
Gestational Diabetes Mellitus: The Crosslink among Inflammation, Nitroxidative Stress, Intestinal Microbiota and Alternative Therapies

 ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Methods
2.1. MESH Terms
2.1.1. GDM
2.1.2. Inflammatory Aspects in GDM
2.1.3. Nitroxidative Stress in GDM
2.1.4. Intestinal Microbiota and GDM
2.1.5. Alternative Therapies in GDM
Randomized Clinical Trials
Experimental Studies in Animal Models
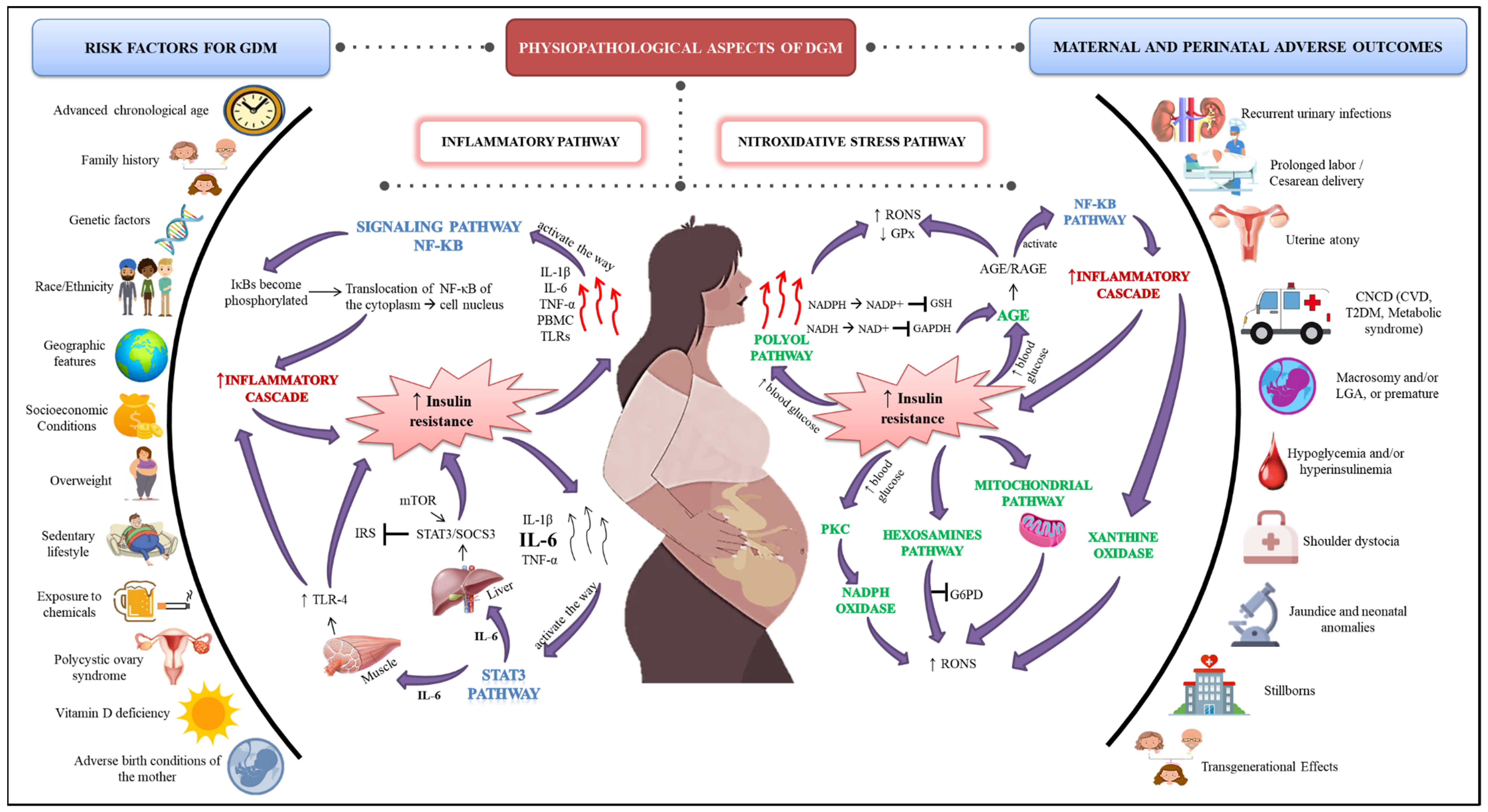
3. GDM
| Global Prevalence | Prevalence by Region | ||
| Hyperglycemia in pregnancy | 16.2% | Africa | 10.4% |
| Western Pacific | 12.6% | ||
| South America and Central America | 13.1% | ||
| North America and Caribbean | 14.6% | ||
| Europe | 16.2% | ||
| Middle East and North Africa | 21.8% | ||
| South East Asia | 24.2% | ||
| Source: Adapted from International Diabetes Federation [19]. | |||
3.1. Screening and Diagnosis
| Criteria | Time Course | Fasting Glucose (mg/dL) | Glucose Overload | Oral Glucose Tolerance Test (mg/dL) | ||
| 1 h | 2 h | 3 h | ||||
| O’Sullivan & Mahan (1964) [21] | Detected at any time during pregnancy | 90 | 100 g of glucose | 165 | 145 | 125 |
| O’Sullivan & Mahan (1964) [21] adapted by National Diabetes Data Group (NDDG) (1979) [22] | 105 | 100 g of glucose | 190 | 165 | 145 | |
| Carpenter & Coustan (1982) [23] | 95 | 100 g of glucose | 180 | 155 | 140 | |
| World Health Organization (WHO) (1999) [26] | 126 | 75 g of Glucose | Not measured | 140 | Not measured | |
| International Association of Diabetes in Pregnancy Study Group (IADPSG (2010) [27] | 24–28 gestational weeks | 92 | 75 g of Glucose | 180 | 153 | |
3.2. Etiology
| Risk Factor | Mechanism of Action | Reference |
| Advanced chronological age (>35 years old) |
| [38,39] |
| Family history |
| [40,41] |
| Genetic factors |
| [40,41,42] |
| [40,42,43] | |
| Race/ Ethnicity |
| [44,45] |
| Geographic features |
| [28,32] |
| Socio-economic |
| [45,46] |
| Overweight |
| [3,47] |
| [48,49] | |
| Westernized diet |
| [37,50] |
| Sedentary lifestyle |
| [51,52] |
| Exposure to chemicals |
| [53,54,55,56,57,58] |
| [59,60,61] | |
| Polycystic Ovary Syndrome (POS) |
| [62,63] |
| Vitamin D Deficiency |
| [64,65] |
| Adverse birth conditions of the mother (Fetal program) |
| [66,67,68,69,70] |
3.3. Maternal and Perinatal Outcomes in the GDM
4. Inflammatory Aspects in GDM
4.1. NF-kB Signaling Pathway
4.2. STAT3 Signaling Pathway
5. Nitroxidative Stress in GDM
5.1. Polyol Pathway
5.2. AGEs
5.3. PKC
5.4. Hexosamine Pathway
5.5. NADPH Oxidase
5.6. Xanthine Oxidase
5.7. Mitochondrial Pathway
6. Intestinal Microbiota and GDM

{kind=link}
{kind=link}
| Source Sample | Population | Size * | Supplementation | Dose/Duration | Main Findings |
|---|---|---|---|---|---|
| Karamali et al. (2016) [118] | Iran | I: 30 C: 30 | L. acidophilus + L. casei + B. bifidum | 2 × 109 CFU/ 6 weeks | Supplementation with probiotics ↓FBG, serum insulin, TG, and VLDL-c, and improved insulin resistance indexes. |
| Hajifaraji et al. (2018) [150] | Iran | I: 27 C: 29 | L. acidophilus LA-5 + B. BB-12 + S. thermophilus STY-31 + L. delbrueckii bulgaricus + LBY-27 | >4 × 109 CFU/ 8 weeks | Supplementation with probiotics significantly ↓CRP and TNF-α. MDA, GPx and GR in women in the intervention group. |
| Kijmanawat et al. (2019) [153] | Thailand | I: 28 C: 29 | Bifidobacterium + Lactobacillus | 2 × 109 CFU/ 4 weeks | In women with diet-controlled GDM, supplementation with probiotics ↓FBG and insulin resistance compared with the control. |
| Babadi et al. (2018) [154] | Iran | I: 24 C: 24 | L. casei + B. bifidum + L. fermentum + L. acidophilus | 2 × 109 CFU/ 6 weeks | Probiotic supplementation improved the expression of genes related to insulin; glycemic control; inflammation; lipid profile, and oxidative stress markers, such as ↓MDA and ↑TAC, compared with the control. |
| Badehnoosh et al. (2018) [155] | Iran | I: 30 C: 30 | L. acidophilus + L. casei + B. bifidum | 2 × 109 CFU/ 6 weeks | Probiotic supplementation improved FBG, and CRP, ↑TAC, and ↓MDA, without affecting pregnancy outcomes. |
| Nabhani et al. (2018) [156] | Iran | I: 45 C: 45 | L. acidophilus + L. plantarum + L. fermentum + L. gasseri + FOS | 1.5–7.0 × 109–10 CFU + 38.5 mg/ 6 weeks | Symbiotics had no effect on FBG and insulin resistance/sensitivity indexes. However, an ↑ in HDL-c and TAC was seen, and a ↓ was seen in blood pressure in the intervention group. |
| Jamilian et al. (2019) [157] | Iran | I: 29 C: 28 | L. acidophilus + B. bifidum + L. reuteri + L. fermentum + Vitamin D | 8 × 109 CFU/ 6 weeks +50.000 UI every 2 weeks | ↓FBG, serum insulin, CRP, and MDA; ↑TAC and GSH; and improved insulin resistance scores. |
| Karamali et al. (2018) [158] | Iran | I: 30 C: 30 | L. acidophilus + L. casei + B. bifidum + Inulin | 2 × 109 CFU/ 6 weeks +800 mg | Symbiotic supplementation ↓CRP and MDA; ↑TAC and GSH; and↓ the rates of cesarean section, hyperbilirubinemia and hospitalization in NB, without affecting other pregnancy outcomes. |
| Ahmadi et al. (2016) [159] | Iran | I: 35 C: 35 | L. acidophilus + L. casei + B. bifidum + inulin | 2 × 109 CFU/ 6 weeks +800 mg | Symbiotics ↑ insulin metabolism markers, and the insulin sensitivity index as well as ↓VLDL-c and TG. |
| Jafarnejad et al. (2016) [160] | Iran | I: 41 C: 41 | S. thermophilus + B. breve + B. longum + B. infantis + L. acidophilus + L. plantarum + L. paracasei + L. delbrueckii subsp. Bulgaricus | 15 × 109 CFU/ 8 weeks | No differences were observed in FBG, glycated hemoglobin, serum insulin, and insulin resistance indices. However, ↓CRP, IL-6, and TNF-α were observed, without changes in IL-10 and IFN-γ. |
| Dolatkhah et al. (2015) [161] | Turkey | I: 29 C: 27 | L. acidophilus LA-5 + B. BB-12 + Streptococcus thermophilus + STY-31 + L. delbrueckii bulgaricus LBY-27 | >4 × 109 CFU/ 8 weeks | ↓FBG and insulin resistance index, and less weight gain in those in the intervention group. |
| Lindsay et al. (2015) [162] | Ireland | I: 74 C: 75 | L. salivarius | 1 × 109 CFU/ 6 weeks | No beneficial effect on glycemic control or pregnancy outcomes. ↓ in total and LDL-c in the supplemented group. |
7. Alternative Therapies in GDM
7.1. Randomized Clinical Trials
| Source | Type of Study | Population | Sample Size * | Intervention | Main Findings |
|---|---|---|---|---|---|
| Karamali et al. (2020) [165] | Randomized clinical trial, double blind | Teerã, Iran | I: 18 C: 18 | Selenium supplementation (200 μg/day) | ↑expression of PPAR-γ and GLUT-1, but did not affect the gene expression of LDLR and LP(a) (p < 0.05), compared with the control. |
| Gomez et al. (2020) [166] | Randomized clinical trial, double blind | Buenos Aires, Argentina | I: 15 C: 15 | 3 tablespoons of extra virgin olive oil daily | The placenta of pregnant women showed regulation on PPARα expression and pro-inflammatory markers (IL-1β and TNF-α), and ↑the expression of miR-518d (p < 0.05), compared with the control. |
| Gunasegaran et al. (2020) [176] | Randomized clinical trial, double blind | Puducherry, India | I1: 34 I2: 36 | I1: Vit D (1000 UI/day) + calcium (1000 mg/day) I2: Vit D (250 UI/day) + calcium (500 mg) | I1↓the serum levels of glucose, insulin, LDL-c, and total cholesterol and ↑the levels of HDL-c and total GSH (p < 0.05), compared with I2. |
| Jamilian et al. (2020) [179] | Randomized clinical trial, double blind | Arak, Iran | I: 30 C: 30 | 2 flaxseed oil (1000 mg) capsules/day, containing 400 mg of α-linolenic acid | Seriously ↓ levels of glucose, insulin, insulin resistance, VLDL-c, total cholesterol, CRP, and MDA, and ↑ sensitivity to insulin, nitrite, and GSH (p < 0.05), compared with the control. |
| Hajimoosayi et al. (2020) [174] | Randomized clinical trial, double blind | Teerã, Iran | I: 37 C: 33 | 3 ginger tablets/day (1500 mg), with consumption after main meals | ↓serum levels of blood glucose, insulin, and the HOMA-IR index (p < 0.05), compared with control. |
| Yang et al. (2019) [178] | Randomized clinical trial, double blind | Changchun, China | I: 268 C: 270 | 1 capsule/day of cod liver oil (500 mg) | ↓serum levels of glucose, lipids, CRP, HOMA-IR, and adverse perinatal outcomes (p < 0.05) compared with the control. |
| Ostadmohammadi et al. (2019) [177] | Randomized clinical trial, double blind | Kashan, Iran | I: 27 C: 27 | Zinc co-supplementation (zinc gluconate 233 mg/day) + Vit E (400 IU/day) | Supplemented group ↓ the serum levels of HOMA-IR insulin, total cholesterol, LDL-c, and QUICKI (p < 0.05), compared with the control group. It positively regulated the gene expression of PPAR-γ and LDLR (p < 0.05), compared with the control. |
| Jamilian et al., (2018) [167] | Randomized clinical trial, double blind | Arak, Iran | I: 20 C: 20 | 2 capsules/day of fish oil (1000 mg), containing 180 mg EPA + 120 mg DHA | ↑ expression of PPAR-γ, IL-1, and TNF-α (p < 0.05), compared with the control. |
| Karamali et al. (2018) [172] | Randomized clinical trial, double blind | Teerã, Iran | I: 30 C: 30 | Magnesium–zinc–calcium–vitamin D co-supplementation | Co-supplementation: ↓ serum glucose, insulin, HOMA-IR, TG, and VLDL-c, while increasing the QUICKI index (p < 0.05) compared with the control. |
| Gao et al., 2017 [181] | Randomized clinical trial, double blind | Hebei, China | I: 123 C: 121 | Enrichment of margarine with phytosterols (every 10 g of margarine, 2 g of phytosterols, twice a day) | Supplementation ↑ maternal and neonatal outcomes in patients with GDM. |
| Zhang et al., 2017 [182] | Randomized clinical trial, double blind | Shandong, China | I: 176 C: 150 | 1 capsule/day of EGCG (500 mg) | Supplementation ↑ maternal diabetes parameters and ↓ risk of neonatal complications (p < 0.05), compared with the control. |
| Yuan et al. 2016 [169] | Randomized clinical trial | Chongqing, China | I: 20 C: 22 | Pepper with capsaicin (5 mg/day) | Supplementation ↓ serum concentrations of glucose, insulin, HOMA-IR, OGTT, total cholesterol, TG, and the risk for LGA births and ↑ the serum levels of apolipoprotein B and CGRP (p < 0.05), compared with the control. |
| Jamilian et al. 2016 [173] | Randomized clinical trial | Arak, Iran | I: 30 C: 30 | 1 capsule/day formulated with vit D3 (1000 IU) + evening primrose oil (1000 mg) | Supplementation ↓ serum levels of glucose, insulin, HOMA-IR, QUICKI, VLDL-c, LDL-c, and total cholesterol (p < 0.05), compared with the control. |
| Sun et al. 2016 [170] | Randomized clinical trial | Shandong, China | I: 64 C: 65 | 2 Artemisia extract tablets/day (200 mg) | Supplementation ↑ insulin sensitivity through upregulation of adiponectin. |
| Asemi et al. 2015 [180] | Randomized clinical trial, double blind | Kashan, Iran | I: 35 C: 35 | 1 tablet/day of selenium (200 μg) | Significantly ↓ serum glucose, insulin, HOMA-IR, CRP, MDA, and plasma GSH concentrations (p < 0.05), compared with the control. |
| Fei et al. 2014 [168] | Randomized clinical trial | Suzhou, China | I: 46 C: 51 | Soy oligosaccharide (10 g/day) in 200–300 mL of warm water, given orally before bedtime | Soybean oligosaccharide: ↑ serum activities of SOD, CAT, and GPx, while ↓ serum levels of TBARS and HOMA-IR (p < 0.05), compared with the control. |
| Asemi et al. 2014 [175] | Randomized clinical trial | Kashan, Iran | I: 25 C: 26 | Co-supplementation of calcium (1000 mg/day), and Vit D3 (50,000 U, in two moments, on the 1st and 21st days of the study) | Co-supplemented: significantly ↓ serum concentrations of glucose, insulin, HOMA-IR, LDL-c, and total cholesterol and ↑ in HDL-c levels; it was possible to avoid the increase in MDA, compared with the control. |
| Asemi et al. 2013 [171] | Randomized clinical trial | Kashan, Iran | I: 16 C: 16 | DASH diet (rich in fruits, vegetables, whole grains, and low-fat dairy but low in total fat, saturated, cholesterol, refined grains, and sweets, with a total of 2400 mg/day Na) | The intervention group with DASH diet ↓plasma concentrations of glucose, insulin, and HOMA-IR, (p < 0.05), compared with the control. |
7.2. Experimental Studies in Animal Models
| Source | Study Type | Animal Model | Diabetogenic Drug | Intervention | Main Findings |
|---|---|---|---|---|---|
| Yessoufou et al. (2013) [184] | Experimental study | Wistar rats | Streptozotocin (20 mg/kg) | I1: Picralima nitida (seeds); I2: Nauclea latifolia (root and stem); I3: Oxytenanthera abyssinica (leaves). All of these extracts were injected intraperitoneally at a concentration of 25 mg/kg (12–15 days) | Groups I1, I2, and I3 showed attenuation of GDM-induced hyperglycemia in pregnant rats. Among the three groups, the extract from group I1 exhibited ↑antioxidant capacity compared with the others. |
| Du et al. (2020) [185] | Experimental study | Wistar rats albinos | Streptozotocin (45 mg/kg) | Nano resveratrol-zinc oxide complex, encapsulated with chitosan (200 mg/kg), daily for 28 days | Supplementation: ↓serum glucose levels, maintained the lipid content compared with the control; ↓inflammatory factors (IL-6) and endoplasmic reticulum stress (GRP78, p-IRE1α, p-eIF2α, and p-PERK). |
| Usman et al. (2018) [186] | Experimental study | Sprague Dawley rats | Streptozotocin (60 mg/kg) | Propolis (Heterotrigona itama) orally: 300 mg/kg/day subcutaneously: 5.0 IU/kg/day | Treatment: ↓serum glucose and MDA levels, similar to the insulin-treated group. |
| Badr et al. (2013) [187] | Experimental study | Swiss albino mice | Streptozotocin (50 mg/kg) | Thymoquinone (20 mg/kg/day) | Supplementation significantly restored serum levels of glucose, insulin, reactive oxygen species, proinflammatory cytokines (IL-1β, IL-6, and TNF-α) and lipids, as well as lymphocyte proliferation in offspring. |
| Afiune et al. (2017) [188] | Experimental study | Wistar rats | Streptozotocin (40 mg/kg) | Oral aqueous extract of Hibiscus rosa-sinensis (flower) (100 mg/kg; 0–7 days/200 mg/kg; 8–14 days/400 mg/kg; 15–21 days) | Supplementation: ↑maternal and fetal weight, ↓atherogenic index and coronary artery risk index, while ↓preimplantation loss rate compared with the untreated diabetic group. |
| Lokman et al. (2019) [189] | Experimental study | Sprague Dawley rats | Streptozotocin (30 mg/kg) | Orthosiphon stamineus (0.1 g/100 g of peso) | Supplementation: significantly ↓serum glucose level. No mortality or signs of toxicity were recorded throughout the study, either for the female rat or her offspring. |
| Laurino et al. (2019) [190] | Experimental study | Wistar rats | Streptozotocin (40 mg/kg) | Lentinus edodes (100 mg/kg) | Supplementation: did not reduce hyperglycemia in rats with induced GDM, but ↑maternal insulin levels and ↓ALT, AST, triglyceride, and total cholesterol levels. |
| Barbalho et al. (2011) [191] | Experimental study | Wistar rats | Streptozotocin (40 mg/kg) | Mentha piperita (Peppermint) (Juice, 100 g/L) | Significantly ↓levels of glucose, cholesterol, LDL-c, and triglycerides and significantly ↑HDL-c levels. |
8. Limitations
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADA | American Diabetes Association |
| AGE | advanced glycation end products |
| ALA | α-linolenic acid |
| ALT | alanine aminotransferase |
| AST | aspartate aminotransferase |
| ATP | adenosine triphosphate |
| BAFF | B cell activating factor |
| BW/GA | relationship between BW and GA |
| BW | birth weight |
| CAT | catalase |
| CD40 | cluster of differentiation 40 |
| CDKAL1 | CDK5 regulatory subunit-associated protein 1-like 1 |
| CDKN2A/2B | cyclin-dependent kinase inhibitor 2A/2B |
| CFU | colony forming unit |
| CGRP | calcitonin gene-related peptide |
| CNCD | chronic non-communicable diseases |
| CRP | C-reactive protein |
| CVD | cardiovascular diseases |
| DASH | dietary approaches to stop hypertension |
| DHA | docosahexaenoic acid |
| DNA | deoxyribonucleic acid |
| EGCG | 3-gallate epigallocatechin |
| EPA | eicosapentaenoic acid |
| ERK | extracellular signal-regulated protein kinase |
| FADH2 | reduced flavin adenine dinucleotide |
| FBG | fasting blood glucose |
| FTO | fat mass- and obesity-associated gene |
| G6PD | glucose-6-phosphate dehydrogenase |
| GA | gestational age |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| GDM | gestational diabetes mellitus |
| GH | growth hormone |
| GLUT | glucose transporters |
| GLUT-1 | glucose transporter 1 |
| GPCRs | G-protein-coupled receptors |
| GPx | glutathione peroxidase |
| GR | glutathione reductase |
| GRP78 | 78-kDa glucose-regulated protein |
| GSH | reduced glutathione |
| GSSG | reduces oxidized glutathione disulfide |
| GST | glutathione S-transferase |
| HAPO | hyperglycemia and adverse pregnancy outcomes |
| HDACs | histone deacetylases |
| HDL-c | high-density lipoprotein cholesterol |
| HHEX | hematopoietically expressed homeobox |
| HOMA-IR | homeostatic assessment model for insulin resistance |
| IADPSG | International Association of Diabetes in Pregnancy Study Group |
| IDF | International Diabetes Federation |
| IFGO | International Federation of Gynecology and Obstetrics |
| IFN-γ | interferon gama |
| IGF-1 | type 1 insulin-like growth factor |
| IGF2BP2 | insulin-like growth factor 2 MRNA binding protein 2 |
| IKK | IκB kinase complex |
| IKKα | IKK alpha subunit |
| IKKβ | IKK beta subunit |
| IKKγ | IKK gamma subunit |
| IL-1β | interleukin 1β |
| IL-6 | Interleukin 6 |
| IRS | insulin receptor substrates |
| IκBs | inhibitory regulators of NF-Kb |
| JAK | Janus kinase pathway |
| LBW | low weight at birth |
| LDL-c | low-density lipoprotein cholesterol |
| LDLR | low-density lipoprotein receptor |
| LGA | large for gestational age |
| LP(a) | lipoprotein a |
| LPS | lipopolysaccharides |
| MCP-1 | monocyte chemoattractant protein-1 |
| MDA | malondialdehyde |
| MnSOD | manganese superoxide dismutase |
| MODY | maturity onset diabetes of the young |
| mTOR | mammalian target of rapamycin |
| MUFA | monounsaturated fatty acids |
| NAD+ | nicotinamide adenine dinucleotide oxidized |
| NADH | nicotinamide adenine dinucleotide |
| NADP+ | nicotinamide adenine dinucleotide phosphate oxidized |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| NB | newborns |
| NDDG | National Diabetes Data Group |
| NF-Kb | nuclear factor kappa B |
| OGTT | Oral Glucose Tolerance Test |
| p38 MAPK | mitogen-activated protein kinase |
| PBMC | peripheral blood mononuclear cells |
| p-eIF2α | phosphorylation of eukaryotic translation initiation factor 2 alpha |
| PFOA | perfluorooctanoic acid |
| PI3K | phosphatidylinositol-3-kinase |
| p-IRE1α | phosphorylation of inositol-requiring enzyme-1alpha |
| PKB | protein kinase B |
| PKC | activation of protein kinase C |
| POS | polycystic ovary syndrome |
| PPARs | peroxisome proliferator-activated receptors |
| PPAR-γ | peroxisome proliferator |
| pPERK | phosphorylation of protein kinase R-like endoplasmic reticulum kinase |
| PTH | parathormone |
| PUFAS | polyunsaturated fatty acids |
| QUICKI | Quantitative Insulin Sensitivity Check Index |
| RAGE | receptor of AGE |
| RNA | ribonucleic acid |
| RNS | reactive nitrogen species |
| RONS | reactive oxygen and nitrogen species |
| ROS | reactive oxygen species |
| SCFA | short-chain fatty acids |
| SCL30A8 | solute carrier family 30 member 8 gene |
| Ser727 | Serine 727 |
| SGA | small for gestational age |
| SOD | superoxide dismutase |
| SOSC3 | suppressor of cytokine signaling 3 |
| STAT | signal transducers and activators of transcription |
| STAT3 | signal transducers and activators of transcription 3 |
| T2DM | type 2 diabetes mellitus |
| TAC | total antioxidant capacity |
| TCA | tricarboxylic acid |
| TCF2 | transcription factor 2 |
| TCF7L2 | transcription factor 7-like 2 |
| TG | triglyceride |
| TLR-4 | toll-like receptors type 4 |
| TLRs | toll-like receptors |
| TMAO | trimethylamine N-oxide |
| TNF-α | tumor necrosis factor α |
| TNF-β | cytokines of the lymphotoxin family |
| Tyr705 | tyrosine 705 |
| Tyr727 | tyrosine 727 |
| VLDL-c | very low-density lipoprotein cholesterol |
| WHO | World Health Organization |
| ZO-1 | zonulin |
| ω-3 | omega-3 |
References
- Lappas, M.; Hiden, U.; Desoye, G.; Froehlich, J.; Hauguel-de Mouzon, S.; Jawerbaum, A. The role of oxidative stress in the pathophysiology of gestational diabetes mellitus. Antioxid. Redox Signal. 2011, 15, 3061–3100. [Google Scholar] [CrossRef]
- Pantham, P.; Aye, I.L.; Powell, T.L. Inflammation in maternal obesity and gestational diabetes mellitus. Placenta 2015, 36, 709–715. [Google Scholar] [CrossRef] [Green Version]
- Plows, J.F.; Stanley, J.L.; Baker, P.N.; Reynolds, C.M.; Vickers, M.H. The Pathophysiology of Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2018, 19, 3342. [Google Scholar] [CrossRef] [Green Version]
- McIntyre, H.D.; Catalano, P.; Zhang, C.; Desoye, G.; Mathiesen, E.R.; Damm, P. Gestational diabetes mellitus. Nat. Rev. Dis. Primers 2019, 5, 47. [Google Scholar] [CrossRef]
- American Diabetes Association. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes—2018. Diabetes Care 2018, 41 (Suppl. 1), S13–S27. [Google Scholar] [CrossRef] [Green Version]
- Szmuilowicz, E.D.; Josefson, J.L.; Metzger, B.E. Gestational Diabetes Mellitus. Endocrinol. Metab. Clin. N. Am. 2019, 48, 479–493. [Google Scholar] [CrossRef]
- Chiefari, E.; Arcidiacono, B.; Foti, D.; Brunetti, A. Gestational diabetes mellitus: An updated overview. J. Endocrinol. Investig. 2017, 40, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Dirar, A.M.; Doupis, J. Gestational diabetes from A to Z. World J. Diabetes 2017, 8, 489–511. [Google Scholar] [CrossRef] [PubMed]
- Li, B.Y.; Xu, X.Y.; Gan, R.Y.; Sun, Q.C.; Meng, J.M.; Shang, A.; Mao, Q.Q.; Li, H.B. Targeting Gut Microbiota for the Prevention and Management of Diabetes Mellitus by Dietary Natural Products. Foods 2019, 8, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Wu, W.; Yang, H.; Zhang, P.; Feng, Y.; Wang, K.; Wang, Y.; Wang, S.; Zhang, Y. A Vegetable Dietary Pattern Is Associated with Lowered Risk of Gestational Diabetes Mellitus in Chinese Women. Diabetes Metab. J. 2020, 44, 887–896. [Google Scholar] [CrossRef]
- Wei, B.; Wang, S.; Wang, Y.; Ke, S.; Jin, W.; Chen, J.; Zhang, H.; Sun, J.; Henning, S.M.; Wang, J.; et al. Gut microbiota-mediated xanthine metabolism is associated with resistance to high-fat diet-induced obesity. J. Nutr. Biochem. 2021, 88, 108533. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M.; Arqués, J.; Medina, M.; Gaya, P.; de Las Rivas, B.; Muñoz, R. Bioactivation of Phytoestrogens: Intestinal Bacteria and Health. Crit. Rev. Food Sci. Nutr. 2016, 56, 1826–1843. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Yang, H.; Geng, Q.; Ma, Q.; Long, Y.; Zhou, C.; Chen, M. Association of oxidative stress biomarkers with gestational diabetes mellitus in pregnant women: A case-control study. PLoS ONE 2015, 10, e0126490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.C.; Lai, Y.N.; Wang, W.H.; Xu, X.M.; Li, X.Q.; Wang, H.; Zheng, J.Y.; Zheng, J.Q. Metformin Ameliorates Gestational Diabetes Mellitus-Induced Endothelial Dysfunction via Downregulation of p65 and Upregulation of Nrf2. Front. Pharmacol. 2020, 11, 575390. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, J.; Shi, W.; Du, N.; Xu, X.; Zhang, Y.; Ji, P.; Zhang, F.; Jia, Z.; Wang, Y.; et al. Dysbiosis of maternal and neonatal microbiota associated with gestational diabetes mellitus. Gut 2018, 67, 1614–1625. [Google Scholar] [CrossRef]
- Brawerman, G.M.; Dolinsky, V.W. Therapies for gestational diabetes and their implications for maternal and offspring health: Evidence from human and animal studies. Pharmacol. Res. 2018, 130, 52–73. [Google Scholar] [CrossRef]
- Ponzo, V.; Fedele, D.; Goitre, I.; Leone, F.; Lezo, A.; Monzeglio, C.; Finocchiaro, C.; Ghigo, E.; Bo, S. Diet-Gut Microbiota Interactions and Gestational Diabetes Mellitus (GDM). Nutrients 2019, 11, 330. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes—2021. Diabetes Care 2021, 44 (Suppl. 1), S15–S33. [Google Scholar] [CrossRef]
- International Diabetes Federation. IDF Diabetes Atlas, 8th ed.; International Diabetes Federation: Brussels, Belgium, 2017; ISBN 978-2-930229-87-4. [Google Scholar]
- Organização Pan-Americana de Saúde; Ministério da Saúde; Federação Brasileira das Associações de Ginecologia e Obstetrícia; Sociedade Brasileira de Diabetes. Rastreamento e Diagnóstico de Diabetes Mellitus Gestacional no Brasil—Brasília. 2017. E-book, pp. 15–19. Available online: https://www.febrasgo.org.br/images/pec/CNE_pdfs/Rastreamento-Diabetes.pdf (accessed on 15 October 2021).
- O’Sullivan, J.B.; Mahan, C.M. Criteria for the oral glucose tolerance test in pregnancy. Diabetes 1964, 13, 278–285. [Google Scholar]
- National Diabetes Data Group. Classification and diagnosis of diabetes mellitus and other categories of glucose intolerance. National Diabetes Data Group. Diabetes 1979, 28, 1039–1057. [Google Scholar] [CrossRef]
- Carpenter, M.W.; Coustan, D.R. Criteria for screening tests for gestational diabetes. Am. J. Obstet. Gynecol. 1982, 144, 768–773. [Google Scholar] [CrossRef]
- American College of Obstetrics and Gynecology. Proceedings of the Third International Workshop-Conference on Gestational Diabetes Mellitus. November 8–10,1990, Chicago, Illinois. Diabetes 1991, 40 (Suppl. 2), 1–201. [Google Scholar]
- HAPO Study Cooperative Research Group. Hyperglycemia and adverse pregnancy outcomes. N. Engl. J. Med. 2008, 358, 1991–2002. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Definition, Diagnosis and Classification of Diabetes Mellitus and its Complications; World Health Organization: Geneva, Switzerland, 1999. [Google Scholar]
- International Association of Diabetes and Pregnancy Study Groups Consensus Panel. International association of diabetes and pregnancy study groups recommendations on the diagnosis and classification of hyperglycemia in pregnancy. Diabetes Care 2010, 33, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Organização Pan-Americana da Saúde. Rastreamento e Diagnóstico de Diabetes Mellitus Gestacional No Brasil. Ministério da Saúde. Federação Brasileira das Associações de Ginecologia e Obstetrícia. (DF): OPAS; Sociedade Brasileira de Diabetes: São Paulo, Brazil, 2016. [Google Scholar]
- HAPO Study Cooperative Research Group. The Hyperglycemia and Adverse Pregnancy Outcome (HAPO) Study. Int. J. Gynaecol. Obstet. 2002, 78, 69–77. [Google Scholar] [CrossRef]
- Blumer, I.; Hadar, E.; Hadden, D.R.; Jovanovic, L.; Mestman, J.H.; Murad, M.H.; Yogev, Y. Diabetes and pregnancy: An endocrine society clinical practice guideline. J. Clin. Endocrinol. Metab. 2013, 98, 4227–4249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Diagnostic Criteria and Classification of Hyperglycaemia First Detected in Pregnancy: A World Health Organization Guideline; WHO Press: Geneva, Switzerland, 2013; Available online: http://apps.who.int/iris/bitstream/10665/85975/1/WHO_NMH_MND_13.2_eng.pdf (accessed on 15 October 2021).
- Hod, M.; Kapur, A.; Sacks, D.A.; Hadar, E.; Agarwal, M.; Di Renzo, G.C.; Roura, L.C.; McIntyre, H.D.; Morris, J.L.; Divakar, H. The International Federation of Gynecology and Obstetrics (FIGO) initiative on gestational diabetes mellitus: A pragmatic guide for diagnosis, management, and care. Int. J. Gynaecol. Obstet. 2015, 131 (Suppl. 3), S173–S211. [Google Scholar] [CrossRef] [Green Version]
- McIntyre, H.D.; Sacks, D.A.; Barbour, L.A.; Feig, D.S.; Catalano, P.M.; Damm, P.; McElduff, A. Issues with the diagnosis and classification of hyperglycemia in early pregnancy. Diabetes Care 2016, 39, 53–54. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.I.; Duncan, B.B.; Reichelt, A.J.; Branchtein, L.; Matos, M.C.; e Forti, A.C.; Spichler, E.R.; Pousada, J.M.; Teixeira, M.M.; Yamashita, T.; et al. Gestational diabetes mellitus diagnosed with a 2-h 75-g oral glucose tolerance test and adverse pregnancy outcomes. Diabetes Care 2001, 24, 1151–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trujillo, J.; Vigo, A.; Reichelt, A.; Duncan, B.B.; Schmidt, M.I. Fasting plasma glucose to avoid a full OGTT in the diagnosis of gestational diabetes. Diabetes Res. Clin. Pract. 2016, 105, 322–326. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Wang, S.; Ji, J.; Ge, A.; Chen, C.; Zhu, Y.; Xie, N.; Wang, Y. Risk factors and management of gestational diabetes. Cell Biochem. Biophys. 2016, 71, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Rawal, S.; Chong, Y.S. Risk factors for gestational diabetes: Is prevention possible? Diabetologia 2016, 59, 1385–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carolan, M. Maternal age ≥45 years and maternal and perinatal outcomes: A review of the evidence. Midwifery 2013, 29, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Lean, S.C.; Derricott, H.; Jones, R.L.; Heazell, A.E.P. Advanced maternal age and adverse pregnancy outcomes: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0186287. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.M.; Kim, T.H.; Lim, S.; Choi, S.H.; Shin, H.D.; Lee, H.K.; Park, K.S.; Jang, H.C. Type 2 diabetes-associated genetic variants discovered in the recent genome-wide association studies are related to gestational diabetes mellitus in the Korean population. Diabetologia 2009, 52, 253–261. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Xie, N.; Zhang, L.; Chen, D. Impact of family history of diabetes on blood glucose, lipid levels and perinatal outcomes in pregnant women with gestational diabetes mellitus. J. Zhejiang Univ. 2021, 50, 329–334. [Google Scholar] [CrossRef]
- Lauenborg, J.; Grarup, N.; Damm, P.; Borch-Johnsen, K.; Jørgensen, T.; Pedersen, O.; Hansen, T. Common type 2 diabetes risk gene variants associate with gestational diabetes. J. Clin. Endocrinol. Metab. 2009, 94, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Urbanová, J.; Brunerová, L.; Nunes, M.A.; Brož, J. MODY diabetes and screening of gestational diabetes. Ceska Gynekol. 2020, 85, 124–130. [Google Scholar]
- Pu, J.; Zhao, B.; Wang, E.J.; Nimbal, V.; Osmundson, S.; Kunz, L.; Popat, R.A.; Chung, S.; Palaniappan, L.P. Racial/Ethnic Differences in Gestational Diabetes Prevalence and Contribution of Common Risk Factors. Paediatr. Perinat. Epidemiol. 2015, 29, 436–443. [Google Scholar] [CrossRef]
- Jaffe, A.; Giveon, S.; Rubin, C.; Novikov, I.; Ziv, A.; Kalter-Leibovici, O. Gestational diabetes risk in a multi-ethnic population. Acta Diabetol. 2020, 57, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, N.; Nachum, Z.; Green, M.S. The prevalence of gestational diabetes mellitus recurrence—Effect of ethnicity and parity: A metaanalysis. Am. J. Obstet. Gynecol. 2015, 213, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Shang, M.; Dong, X.; Hou, L. Correlation of adipokines and markers of oxidative stress in women with gestational diabetes mellitus and their newborns. J. Obstet. Gynaecol. Res. 2018, 44, 637–646. [Google Scholar] [CrossRef]
- LifeCycle Project-Maternal Obesity and Childhood Outcomes Study Group. Association of Gestational Weight Gain with Adverse Maternal and Infant Outcomes. JAMA 2019, 321, 1702–1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barakat, R.; Refoyo, I.; Coteron, J.; Franco, E. Exercise during pregnancy has a preventative effect on excessive maternal weight gain and gestational diabetes. A randomized controlled trial. Braz. J. Phys. Ther. 2019, 23, 148–155. [Google Scholar] [CrossRef]
- Zhang, C.; Schulze, M.B.; Solomon, C.G.; Hu, F.B. A prospective study of dietary pattRNS, meat intake and the risk of gestational diabetes mellitus. Diabetologia 2006, 49, 2604–2613. [Google Scholar] [CrossRef] [Green Version]
- Badon, S.E.; Wartko, P.D.; Qiu, C.; Sorensen, T.K.; Williams, M.A.; Enquobahrie, D.A. Leisure Time Physical Activity and Gestational Diabetes Mellitus in the Omega Study. Med. Sci. Sports Exerc. 2016, 48, 1044–1052. [Google Scholar] [CrossRef] [Green Version]
- Mijatovic-Vukas, J.; Capling, L.; Cheng, S.; Stamatakis, E.; Louie, J.; Cheung, N.W.; Markovic, T.; Ross, G.; Senior, A.; Brand-Miller, J.C.; et al. Associations of Diet and Physical Activity with Risk for Gestational Diabetes Mellitus: A Systematic Review and Meta-Analysis. Nutrients 2018, 10, 698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, C.; Thibodeaux, J.R.; Hanson, R.G.; Narotsky, M.G.; Rogers, J.M.; Lindstrom, A.B.; Strynar, M.J. Effects of perfluorooctanoic acid exposure during pregnancy in the mouse. Toxicol. Sci. 2006, 90, 510–518. [Google Scholar] [CrossRef] [Green Version]
- Fenton, S.E.; Reiner, J.L.; Nakayama, S.F.; Delinsky, A.D.; Stanko, J.P.; Hines, E.P.; White, S.S.; Lindstrom, A.B.; Strynar, M.J.; Petropoulou, S.E. Analysis of PFOA in dosed CD-1 mice. Part 2. Disposition of PFOA in tissues and fluids from pregnant and lactating mice and their pups. Reprod. Toxicol. 2009, 27, 365–372. [Google Scholar] [CrossRef] [Green Version]
- Steenland, K.; Tinker, S.; Shankar, A.; Ducatman, A. Association of perfluorooctanoic acid (PFOA) and perfluorooctane sulfonate (PFOS) with uric acid among adults with elevated community exposure to PFOA. Environ. Health Perspect. 2010, 118, 229–233. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Sundaram, R.; Maisog, J.; Calafat, A.M.; Barr, D.B.; Buck Louis, G.M. A prospective study of prepregnancy serum concentrations of perfluorochemicals and the risk of gestational diabetes. Fertil. Steril. 2015, 103, 184–189. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, K.T.; Raaschou-Nielsen, O.; McLaughlin, J.K.; Lipworth, L.; Tjønneland, A.; Overvad, K.; Sørensen, M. Association between plasma PFOA and PFOS levels and total cholesterol in a middle-aged Danish population. PLoS ONE 2013, 8, e56969. [Google Scholar] [CrossRef]
- Wang, H.; Yang, J.; Du, H.; Xu, L.; Liu, S.; Yi, J.; Qian, X.; Chen, Y.; Jiang, Q.; He, G. Perfluoroalkyl substances, glucose homeostasis, and gestational diabetes mellitus in Chinese pregnant women: A repeat measurement-based prospective study. Environ. Int. 2018, 114, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Han, K.; You, S.Y.; Kwon, H.S.; Yoon, K.H.; Lee, S.H. Prepregnancy smoking and the risk of gestational diabetes requiring insulin therapy. Sci. Rep. 2020, 10, 13901. [Google Scholar] [CrossRef]
- Bar-Zeev, Y.; Haile, Z.T.; Chertok, I.A. Association between Prenatal Smoking and Gestational Diabetes Mellitus. Obstet. Gynecol. 2020, 135, 91–99. [Google Scholar] [CrossRef]
- Hinkle, S.N.; Bao, W.; Wu, J.; Sun, Y.; Ley, S.H.; Tobias, D.K.; Qian, F.; Rawal, S.; Zhu, Y.; Chavarro, J.E.; et al. Association of Habitual Alcohol Consumption with Long-term Risk of Type 2 Diabetes Among Women With a History of Gestational Diabetes. JAMA Netw. Open 2021, 4, e2124669. [Google Scholar] [CrossRef] [PubMed]
- Elting, M.W.; Korsen, T.J.M.; Bezemer, P.D.; Schoemaker, J. Prevalence of diabetes mellitus, hypertension and cardiac complaints in a follow-up study of a Dutch PCOS population. Hum. Reprod. 2001, 16, 556–560. [Google Scholar] [CrossRef]
- Rojas, J.; Chávez-Castillo, M.; Bermúdez, V. O papel da metformina nos distúrbios metabólicos durante a gravidez: Síndrome dos Ovários Policísticos e Diabetes Mellitus Gestacional. Int. J. Reprod. Med. 2014, 2014, 797681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, G.; Garzon, S.; Fichera, M.; Panella, M.M.; Catena, U.; Schiattarella, A.; de Franciscis, P.; Vilos, G.; Tesarik, J.; Török, P.; et al. Vitamin D and Gestational Diabetes Mellitus: Is There a Link? Antioxidants 2019, 8, 511. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, C.; Song, Y.; Zhang, Z. Deficiência de vitamina D no soro e risco de diabetes mellitus gestacional: Uma meta-análise. Arch. Med. Sci. 2020, 16, 742–751. [Google Scholar] [CrossRef]
- Barker, D.J.; Bull, A.R.; Osmond, C.; Simmonds, S.J. Fetal and placental size and risk of hypertension in adult life. BMJ 1990, 301, 259–262. [Google Scholar] [CrossRef]
- Yan, J.; Yang, H. Gestational diabetes mellitus, programing and epigenetics. J. Matern. Fetal Neonatal Med. 2014, 27, 1266–1269. [Google Scholar] [CrossRef]
- Franzago, M.; Fraticelli, F.; Stuppia, L.; Vitacolonna, E. Nutrigenetics, epigenetics and gestational diabetes: Consequences in mother and child. Epigenetics 2019, 14, 215–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Q.; Cai, H.; Xiang, Y.B.; Zhou, P.; Li, H.; Yang, G.; Jiang, Y.; Shu, X.O.; Zheng, W.; Xu, W.H. Prospective cohort studies of birth weight and risk of obesity, diabetes, and hypertension in adulthood among the Chinese population. J. Diabetes 2019, 11, 55–64. [Google Scholar] [CrossRef]
- Mendonça, E.L.S.S.; de Lima Macêna, M.; Bueno, N.B.; de Oliveira, A.C.M.; Mello, C.S. Premature birth, low birth weight, small for gestational age and chronic non-communicable diseases in adult life: A systematic review with meta-analysis. Early Hum. Dev. 2020, 149, 105154. [Google Scholar] [CrossRef] [PubMed]
- Kamana, K.C.; Shakya, S.; Zhang, H. Gestational diabetes mellitus and macrosomia: A literature review. Ann. Nutr. Metab. 2015, 66 (Suppl. 2), 14–20. [Google Scholar] [CrossRef]
- Gascho, C.L.; Leandro, D.M.; e Silva, T.R.; Silva, J.C. Predictors of cesarean delivery in pregnant women with gestational diabetes mellitus. Rev. Bras. Ginecol. Obstet. 2017, 39, 60–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, J. Weight and length at birth of infants of diabetic mothers. Acta Endocrinol. 1954, 16, 330–342. [Google Scholar] [CrossRef]
- Freinkel, N. Banting Lecture 1980. Of pregnancy and progeny. Diabetes 1980, 29, 1023–1035. [Google Scholar] [CrossRef]
- Moore, T.R. A comparison of amniotic fluid fetal pulmonary phospholipids in normal and diabetic pregnancy. Am. J. Obstet. Gynecol. 2002, 186, 641–650. [Google Scholar] [CrossRef]
- McFarland, L.V.; Raskin, M.; Daling, J.R.; Benedetti, T.J. Erb/Duchenne’s palsy: A consequence of fetal macrosomia and method of delivery. Obstet. Gynecol. 1986, 68, 784–788. [Google Scholar]
- Vaquero, G.; Ramos, A.; Martinez, J.C.; Valero, P.; Nunez-Enamorado, N.; Simon-De Las Heras, R.; Camacho-Salas, A. Paralisis braquial obstetrica: Incidencia, seguimiento evolutivo y factores pronosticos [Obstetric brachial plexus palsy: Incidence, monitoring of progress and prognostic factors]. Rev. Neurol. 2017, 65, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Araújo Júnior, E.; Peixoto, A.B.; Zamarian, A.C.; Elito Júnior, J.; Tonni, G. Macrosomia. Best Pract. Res. Clin. Obstet. Gynaecol. 2017, 38, 83–96. [Google Scholar] [CrossRef]
- Silva, L.; Plösch, T.; Toledo, F.; Faas, M.M.; Sobrevia, L. Adenosine kinase and cardiovascular fetal programming in gestational diabetes mellitus. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165397. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, D.L. Morbidity and mortality in Brazil. Cad. Saúde Colet. 2015, 23, 1. [Google Scholar] [CrossRef] [Green Version]
- Lowe, W.L.; Scholtens, D.M.; Lowe, L.P.; Kuang, A.; Nodzenski, M.; Talbot, O.; Catalano, P.M.; Linder, B.; Brickman, W.J.; Clayton, P.; et al. Association of Gestational Diabetes with Maternal Disorders of Glucose Metabolism and Childhood Adiposity. JAMA 2018, 320, 1005–1016. [Google Scholar] [CrossRef]
- Prates, T. Nutrição no Início da Vida, Epigenética e Prevenção das Doenças Crônicas não Transmissíveis: Uma Janela de Oportunidades para Pediatras/São Paulo: ILSI Brasil—International Life Sciences Institute do Brasil: São Paulo, Brasil, 2018. Available online: https://ilsibrasil.org/wp-content/uploads/sites/9/2019/02/Fasc%C3%ADculo-EPIGEN%C3%89TICA.pdf (accessed on 15 October 2021).
- Liu, T.; Zheng, W.; Wang, L.; Wang, L.; Zhang, Y. TLR4/NF-κB Signaling Pathway Participates in the Protective Effects of Apocynin on Gestational Diabetes Mellitus Induced Placental Oxidative Stress and Inflammation. Reprod. Sci. 2020, 27, 722–730. [Google Scholar] [CrossRef]
- Pasternak, Y.; Ohana, M.; Biron-Shental, T.; Cohen-Hagai, K.; Benchetrit, S.; Zitman-Gal, T. Thioredoxin, thioredoxin interacting protein and transducer and activator of transcription 3 in gestational diabetes. Mol. Biol. Rep. 2020, 47, 1199–1206. [Google Scholar] [CrossRef]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB system. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzmicki, M.; Telejko, B.; Wawrusiewicz-Kurylonek, N.; Lipinska, D.; Pliszka, J.; Wilk, J.; Zielinska, A.; Skibicka, J.; Szamatowicz, J.; Kretowski, A.; et al. The expression of genes involved in NF-κB activation in peripheral blood mononuclear cells of patients with gestational diabetes. Eur. J. Endocrinol. 2013, 168, 419–427. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Ghanim, H.; Aljada, A.; Daoud, N.; Deopurkar, R.; Chaudhuri, A.; Dandona, P. Role of inflammatory mediators in the suppression of insulin receptor phosphorylation in circulating mononuclear cells of obese subjects. Diabetologia 2007, 50, 278–285. [Google Scholar] [CrossRef] [Green Version]
- Mello, V.D.; Kolehmainen, M.; Pulkkinen, L.; Schwab, U.; Mager, U.; Laaksonen, D.E.; Niskanen, L.; Gylling, H.; Atalay, M.; Rauramaa, R.; et al. Downregulation of genes involved in NFkappaB activation in peripheral blood mononuclear cells after weight loss is associated with the improvement of insulin sensitivity in individuals with the metabolic syndrome: The GENOBIN study. Diabetologia 2008, 51, 2060–2067. [Google Scholar] [CrossRef] [PubMed]
- Abell, S.K.; De Courten, B.; Boyle, J.A.; Teede, H.J. Inflammatory and Other Biomarkers: Role in Pathophysiology and Prediction of Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2015, 16, 13442–13473. [Google Scholar] [CrossRef]
- Nguyen-Ngo, C.; Jayabalan, N.; Salomon, C.; Lappas, M. Molecular pathways disrupted by gestational diabetes mellitus. J. Mol. Endocrinol. 2019, 63, R51–R72. [Google Scholar] [CrossRef] [Green Version]
- Bossick, A.S.; Peters, R.M.; Burmeister, C.; Kakumanu, N.; Shill, J.E.; Cassidy-Bushrow, A.E. Antenatal inflammation and gestational diabetes mellitus risk among pregnant African-American women. J. Reprod. Immunol. 2016, 115, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Loh, C.Y.; Arya, A.; Naema, A.F.; Wong, W.F.; Sethi, G.; Looi, C.Y. Signal Transducer and Activator of Transcription (STATs) Proteins in Cancer and Inflammation: Functions and Therapeutic Implication. Front. Oncol. 2019, 9, 48. [Google Scholar] [CrossRef] [Green Version]
- Gianotti, T.F.; Sookoian, S.; Gemma, C.; Burgueño, A.L.; González, C.D.; Pirola, C.J. Study of genetic variation in the STAT3 on obesity and insulin resistance in male adults. Obesity 2008, 16, 1702–1707. [Google Scholar] [CrossRef] [PubMed]
- Recio, C.; Guerra, B.; Guerra-Rodríguez, M.; Aranda-Tavío, H.; Martín-Rodríguez, P.; de Mirecki-Garrido, M.; Brito-Casillas, Y.; García-Castellano, J.M.; Estévez-Braun, A.; Fernández-Pérez, L. Signal transducer and activator of transcription (STAT)-5: An opportunity for drug development in oncohematology. Oncogene 2019, 38, 4657–4668. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, J.E.; Liu, H.Y.; Cao, W.; Chen, J. Regulation of interleukin-6-induced hepatic insulin resistance by mammalian target of rapamycin through the STAT3-SOCS3 pathway. J. Biol. Chem. 2008, 283, 708–715. [Google Scholar] [CrossRef] [Green Version]
- Heo, Y.J.; Choi, S.E.; Jeon, J.Y.; Han, S.J.; Kim, D.J.; Kang, Y.; Lee, K.W.; Kim, H.J. Visfatin Induces Inflammation and Insulin Resistance via the NF-κB and STAT3 Signaling Pathways in Hepatocytes. J. Diabetes Res. 2019, 2019, 4021623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashili, F.; Chibalin, A.V.; Krook, A.; Zierath, J.R. Constitutive STAT3 phosphorylation contributes to skeletal muscle insulin resistance in type 2 diabetes. Diabetes 2013, 62, 457–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, A.; Van Dyke, T.E. Role of suppressors of cytokine signaling 3 in bone inflammatory responses. Front. Immunol. 2014, 4, 506. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.H.; Choi, S.E.; Ha, E.S.; Jung, J.G.; Han, S.J.; Kim, H.J.; Kim, D.J.; Kang, Y.; Lee, K.W. IL-6 induction of TLR-4 gene expression via STAT3 has an effect on insulin resistance in human skeletal muscle. Acta Diabetol. 2013, 50, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Xu, J.; Wei, G.; Zhang, L.; Sun, L.; Wang, G.; Li, F.; Jiang, F. HNF1α Controls Liver Lipid Metabolism and Insulin Resistance via Negatively Regulating the SOCS-3-STAT3 Signaling Pathway. J. Diabetes Res. 2019, 2019, 5483946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Chen, Z.; Wang, Y.; Tweardy, D.J.; Mitch, W.E. Stat3 activation induces insulin resistance via a muscle-specific E3 ubiquitin ligase Fbxo40. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E625–E635. [Google Scholar] [CrossRef]
- Nourbakhsh, M.; Nourbakhsh, M.; Gholinejad, Z.; Razzaghy-Azar, M. Visfatin in obese children and adolescents and its association with insulin resistance and metabolic syndrome. Scand. J. Clin. Lab. Investig. 2015, 75, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Radzicka, S.; Pietryga, M.; Iciek, R.; Brązert, J. The role of visfatin in pathogenesis of gestational diabetes (GDM). Ginekol. Pol. 2018, 89, 518–521. [Google Scholar] [CrossRef] [Green Version]
- Decker, T.; Kovarik, P. Serine phosphorylation of STATs. Oncogene 2000, 19, 2628–2637. [Google Scholar] [CrossRef] [Green Version]
- Reyna, S.M.; Ghosh, S.; Tantiwong, P.; Meka, C.S.; Eagan, P.; Jenkinson, C.P.; Cersosimo, E.; Defronzo, R.A.; Coletta, D.K.; Sriwijitkamol, A.; et al. Elevated toll-like receptor 4 expression and signaling in muscle from insulin-resistant subjects. Diabetes 2008, 57, 2595–2602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Al-Shehri, S.S. Reactive oxygen and nitrogen species and innate immune response. Biochimie 2021, 181, 52–64. [Google Scholar] [CrossRef]
- Gauster, M.; Majali-Martinez, A.; Maninger, S.; Gutschi, E.; Greimel, P.H.; Ivanisevic, M.; Djelmis, J.; Desoye, G.; Hiden, U. Maternal Type 1 diabetes activates stress response in early placenta. Placenta 2017, 50, 110–116. [Google Scholar] [CrossRef]
- Tenório, M.B.; Ferreira, R.C.; Moura, F.A.; Bueno, N.B.; de Oliveira, A.C.M.; Goulart, M.O.F. Cross-Talk between Oxidative Stress and Inflammation in Preeclampsia. Oxid. Med. Cell. Longev. 2019, 2019, 8238727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taysi, S.; Tascan, A.S.; Ugur, M.G.; Demir, M. Radicals, Oxidative/Nitrosative Stress and Preeclampsia. Mini Rev. Med. Chem. 2019, 19, 178–193. [Google Scholar] [CrossRef]
- Marrocco, I.; Altieri, F.; Peluso, I. Measurement and Clinical Significance of Biomarkers of Oxidative Stress in Humans. Oxid. Med. Cell. Longev. 2017, 2017, 6501046. [Google Scholar] [CrossRef]
- Hastie, R.; Lappas, M. The effect of pre-existing maternal obesity and diabetes on placental mitochondrial content and electron transport chain activity. Placenta 2014, 35, 673–683. [Google Scholar] [CrossRef]
- Vidal, Z.E.; Rufino, S.C.; Tlaxcalteco, E.H.; Trejo, C.H.; Campos, R.M.; Meza, M.N.; Rodríguez, R.C.; Arroyo-Helguera, O. Oxidative stress increased in pregnant women with iodine deficiency. Biol. Trace Elem. Res. 2014, 157, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Sun, F.; Que, S.; Li, Y.; Yang, S.; Liu, G. Prepregnancy maternal diabetes combined with obesity impairs placental mitochondrial function involving Nrf2/ARE pathway and detrimentally alters metabolism of offspring. Obes. Res. Clin. Pract. 2018, 12 (Suppl. 2), 90–100. [Google Scholar] [CrossRef]
- Ramiro-Cortijo, D.; Herrera, T.; Rodríguez-Rodríguez, P.; López De Pablo, Á.L.; De La Calle, M.; López-Giménez, M.R.; Mora-Urda, A.I.; Gutiérrez-Arzapalo, P.Y.; Gómez-Rioja, R.; Aguilera, Y.; et al. Maternal plasma antioxidant status in the first trimester of pregnancy and development of obstetric complications. Placenta 2016, 47, 37–45. [Google Scholar] [CrossRef]
- Agarwal, A.; Gupta, S.; Sharma, R.K. Role of oxidative stress in female reproduction. Reprod. Biol. Endocrinol. 2005, 3, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamali, M.; Dadkhah, F.; Sadrkhanlou, M.; Jamilian, M.; Ahmadi, S.; Tajabadi-Ebrahimi, M.; Jafari, P.; Asemi, Z. Effects of probiotic supplementation on glycaemic control and lipid profiles in gestational diabetes: A randomized, double-blind, placebo-controlled trial. Diabetes Metab. 2016, 42, 234–241. [Google Scholar] [CrossRef]
- Papachristoforou, E.; Lambadiari, V.; Maratou, E.; Makrilakis, K. Association of Glycemic Indices (Hyperglycemia, Glucose Variability, and Hypoglycemia) with Oxidative Stress and Diabetic Complications. J. Diabetes Res. 2020, 2020, 7489795. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M. Molecular pathways associated with oxidative stress in diabetes mellitus. Biomed. Pharmacother. 2018, 108, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Frijhoff, J.; Winyard, P.G.; Zarkovic, N.; Davies, S.S.; Stocker, R.; Cheng, D.; Knight, A.R.; Taylor, E.L.; Oettrich, J.; Ruskovska, T.; et al. Clinical Relevance of Biomarkers of Oxidative Stress. Antioxid. Redox Signal. 2015, 23, 1144–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belfiore, A.; Malaguarnera, R.; Vella, V.; Lawrence, M.C.; Sciacca, L.; Frasca, F.; Morrione, A.; Vigneri, R. Insulin Receptor Isoforms in Physiology and Disease: An Updated View. Endocr. Rev. 2017, 38, 379–431. [Google Scholar] [CrossRef] [PubMed]
- Volpe, C.M.O.; Villar-Delfino, P.H.; Dos Anjos, P.M.F.; Nogueira-Machado, J.A. Cellular death, reactive oxygen species (ROS) and diabetic complications. Cell Death Dis. 2018, 9, 119. [Google Scholar] [CrossRef]
- Sánchez-Santos, A.; Martínez-Hernández, M.G.; Contreras-Ramos, A.; Ortega-Camarillo, C.; Baiza-Gutman, L.A. Hyperglycemia-induced mouse trophoblast spreading is mediated by reactive oxygen species. Mol. Reprod. Dev. 2018, 85, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Chiarello, D.I.; Abad, C.; Rojas, D.; Toledo, F.; Vázquez, C.M.; Mate, A.; Sobrevia, L.; Marín, R. Oxidative stress: Normal pregnancy versus preeclampsia. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165354. [Google Scholar] [CrossRef]
- Little, B.D.; Hopkins, R.Z. Superoxide Dismutases in Biology and Medicine: Essentials and Recent Advances. React. Oxyg. Species 2020, 9, 13–21. [Google Scholar] [CrossRef]
- Giustarini, D.; Tsikas, D.; Colombo, G.; Milzani, A.; Dalle-Donne, I.; Fanti, P.; Rossi, R. Pitfalls in the analysis of the physiological antioxidant glutathione (GSH) and its disulfide (GSSG) in biological samples: An elephant in the room. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2016, 1019, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Bartolini, D.; Giustarini, D.; Pietrella, D.; Rossi, R.; Galli, F. Glutathione S-transferase P influences the Nrf2-dependent response of cellular thiols to seleno-compounds. Cell Biol. Toxicol. 2020, 36, 379–386. [Google Scholar] [CrossRef]
- Romo, A.I.B.; Dibo, V.S.; Abreu, D.S.; Carepo, M.S.P.; Neira, A.C.; Castillo, I.; Lemus, L.; Nascimento, O.R.; Bernhardt, P.V.; Sousa, E.H.S.; et al. Ascorbyl and hydroxyl radical generation mediated by a copper complex adsorbed on gold. Dalton Trans. 2019, 48, 14128–14137. [Google Scholar] [CrossRef]
- Di Vincenzo, A.; Tana, C.; El Hadi, H.; Pagano, C.; Vettor, R.; Rossato, M. Antioxidant, Anti-Inflammatory, and Metabolic Properties of Tocopherols and Tocotrienols: Clinical Implications for Vitamin E Supplementation in Diabetic Kidney Disease. Int. J. Mol. Sci. 2019, 20, 5101. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Gutteridge, J.M.C. Free Radical in Biology and Medicine, 5th ed.; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Clarke, G.; Stilling, R.M.; Kennedy, P.J.; Stanton, C.; Cryan, J.F.; Dinan, T.G. Minireview: Gut microbiota: The neglected endocrine organ. Mol. Endocrinol. 2014, 28, 1221–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijnikman, A.S.; Gerdes, V.E.; Nieuwdorp, M.; Herrema, H. Evaluating Causality of Gut Microbiota in Obesity and Diabetes in Humans. Endocr. Rev. 2018, 39, 133–153. [Google Scholar] [CrossRef] [PubMed]
- Sircana, A.; Framarin, L.; Leone, N.; Berrutti, M.; Castellino, F.; Parente, R.; De Michieli, F.; Paschetta, E.; Musso, G. Altered Gut Microbiota in Type 2 Diabetes: Just a Coincidence? Curr. Diab. Rep. 2018, 18, 98. [Google Scholar] [CrossRef]
- Hu, C.; Wong, F.S.; Wen, L. Type 1 diabetes and gut microbiota: Friend or foe? Pharmacol. Res. 2015, 98, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crusell, M.K.W.; Hansen, T.H.; Nielsen, T.; Allin, K.H.; Rühlemann, M.C.; Damm, P.; Vestergaard, H.; Rørbye, C.; Jørgensen, N.R.; Christiansen, O.B.; et al. Gestational diabetes is associated with change in the gut microbiota composition in third trimester of pregnancy and postpartum. Microbiome 2018, 6, 89. [Google Scholar] [CrossRef] [PubMed]
- Serino, M.; Fernández-Real, J.M.; García-Fuentes, E.; Queipo-Ortuño, M.; Moreno-Navarrete, J.M.; Sánchez, A.; Burcelin, R.; Tinahones, F. The gut microbiota profile is associated with insulin action in humans. Acta Diabetol. 2013, 50, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Crommen, S.; Simon, M.C. Microbial Regulation of Glucose Metabolism and Insulin Resistance. Genes 2017, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, W.; Bowers, K.; Tobias, D.K.; Olsen, S.F.; Chavarro, J.; Vaag, A.; Kiely, M.; Zhang, C. Prepregnancy low-carbohydrate dietary pattern and risk of gestational diabetes mellitus: A prospective cohort study. Am. J. Clin. Nutr. 2014, 99, 1378–1384. [Google Scholar] [CrossRef] [Green Version]
- Mokkala, K.; Tertti, K.; Rönnemaa, T.; Vahlberg, T.; Laitinen, K. Evaluation of serum zonulin for use as an early predictor for gestational diabetes. Nutr. Diabetes 2017, 7, e253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayashree, B.; Bibin, Y.S.; Prabhu, D.; Shanthirani, C.S.; Gokulakrishnan, K.; Lakshmi, B.S.; Mohan, V.; Balasubramanyam, M. Increased circulatory levels of lipopolysaccharide (LPS) and zonulin signify novel biomarkers of proinflammation in patients with type 2 diabetes. Mol. Cell. Biochem. 2014, 388, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Cortez, R.V.; Taddei, C.R.; Sparvoli, L.G.; Ângelo, A.G.S.; Padilha, M.; Mattar, R.; Daher, S. Microbiome and its relation to gestational diabetes. Endocrine 2019, 64, 254–264. [Google Scholar] [CrossRef]
- Hasain, Z.; Mokhtar, N.M.; Kamaruddin, N.A.; Mohamed Ismail, N.A.; Razalli, N.H.; Gnanou, J.V.; Raja Ali, R.A. Gut Microbiota and Gestational Diabetes Mellitus: A Review of Host-Gut Microbiota Interactions and Their Therapeutic Potential. Front. Cell. Infect. Microbiol. 2020, 10, 188. [Google Scholar] [CrossRef]
- Ferrocino, I.; Ponzo, V.; Gambino, R.; Zarovska, A.; Leone, F.; Monzeglio, C.; Goitre, I.; Rosato, R.; Romano, A.; Grassi, G.; et al. Changes in the gut microbiota composition during pregnancy in patients with gestational diabetes mellitus (GDM). Sci. Rep. 2018, 8, 12216. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.C.; Bueno, A.A.; de Souza, R.G.; Mota, J.F. Gut microbiota, probiotics and diabetes. Nutr. J. 2014, 13, 60. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Cao, H.; Cover, T.L.; Whitehead, R.; Washington, M.K.; Polk, D.B. Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth. Gastroenterology 2007, 132, 562–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banan, A.; Keshavarzian, A.; Zhang, L.; Shaikh, M.; Forsyth, C.B.; Tang, Y.; Fields, J.Z. NF-kappaB activation as a key mechanism in ethanol-induced disruption of the F-actin cytoskeleton and monolayer barrier integrity in intestinal epithelium. Alcohol 2007, 41, 447–460. [Google Scholar] [CrossRef]
- Hajifaraji, M.; Jahanjou, F.; Abbasalizadeh, F.; Aghamohammadzadeh, N.; Abbasi, M.M.; Dolatkhah, N. Effect of probiotic supplements in women with gestational diabetes mellitus on inflammation and oxidative stress biomarkers: A randomized clinical trial. Asia Pac. J. Clin. Nutr. 2018, 27, 581–591. [Google Scholar] [CrossRef]
- Kinalski, M.; Sledziewski, A.; Telejko, B.; Zarzycki, W.; Kinalska, I. Lipid peroxidation and scavenging enzyme activity in streptozotocin-induced diabetes. Acta Diabetol. 2000, 37, 179–183. [Google Scholar] [CrossRef]
- Fugmann, M.; Breier, M.; Rottenkolber, M.; Banning, F.; Ferrari, U.; Sacco, V.; Grallert, H.; Parhofer, K.G.; Seissler, J.; Clavel, T.; et al. The stool microbiota of insulin resistant women with recent gestational diabetes, a high risk group for type 2 diabetes. Sci. Rep. 2015, 5, 13212. [Google Scholar] [CrossRef] [PubMed]
- Kijmanawat, A.; Panburana, P.; Reutrakul, S.; Tangshewinsirikul, C. Effects of probiotic supplements on insulin resistance in gestational diabetes mellitus: A double-blind randomized controlled trial. J. Diabetes Investig. 2019, 10, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Babadi, M.; Khorshidi, A.; Aghadavood, E.; Samimi, M.; Kavossian, E.; Bahmani, F.; Mafi, A.; Shafabakhsh, R.; Satari, M.; Asemi, Z. The Effects of Probiotic Supplementation on Genetic and Metabolic Profiles in Patients with Gestational Diabetes Mellitus: A Randomized, Double-Blind, Placebo-Controlled Trial. Probiotics Antimicrob. Proteins 2019, 11, 1227–1235. [Google Scholar] [CrossRef] [PubMed]
- Badehnoosh, B.; Karamali, M.; Zarrati, M.; Jamilian, M.; Bahmani, F.; Tajabadi-Ebrahimi, M.; Jafari, P.; Rahmani, E.; Asemi, Z. The effects of probiotic supplementation on biomarkers of inflammation, oxidative stress and pregnancy outcomes in gestational diabetes. J. Matern. Fetal Neonatal Med. 2018, 31, 1128–1136. [Google Scholar] [CrossRef] [Green Version]
- Nabhani, Z.; Hezaveh, S.J.G.; Razmpoosh, E.; Asghari-Jafarabadi, M.; Gargari, B.P. The effects of symbiotic supplementation on insulin resistance/sensitivity, lipid profile and total antioxidant capacity in women with gestational diabetes mellitus: A randomized double blind placebo controlled clinical trial. Diabetes Res. Clin. Pract. 2018, 138, 149–157. [Google Scholar] [CrossRef]
- Jamilian, M.; Amirani, E.; Asemi, Z. The effects of vitamin D and probiotic co-supplementation on glucose homeostasis, inflammation, oxidative stress and pregnancy outcomes in gestational diabetes: A randomized, double-blind, placebo-controlled trial. Clin. Nutr. 2019, 38, 2098–2105. [Google Scholar] [CrossRef]
- Karamali, M.; Nasiri, N.; Taghavi Shavazi, N.; Jamilian, M.; Bahmani, F.; Tajabadi-Ebrahimi, M.; Asemi, Z. The Effects of Synbiotic Supplementation on Pregnancy Outcomes in Gestational Diabetes. Probiotics Antimicrob. Proteins 2018, 10, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, S.; Jamilian, M.; Tajabadi-Ebrahimi, M.; Jafari, P.; Asemi, Z. The effects of synbiotic supplementation on markers of insulin metabolism and lipid profiles in gestational diabetes: A randomised, double-blind, placebo-controlled trial. Br. J. Nutr. 2016, 116, 1394–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jafarnejad, S.; Saremi, S.; Jafarnejad, F.; Arab, A. Effects of a Multispecies Probiotic Mixture on Glycemic Control and Inflammatory Status in Women with Gestational Diabetes: A Randomized Controlled Clinical Trial. J. Nutr. Metab. 2016, 2016, 5190846. [Google Scholar] [CrossRef] [Green Version]
- Dolatkhah, N.; Hajifaraji, M.; Abbasalizadeh, F.; Aghamohammadzadeh, N.; Mehrabi, Y.; Abbasi, M.M. Is there a value for probiotic supplements in gestational diabetes mellitus? A randomized clinical trial. J. Health Popul. Nutr. 2015, 33, 25. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, K.L.; Brennan, L.; Kennelly, M.A.; Maguire, O.C.; Smith, T.; Curran, S.; Coffey, M.; Foley, M.E.; Hatunic, M.; Shanahan, F.; et al. Impact of probiotics in women with gestational diabetes mellitus on metabolic health: A randomized controlled trial. Am. J. Obstet. Gynecol. 2015, 212, e1–e11. [Google Scholar] [CrossRef]
- Guo, L.; Ma, J.; Tang, J.; Hu, D.; Zhang, W.; Zhao, X. Comparative Efficacy and Safety of Metformin, Glyburide, and Insulin in Treating Gestational Diabetes Mellitus: A Meta-Analysis. J. Diabetes Res. 2019, 2019, 9804708. [Google Scholar] [CrossRef] [Green Version]
- Koh, L.M.; Percival, B.; Pauley, T.; Pathak, S. Complementary therapy and alternative medicine: Effects on induction of labour and pregnancy outcome in low risk post-dates women. Heliyon 2019, 5, e02787. [Google Scholar] [CrossRef] [Green Version]
- Karamali, M.; Dastyar, F.; Badakhsh, M.H.; Aghadavood, E.; Amirani, E.; Asemi, Z. The Effects of Selenium Supplementation on Gene Expression Related to Insulin and Lipid Metabolism, and Pregnancy Outcomes in Patients with Gestational Diabetes Mellitus: A Randomized, Double-Blind, Placebo-Controlled Trial. Biol. Trace Elem. Res. 2020, 195, 1–8. [Google Scholar] [CrossRef]
- Gomez Ribot, D.; Diaz, E.; Fazio, M.V.; Gómez, H.L.; Fornes, D.; Macchi, S.B.; Gresta, C.A.; Capobianco, E.; Jawerbaum, A. An extra virgin olive oil-enriched diet improves maternal, placental, and cord blood parameters in GDM pregnancies. Diabetes Metab. Res. Rev. 2020, 36, e3349. [Google Scholar] [CrossRef]
- Jamilian, M.; Samimi, M.; Mirhosseini, N.; Afshar Ebrahimi, F.; Aghadavod, E.; Taghizadeh, M.; Asemi, Z. A Randomized Double-Blinded, Placebo-Controlled Trial Investigating the Effect of Fish Oil Supplementation on Gene Expression Related to Insulin Action, Blood Lipids, and Inflammation in Gestational Diabetes Mellitus-Fish Oil Supplementation and Gestational Diabetes. Nutrients 2018, 10, 163. [Google Scholar] [CrossRef] [Green Version]
- Fei, B.B.; Ling, L.; Hua, C.; Ren, S.Y. Effects of soybean oligosaccharides on antioxidant enzyme activities and insulin resistance in pregnant women with gestational diabetes mellitus. Food Chem. 2014, 158, 429–432. [Google Scholar] [CrossRef]
- Yuan, L.J.; Qin, Y.; Wang, L.; Zeng, Y.; Chang, H.; Wang, J.; Wang, B.; Wan, J.; Chen, S.H.; Zhang, Q.Y.; et al. Capsaicin-containing chili improved postprandial hyperglycemia, hyperinsulinemia, and fasting lipid disorders in women with gestational diabetes mellitus and lowered the incidence of large-for-gestational-age newborns. Clin. Nutr. 2016, 35, 388–393. [Google Scholar] [CrossRef]
- Sun, X.; Sun, H.; Zhang, J.; Ji, X. Artemisia Extract Improves Insulin Sensitivity in Women With Gestational Diabetes Mellitus by Up-Regulating Adiponectin. J. Clin. Pharmacol. 2016, 56, 1550–1554. [Google Scholar] [CrossRef]
- Asemi, Z.; Samimi, M.; Tabassi, Z.; Sabihi, S.S.; Esmaillzadeh, A. A randomized controlled clinical trial investigating the effect of DASH diet on insulin resistance, inflammation, and oxidative stress in gestational diabetes. Nutrition 2013, 29, 619–624. [Google Scholar] [CrossRef]
- Karamali, M.; Bahramimoghadam, S.; Sharifzadeh, F.; Asemi, Z. Magnesium-zinc-calcium-vitamin D co-supplementation improves glycemic control and markers of cardiometabolic risk in gestational diabetes: A randomized, double-blind, placebo-controlled trial. Appl. Physiol. Nutr. Metab. 2018, 43, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Jamilian, M.; Karamali, M.; Taghizadeh, M.; Sharifi, N.; Jafari, Z.; Memarzadeh, M.R.; Mahlouji, M.; Asemi, Z. Vitamin D and Evening Primrose Oil Administration Improve Glycemia and Lipid Profiles in Women with Gestational Diabetes. Lipids 2016, 51, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Hajimoosayi, F.; Jahanian Sadatmahalleh, S.; Kazemnejad, A.; Pirjani, R. Effect of ginger on the blood glucose level of women with gestational diabetes mellitus (GDM) with impaired glucose tolerance test (GTT): A randomized double-blind placebo-controlled trial. BMC Complement. Med. Ther. 2020, 20, 116. [Google Scholar] [CrossRef]
- Asemi, Z.; Karamali, M.; Esmaillzadeh, A. Effects of calcium-vitamin D co-supplementation on glycaemic control, inflammation and oxidative stress in gestational diabetes: A randomised placebo-controlled trial. Diabetologia 2014, 57, 1798–1806. [Google Scholar] [CrossRef]
- Gunasegaran, P.; Tahmina, S.; Daniel, M.; Nanda, S.K. Role of vitamin D-calcium supplementation on metabolic profile and oxidative stress in gestational diabetes mellitus: A randomized controlled trial. J. Obstet. Gynaecol. Res. 2021, 47, 1016–1022. [Google Scholar] [CrossRef]
- Ostadmohammadi, V.; Samimi, M.; Mobini, M.; Zarezade Mehrizi, M.; Aghadavod, E.; Chamani, M.; Dastorani, M.; Asemi, Z. The effect of zinc and vitamin E cosupplementation on metabolic status and its related gene expression in patients with gestational diabetes. J. Matern. Fetal Neonatal Med. 2019, 32, 4120–4127. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Lin, R.; Si, L.; Li, Z.; Jian, W.; Yu, Q.; Jia, Y. Cod-Liver Oil Improves Metabolic Indices and hs-CRP Levels in Gestational Diabetes Mellitus Patients: A Double-Blind Randomized Controlled Trial. J. Diabetes Res. 2019, 2019, 7074042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamilian, M.; Tabassi, Z.; Reiner, Ž.; Panahandeh, I.; Naderi, F.; Aghadavod, E.; Amirani, E.; Taghizadeh, M.; Shafabakhsh, R.; Satari, M.; et al. The effects of n-3 fatty acids from flaxseed oil on genetic and metabolic profiles in patients with gestational diabetes mellitus: A randomised, double-blind, placebo-controlled trial. Br. J. Nutr. 2020, 123, 792–799. [Google Scholar] [CrossRef]
- Asemi, Z.; Jamilian, M.; Mesdaghinia, E.; Esmaillzadeh, A. Effects of selenium supplementation on glucose homeostasis, inflammation, and oxidative stress in gestational diabetes: Randomized, double-blind, placebo-controlled trial. Nutrition 2015, 31, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Wang, G.; Wang, L.; Guo, N. Phytosterol nutritional supplement improves pregnancy and neonatal complications of gestational diabetes mellitus in a double-blind and placebo-controlled clinical study. Food Funct. 2017, 8, 424–428. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Su, S.; Yu, X.; Li, Y. Dietary epigallocatechin 3-gallate supplement improves maternal and neonatal treatment outcome of gestational diabetes mellitus: A double-blind randomised controlled trial. J. Hum. Nutr. Diet. 2017, 30, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekhar, D.; Jose, S.M.; Jomy, A.; Joseph, A.; Pradeep, A.; Geoji, A.S. Antiglycation property of passiflora edulis f. Flavicarpa deg. foliage in type 2 diabetic patients. Clin. Epidemiol. Glob. Health 2018, 7, P409–P412. [Google Scholar] [CrossRef] [Green Version]
- Yessoufou, A.; Gbenou, J.; Grissa, O.; Hichami, A.; Simonin, A.M.; Tabka, Z.; Moudachirou, M.; Moutairou, K.; Khan, N.A. Anti-hyperglycemic effects of three medicinal plants in diabetic pregnancy: Modulation of T cell proliferation. BMC Complement. Altern. Med. 2013, 13, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, S.; Lv, Y.; Li, N.; Huang, X.; Liu, X.; Li, H.; Wang, C.; Jia, Y.F. Biological investigations on therapeutic effect of chitosan encapsulated nano resveratrol against gestational diabetes mellitus rats induced by streptozotocin. Drug Deliv. 2020, 27, 953–963. [Google Scholar] [CrossRef] [PubMed]
- Usman, U.Z.; Bakar, A.B.A.; Mohamed, M. Propolis improves pregnancy outcomes and placental oxidative stress status in streptozotocin-induced diabetic rats. BMC Complement. Altern. Med. 2018, 18, 324. [Google Scholar] [CrossRef]
- Badr, G.; Mahmoud, M.H.; Farhat, K.; Waly, H.; Al-Abdin, O.Z.; Rabah, D.M. Maternal supplementation of diabetic mice with thymoquinone protects their offspring from abnormal obesity and diabetes by modulating their lipid profile and free radical production and restoring lymphocyte proliferation via PI3K/AKT signaling. Lipids Health Dis. 2013, 12, 37. [Google Scholar] [CrossRef] [Green Version]
- Afiune, L.A.F.; Leal-Silva, T.; Sinzato, Y.K.; Moraes-Souza, R.Q.; Soares, T.S.; Campos, K.E.; Fujiwara, R.T.; Herrera, E.; Damasceno, D.C.; Volpato, G.T. Beneficial effects of Hibiscus rosa-sinensis, L. flower aqueous extract in pregnant rats with diabetes. PLoS ONE 2017, 12, e0179785. [Google Scholar] [CrossRef] [PubMed]
- Lokman, E.F.; Saparuddin, F.; Muhammad, H.; Omar, M.H.; Zulkapli, A. Orthosiphon stamineus as a potential antidiabetic drug in maternal hyperglycemia in streptozotocin-induced diabetic rats. Integr. Med. Res. 2019, 8, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Laurino, L.F.; Viroel, F.J.M.; Caetano, E.; Spim, S.; Pickler, T.B.; Rosa-Castro, R.M.; Vasconcelos, E.A.; Jozala, A.F.; Hataka, A.; Grotto, D.; et al. Lentinus edodes Exposure before and after Fetus Implantation: Materno-Fetal Development in Rats with Gestational Diabetes Mellitus. Nutrients 2019, 11, 2720. [Google Scholar] [CrossRef] [Green Version]
- Barbalho, S.M.; Damasceno, D.C.; Spada, A.P.; da Silva, V.S.; Martuchi, K.A.; Oshiiwa, M.; Machado, F.M.; Mendes, C.G. Metabolic Profile of Offspring from Diabetic Wistar Rats Treated with Mentha piperita (Peppermint). Evid. Based Complement. Alternat. Med. 2011, 2011, 430237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abukhader, M.M. The effect of route of administration in thymoquinone toxicity in male and female rats. Indian J. Pharm. Sci. 2012, 74, 195–200. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Mendonça, E.L.S.S.; Fragoso, M.B.T.; de Oliveira, J.M.; Xavier, J.A.; Goulart, M.O.F.; de Oliveira, A.C.M. Gestational Diabetes Mellitus: The Crosslink among Inflammation, Nitroxidative Stress, Intestinal Microbiota and Alternative Therapies. Antioxidants 2022, 11, 129. https://doi.org/10.3390/antiox11010129
de Mendonça ELSS, Fragoso MBT, de Oliveira JM, Xavier JA, Goulart MOF, de Oliveira ACM. Gestational Diabetes Mellitus: The Crosslink among Inflammation, Nitroxidative Stress, Intestinal Microbiota and Alternative Therapies. Antioxidants. 2022; 11(1):129. https://doi.org/10.3390/antiox11010129
Chicago/Turabian Stylede Mendonça, Elaine Luiza Santos Soares, Marilene Brandão Tenório Fragoso, Jerusa Maria de Oliveira, Jadriane Almeida Xavier, Marília Oliveira Fonseca Goulart, and Alane Cabral Menezes de Oliveira. 2022. "Gestational Diabetes Mellitus: The Crosslink among Inflammation, Nitroxidative Stress, Intestinal Microbiota and Alternative Therapies" Antioxidants 11, no. 1: 129. https://doi.org/10.3390/antiox11010129