
Antioxidant Baccharis trimera Leaf Extract Suppresses Lipid Accumulation in C. elegans Dependent on Transcription Factor NHR-49
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Acquisition of B. trimera Leaf Extract (BT)
2.3. Physicochemical Characterization of BT
2.4. Assessment of BT Cytotoxicity
2.5. In Vitro Antioxidant Assays
2.6. Assays with C. elegans
2.6.1. Conditions for the Maintenance and Treatment of C. elegans
2.6.2. Bacterial Growth Assay
2.6.3. Evaluation of BT Toxicity in C. elegans
2.6.4. Intracellular Accumulation of ROS in C. elegans
2.6.5. Oxidative Stress Survival Assay
2.6.6. Oil Red O Staining
2.6.7. Triglyceride Quantification Assay
2.7. Statistical Analyzes
3. Results
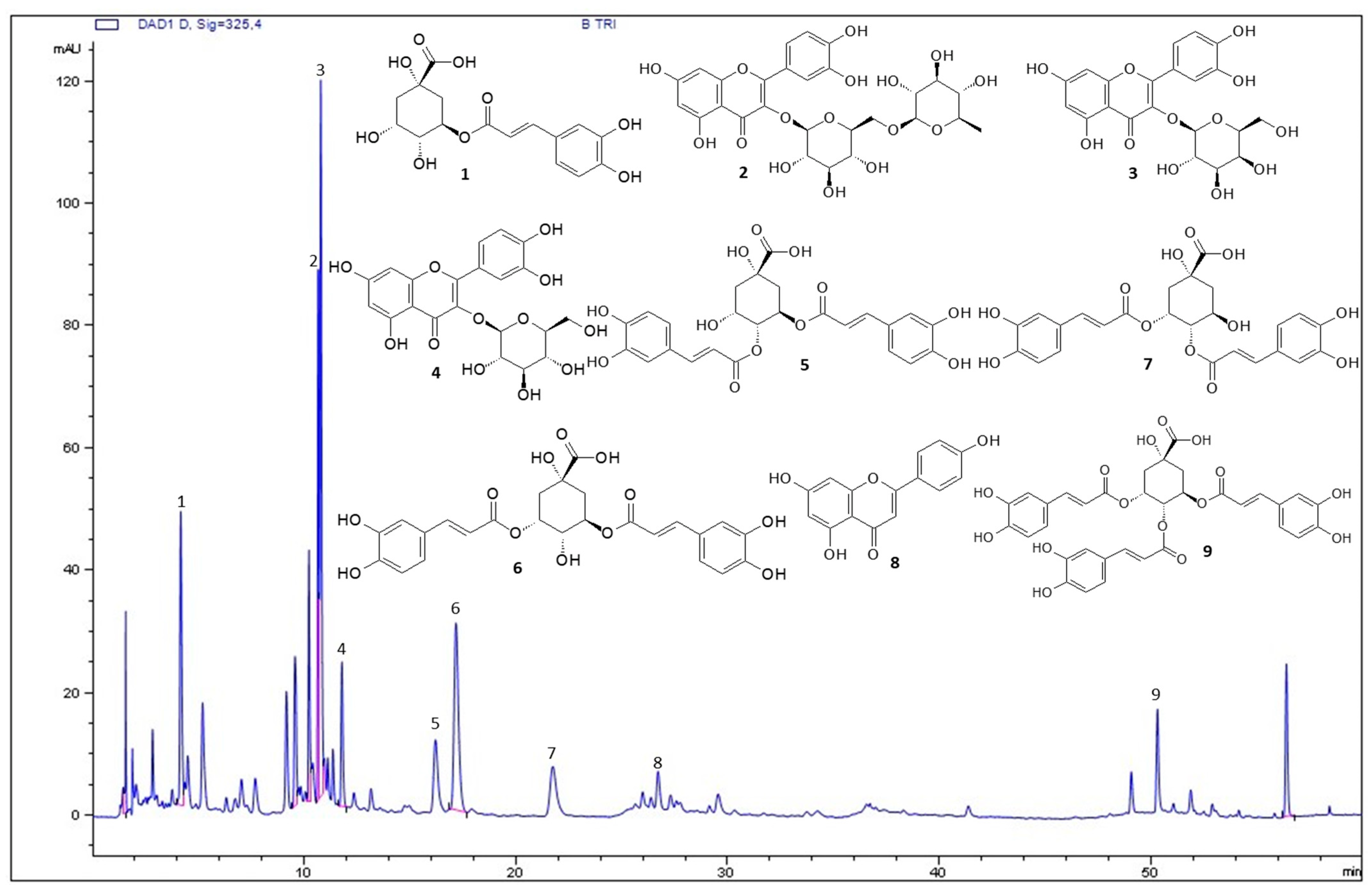
3.1. Characterization of BT
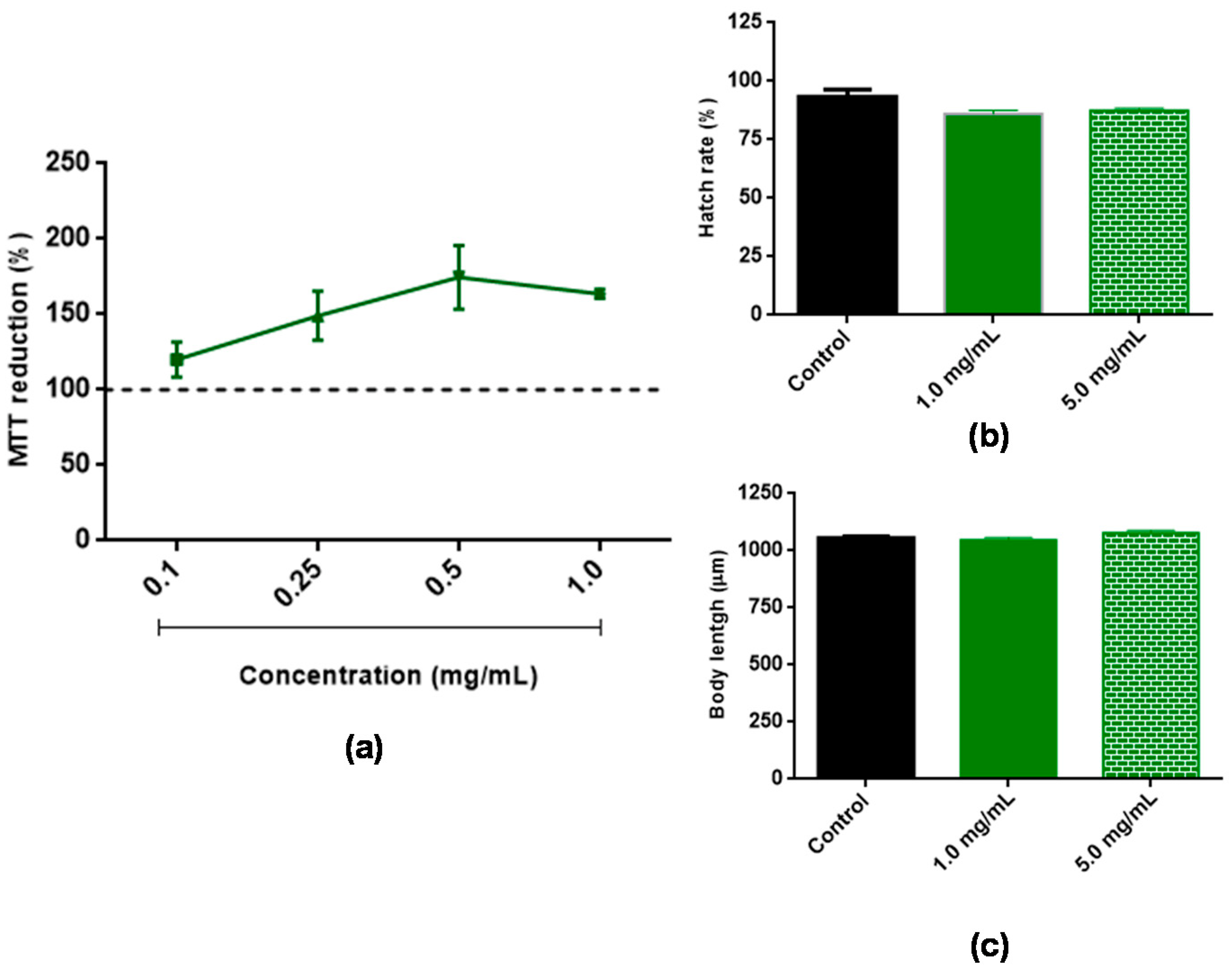
3.2. Assessment of BT Toxicity
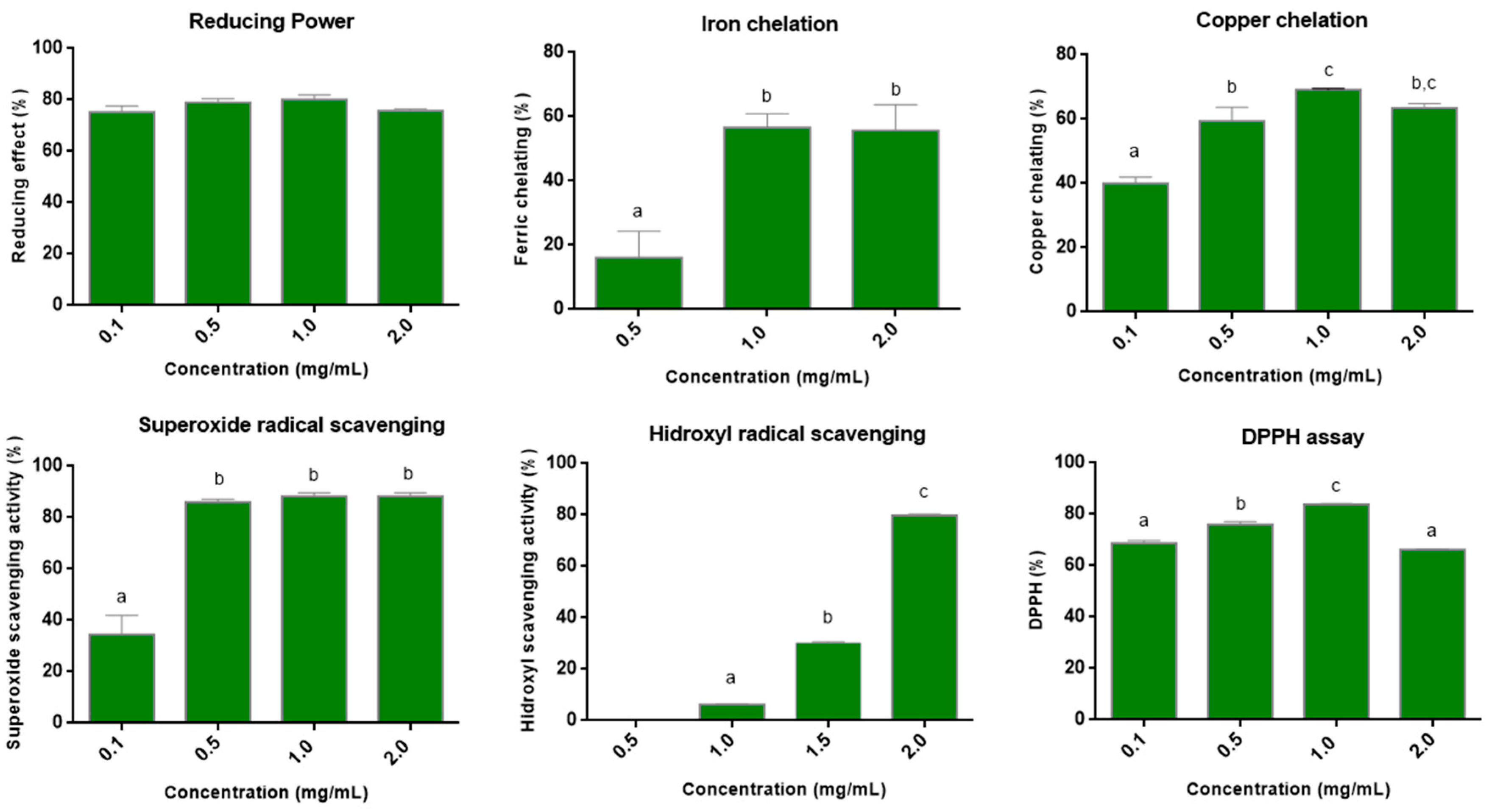
3.3. Antioxidant Assays In Vitro
3.4. Bacterial Growth Assay
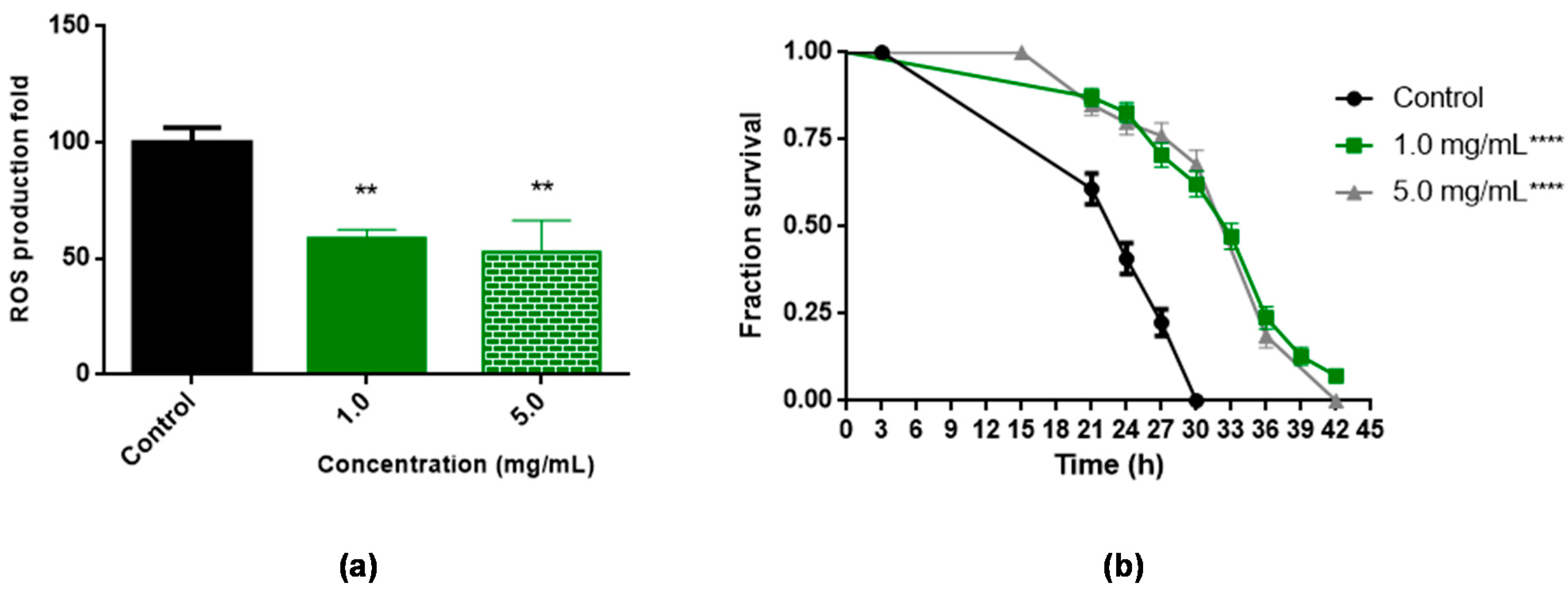
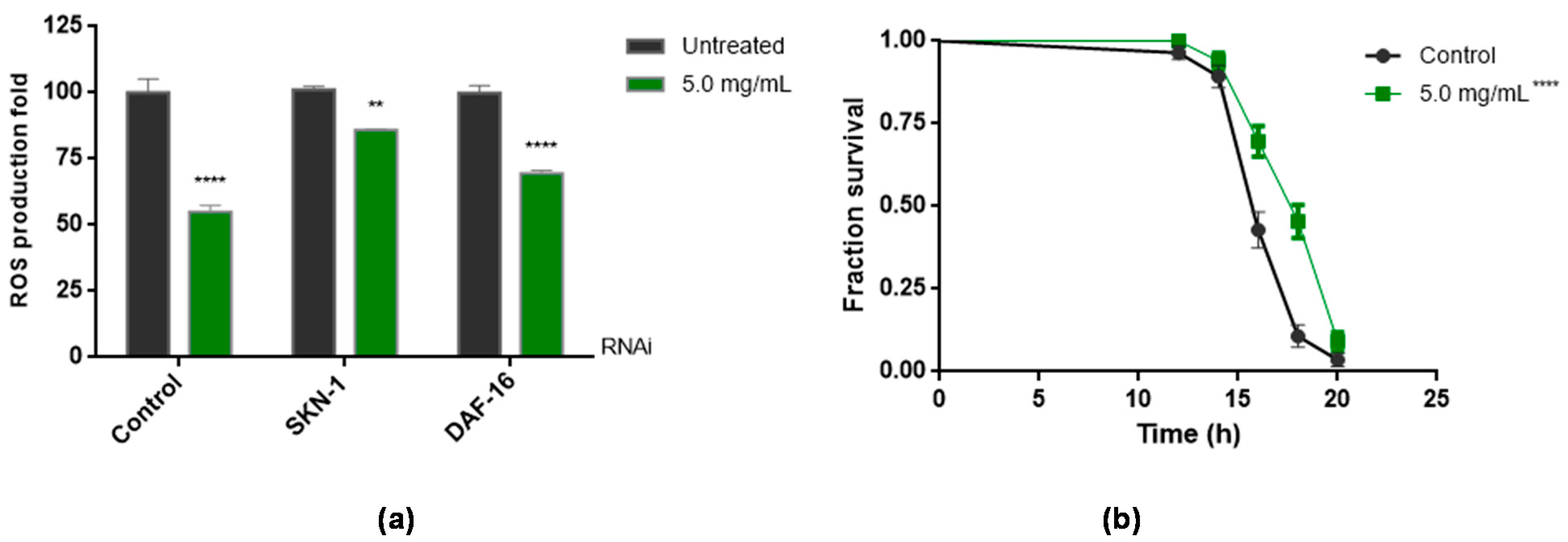
3.5. Antioxidant Assays In Vivo Using C. elegans
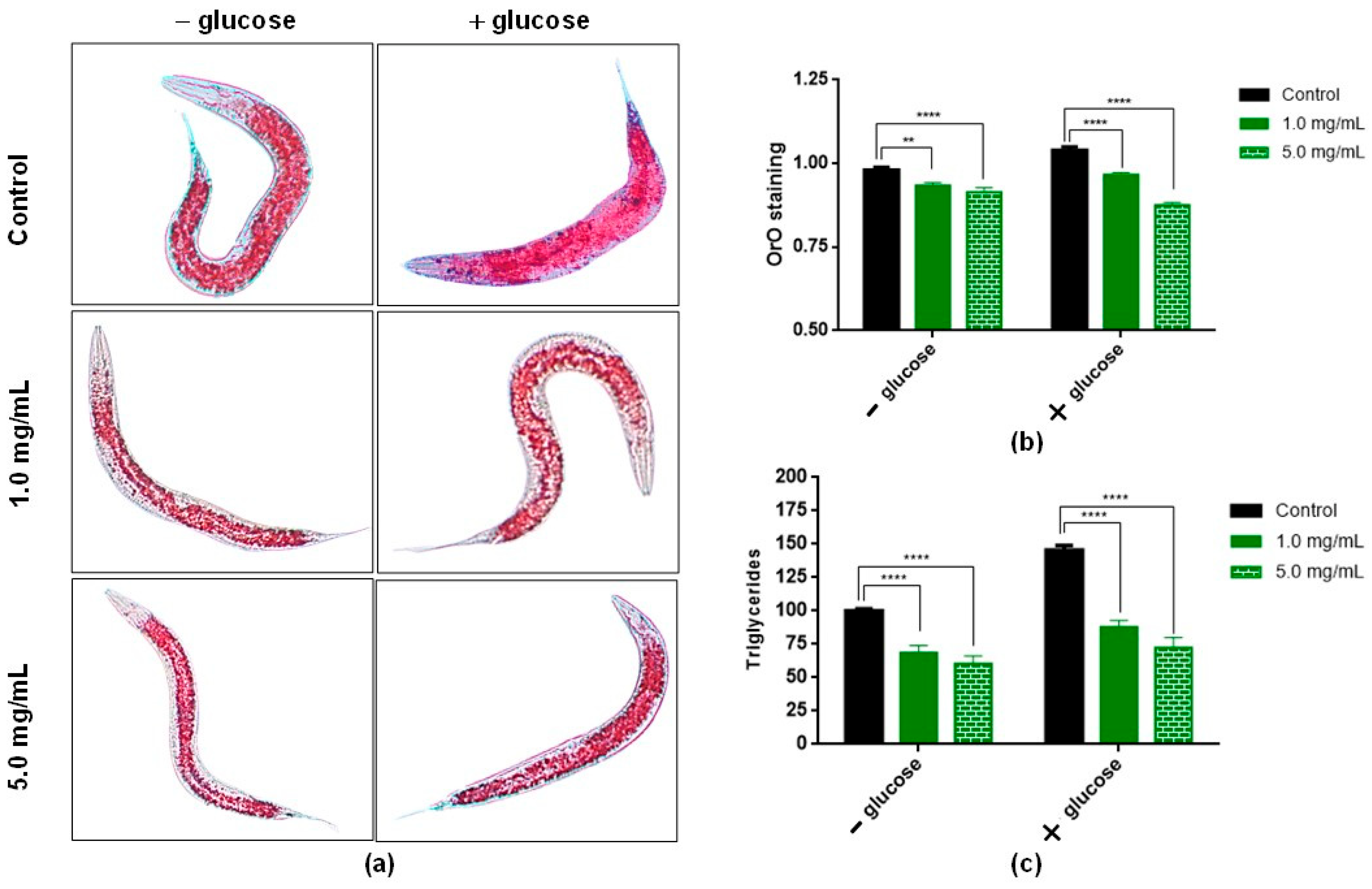
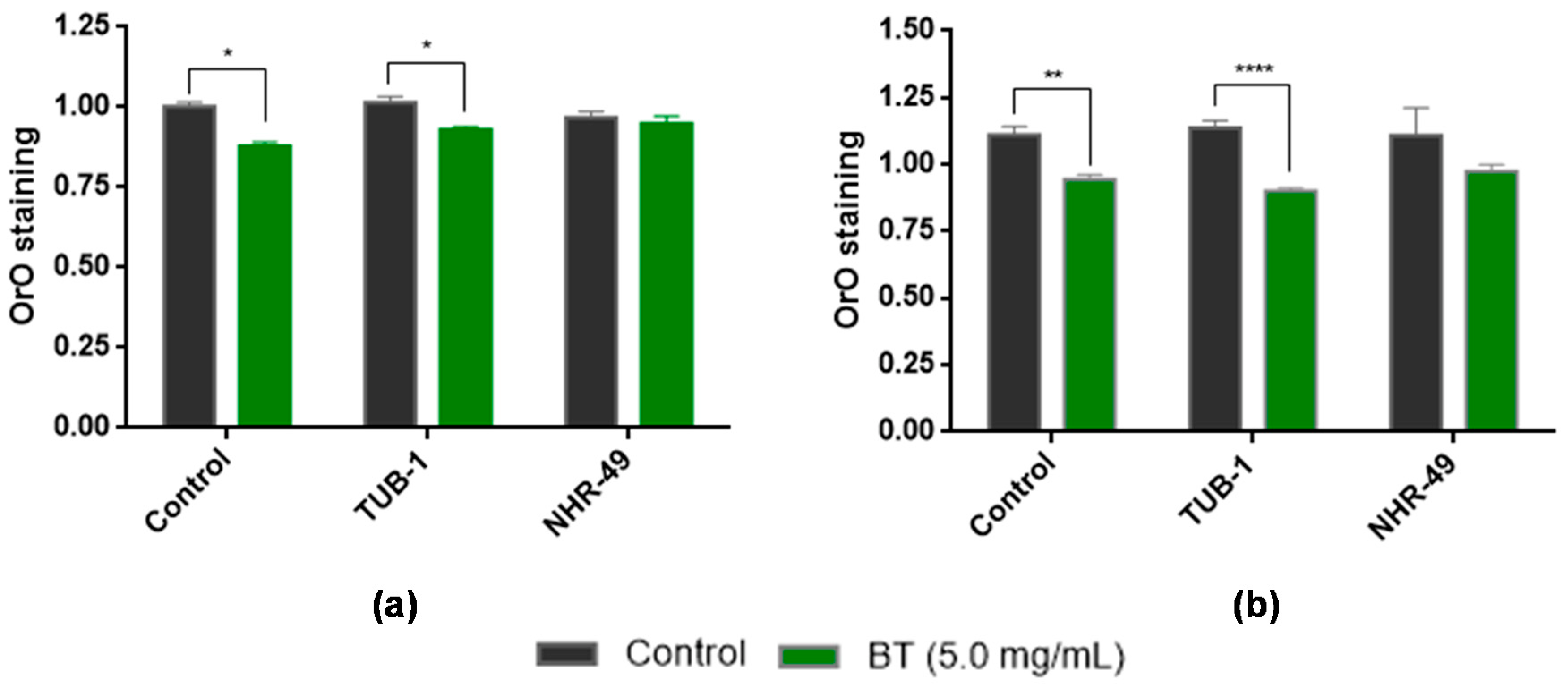
3.6. In Vivo Assays to Assess Lipid Accumulation Using C. elegans
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Obesity and Overweight. 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 18 July 2022).
- World Obesity Atlas 2022. Available online: https://www.worldobesity.org/resources/resource-library/world-obesity-atlas-2022 (accessed on 18 July 2022).
- Kotsis, V.; Stabouli, S.; Bouldin, M.; Low, A.; Toumanidis, S.; Zakopoulos, N. Impact of obesity on 24-h ambulatory blood pressure and hypertension. Hypertension 2005, 45, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Stabouli, S.; Kotsis, V.; Papamichael, C.; Constantopoulos, A.; Zakopoulos, N. Adolescent obesity is associated with high ambulatory blood pressure and increased carotid intimal medial thickness. J. Pediatr. 2005, 147, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Renehan, A.G.; Tyson, M.; Egger, M.; Heller, R.F.; Zwahlen, M. Body-mass index and incidence of cancer: A systematic review and meta-analysis of prospective observational studies. Lancet 2008, 371, 569–578. [Google Scholar] [CrossRef]
- Zalensin, K.C.; Franklin, B.A.; Miller, W.M.; Peterson, E.D.; McCullough, P.A. The impact of obesity on cardiovascular disease. Med. Clin. N. 2011, 95, 919–937. [Google Scholar] [CrossRef]
- Marbniang, S.P.; Lhungdim, H.; Yadav, B.; Yajurvedi, V.K. Overweight/obesity risks and prevalence of diabetes and hypertension in Northeastern India: An analysis using seemingly unrelated probit model. Clin. Epidemiol. Glob. Health Clin. Epidemiol. 2021, 11, 100764. [Google Scholar] [CrossRef]
- Kitabchi, A.E.; McDaniel, K.A.; Wan, J.Y.; Tylavsky, F.A.; Jacovino, C.A.; Sands, C.W.; Nyenwe, E.A.; Stentz, F.B. Effects of high-protein versus high-carbohydrate diets on markers of beta-cell function, oxidative stress, lipid peroxidation, proinflammatory cytokines, and adipokines in obese, premenopausal women without diabetes: A randomized controlled trial. Diabetes Care 2013, 36, 1919–1925. [Google Scholar] [CrossRef]
- Savini, I.; Catani, M.V.; Evangelista, D.; Gasperi, V.; Avigliano, L. Obesity-associated oxidative stress: Strategies finalized to improve redox state. Int. J. Mol. Sci. 2013, 14, 10497–10538. [Google Scholar] [CrossRef]
- Lee, H.; Lee, Y.J.; Choi, H.; Ko, E.H.; Kim, J.-W. Reactive oxygen species facilitate adipocyte differentiation by accelerating mitotic clonal expansion. J. Biol. Chem. 2009, 17, 284. [Google Scholar] [CrossRef]
- De Ferranti, S.; Mozaffarian, D. The Perfect Storm: Obesity, Adipocyte Dysfunction, and Metabolic Consequences. Clin. Chem. 2008, 54, 945–955. [Google Scholar] [CrossRef]
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, Á.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, oxidative stress, an obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef] [Green Version]
- Zielinska-Blizniewska, H.; Sitarek, P.; Merecz-Sadowska, A.; Malinowska, K.; Zajdel, K.; Jablonska, M.; Sliwinski, T.; Zajdel, R. Plant Extracts and Reactive Oxygen Species as Two Counteracting Agents with Anti- and Pro-Obesity Properties. Int. J. Mol. Sci. 2019, 20, 4556. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Jiang, Y.; Guo, J.; Su, Z. Natural Products with anti-obesity effects and different mechanisms of action. J. Food Sci. 2016, 64, 9571–9585. [Google Scholar] [CrossRef] [PubMed]
- Lavie, C.J.; Laddu, D.; Arena, R.; Ortega, F.B.; Alpert, A.A.; Kushner, R.F. Healthy Weight and Obesity Prevention: JACC Health Promotion Series. JACC J. 2018, 72, 1506–1531. [Google Scholar] [CrossRef] [PubMed]
- Saunders, K.H.; Umashanker, D.; Igel, L.I.; Kumar, R.B.; Aronne, L.J. Obesity Pharmacotherapy. Med. Clin. N. 2018, 102, 135–148. [Google Scholar] [CrossRef]
- Zhang, W.L.; Zhu, L.; Jiang, J.G. Active ingredients from natural botanicals in the treatment of obesity. Obes. Rev. 2014, 15, 957–967. [Google Scholar] [CrossRef]
- Mozaffarian, D. Dietary and Policy Priorities for Cardiovascular Disease, Diabetes, and Obesity: A Comprehensive Review. Circulation 2016, 133, 187–225. [Google Scholar] [CrossRef]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- Hassan, W.; Noreen, H.; Rehman, S.; Gul, S.; Amjad Kamal, M.; Paul Kamdem, J.; Zaman, B.; da Rocha, J.B.T. Oxidative Stress and Antioxidant Potential of One Hundred Medicinal Plants. Curr. Top Med. Chem. 2017, 17, 1336–1370. [Google Scholar] [CrossRef]
- Limanaqi, F.; Biagioni, F.; Mastroiacovo, F.; Polzella, M.; Lazzeri, G.; Fornai, F. Merging the Multi-Target Effects of Phytochemicals in Neurodegeneration: From Oxidative Stress to Protein Aggregation and Inflammation. Antioxidants 2020, 20, 1022. [Google Scholar] [CrossRef]
- Cercato, L.M.; White, P.A.S.; Nampo, F.K.; Santos, M.R.V.; Camargo, E.A. A systematic review of medicinal plants used for weight loss in Brazil: Is there potential for obesity treatment? J. Ethnopharmacol. 2015, 24, 286–296. [Google Scholar] [CrossRef]
- Rabelo, A.C.S.; Costa, D.C. A review of biological and pharmacological activities of Baccharis trimera. Chem. Biol. Interact. 2018, 296, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.J.A.; Bessa, A.L.; Benito, P.B. Biologically active substances from the genus Baccharis L. (Compositae). Stud. Nat. Prod. Chem. 2005, 30, 703–759. [Google Scholar] [CrossRef]
- Barbosa, R.J.; Silva, G.R.; Cola, I.M.; Kuchler, J.C.; Coelho, N.; Barboza, L.N.; Menetrier, J.V.; De Souza, R.; Zonta, F.N.; Froehlich, D.L.; et al. Promising therapeutic use of Baccharis trimera (less.) DC. as a natural hepatoprotective agent against hepatic lesions that are caused by multiple risk factors. J. Ethnopharmacol. 2020, 254, 112729. [Google Scholar] [CrossRef] [PubMed]
- Lívero, F.A.R.; Da Silva, L.M.; Ferreira, D.M.; Galuppo, L.F.; Borato, D.G.; Prando, T.B.; Lourenço, E.L.; Strapasson, R.L.; Stefanello, M.É.; Werner, M.F.; et al. Hydroethanolic extract of Baccharis trimera promotes gastroprotection and healing of acute and chronic gastric ulcers induced by ethanol and acetic acid. Naunyn-Schmiedeb. Arch. Pharmacol. 2016, 389, 985–998. [Google Scholar] [CrossRef] [PubMed]
- Kaut, N.N.N.; Rabelo, A.C.S.; Araujo, G.R.; Taylor, J.G.; Silva, M.E.; Pedrosa, M.L.; Chaves, M.M.; Junior, J.V.R.; Costa, D.C. Baccharis trimera (Carqueja) Improves Metabolic and Redox Status in an Experimental Model of Type 1 Diabetes. Evid. Based Complement. Altern. Med. 2018, 2018, 6532637. [Google Scholar] [CrossRef]
- Souza, S.P.; Pereira, L.L.S.; Souza, A.A.; Souza, R.V.; Dos Santos, C.D. Estudo da atividade antiobesidade do extrato metanólico de Baccharis trimera (Less.) DC. Braz. J. Pharm. Sci. 2012, 93, 27–32. [Google Scholar]
- Figueiredo, A.P.; Pereira, R.S. Estudo dos efeitos de cápsulas de carqueja (Baccharis trimera (Less) D.C.), sobre o metabolismo lipídico de pacientes em processo de emagrecimento. Conex. Ciência 2009, 4, 15. [Google Scholar] [CrossRef]
- Souza, S.P.; Pereira, L.L.S.; Souza, A.A.; Santos, C.D. Inhibition of pancreatic lipase by extracts of Baccharis trimera: Evaluation of antinutrients and effect on glycosidases. Braz. J. Pharmacog 2011, 21, 450–455. [Google Scholar] [CrossRef]
- Nascimento, D.S.M.; Oliveira, R.M.; Camara, R.B.G.; Gomes, D.L.; Monte, J.F.S.; Costa, M.S.S.P.; Fernandes, J.M.; Langassner, S.M.Z.; Rocha, H.A.O. Baccharis trimera (Less.) DC exhibits an anti-adipogenic effect by inhibiting the expression of proteins involved in adipocyte differentiation. Molecular 2017, 22, 972. [Google Scholar] [CrossRef]
- Fabri, R.L.; Nogueira, M.S.; Dutra, L.B.; Bouzada, M.L.M.; Scio, E. Antioxidant and antimicrobial potential of Asteraceae species. Braz. J. Med. Biol. 2011, 13, 183–189. [Google Scholar] [CrossRef]
- Paiva, F.A.; Bonomo, L.F.; Boasquivis, P.F.; Paula, I.T.B.R.; Guerra, J.F.C.; Leal, W.M.; Silva, M.E.; Pedrosa, M.L.; Oliveira, R.P. Carqueja (Baccharis trimera) Protects against Oxidative Stress and 𝛽-Amyloid-Induced Toxicity in Caenorhabditis elegans. Oxidative Med. Cell Longev. 2015, 2015, 740162. [Google Scholar] [CrossRef] [Green Version]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxidative Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef]
- Gubert, P.; Puntel, B.; Lehmen, T.; Bornhorst, J.; Avila, D.S.; Aschner, M.; Soares, F.A.A. Reversible reprotoxic effects of manganese through DAF-16 transcription fator activation and vitellogenin downregulation in Caenorhabditis elegans. Life Sci. 2016, 151, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zou, X.; Ding, Y.; Wang, H.; Wu, X.; Liang, B. Comparative genomics and functional study of lipid metabolic genes in Caenorhabditis elegans. BMC Genom. 2013, 14, 164. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhou, B.; Oshiro-Rapley, N.; Li, M.; Paulo, J.A.; Webster, C.M.; Mou, F.; Kacergis, M.C.; Talkowski, M.E.; Carr, C.E.; et al. An ancient, unified mechanism for metformin growth inhibition in C. elegans and cancer. Cell 2016, 167, 1705–1718. [Google Scholar] [CrossRef] [PubMed]
- Walts, C.T.; Hanson, E.D.; Delmonico, M.J.; Yao, L.; Wang, M.Q.; Hurley, B.F. Do sex or race differences influence strength training effects on muscle or fat? Med. Sci. Sports Exerc. 2008, 40, 669–676. [Google Scholar] [CrossRef]
- Caland, R.B.O.; Cadavid, C.O.M.; Carmona, L.; Peña, L.; Oliveira, R.P. Pasteurized Orange Juice Rich in Carotenoids Protects Caenorhabditis elegans against Oxidative Stress and Amyloid Toxicity through Direct and Indirect Mechanisms. Oxidative Med. Cell Longev. 2019, 2019, 5046280. [Google Scholar] [CrossRef]
- Barreto, S.M.A.G.; Cadavid, C.O.M.; Moura, R.A.O.; Silva, G.M.M.; Araújo, V.F.; Filho, J.A.A.S.; Rocha, H.A.O.; Oliveira, R.P.; Giordani, R.B.; Ferrari, M. In Vitro and In Vivo Antioxidant Activity of Agave sisalana Agro-Industrial Residue. Biomolecules 2020, 10, 1435. [Google Scholar] [CrossRef]
- Menzel, R.; Rodel, M.; Kulas, J.; Steinberg, C.E.W. CYP35: Xenobiotically induced gene expression in the nematode Caenorhabditis elegans. Arch. Biochem. Biophys. 2005, 438, 93–102. [Google Scholar] [CrossRef]
- Oliveira, R.P.; Abate, J.P.; Dilks, K.; Landis, J.; Ashraf, J.; Murphy, C.T.; Blackwell, T.K. Condition adapted stress and longevity gene regulation by Caenorhabditis elegans SKN1/Nrf. Aging Cell 2009, 8, 524–541. [Google Scholar] [CrossRef]
- Tullet, J.M.A.; Green, J.W.; Au, C.; Benedetto, A.; Thompson, M.A.; Clark, E.; Gilliat, A.F.; Young, A.; Schmeisser, K.; Gems, D. The SKN-1/Nrf2 transcription factor can protect against oxidative stress and increase lifespan in C. elegans by distinct mechanisms. Aging Cell 2017, 16, 1191–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Gilst, M.R.; Hadjivassiliou, H.; Jolly, A.; Yamamoto, K.R. Nuclear hormone receptor NHR-49 controls fat consumption and fatty acid composition in C. elegans. PLoS Biol. 2005, 3, e53. [Google Scholar] [CrossRef] [PubMed]
- Mukhopzdhyay, A.; Pan, X.; Lambright, G.; Tissenbaum, H. An endocytic pathway as a target of tubby for regulation of fat storage. EMBO Rep. 2007, 8, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D. The transcriptional basis of adipocyte development. Prostaglandins Leukot. Essent. Fatty Acids 2005, 73, 31–34. [Google Scholar] [CrossRef]
- Ashrafi, K.; Chang, F.Y.; Watts, J.L.; Fraser, A.G.; Kamath, R.S.; Ahringer, J.; Ruvkun, G. Genome wide RNAi analysis of Caenorhabditis elegans fat regulatory genes. Nature 2003, 421, 268–272. [Google Scholar] [CrossRef]
- Goh, G.Y.S.; Winter, J.J.; Bhanshali, F.; Doering, K.R.S.; Lai, R.; Lee, K.; Veal, E.A.; Taubert, S. NHR-49/HNF4 integrates regulation of fatty acid metabolism with a protective transcriptional response to oxidative stress and fasting. Aging Cell 2018, 17, e12743. [Google Scholar] [CrossRef]
- Pathare, P.P.; Lin, A.; Bornfeldt, K.E.; Tauberts, S.; Van Gilst, M.R. Coordinate Regulation of Lipid Metabolism by Novel Nuclear Receptor Partnerships. PLoS Genet. 2012, 8, e1002645. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 250–256. [Google Scholar] [CrossRef]
- Spector, J. Refinement of the coomassie blue method of protein quantification. A simple and linear spectrophotometric assay of 0.5 to 50 µg of protein. Anal. Chem. 1978, 86, 142–143. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 5563. [Google Scholar] [CrossRef]
- Paiva, W.S.; de Souza Neto, F.E.; Queiroz, M.F.; Batista, L.A.N.C.; Rocha, H.A.O.; de Lima Batista, A.C. Oligochitosan Synthesized by Cunninghamella elegans, a Fungus from Caatinga (The Brazilian Savanna) Is a Better Antioxidant than Animal Chitosan. Molecules 2021, 27, 171. [Google Scholar] [CrossRef] [PubMed]
- Shimada, K.; Fujikawa, K.; Yahara, K.; Nakamura, T. Antioxidative properties of xanthan on the autoxidation of soybean oil in cyclodextrin emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Escorcia, W.; Ruter, D.L.; Nhan, J.; Curran, S.P. Quantification of lipid abundance and evaluation of lipid distribution in Caenorhabditis elegans by nile red and oil red O staining. J. Vis. Exp. 2018, 133, e57352. [Google Scholar] [CrossRef]
- Sabir, S.M.; Athayde, M.L.; Boligon, A.A.; Rocha, J.B.T. Antioxidant activities and phenolic profile of Baccharis trimera, a commonly used medicinal plant from Brazil. S. Afr. J. Bot. 2017, 113, 318–323. [Google Scholar] [CrossRef]
- Gené, R.M.; Cartaña, C.; Adzet, T.; Marin, E.; Parelha, T.; Cañigueral, S. Anti-inflammatory and analgesic activity of Baccharis trimera: Identification of its active constituents. Planta Med. 1996, 62, 232–235. [Google Scholar] [CrossRef]
- Silva, A.R.H.; Reginato, F.Z.; Guex, C.G.; Figueredo, K.C.; Araldi, I.C.C.; Freitas, R.B.; Boligon, A.A.; Athayde, M.L.; Mazzanti, C.M.A.; Hubscher, G.H.; et al. Acute and sub-chronic (28 days) oral toxicity evaluation of tincture Baccharis trimera (Less) Backer in male and female rodent animals. Regulat. Toxicol. Pharmacol. 2016, 74, 170–177. [Google Scholar] [CrossRef]
- Soicke, H.; Leng-Peschlow, E. Characterization of Flavonoids from Baccharis trimera and their Antihepatotoxic Properties. Stud. Nat. Prod. Chem. 1987, 30, 37–39. [Google Scholar] [CrossRef]
- Rodriguez, M.V.; Sortino, M.A.; Ivancovich, J.J.; Pellegrino, J.M.; Favier, L.S.; Raimondi, M.P.; Gattuso, M.A.; Zacchino, S.A. Detection of synergistic combinations of Baccharis extracts with Terbinafine against Trichophyton rubrum with high throughput screening synergy assay (HTSS) followed by 3D graphs. Behavior of some of their components. Phytomedicine 2013, 20, 1230–1239. [Google Scholar] [CrossRef]
- Rabelo, A.C.S.; Lucio, K.P.; Araujo, C.M.; Araujo, G.R.; Miranda, P.H.A.; Carneiro, A.C.A.; Ribeiro, E.M.; Silva, B.M.; Lima, W.G.; Costa, D.C. Baccharis trimera protects against ethanol induced hepatotoxicity in vitro and in vivo. J. Ethnopharmacol. 2017, 215, 1–13. [Google Scholar] [CrossRef]
- Pádua, B.C.; Rossoni-Júnior, J.V.; Brito Magalhães, C.L.; Chaves, M.M.; Silva, M.E.; Pedrosa, M.L.; Bianco, G.H.; Brandão, G.C.; Rodrigues, I.V.; Lima, W.G.; et al. Protective Effect of Baccharis trimera Extract on Acute Hepatic Injury in a Model of Inflammation Induced by Acetaminophen. Mediat. Inflamm. 2014, 2014, 196598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Cruz Pádua, B.; Silva, L.D.; Rossoni Júnior, J.V.; Humberto, L.; Chaves, M.M.; Silva, M.E.; Pedrosa, M.L.; Costa, D.C. Antioxidant properties of Baccharis trimera in the neutrophils of Fisher rats. J. Ethnopharmacol. 2010, 129, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Da Cruz Pádua, B.; Victor Rossoni Junior, J.; Lopes de Brito Magalhaes, C.; Brandao Seiberf, J.; Morais Araujo, C.; Henrique Bianco de Souza, G.; Martins Chaves, M.; Eustaquio Silva, M.; Lucia Pedrosa, M.; Caldeira Costa, D. Baccharis trimera improves the antioxidant defense system and inhibits iNOS and NADPH oxidase expression in a rat model of inflammation. Curr. Pharm. Biotechnol. 2013, 14, 975–984. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Badana, A.K.; Mohan, M.; Shailender, G.; Malla, R.R. Reactive Oxygen Species: A Key Constituent in Cancer Survival. Biomark. Insights 2018, 13, 1177271918755391. [Google Scholar] [CrossRef]
- Sahreen, S.; Khan, M.R.; Khan, R.A. Evaluation of antioxidant activities of various solvent extracts of Carissa opaca fruits. Food Chem. 2010, 122, 1205–1211. [Google Scholar] [CrossRef]
- McMurray, F.; Patten, D.A.; Harper, M.E. Reactive Oxygen Species and Oxidative Stress in Obesity-Recent Findings and Empirical Approaches. Obesity 2016, 24, 2301–2310. [Google Scholar] [CrossRef]
- Yuan, Y.; Kang, N.; Li, Q.; Zhang, Y.; Liu, Y.; Tan, P. Study of the Effect of Neutral Polysaccharides from Rehmannia glutinosa on Lifespan of Caenorhabditis elegans. Molecules 2019, 24, 4592. [Google Scholar] [CrossRef]
- Ribeiro, A.R.C.; Cordeiro, M.L.S.; Silva, L.M.P.; Cadavid, C.O.M.; Caland, R.B.O.; Fernandes-Negreiros, M.M.; Queiroz, M.F.; Barbosa, J.S.; Aragão, C.F.S.; Zucolotto, S.M.; et al. Myrciaria tenella (DC.) O. Berg (Myrtaceae) Leaves as a Source of Antioxidant Compounds. Antioxidants 2019, 8, 310. [Google Scholar] [CrossRef]
- Moraes, D.P.; Lozano-Sanchez, J.; Machado, M.L.; Vizzotto, M.; Lazzaretti, M.; LeyvaJimenez, F.J.J.; da Silveira, T.L.; Ries, E.F.; Barcia, M.T. Characterization of a new blackberry cultivar BRS Xingu: Chemical composition, phenolic compounds, and antioxidant capacity in vitro and in vivo. Food Chem. 2020, 322, 126783. [Google Scholar] [CrossRef]
- Davila-Trujillo, R.; Fernandes, S.S.; Lima Dora, C.; Monserrat, J.M.; Prentice, C.; de las Mercedes Salas-Mellado, M. Physical, chemical and biological evaluation of nanoparticles containing phenolic compounds from wine production residues. J. Food Process. 2021, 45, e15629. [Google Scholar] [CrossRef]
- Rossi, L.; Mazzitelli, S.; Arciello, M.; Capo, C.R.; Rotilio, G. Benefits from dietary polyphenols for brain aging and Alzheimer’s disease. Neurochem. Res. 2008, 33, 2390–2400. [Google Scholar] [CrossRef] [PubMed]
- Koch, K.; Havermann, S.; Buchter, C.; Watjen, W. Caenorhabditis elegans as model system in pharmacology and toxicology: Effects of flavonoids on redox-sensitive signalling pathways and ageing. Sci. World J. 2014, 2014, 920398. [Google Scholar] [CrossRef] [PubMed]
- Tobore, T.O. Towards a comprehensive theory of obesity and a healthy diet: The causal role of oxidative stress in food addiction and obesity. Behav. Brain Res. 2020, 20, 112560. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 3, 423–444. [Google Scholar] [CrossRef] [PubMed]
- Aranaz, P.; Navarro-Herrera, D.; Zabala, M.; Romo-Hualde, A.; Lopez-Yoldi, M.; Vizmanos, J.L.; Milagro, F.I.; Gonzalez-Navarro, C.J. Phenolic compounds reduce the fat content in Caenorhabditis elegans by affecting lipogenesis, lipolysis, and different stress responses. Pharmaceuticals 2020, 13, 355. [Google Scholar] [CrossRef]
- Oliveira, A.C.P.; Endringer, D.C.; Amorim, L.A.S.; Brandão, M.G.L.; Coelho, M.M. Effect of the extracts and fractions of Baccharis trimera and Syzygium cumini on glycaemia of diabetic and non-diabetic mice. J. Ethnopharmacol. 2005, 102, 465–469. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.; Deplancke, B.; Walhout, A.J.; Tissenbaum, H.A. C. elegans tubby regulates life span and fat storage by two independent mechanisms. Cell Metab. 2005, 2, 35–42. [Google Scholar] [CrossRef]
- Mak, H.Y.; Nelson, L.S.; Basson, M.; Johnson, C.D.; Ruykun, G. Polygenic control of Caenorhabditis elegans fat storage. Nat Genetics 2006, 38, 363–368. [Google Scholar] [CrossRef]
- Botta, M.; Audano, M.; Sahebkar, A.; Sirtori, C.R.; Mitro, N.; Ruscica, M. PPAR Agonists and Metabolic Syndrome: An Established Role? Int. J. Mol. Sci. 2018, 19, 1197. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Survival (hours ± SEM) | p Value (Long Rank) Extract vs. Untreated | n | |
|---|---|---|---|
| Control | 24.73 ± 0.33 a | < 0.0001 | 121 (3) |
| 1.0 mg/mL | 32.60 ± 0.49 b | < 0.0001 | 127 (3) |
| 5.0 mg/mL | 32.70 ± 0.56 b | < 0.0001 | 136 (3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza, F.R.M.; Silva, G.M.M.; Cadavid, C.O.M.; Lisboa, L.d.S.; Silva, M.M.C.L.; Paiva, W.S.; Ferreira, M.J.P.; de Paula Oliveira, R.; Rocha, H.A.O. Antioxidant Baccharis trimera Leaf Extract Suppresses Lipid Accumulation in C. elegans Dependent on Transcription Factor NHR-49. Antioxidants 2022, 11, 1913. https://doi.org/10.3390/antiox11101913
Souza FRM, Silva GMM, Cadavid COM, Lisboa LdS, Silva MMCL, Paiva WS, Ferreira MJP, de Paula Oliveira R, Rocha HAO. Antioxidant Baccharis trimera Leaf Extract Suppresses Lipid Accumulation in C. elegans Dependent on Transcription Factor NHR-49. Antioxidants. 2022; 11(10):1913. https://doi.org/10.3390/antiox11101913
Chicago/Turabian StyleSouza, Flávia Roberta Monteiro, Giovanna Melo Martins Silva, Cesar Orlando Muñoz Cadavid, Lucas dos Santos Lisboa, Maylla Maria Correia Leite Silva, Weslley Souza Paiva, Marcelo José Pena Ferreira, Riva de Paula Oliveira, and Hugo Alexandre Oliveira Rocha. 2022. "Antioxidant Baccharis trimera Leaf Extract Suppresses Lipid Accumulation in C. elegans Dependent on Transcription Factor NHR-49" Antioxidants 11, no. 10: 1913. https://doi.org/10.3390/antiox11101913