

Novel Cinnamaldehyde Derivatives Inhibit Peripheral Nerve Degeneration by Targeting Schwann Cells
 , ,
, , 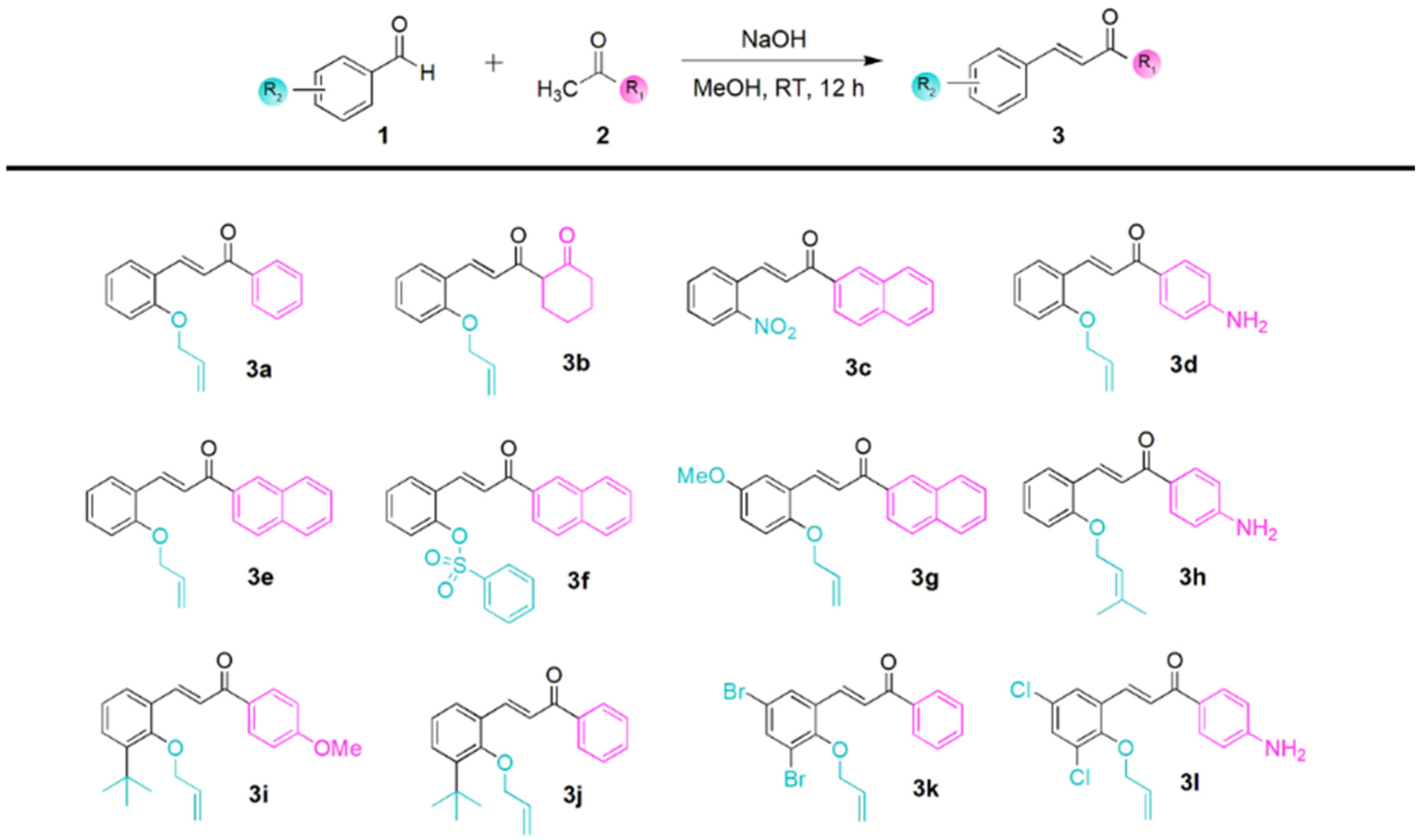{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Instruments
2.2. General Procedure
2.3. Cell Culture
2.4. Cell Viability Assay
2.5. Trypan Blue Exclusion Assay
2.6. Drug Efficacy Indices
2.7. Molecular Docking
2.8. Animals
2.9. Sciatic Nerve Explant Culture
2.10. Immunohistochemistry
2.11. Morphometric Indices
2.12. Bioinformatics and Validation
2.12.1. Functional Enrichment Analysis
2.12.2. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.13. Zebrafish Husbandry
2.14. Exposure of Zebrafish Embryos to 3f
2.15. Statistical Analysis
3. Results and Discussion
3.1. Synthesis of CAH Derivative 3
3.2. In Vitro Assay
3.3. Molecular Docking Studies
3.4. Ex Vivo and In Vivo Efficacy Assays
3.5. Inhibitory Phenotypes of PND Ex Vivo via 3f
3.6. Pharmacological Mechanism of 3f
3.7. In Vivo Embryo and Adult Zebrafish Toxicity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jessen, K.R.; Mirsky, R. The success and failure of the Schwann cell response to nerve injury. Front. Cell. Neurosci. 2019, 13, 33. [Google Scholar]
- Gandhi, M.; Fargo, E.; Prasad-Reddy, L.; Mahoney, K.M.; Isaacs, D. Diabetes: How to manage diabetic peripheral neuropathy. Drugs Context. 2022, 11, 2021. [Google Scholar]
- Muller, J.L.; Gutzeit, H.O. Plant Natural Products: Synthesis, Biological Functions and Practical Applications, 1st ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2014. [Google Scholar]
- Shreaz, S.; Wani, W.A.; Behbehani, J.M.; Raja, V.; Irshad, M.; Karched, M.; Ali, I.; Siddiqi, W.B.; Hun, L.T. Cinnamaldehyde and its derivatives, a novel class of antifungal agents. Fitoterapia 2016, 112, 116–131. [Google Scholar]
- Doyle, A.A.; Stephens, J.C. A review of Cinnamaldehyde and its derivatives as antibacterial agents. Fitoterapia 2019, 139, 104405. [Google Scholar]
- Gürer, B.; Kertmen, H.; Kuru Bektaşoğlu, P.; Öztürk, Ö.Ç.; Bozkurt, H.; Karakoç, A.; Arikok, A.T.; Celikoglu, E. The effects of Cinnamaldehyde on early brain injury and cerebral vasospasm following experimental subarachnoid hemorrhage in rabbits. Metab. Brain Dis. 2019, 34, 1737–1746. [Google Scholar]
- Hajinejad, M.; Ghaddaripouri, M.; Dabzadeh, M.; Forouzanfar, F.; Sahab-Negah, S. Natural Cinnamaldehyde and its derivatives ameliorate neuroinflammatory pathways in neurodegenerative diseases. Biomed. Res. Int. 2020, 2020, 1034325. [Google Scholar]
- Iranshahy, M.; Javadi, B.; Sahebkar, A. Protective effects of functional foods against Parkinson’s disease: A narrative review on pharmacology, phytochemistry, and molecular mechanisms. Phyther. Res. 2022, 36, 1952–1989. [Google Scholar]
- Rao, P.V.; Gan, S.H. Cinnamon: A multifaceted medicinal plant. Evid. Based Complement. Altern. Med. 2014, 2014, 642942. [Google Scholar]
- Chen, B.J.; Fu, C.S.; Li, G.H.; Wang, X.N.; Lou, H.X.; Ren, D.M.; Shen, T. Cinnamaldehyde analogues as potential therapeutic agents. Mini-Rev. Med. Chem. 2016, 17, 33–43. [Google Scholar]
- Manneck, D.; Manz, G.; Braun, H.; Rosendahl, J.; Stumpff, F. The TRPA1 agonist Cinnamaldehyde induces the secretion of HCO3− by the porcine colon. Int. J. Mol. Sci. 2021, 22, 5198. [Google Scholar]
- Legrand, C.; Merlini, J.M.; Senarclens-Bezencon, C.; Michlig, C. New natural agonists of the transient receptor potential Ankyrin1 (TRPA1) channel. Sci. Rep. 2020, 10, 11238. [Google Scholar]
- De Logu, F.; Nassini, R.; Materazzi, S.; Carvalho Gonçalves, M.; Nosi, D.; Rossi Degl’Innocenti, D.; Marone, I.M.; Ferreira, J.; Puma, S.L.; Benemei, S. Schwann cell TRPA1 mediates neuroinflammation that sustains macrophage-dependent neuropathic pain in mice. Nat. Commun. 2017, 8, 1887. [Google Scholar]
- De Logu, F.; De Pra, S.D.; Antoniazzi, C.T.D.; Kudsi, S.Q.; Ferro, P.R.; Landini, L.; Rigo, F.K.; Silveira, G.B.; Silveira, P.C.L.; Oliveira, S.M.; et al. Macrophages and Schwann cell TRPA1 mediate chronic allodynia in a mouse model of complex regional pain syndrome type I. Brain Behav. Immun. 2020, 88, 535–546. [Google Scholar]
- Carenini, S.; Maurer, M.; Werner, A.; Blazyca, H.; Toyka, K.V.; Schmid, C.D.; Raivich, G.; Martini, R. The role of macrophages in demyelinating peripheral nervous system of mice heterozygously deficient in p0. J. Cell Biol. 2001, 152, 301–308. [Google Scholar]
- Powers, D.G.; Casebier, D.S.; Fokas, D.; Ryan, W.J.; Troth, J.R.; Coffen, D.L. Automated paraller synthesis of chalcone-based screening libraries. Tetrahedron 1998, 54, 4085–4096. [Google Scholar]
- Lee, J.I. A review of the syntheses of flavanones, thioflavanones, andazaflavanones from 2′-substituted chalcones. Bull. Korean Chem. Soc. 2022, 43, 117–128. [Google Scholar]
- Brooks, E.A.; Galarza, S.; Gencoglu, M.F.; Cornelison, R.C.; Munson, J.M.; Peyton, S.R. Applicability of drug response metrics forcancer studies using biomaterials. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180226. [Google Scholar]
- Zhu, H.S.; Yang, X.L.; Wang, L.B.; Zhao, D.X.; Chen, L. Effects of extracts from sporoderm-broken spores of Ganoderma lucidum on HeLa cells. Cell Biol. Toxicol. 2000, 16, 201–206. [Google Scholar]
- Park, B.S.; Kim, H.W.; Rhyu, I.J.; Park, C.; Yeo, S.G.; Huh, Y.B.; Jeong, N.Y.; Jung, J.Y. Hydrogen sulfide is essential for Schwann cell responses to peripheral nerve injury. J. Neurochem. 2015, 132, 230–242. [Google Scholar]
- Chen, R.; Wu, J.; Lu, C.; Yan, T.; Qian, Y.; Shen, H.; Zhao, Y.; Wang, J.; Kong, P.; Zhang, X. Systematic transcriptome analysis reveals the inhibitory function of Cinnamaldehyde in non-small cell lung cancer. Front. Pharmacol. 2021, 11, 611060. [Google Scholar]
- Dennis, G., Jr.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for annotation, visualization, and integrated discovery. Genome Biol. 2003, 4, P3. [Google Scholar]
- von Hellfeld, R.; Brotzmann, K.; Baumann, L.; Strecker, R.; Braunbeck, T. Adverse effects in the fish embryo acute toxicity (FET) test: A catalogue of unspecific morphological changes versus more specific effects in zebrafish (Danio rerio) embryos. Environ. Sci. Eur. 2020, 32, 122. [Google Scholar]
- Talavera, K.; Startek, J.B.; Alvarez-Collazo, J.; Boonen, B.; Alpizar, Y.A.; Sanchez, A.; Naert, R.; Nilius, B. Mammalian transient receptor potential TRPA1 channels: From structure to disease. Physiol. Rev. 2020, 100, 725–803. [Google Scholar]
- Kim, Y.H.; Lee, S.M.; Yang, H.J.; Chun, Y.L.; Kim, D.K.; Yeo, S.G.; Park, C.; Jung, J.Y.; Huh, Y.B. Inhibition of transient receptor potential melastatin 7 (TRPM7) protects against Schwann cell trans-dedifferentiation and proliferation during Wallerian degeneration. Anim. Cells Syst. 2020, 24, 189–196. [Google Scholar]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron 2004, 41, 849–857. [Google Scholar]
- Lieder, B.; Hoi, J.; Burian, N.; Hans, J.; Holik, A.K.; Beltran Marquez, L.R.; Ley, J.P.; Hatt, H.; Somoza, V. Structure-dependent effects of Cinnamaldehyde derivatives on TRPA1-induced serotonin release in human intestinal cell models. J. Agric. Food Chem. 2020, 68, 3924–3932. [Google Scholar]
- Allouche, A. Gabedit—A graphical user interface for computational chemistry softwares. J. Comput. Chem. 2012, 32, 174–182. [Google Scholar]
- Zhuang, C.; Zhang, W.; Sheng, C.; Zhang, W.; Xing, C.; Miao, Z. Chalcone: A privileged structure in medicinal chemistry. Chem. Rev. 2017, 117, 7762–7810. [Google Scholar]
- Mahapatra, D.K.; Bharti, S.K.; Asati, V. Anti-cancer chalcones: Structural and molecular target perspectives. Eur. J. Med. Chem. 2015, 98, 69–114. [Google Scholar]
- Stettner, M.; Wolffram, K.; Mausberg, A.K.; Albrecht, P.; Derksen, A.; Methner, A.; Dehmel, T.; Hartung, H.P.; Dietrich, H.; Kieseier, B.C. Promoting myelination in an in vitro mouse model of the peripheral nervous system: The effect of wine ingredients. PLoS ONE 2013, 8, e66079. [Google Scholar]
- Chen, Y.; Wang, H.; Yoon, S.O.; Xu, X.; Hottiger, M.O.; Svaren, J.; Nave, K.A.; Kim, H.A.; Olson, E.N.; Lu, Q.R. HDAC-mediated deacetylation of NF-κB is critical for Schwann cell myelination. Nat. Neurosci. 2011, 14, 437–441. [Google Scholar]
- Yokoi, H.; Tsuruo, Y.; Ishimura, K. Steroid 5alpha-reductase type 1 immunolocalized in the rat peripheral nervous system and paraganglia. Histochem. J. 1998, 30, 731–739. [Google Scholar]
- Smit, J.J.; Baas, F.; Hoogendijk, J.E.; Jansen, G.H.; van der Valk, M.A.; Schinkel, A.H.; Berns, A.J.; Acton, D.; Nooter, K.; Burger, H.; et al. Peripheral neuropathy in mice transgenic for a human MDR3 P-glycoprotein mini-gene. J. Neurosci. 1996, 16, 6386–6393. [Google Scholar]
- Weerasuriya, A.; Mizisin, A.P. The blood-nerve barrier: Structure and functional significance. Methods Mol. Biol. 2011, 686, 149–173. [Google Scholar]
- Jung, S.H.; Kim, S.H.; Chung, A.Y.; Kim, H.T.; So, J.H.; Ryu, J.H.; Park, H.C.; Kim, C.H. Visualization of myelination in GFP-transgenic zebrafish. Dev. Dyn. 2010, 239, 592–597. [Google Scholar]
- Jessen, K.R.; Mirsky, R. The repair Schwann cell and its function in regenerating nerves. J. Physiol. 2016, 594, 3521–3531. [Google Scholar]
- Nocera, G.; Jacob, C. Mechanisms of Schwann cell plasticity involved in peripheral nerve repair after injury. Cell. Mol. Life Sci. 2020, 77, 3977–3989. [Google Scholar]
- Chun, Y.L.; Lee, S.; Park, K.H.; Park, C.; Huh, Y.; Jeong, N.Y.; Jung, J. Protective and therapeutic effect of (S)-ginsenoside F1 on peripheral nerve degeneration targeting Schwann cells: A pharmaco-neuroanatomical approach. Anat. Sci. Int. 2022, 97, 79–89. [Google Scholar]
- Jessen, K.R.; Arthur-Farraj, P. Repair Schwann cell update: Adaptive reprogramming, EMT, and stemness in regenerating nerves. Glia 2019, 67, 421–437. [Google Scholar]
- Vincent, A.M.; Kato, K.; McLean, L.L.; Soules, M.E.; Feldman, E.L. Sensory neurons and Schwann cells respond to oxidative stress by increasing antioxidant defense mechanisms. Antioxid. Redox Signal. 2009, 11, 425–438. [Google Scholar]
- Jeanette, H.; Marziali, L.N.; Bhatia, U.; Hellman, A.; Herron, J.; Kopec, A.M.; Feltri, M.L.; Poitelon, Y.; Belin, S. YAP and TAZ regulate Schwann cell proliferation and differentiation during peripheral nerve regeneration. Glia 2021, 69, 1061–1074. [Google Scholar]
- Li, J.Q.; Jiang, H.J.; Su, X.Y.; Feng, L.; Zhan, N.Z.; Li, S.S.; Chen, Z.J.; Chang, B.H.; Cheng, P.Z.; Yang, L. Schwann cells accelerate osteogenesis via the Mif/CD74/FOXO1 signaling pathway in vitro. Stem Cells Int. 2022, 2022, 4363632. [Google Scholar]
- Hackett, A.R.; Strickland, A.; Milbrandt, J. Disrupting insulin signaling in Schwann cells impairs myelination and induces a sensory neuropathy. Glia 2020, 68, 963–978. [Google Scholar]
- Newbern, J.; Birchmeier, C. Nrg1/ErbB signaling networks in Schwann cell development and myelination. Semin. Cell Dev. Biol. 2010, 21, 922–928. [Google Scholar]
- Ishii, A.; Furusho, M.; Bansal, R. Mek/ERK1/2-MAPK and PI3K/Akt/mTOR signaling plays both independent and cooperative roles in Schwann cell differentiation, myelination and dysmyelination. Glia 2021, 69, 2429–2446. [Google Scholar]
- Hardy, M.; Reddy, U.R.; Pleasure, D. Platelet-derived growth factor and regulation of Schwann cell proliferation in vivo. J. Neurosci. Res. 1992, 31, 254–262. [Google Scholar]
- Jessen, K.R.; Mirsky, R. The origin and development of glial cells in peripheral nerves. Nat. Rev. Neurosci. 2005, 6, 671–682. [Google Scholar]
- Mather, B.D.; Viswanathan, K.; Miller, K.M.; Long, T.E. Michael addition reactions in macromolecular design for emerging technologies. Prog. Polym. Sci. 2006, 31, 487–531. [Google Scholar]
- Babtie, A.C.; Lima, M.F.; Kirby, A.J.; Hollfelder, F. Kinetic and computational evidence for an intermediate in the hydrolysis of sulfonate esters. Org. Biomol. Chem. 2012, 10, 8095–8101. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chun, Y.L.; Park, K.-H.; Pallavi, B.; Eom, W.-J.; Park, C.; Huh, Y.; Lee, Y.; Lee, J.; Kim, S.H.; Yeo, S.G.; et al. Novel Cinnamaldehyde Derivatives Inhibit Peripheral Nerve Degeneration by Targeting Schwann Cells. Antioxidants 2022, 11, 1846. https://doi.org/10.3390/antiox11101846
Chun YL, Park K-H, Pallavi B, Eom W-J, Park C, Huh Y, Lee Y, Lee J, Kim SH, Yeo SG, et al. Novel Cinnamaldehyde Derivatives Inhibit Peripheral Nerve Degeneration by Targeting Schwann Cells. Antioxidants. 2022; 11(10):1846. https://doi.org/10.3390/antiox11101846
Chicago/Turabian StyleChun, Yoo Lim, Ki-Hoon Park, Badvel Pallavi, Won-Joon Eom, Chan Park, Youngbuhm Huh, Yeonjoo Lee, Jimin Lee, Sang Hoon Kim, Seung Geun Yeo, and et al. 2022. "Novel Cinnamaldehyde Derivatives Inhibit Peripheral Nerve Degeneration by Targeting Schwann Cells" Antioxidants 11, no. 10: 1846. https://doi.org/10.3390/antiox11101846