
Human Milk Antioxidative Modifications in Mastitis: Further Beneficial Effects of Cranberry Supplementation

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Collection of Samples
2.3. Methods
2.4. TAC Determination
2.5. Assay of GPx Activity
2.6. Assay of GSH Concentration
2.7. TPP Determination
2.8. Assay of MDA Concentration
2.9. Quantification of CGC
2.10. Protein Concentration Measurement
2.11. Statistical Analysis
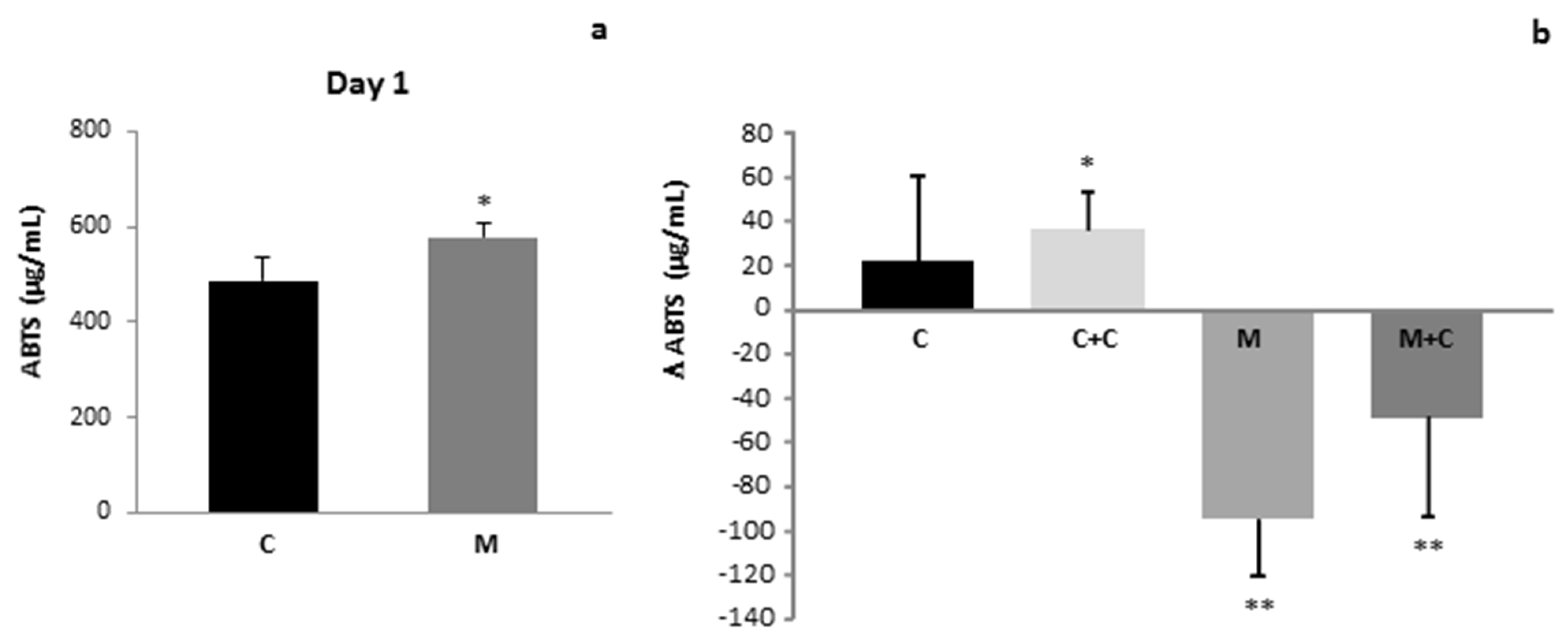
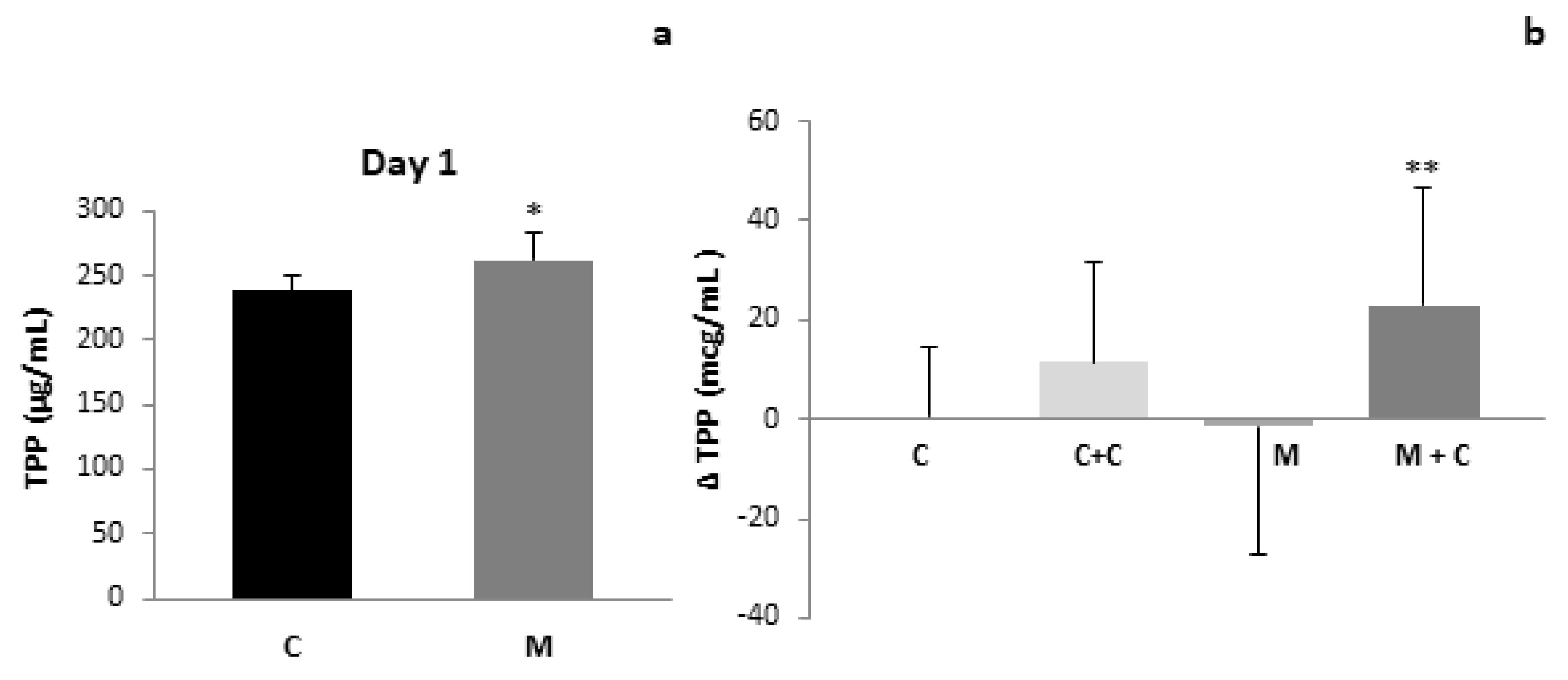
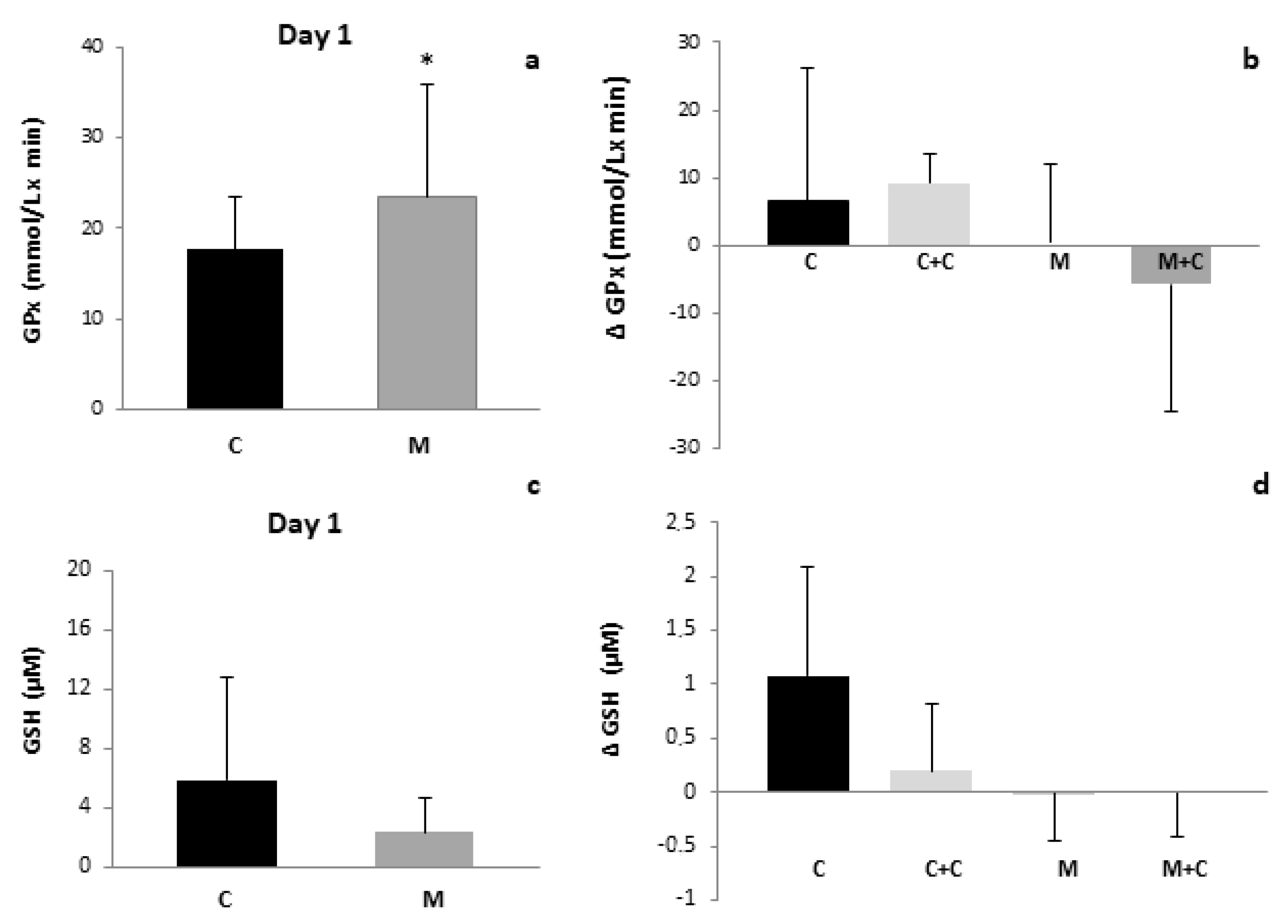
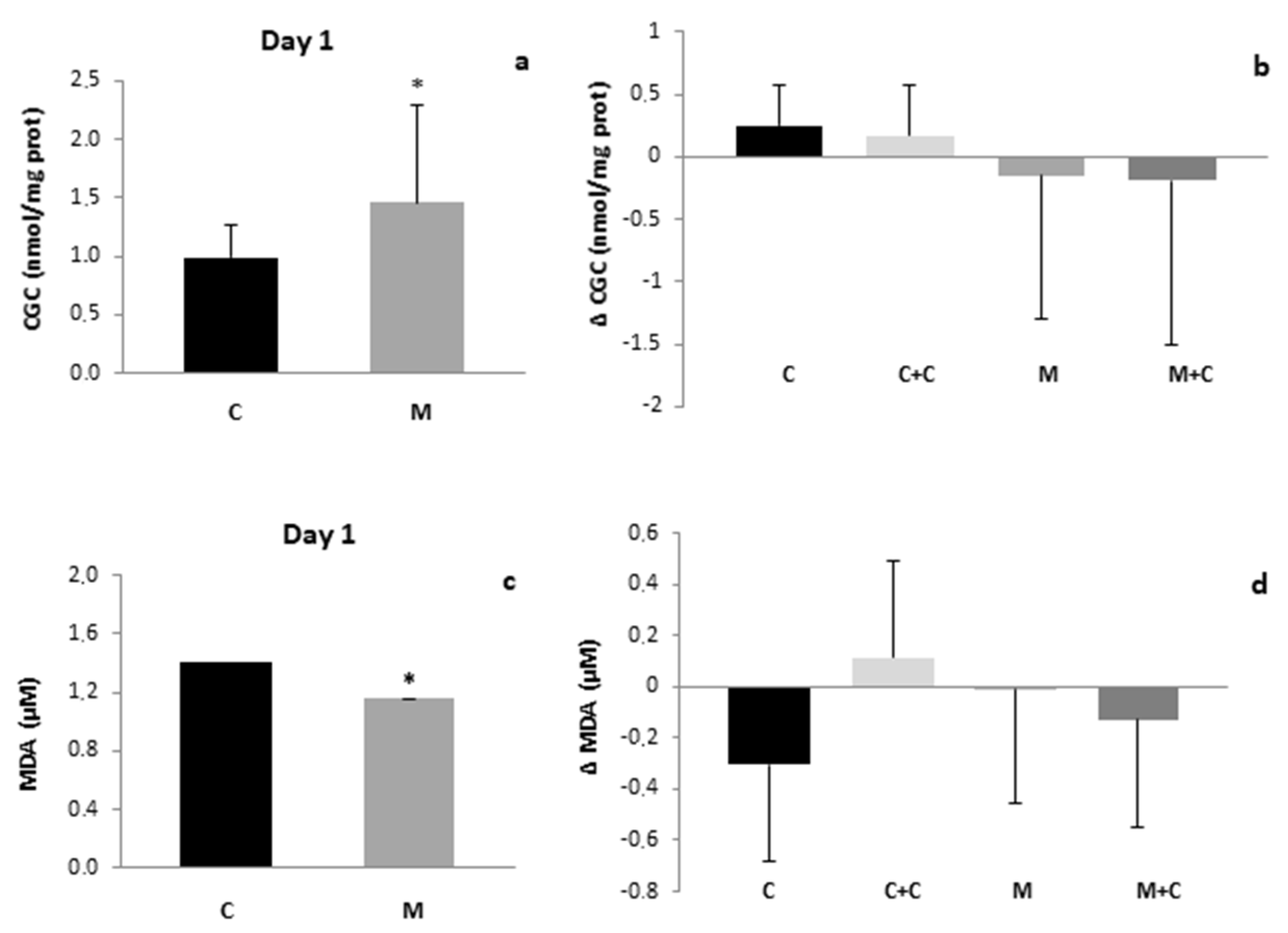
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fanaro, S. The biological specificity and superiority of human milk. Scientific basis, guarantees and safety controls. Minerva Pediatr. 2002, 54, 113–129. [Google Scholar]
- Ojo-Okunola, A.; Nicol, M.; du Toit, E. Human breast milk bacteriome in health and disease. Nutrients 2018, 10, 1643. [Google Scholar] [CrossRef] [Green Version]
- WHO. Healthy Diet. Fact Sheet, nº 394; WHO: Geneva, Switzerland, August 2018. [Google Scholar]
- Abbe, M.R.; Friel, J.K.J. Superoxide dismutase and glutathione peroxidase content of human milk from mothers of premature and full-term infants during the first 3 months of lactation. Gastroenterol. Nutr. 2000, 31, 174–270. [Google Scholar]
- Turoli, D.; Testolin, G.; Zanini, R.; Bellù, R. Determination of oxidative status in breast and formula milk. Acta Paediatr. 2004, 93, 1569–1574. [Google Scholar] [CrossRef]
- Hanson, C.; Lyden, E.; Furtado, J.; Van Ormer, M.; Anderson-Berry, A. A comparison of nutritional antioxidant content in breast milk, donor milk, and infant formulas. Nutrients 2016, 8, 681. [Google Scholar] [CrossRef] [Green Version]
- Tsopmo, A. Phytochemicals in human milk and their potential antioxidative protection. Antioxidants 2018, 7, 32. [Google Scholar] [CrossRef] [Green Version]
- Silvestre, D.; Miranda, M.; Muriach, M.; Almansa, I.; Jareño, E.; Romero, F.J. Frozen breast milk at −20 °C and −80 °C: A longitudinal study of glutathione peroxidase activity and malondialdehyde concentration. J. Hum. Lactation 2010, 26, 35–41. [Google Scholar] [CrossRef]
- Silvestre, D.; Miranda, M.; Muriach, M.; Almansa, I.; Jareno, E.; Romero, F.J. Antioxidant capacity of human milk: Effect of thermal conditions for the pasteurization. Acta Paediatr. Int. J. Paediatr. 2008, 97, 1070–1074. [Google Scholar] [CrossRef]
- Miranda, M.; Muriach, M.; Almansa, I.; Jareño, E.; Bosch-Morell, F.; Romero, F.J.; Silvestre, D. Oxidative status of human milk and its variations during cold storage. BioFactors 2004, 20, 129–137. [Google Scholar] [CrossRef]
- Codoñer-Franch, P.; Hernández-Aguilar, M.T.; Navarro-Ruiz, A.; López-Jaén, A.B.; Borja-Herrero, C.; Valls-Bellés, V. Diet supplementation during early lactation with non-alcoholic beer increases the antioxidant properties of breastmilk and decreases the oxidative damage in breastfeeding mothers. Breastfeed. Med. 2013, 8, 164–169. [Google Scholar] [CrossRef]
- Mutinati, M.; Pantaleo, M.; Roncetti, M.; Piccinno, M.; Rizzo, A.; Sciorsci, R.L. Oxidative stress in neonatology: A review. Reprod. Domest. Anim. 2014, 49, 7–16. [Google Scholar] [CrossRef]
- Ledo, A.; Arduini, A.; Asensi, M.A.; Sastre, J.; Escrig, R.; Brugada, M.; Aguar, M.; Saenz, P.; Vento, M. Human milk enhances antioxidant defenses against hydroxyl radical aggression in preterm infants. Am. J. Clin. Nutr. 2009, 89, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; Davis, J.M. Future applications of antioxidants in premature infants. Curr. Opin. Pediatr. 2011, 23, 161–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkinson, J.R.C.; Hyde, M.J.; Gale, C.; Santhakumaran, S.; Modi, N. Preterm birth and the metabolic syndrome in adult life: A systematic review and meta-analysis. Pediatrics 2013, 131, e1240–e1263. [Google Scholar] [CrossRef] [Green Version]
- Lapillonne, A.; Griffin, I.J. Feeding preterm infants today for later metabolic and cardiovascular outcomes. J. Pediatr. 2013, 162, S7. [Google Scholar] [CrossRef] [PubMed]
- Gila-Diaz, A.; Arribas, S.M.; Algara, A.; Martín-Cabrejas, M.A.; de Pablo, Á.L.L.; de Pipaón, M.S.; Ramiro-Cortijo, D. A review of bioactive factors in human breastmilk: A focus on prematurity. Nutrients 2019, 11, 1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, H. Benign breast disorders: The clinician’s view. Cancer Detect. Prev. 1992, 16, 1–5. [Google Scholar]
- Contreras, G.A.; Rodríguez, J.M. Mastitis: Comparative etiology and epidemiology. J. Mammary Gland Biol. Neoplasia 2011, 16, 339–356. [Google Scholar] [CrossRef]
- Patel, S.H.; Vaidya, Y.H.; Patel, R.J.; Pandit, R.J.; Joshi, C.G.; Kunjadiya, A.P. Culture independent assessment of human milk microbial community in lactational mastitis. Sci. Rep. 2017, 7, 7804. [Google Scholar] [CrossRef] [Green Version]
- Jahanfar, S.; Ng, C.J.; Teng, C.L. Antibiotics for mastitis in breastfeeding women. Sao Paulo Med. J. 2016, 134, 273. [Google Scholar] [CrossRef] [Green Version]
- Bond, D.M.; Morris, J.M.; Nassar, N. Study protocol: Evaluation of the probiotic Lactobacillus Fermentum CECT5716 for the prevention of mastitis in breastfeeding women: A randomised controlled trial. BMC Pregnancy Childbirth 2017, 17, 148. [Google Scholar] [CrossRef] [Green Version]
- Johnson, H.M.; Mitchell, K.B. Lactational phlegmon: A distinct clinical entity affecting breastfeeding women within the mastitis-abscess spectrum. Breast J. 2020, 26, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Gelasakis, A.I.; Angelidis, A.S.; Giannakou, R.; Filioussis, G.; Kalamaki, M.S.; Arsenos, G. Bacterial subclinical mastitis and its effect on milk yield in low-input dairy goat herds. J. Dairy Sci. 2016, 99, 3698–3708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Gao, J.; Ali, T.; Yu, D.; Zhang, S.; Khan, S.U.; Fanning, S.; Han, B. Characteristics of Aerococcus viridans isolated from bovine subclinical mastitis and its effect on milk SCC, yield, and composition. Trop. Anim. Health Prod. 2017, 49, 843–849. [Google Scholar] [CrossRef]
- Silanikove, N.; Merin, U.; Shapiro, F.; Leitner, G. Subclinical mastitis in goats is associated with upregulation of nitric oxide-derived oxidative stress that causes reduction of milk antioxidative properties and impairment of its quality. J. Dairy Sci. 2014, 97, 3449–3455. [Google Scholar] [CrossRef]
- Samuel, T.M.; De Castro, C.A.; Dubascoux, S.; Affolter, M.; Giuffrida, F.; Billeaud, C.; Picaud, J.-C.; Agosti, M.; Al-Jashi, I.; Barroso Pereira, A.; et al. Subclinical Mastitis in a European Mul-ticenter Cohort: Prevalence, Impact on Human Milk (HM) Composition, and Association with Infant HM Intake and Growth. Nutrients 2020, 12, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Say, B.; Dizdar, E.A.; Degirmencioglu, H.; Uras, N.; Sari, F.N.; Oguz, S.; Canpolat, F.E. The effect of lactational mastitis on the macronutrient content of breast milk. Early Hum. Dev. 2016, 98, 7–9. [Google Scholar] [CrossRef]
- Perez, M.; Ladero, V.; Redruello, B.; Del Rio, B.; Fernandez, L.; Rodriguez, J.M.; Martín, M.C.; Fernandez, M.; Alvarez, M.A. Mastitis modifies the biogenic amines profile in human milk, with significant changes in the presence of histamine, putrescine and spermine. PLoS ONE 2016, 11, e0162426. [Google Scholar] [CrossRef]
- Buescher, E.S.; Hair, P.S. Human milk anti-inflammatory component contents during acute mastitis. Cell. Immunol. 2001, 210, 87–95. [Google Scholar] [CrossRef]
- Nardi, G.M.; De Farias Januário, A.G.; Freire, C.G.; Megiolaro, F.; Schneider, K.; Perazzoli, M.R.A.; Do Nascimento, S.R.; Gon, A.C.; Mariano, L.N.B.; Wagner, G.; et al. Anti-inflammatory activity of berry fruits in mice model of inflammation is based on oxidative stress modulation. Pharmacogn. Res. 2016, 8, S42–S49. [Google Scholar] [CrossRef] [Green Version]
- Grace, M.H.; Esposito, D.; Dunlap, K.L.; Lila, M.A. Comparative analysis of phenolic content and profile, antioxidant capacity, and anti-inflammatory bioactivity in wild alaskan and commercial vaccinium berries. J. Agric. Food Chem. 2014, 62, 4007–4017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, N.J.; Rice-Evans, C.A. Factors influencing the antioxidant activity determined by the ABTS•+ radical cation assay. Free Radic. Res. 1997, 26, 195–199. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Lawrence, R.A.; Parkhill, L.K.; Burk, R.F. Hepatic cytosolic non selenium-dependent glutathione peroxidase activity: Its nature and the effect of selenium deficiency. J. Nutr. 1978, 108, 981–987. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.J.; Babson, J.R.; Beatty, P.W.; Brodie, A.E.; Ellis, W.W.; Potter, D.W. High-performance liquid chromatography analysis of nanomole levels of glutathione, glutathione disulfide, and related thiols and disulfides. Anal. Biochem. 1980, 106, 55–62. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A.J. Colorimetry to total phenolics with phosphomolybdic acid reagents. Am. J. Enol. Vinic. 1965, 16, 144–158. [Google Scholar]
- Romero, M.J.; Bosch-Morell, F.; Romero, B.; Rodrigo, J.M.; Serra, M.A.; Romero, F.J. Serum malondialdehyde: Possible use for the clinical management of chronic hepatitis C patients. Free Radic. Biol. Med. 1998, 25, 993–997. [Google Scholar] [CrossRef]
- Richard, M.J.; Guiraud, P.; Meo, J.; Favier, A. High-performance liquid chromatographic separation of malondialdehyde-thiobarbituric acid adduct in biological materials (plasma and human cells) using a commercially available reagent. J. Chromatogr. B Biomed. Sci. Appl. 1992, 577, 9–18. [Google Scholar] [CrossRef]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.G.; Ahn, B.W.; Shaltiel, S.; Stadtman, E.R. Determination of Carbonyl Content in Oxidatively Modified Proteins. Methods Enzymol. 1990, 186, 464–478. [Google Scholar] [CrossRef]
- Calder, P.C. Very long-chain n-3 fatty acids and human health: Fact, fiction and the future. Proc. Nutr. Soc. 2018, 77, 52–72. [Google Scholar] [CrossRef] [Green Version]
- Steffl, M.; Kinkorova, I.; Kokstejn, J.; Petr, M. Macronutrient intake in soccer players—A meta-analysis. Nutrients 2019, 11, 1305. [Google Scholar] [CrossRef] [Green Version]
- Espinosa-Martos, I.; Jiménez, E.; De Andrés, J.; Rodríguez-Alcalá, L.M.; Tavárez, S.; Manzano, S.; Fernández, L.; Alonso, E.; Fontecha, J.; Rodríguez, J.M. Milk and blood biomarkers associated to the clinical efficacy of a probiotic for the treatment of infectious mastitis. Benef. Microbes 2016, 7, 305–318. [Google Scholar] [CrossRef] [Green Version]
- Sadek, K.; Saleh, E.; Ayoub, M. Selective, reliable blood and milk bio-markers for diagnosing clinical and subclinical bovine mastitis. Trop. Anim. Health Prod. 2017, 49, 431–437. [Google Scholar] [CrossRef]
- Alba, D.F.; da Rosa, G.; Hanauer, D.; Saldanha, T.F.; Souza, C.F.; Baldissera, M.D.; da Silva dos Santos, D.; Piovezan, A.P.; Girardini, L.K.; Schafer Da Silva, A. Subclinical mastitis in Lacaune sheep: Causative agents, impacts on milk production, milk quality, oxidative profiles and treatment efficacy of ceftiofur. Microb. Pathog. 2019, 137, 103732. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Chi, X.; Ma, X.; Xu, W.; Shi, F.; Hu, S. Anti-inflammatory mechanism of ginsenoside Rg1: Proteomic analysis of milk from goats with mastitis induced with lipopolysaccharide. Int. Immunopharmacol. 2019, 71, 382–391. [Google Scholar] [CrossRef]
- Baliño, P.; Romero-Cano, R.; Sánchez-Andrés, J.V.; Valls, V.; Aragón, C.G.; Muriach, M. Effects of Acute Ethanol Administration on Brain Oxidative Status: The Role of Acetaldehyde. Alcohol. Clin. Exp. Res. 2019, 43, 1672–1681. [Google Scholar] [CrossRef]
- Fromenty, B.; Vadrot, N.; Massart, J.; Turlin, B.; Barri-Ova, N.; Lettéron, P.; Fautrel, A.; Robin, M.A. Chronic ethanol consumption lessens the gain of body weight, liver triglycerides, and diabetes in obese ob/ob mice. J. Pharmacol. Exp. Ther. 2009, 331, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Fetherston, C.M.; Lai, C.T.; Hartmann, P.E. Relationships between symptoms and changes in breast physiology during lactation mastitis. Breastfeed. Med. Off. J. Acad. Breastfeed. Med. 2006, 1, 136–145. [Google Scholar] [CrossRef]
- Kruzel, M.L.; Zimecki, M.; Actor, J.K. Lactoferrin in a context of inflammation-induced pathology. Front. Immunol. 2017, 8, 1438. [Google Scholar] [CrossRef]
- Han, N.; Li, H.; Li, G.; Shen, Y.; Fei, M.; Nan, Y. Effect of bovine lactoferrin as a novel therapeutic agent in a rat model of sepsis-induced acute lung injury. AMB Express 2019, 9, 177. [Google Scholar] [CrossRef] [Green Version]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W.G. Selenium: Biochemical role as a component of glatathione peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef]
- Song, B.J.; Jouni, Z.E.; Ferruzzi, M.G. Assessment of phytochemical content in human milk during different stages of lactation. Nutrition 2013, 29, 195–202. [Google Scholar] [CrossRef]
- Li, W.; Hosseinian, F.S.; Tsopmo, A.; Friel, J.K.; Beta, T. Evaluation of antioxidant capacity and aroma quality of breast milk. Nutrition 2009, 25, 105–114. [Google Scholar] [CrossRef]
- Dimri, U.; Sharma, M.C.; Singh, S.K.; Kumar, P.; Jhambh, R.; Singh, B.; Bandhyopadhyay, S.; Verma, M.R. Amelioration of altered oxidant/antioxidant balance of Indian water buffaloes with subclinical mastitis by vitamins A, D3, E, and H supplementation. Trop. Anim. Health Prod. 2013, 45, 971–978. [Google Scholar] [CrossRef]
- Li, C.; Feng, J.; Huang, W.Y.; An, X.T. Composition of polyphenols and antioxidant activity of rabbiteye blueberry (Vaccinium ashei) in Nanjing. J. Agric. Food Chem. 2013, 61, 523–531. [Google Scholar] [CrossRef]
- Franke, A.A.; Halm, B.M.; Custer, L.J.; Tatsumura, Y.; Hebshi, S. Isoflavones in breastfed infants after mothers consume soy. Am. J. Clin. Nutr. 2006, 84, 406–413. [Google Scholar] [CrossRef]
- Jochum, F.; Alteheld, B.; Meinardus, P.; Dahlinger, N.; Nomayo, A.; Stehle, P. Mothers’ Consumption of Soy Drink but Not Black Tea Increases the Flavonoid Content of Term Breast Milk: A Pilot Randomized, Controlled Intervention Study. Ann. Nutr. Metab. 2017, 70, 147–153. [Google Scholar] [CrossRef]
- Dai, H.; Coleman, D.N.; Hu, L.; Martinez-Cortés, I.; Wang, M.; Parys, C.; Shen, X.; Loor, J.J. Methionine and arginine supplementation alter inflammatory and oxidative stress responses during lipopolysaccharide challenge in bovine mammary epithelial cells in vitro. J. Dairy Sci. 2020, 103, 676–689. [Google Scholar] [CrossRef]
- Alhalwani, A.Y.; Davey, R.L.; Kaul, N.; Barbee, S.A.; Alex Huffman, J. Modification of lactoferrin by peroxynitrite reduces its antibacterial activity and changes protein structure. Proteins Struct. Funct. Bioinform. 2020, 88, 166–174. [Google Scholar] [CrossRef]
- Eslami, H.; Batavani, R.A.; Asr I-Rezaei, S.; Hobbenaghi, R. Changes of stress oxidative enzymes in rat mammary tissue, blood and milk after experimental mastitis induced by E. coli lipopolysaccharide. Vet. Res. Forum Int. Q. J. 2015, 6, 131–136. [Google Scholar]
- Nedić, S.; Vakanjac, S.; Samardžija, M.; Borozan, S. Paraoxonase 1 in bovine milk and blood as marker of subclinical mastitis caused by Staphylococcus aureus. Res. Vet. Sci. 2019, 125, 323–332. [Google Scholar] [CrossRef]
- Abouzed, T.K.; Sadek, K.M.; Ayoub, M.M.; Saleh, E.A.; Nasr, S.M.; El-Sayed, Y.S.; Shoukry, M. Papaya extract upregulates the immune and antioxidants-related genes, and proteins expression in milk somatic cells of Friesian dairy cows. J. Anim. Physiol. Anim. Nutr. 2019, 103, 407–415. [Google Scholar] [CrossRef]
- Mukherjee, R. Selenium and vitamin E increases polymorphonuclear cell phagocytosis and antioxidant levels during acute mastitis in riverine buffaloes. Vet. Res. Commun. 2008, 32, 305–313. [Google Scholar] [CrossRef]
- Politis, I.; Theodorou, G.; Lampidonis, A.D.; Kominakis, A.; Baldi, A. Short communication: Oxidative status and incidence of mastitis relative to blood α-tocopherol concentrations in the postpartum period in dairy cows. J. Dairy Sci. 2012, 95, 7331–7335. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | C + C | M | M + C | |
|---|---|---|---|---|
| Age | 36.5 ± 4.1 | 33.8 ± 3.7 | 33.7 ± 3.9 | 36.4 ± 3.3 |
| Weight (kg) | 60.6 ± 7.2 | 64.6 ± 8.8 | 66.2 ± 10.5 | 63.9 ± 10.5 |
| Height (cm) | 164.6 ± 4.7 | 165.5 ± 6.2 | 163.2 ± 4.8 | 164.6 ± 6.7 |
| BMI | 22.3 ± 2.5 | 23.6 ± 2.6 | 24.8 ± 3.9 | 23.6 ± 4 |
| Kcal | 1595 ± 394 | 1510 ± 265 | 1659 ± 248 | 1631 ± 428 |
| Carbohydrates (g) | 166.2 ± 54.5 | 167.8 ± 48 | 191.2 ± 44.7 | 188.6 ± 40 |
| Proteins (g) | 73.8 ± 15.9 | 71.6 ± 16 | 82.7 ± 12.5 | 80.8 ± 19.6 |
| Fat (g) | 77.6 ± 23 | 67 ± 11.2 | 68.9 ± 16.7 | 68.2 ± 30.1 |
| Cholesterol (mg) | 227.5 ± 76.3 | 238.7 ± 109.2 | 249.7 ± 90.6 | 231.8 ± 86.1 |
| Fiber (g) | 12.4 ± 6 | 9.5 ± 3.5 | 14.6 ± 7.5 | 11.5 ± 5.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valls-Bellés, V.; Abad, C.; Hernández-Aguilar, M.T.; Nacher, A.; Guerrero, C.; Baliño, P.; Romero, F.J.; Muriach, M. Human Milk Antioxidative Modifications in Mastitis: Further Beneficial Effects of Cranberry Supplementation. Antioxidants 2022, 11, 51. https://doi.org/10.3390/antiox11010051
Valls-Bellés V, Abad C, Hernández-Aguilar MT, Nacher A, Guerrero C, Baliño P, Romero FJ, Muriach M. Human Milk Antioxidative Modifications in Mastitis: Further Beneficial Effects of Cranberry Supplementation. Antioxidants. 2022; 11(1):51. https://doi.org/10.3390/antiox11010051
Chicago/Turabian StyleValls-Bellés, Victoria, Cristina Abad, María Teresa Hernández-Aguilar, Amalia Nacher, Carlos Guerrero, Pablo Baliño, Francisco J. Romero, and María Muriach. 2022. "Human Milk Antioxidative Modifications in Mastitis: Further Beneficial Effects of Cranberry Supplementation" Antioxidants 11, no. 1: 51. https://doi.org/10.3390/antiox11010051