–1528. [Google Scholar] [CrossRef] [Green Version]
- Ratcliffe, O.J.; Amaya, I.; Vincent, C.A.; Rothstein, S.; Carpenter, R.; Coen, E.S.; Bradley, D.J. A common mechanism controls the life cycle and architecture of plants. Development 1998, 125, 1609–1615. [Google Scholar] [CrossRef]
- Wang, Y.; Pijut, P.M. Isolation and characterization of a TERMINAL FLOWER 1 homolog from Prunus serotina Ehrh. Tree Physiol. 2013, 33, 855–865. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Zhao, Z.; Chen, J.; Hu, X.; Luo, D. A putative CENTRORADIALIS/TERMINAL FLOWER 1-like gene, Ljcen1, plays a role in phase transition in Lotus japonicus. J. Plant Physiol. 2006, 163, 436–444. [Google Scholar] [CrossRef]
- Higuchi, Y.; Hisamatsu, T. CsTFL1, a constitutive local repressor of flowering, modulates floral initiation by antagonising florigen complex activity in chrysanthemum. Plant Sci. 2015, 237, 1–7. [Google Scholar] [CrossRef]
- Li, C.; Fu, Q.; Niu, L.; Luo, L.; Chen, J.; Xu, Z.F. Three TFL1 homologues regulate floral initiation in the biofuel plant Jatropha curcas. Sci. Rep. 2017, 7, 43090. [Google Scholar] [CrossRef] [Green Version]
- Oda, A.; Narumi, T.; Li, T.; Kando, T.; Higuchi, Y.; Sumitomo, K.; Fukai, S.; Hisamatsu, T. CsFTL3, a chrysanthemum flowering locus t-like gene, is a key regulator of photoperiodic flowering in chrysanthemums. J. Exp. Bot. 2012, 63, 1461–1477. [Google Scholar] [CrossRef] [Green Version]
- Komiya, R.; Yokoi, S.; Shimamoto, K. A gene network for long-day flowering activates RFT1 encoding a mobile flowering signal in rice. Development 2009, 136, 3443–3450. [Google Scholar] [CrossRef] [Green Version]
- Kato-Noguchi, H.; Yasuda, Y.; Sasaki, R. Soluble sugar availability of aerobically germinated barley, oat and rice coleoptiles in anoxia. J. Plant Physiol. 2010, 167, 1571–1576. [Google Scholar] [CrossRef]
- Roldán, M.; Gómez-Mena, C.; Ruiz-García, L.; Salinas, J.; Martínez-Zapater, J.M. Sucrose availability on the aerial part of the plant promotes morphogenesis and flowering of Arabidopsis in the dark. Plant J. 1999, 20, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Ohto, M.A.; Onai, K.; Furukawa, Y.; Aoki, E.; Araki, T.; Nakamura, K. Effects of sugar on vegetative development and floral transition in Arabidopsis. Plant Physiol. 2001, 127, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Liu, Y.; Song, L.; Jacobs, D.F.; Du, X.; Ying, Y.; Shao, Q.; Wu, J. Effect of differential light quality on morphology, photosynthesis, and antioxidant enzyme activity in Camptotheca acuminata seedlings. J. Plant Growth Regul. 2017, 36, 148–160. [Google Scholar] [CrossRef]
- Aalifar, M.; Aliniaeifard, S.; Arab, M.; Zare Mehrjerdi, M.; Dianati Daylami, S.; Serek, M.; Woltering, E.; Li, T. Blue light improves vase life of carnation cut flowers through its effect on the antioxidant defense system. Front. Plant Sci. 2020, 11, 511. [Google Scholar] [CrossRef]
- Nishihara, E.; Kondo, K.; Parvez, M.M.; Takahashi, K.; Watanabe, K.; Tanaka, K. Role of 5-aminolevulinic acid (ALA) on active oxygen-scavenging system in NaCl-treated spinach (Spinacia oleracea). J. Plant Physiol. 2003, 160, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Luthria, D.L.; Mukhopadhyay, S.; Krizek, D.T. Content of total phenolics and phenolic acids in tomato (Lycopersicon esculentum Mill.) fruits as influenced by cultivar and solar UV radiation. J. Food Compos. Anal. 2006, 19, 771–777. [Google Scholar] [CrossRef]
- Johkan, M.; Shoji, K.; Goto, F.; Hashida, S.N.; Yoshihara, T. Blue light-emitting diode light irradiation of seedlings improves seedling quality and growth after transplanting in red leaf lettuce. HortScience 2010, 45, 1809–1814. [Google Scholar] [CrossRef] [Green Version]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esnouf, A.; Latrille, É.; Steyer, J.-P.; Helias, A. Representativeness of environmental impact assessment methods regarding life cycle inventories. Sci. Total Environ. 2018, 621, 1264–1271. [Google Scholar] [CrossRef]
- Ohl, S.; Hedrick, S.A.; Chory, J.; Lamb, C.J. Functional properties of a phenylalanine ammonia-lyase promoter from Arabidopsis. Plant Cell 1990, 2, 837–848. [Google Scholar] [PubMed]
- Helariutta, Y.; Elomaa, P.; Kotilainen, M.; Seppänen, P.; Teeri, T.H. Cloning of cDNA coding for dihydroflavonol-4-reductase (DFR) and characterization of dfr expression in the corollas of Gerbera hybrida var. Regina (Compositae). Plant Mol. Biol. 1993, 22, 183–193. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Abbreviation | Proportion 2 (%) |
|---|---|---|
| 1 day (LD13) × 60 1 cycles | 60 days-LD13 | 0.0 |
| [0 day (LD13) + 1 day (LD13 + 4B)] × 60 cycles | 60 days-(LD13 + 4B) | 100.0 |
| [1 day (LD13) + 1 day (LD13 + 4B)] × 30 cycles | 30 days-(LD13 + 4B) | 50.0 |
| [3 days (LD13) + 1 day (LD13 + 4B)] × 15 cycles | 15 days-(LD13 + 4B) | 25.0 |
| [5 days (LD13) + 1 day (LD13 + 4B)] × 10 cycles | 10 days-(LD13 + 4B) | 16.7 |
| [7 days (LD13) + 1 day (LD13 + 4B)] × 7 cycles + 4 days (LD13) | 7 days-(LD13 + 4B) | 11.7 |
| 1 day (SD10) × 60 cycles | 60 days-SD10 | 0.0 |
| Reference gene | Name | Accession number | Forward primer (5′ to 3′) | Reverse primer (5′ to 3′) |
| CmACTIN [64] | AB205087 | GATGACGCAGATCATGTTCG | AGCATGTGGAAGTGCATACC | |
| CmEF1α [65] | AB548817 | CTTGTTGCTTGATGACTGTGG | CTTGTTGCTTGATGACTGTGG | |
| Flowering-related gene | CmTFL1 [66] | AB839767 | CCATCATCAAGGCACAATTTCA | TTTCCCTTTGGCAGTTGAAGAA |
| CDM111 [67] | AY173054 | GGTCTCAAGAATATTCGCAC | TCATTAGTCATCCCATCAGC | |
| CmAFL1 [68] | AB451218 | CAAGCTCAACCATCAATAGTC | TGCAGCACATGAACGAGTAG | |
| CmFL [68] | AB451217 | CATTGATGCCATATTTAACTC | ACACGGATCATTCATTGTATA | |
| PCR conditions | PCR was performed with an initial denaturing step at 95 °C for 5 min, followed by 40 cycles at 95 °C for 5 s, 60 °C for 20 s, 72 °C for 30 s, and 72 °C for 10 min to final extension. The fluorescence was quantified after the incubation at 72 °C and repeated three times. | |||
| Antioxidant synthesis-related gene | CmPAL [69] | JF806362 | TACATTTTGGACGGGAGTGA | AGCGTATCGGTCTTGTTTGG |
| CmDFR [70] | GU324979 | GACATTATGGAAGGCGGATT | GTGGCAACATGAAACACTC | |
| PCR conditions | 30-s pre-denaturation at 95 °C followed by 5 s at 95 °C, 30 s at 60 °C, 40 cycles; the melting curves were 15 s at 95 °C, 60 s at 60 °C, and 15 s at 95 °C. The fluorescence signal was collected once for every 0.3 °C increase and repeated three times. | |||
| Treatment | Shoot | |||
| Shoot length (cm) | No. of branches/Plant | Dry weight (g) | ||
| 60 days-LD13 | 14.8 ± 0.53 ab 1 | 16.3 ± 1.03 a | 3.72 ± 0.05 bc | |
| 60 days-(LD13 + 4B) | 15.8 ± 0.41 a | 11.4 ± 0.97 b | 6.50 ± 0.10 a | |
| 30 days-(LD13 + 4B) | 15.7 ± 0.47 a | 11.3 ± 1.01 b | 5.81 ± 0.09 ab | |
| 15 days-(LD13 + 4B) | 15.5 ± 0.50 a | 14.4 ± 1.12 ab | 4.63 ± 0.07 b | |
| 10 days-(LD13 + 4B) | 13.5 ± 0.45 b | 14.8 ± 1.07 ab | 4.24 ± 0.06 b | |
| 7 days-(LD13 + 4B) | 13.2 ± 0.43 b | 16.2 ± 1.00 a | 3.92 ± 0.05 bc | |
| 60 days-SD10 | 9.3 ± 0.38 c | 7.7 ± 0.96 c | 1.80 ± 0.05 c | |
| Treatment | Leaf | Flower | ||
| No. of leaves/Plant | Leaf area 2 (cm2) | DVB 3 (day) | No. of flowers/Plant | |
| 60 days-LD13 | 337.2 ± 10.98 a | 3.8 ± 0.09 bc | - | 0.0 ± 0.00 e |
| 60 days-(LD13 + 4B) | 225.4 ± 6.97 c | 6.1 ± 0.11 a | 29.7 ± 1.74 c | 43.9 ± 5.76 a |
| 30 days-(LD13 + 4B) | 230.0 ± 6.24 c | 5.2 ± 0.08 ab | 34.2 ± 2.11 b | 25.7 ± 3.24 b |
| 15 days-(LD13 + 4B) | 276.7 ± 9.36 b | 4.5 ± 0.13 b | 48.6 ± 2.96 ab | 8.4 ± 1.06 d |
| 10 days-(LD13 + 4B) | 280.4 ± 7.01 b | 3.8 ± 0.17 bc | 51.9 ± 2.03 a | 5.6 ± 1.12 d |
| 7 days-(LD13 + 4B) | 301.6 ± 7.64 ab | 3.8 ± 0.20 bc | - | 0.0 ± 0.00 e |
| 60 days-SD10 | 181.2 ± 8.52 d | 2.9 ± 0.10 c | 26.2 ± 1.57 cd | 17.8 ± 3.73 c |
| Treatment | Pn 1 (μmol CO2 m−2·s−1) | Tr 2 (mmol H2O m−2·s−1) | Gs 3 (mol H2O m−2·s−1) | Ci 4 (μmol CO2 mol−1) | Fv/Fm 5 | Fv′/Fm′ 6 | qP7 |
| 60 days-LD13 | 12.02 ± 0.06 bc 8 | 1.52 ± 0.01 c | 0.50 ± 0.007 c | 384.6 ± 2.24 c | 0.81 ± 0.004 ab | 0.43 ± 0.001 bc | 0.45 ± 0.002 bc |
| 60 days-(LD13 + 4B) | 18.03 ± 0.07 a | 2.12 ± 0.02 a | 0.83 ± 0.009 a | 462.7 ± 3.97 a | 0.86 ± 0.003 a | 0.68 ± 0.004 a | 0.67 ± 0.004 a |
| 30 days-(LD13 + 4B) | 16.31 ± 0.07 ab | 1.91 ± 0.04 ab | 0.74 ± 0.006 ab | 443.1 ± 3.09 ab | 0.85 ± 0.006 a | 0.62 ± 0.006 ab | 0.60 ± 0.003 ab |
| 15 days-(LD13 + 4B) | 14.11 ± 0.04 b | 1.80 ± 0.02 b | 0.65 ± 0.004 b | 422.7 ± 4.01 b | 0.82 ± 0.007 ab | 0.51 ± 0.002 b | 0.53 ± 0.007 b |
| 10 days-(LD13 + 4B) | 14.08 ± 0.06 b | 1.65 ± 0.03 bc | 0.58 ± 0.009 bc | 407.1 ± 2.36 bc | 0.81 ± 0.005 ab | 0.51 ± 0.003 b | 0.51 ± 0.008 b |
| 7 days-(LD13 + 4B) | 12.07 ± 0.08 bc | 1.63 ± 0.04 bc | 0.57 ± 0.005 bc | 404.9 ± 1.28 bc | 0.80 ± 0.004 ab | 0.46 ± 0.005 bc | 0.47 ± 0.002 bc |
| 60 days-SD10 | 10.52 ± 0.06 c | 1.49 ± 0.05 c | 0.49 ± 0.007 c | 378.8 ± 3.79 c | 0.80 ± 0.007 ab | 0.37 ± 0.003 c | 0.40 ± 0.001 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Song, J.; Jeong, B.R. Blue Light Supplemented at Intervals in Long-Day Conditions Intervenes in Photoperiodic Flowering, Photosynthesis, and Antioxidant Properties in Chrysanthemums. Antioxidants 2022, 11, 2310. https://doi.org/10.3390/antiox11122310
Yang J, Song J, Jeong BR. Blue Light Supplemented at Intervals in Long-Day Conditions Intervenes in Photoperiodic Flowering, Photosynthesis, and Antioxidant Properties in Chrysanthemums. Antioxidants. 2022; 11(12):2310. https://doi.org/10.3390/antiox11122310
Chicago/Turabian StyleYang, Jingli, Jinnan Song, and Byoung Ryong Jeong. 2022. "Blue Light Supplemented at Intervals in Long-Day Conditions Intervenes in Photoperiodic Flowering, Photosynthesis, and Antioxidant Properties in Chrysanthemums" Antioxidants 11, no. 12: 2310. https://doi.org/10.3390/antiox11122310