
A Novel Insight into Screening for Antioxidant Peptides from Hazelnut Protein: Based on the Properties of Amino Acid Residues

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Analysis
2.2.1. Inhibition Activity Assay of Linoleic Acid Oxidation
2.2.2. Superoxide Radical Scavenging Activity Assay
2.2.3. Metal Ion Chelation Activity Assay
2.3. Synthesis of Dipeptides
2.4. BIOPEP Database Analysis
2.5. Selection of Protease
2.6. Preparation of Hazelnut Protein Hydrolysates
2.7. Analysis of Peptide Sequence
2.8. DFT Calculations
2.9. Inhibition of Hazelnut Oil Oxidation Assay
2.10. Statistical Analysis
3. Results
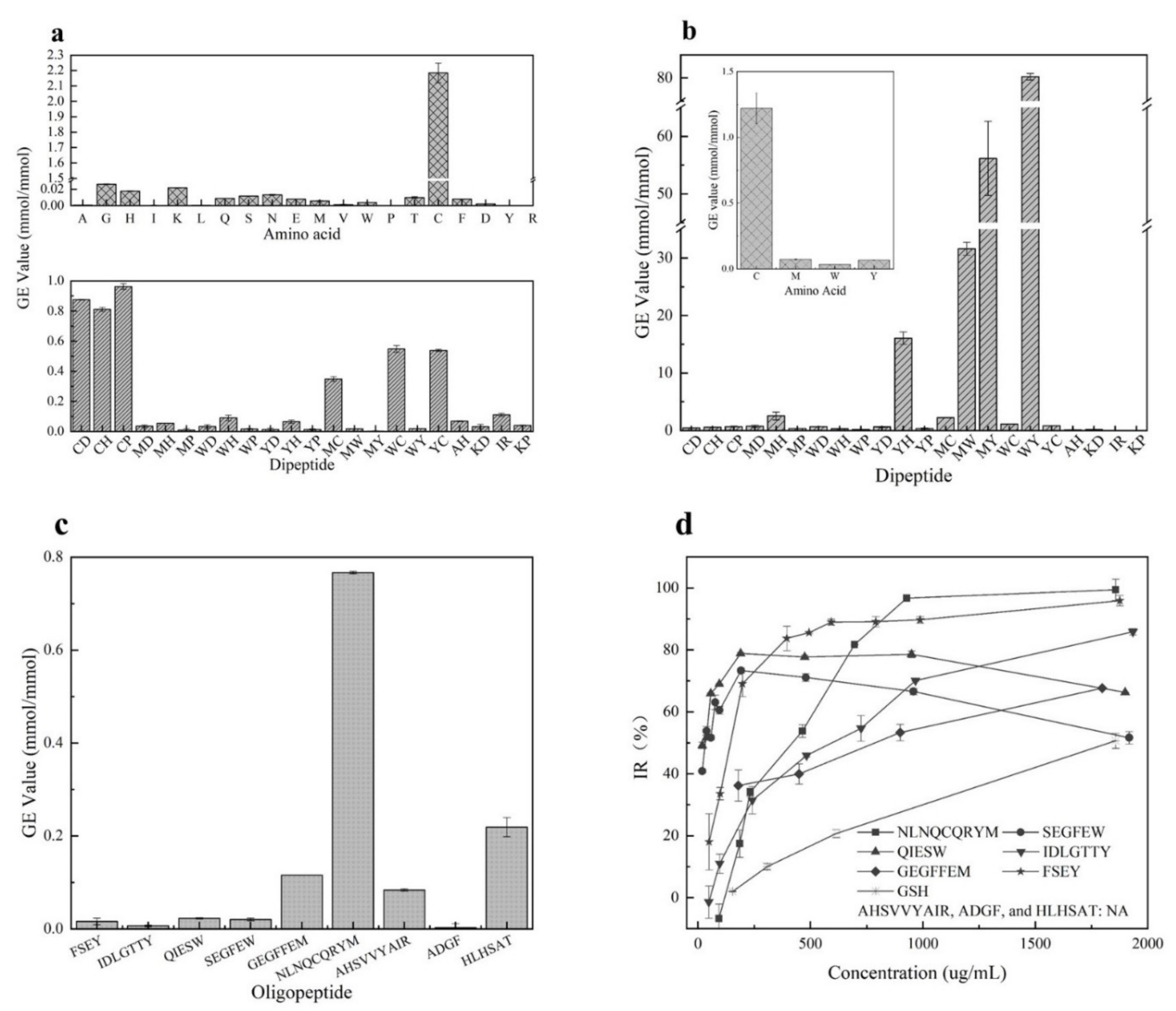
3.1. Antioxidant Activity of Amino Acids and Dipeptides
3.2. BIOPEP Database Analysis
3.3. Selection of Protease
3.4. Screening of Antioxidant Peptides
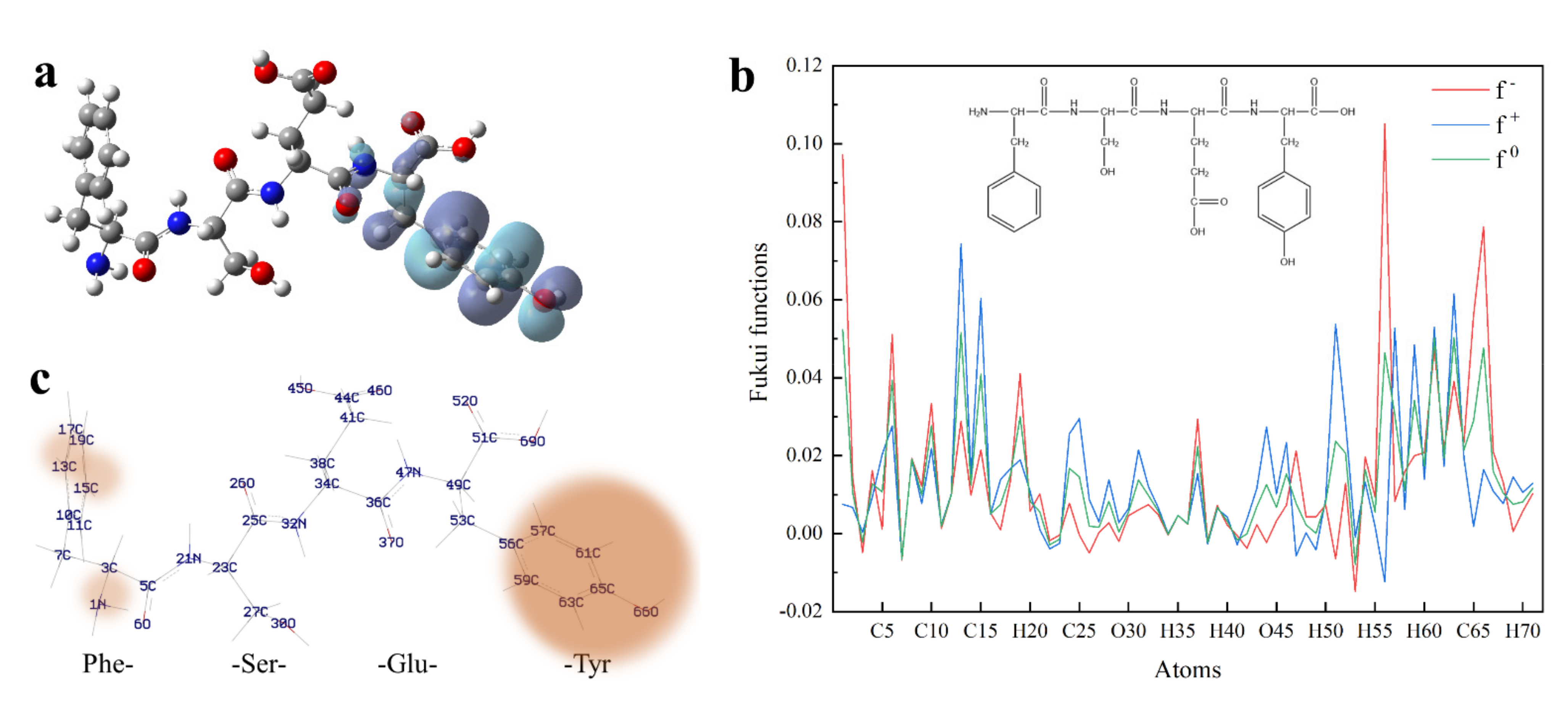
3.5. DFT Calculation of Peptides
3.5.1. Frontier Molecular Orbital Energy of Peptides
3.5.2. Fukui Function
3.6. Effects of Tyrosine Residue’s Location on the Antioxidant Activity of Peptides
3.7. Application of Hazelnut Peptide FSEY in Inhibiting Lipid Oxidation
4. Discussion
4.1. Chemical Test: We Found Four Key Amino Acid Residues
4.2. DFT Calculation: Tyr, Trp, Met, and Cys Are Active Sites
4.3. BIOPEP Database Analysis: Rules of Molecular Weight, GRAVY Value and Active Amino Acid Residue’s Location
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zheng, L. Structure-Activity Relationship and Directional Preparation of Antioxidant Peptide. Ph.D. Thesis, South China University of Technology, Guangzhou, China, 2015. [Google Scholar]
- Wen, C.; Zhang, J.; Zhang, H.; Duan, Y.; Ma, H. Plant protein-derived antioxidant peptides: Isolation, identification, mechanism of action and application in food systems: A review. Trends Food Sci. Technol. 2020, 105, 308–322. [Google Scholar]
- Matsui, R.; Honda, R.; Kanome, M.; Hagiwara, A.; Terashima, M. Designing antioxidant peptides based on the antioxidant properties of the amino acid side-chains. Food Chem. 2017, 245, 750. [Google Scholar]
- Jie, Y.; Zhao, H.; Sun, X.; Lv, X.; Zhang, Z.; Zhang, B. Isolation of antioxidative peptide from the protein hydrolysate of Caragana ambigua seeds and its mechanism for retarding lipid auto-oxidation. J. Sci. Food Agric. 2019, 99, 3078–3085. [Google Scholar]
- Lafarga, T.; O’Connor, P.; Hayes, M. Identification of novel dipeptidyl peptidase-IV and angiotensin-I-converting enzyme inhibitory peptides from meat proteins using in silico analysis. Peptides 2014, 59, 53–62. [Google Scholar]
- Feng, Y.; Wang, Z.; Chen, J.; Li, H.; Wang, Y.; Ren, D.; Lu, J. Separation, identification, and molecular docking of tyrosinase inhibitory peptides from the hydrolysates of defatted walnut (Juglans regia L.) meal. Food Chem. 2021, 353, 129471. [Google Scholar]
- Jiang, J.; Liang, L.; Ma, Q.; Zhao, T. Kernel nutrient composition and antioxidant ability of Corylus spp. in China. Front. Plant Sci. 2021, 12, 690966. [Google Scholar]
- Moscetti, R.; Saeys, W.; Keresztes, J.C.; Goodarzi, M.; Cecchini, M.; Danilo, M.; Massantini, R. Hazelnut Quality Sorting Using High Dynamic Range Short-Wave Infrared Hyperspectral Imaging. Food Bioprocess Technol. 2015, 8, 1593–1604. [Google Scholar]
- Zhong, Y. Hazelnut Protein Physical Modification Methods and the Physical and Chemical Properties Change Research. Master’s Thesis, Shenyang Agricultural university, Shenyang, China, 2017. [Google Scholar]
- Çağlar, A.F.; Çakır, B.; Gülseren, İ. LC-Q-TOF/MS based identification and in silico verification of ACE-inhibitory peptides in Giresun (Turkey) hazelnut cakes. Eur. Food Res. Technol. 2021, 247, 1189–1198. [Google Scholar]
- Sabeena Farvin, K.H.; Andersen, L.L.; Nielsen, H.H.; Jacobsen, C.; Jakobsen, G.; Johansson, I.; Jessen, F. Antioxidant activity of Cod (Gadus morhua) protein hydrolysates: In vitro assays and evaluation in 5% fish oil-in-water emulsion. Food Chem. 2014, 149, 326–334. [Google Scholar]
- Sakanaka, S.; Tachibana, Y.; Ishihara, N.; Raj Juneja, L. Antioxidant activity of egg-yolk protein hydrolysates in a linoleic acid oxidation system. Food Chem. 2004, 86, 99–103. [Google Scholar]
- Alashi, A.M.; Blanchard, C.L.; Mailer, R.J.; Agboola, S.O.; Mawson, A.J.; He, R.; Girgih, A.; Aluko, R.E. Antioxidant properties of Australian canola meal protein hydrolysates. Food Chem. 2014, 146, 500–506. [Google Scholar]
- Li, Y.; Bo, J.; Tao, Z.; Mu, W.; Jian, L. Antioxidant and free radical-scavenging activities of chickpea protein hydrolysate (CPH). Food Chem. 2008, 106, 444–450. [Google Scholar]
- Zhang, T.; Li, Y.; Miao, M.; Jiang, B. Purification and characterisation of a new antioxidant peptide from chickpea (Cicer arietium L.) protein hydrolysates. Food Chem. 2011, 128, 28–33. [Google Scholar]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM database of bioactive peptides: Current opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar]
- Piotr, M.; Jerzy, D.; Małgorzata, D.; Anna, I.; Nałecz, M. Food Peptidomics. Food Technol. Biotechnol. 2008, 46, 1–10. [Google Scholar]
- Piotr, M.; Jerzy, D.; Anna, I.; Marta, D.; Magorzata, D. BIOPEP Database and Other Programs for Processing Bioactive Peptide Sequences. J. AOAC Int. 2019, 4, 965–980. [Google Scholar]
- Iwaniak, A.; Minkiewicz, P.; Darewicz, M.; Sieniawski, K.; Starowicz, P. BIOPEP database of sensory peptides and amino acids. Food Res. Int. 2016, 85, 155–161. [Google Scholar]
- Chatterjee, A.; Kanawjia, S.K.; Khetra, Y.; Saini, P. Discordance between in silico & in vitroanalyses of ACE inhibitory & antioxidative peptides from mixed milk tryptic whey protein hydrolysate. J. Food Sci. Technol. 2015, 52, 5621–5630. [Google Scholar]
- Tatar, F.; Tunç, M.T.; Kahyaoglu, T. Turkish Tombul hazelnut (Corylus avellana L.) protein concentrates: Functional and rheological properties. J. Food Sci. Technol. 2015, 52, 1024–1031. [Google Scholar]
- Liu, C.; Ren, D.; Li, J.; Fang, L.; Wang, J.; Liu, J.; Min, W. Cytoprotective effect and purification of novel antioxidant peptides from hazelnut (C. heterophylla Fisch) protein hydrolysates. J. Funct. Foods 2018, 42, 203–215. [Google Scholar]
- Chen, Y.; Lv, C.; Han, J.; Ji, Y.; Fu, Q.; Liu, L.; Xu, J. Separation, Purification and Sequence Analysis of Antioxidative Peptides from the Halnut Meal Protein Hydrolysate. Sci. Technol. Food Ind. 2018, 39, 114–118. [Google Scholar]
- Wen, C.; Zhang, J.; Zhang, H.; Duan, Y.; Ma, H. Study on the structure–activity relationship of watermelon seed antioxidant peptides by using molecular simulations. Food Chem. 2021, 364, 130432. [Google Scholar]
- Zacharias, A.O.; Varghese, A.; Akshaya, K.B.; Savitha, M.S.; George, L. DFT, spectroscopic studies, NBO, NLO and Fukui functional analysis of 1-(1-(2,4-difluorophenyl)-2-(1H-1,2,4-triazol-1-yl)ethylidene) thiosemicarbazide. J. Mol. Struct. 2018, 1158, 1–13. [Google Scholar] [CrossRef]
- Sevvanthi, S.; Muthu, S.; Raja, M.; Aayisha, S.; Janani, S. PES, molecular structure, spectroscopic (FT-IR, FT-Raman), electronic (UV-Vis, HOMO-LUMO), quantum chemical and biological (docking) studies on a potent membrane permeable inhibitor: Dibenzoxepine derivative. Heliyon 2020, 6, e04724. [Google Scholar]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. Gaussian 09, Revision D.01; Gaussian, Inc.: Wallingford, CT, USA, 2013. [Google Scholar]
- Shi, C.; Liu, M.; Ma, Q.; Zhao, T.; Liang, L.; Zhang, B. Impact of tetrapeptide-FSEY on oxidative and physical stability of hazelnut oil-in-water emulsion. Foods 2021, 10, 1400. [Google Scholar]
- Vaisali, C.; Belur, P.D.; Regupathi, I. Comparison of antioxidant properties of phenolic compounds and their effectiveness in imparting oxidative stability to sardine oil during storage. LWT-Food Sci. Technol. 2016, 69, 153–160. [Google Scholar]
- Di Mattia, C.D.; Sacchetti, G.; Mastrocola, D.; Sarker, D.K.; Pittia, P. Surface properties of phenolic compounds and their influence on the dispersion degree and oxidative stability of olive oil O/W emulsions. Food Hydrocoll. 2010, 24, 652–658. [Google Scholar]
- Pei, Y.; Deng, Q.; McClements, D.; Li, J.; Li, B. Impact of Phytic Acid on the Physical and Oxidative Stability of Protein-Stabilized Oil-in-Water Emulsions. Food Biophys. 2020, 15, 433–441. [Google Scholar]
- Zeng, Z. Study on Structure-Activity Relationships of Antioxidant Peptide through Enzymatic by Quantum Chemistry; Changsha University of Science & Technology: Changsha, China, 2013. [Google Scholar]
- Huang, W.Y.; Majumder, K.; Wu, J. Oxygen radical absorbance capacity of peptides from egg white protein ovotransferrin and their interaction with phytochemicals. Food Chem. 2010, 123, 635–641. [Google Scholar]
- Keil, B. Specificity of Proteolysis; Springer: Berlin/Heidelberg, Germany, 1992. [Google Scholar]
- Tacias-Pascacio, V.G.; Morellon-Sterling, R.; Siar, E.-H.; Tavano, O.; Berenguer-Murcia, Á.; Fernandez-Lafuente, R. Use of alcalase in the production of bioactive peptides: A review. Int. J. Biol. Macromol. 2020, 165, 2143–2196. [Google Scholar]
- Arise, A.K.; Alashi, A.M.; Nwachukwu, I.D.; Malomo, S.A.; Amonsou, E.O. Inhibitory properties of bambara groundnut protein hydrolysate and peptide fractions against angiotensin converting enzymes, renin and free radicals. J. Sci. Food Agric. 2016, 97, 2834–2841. [Google Scholar]
- Ngamsuk, S.; Hsu, J.-L.; Huang, T.-C.; Suwannaporn, P. Ultrasonication of Milky Stage Rice Milk with Bioactive Peptides from Rice Bran: Its Bioactivities and Absorption. Food Bioprocess Technol. 2020, 13, 462–474. [Google Scholar]
- Antonczak, S. Electronic description of four flavonoids revisited by DFT method. J. Mol. Struc.-Theochem 2008, 856, 38–45. [Google Scholar]
- Wang, M.; Li, C.; Li, H.; Wu, Z.; Chen, B.; Lei, Y.; Shen, Y. In Vitro and In Silico Antioxidant Activity of Novel Peptides Prepared from Paeonia Ostii ‘Feng Dan’ Hydrolysate. Antioxidants 2019, 8, 433. [Google Scholar]
- Srivastava, A.K.; Pandey, A.K.; Jain, S.; Misra, N. FT-IR spectroscopy, intra-molecular C−H⋯O interactions, HOMO, LUMO, MESP analysis and biological activity of two natural products, triclisine and rufescine: DFT and QTAIM approaches. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2015, 136, 682–689. [Google Scholar]
- Wu, R.; Huang, J.; Huan, R.; Chen, L.; Yi, C.; Liu, D.; Wang, M.; Liu, C.; He, H. New insights into the structure-activity relationships of antioxidative peptide PMRGGGGYHY. Food Chem. 2021, 337, 127678. [Google Scholar]
- Hougland, J.L.; Darling, J.; Flynn, S. Protein Posttranslational Modification; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013. [Google Scholar]
- Zheng, L.; Zhao, Y.; Dong, H.; Su, G.; Zhao, M. Structure activity relationship of antioxidant dipeptides: Dominant role of Tyr, Trp, Cys and Met residues. J. Funct. Foods 2016, 21, 485–496. [Google Scholar]
- Uno, S.; Kodama, D.; Yukawa, H.; Shidara, H.; Akamatsu, M. Quantitative analysis of the relationship between structure and antioxidant activity of tripeptides. J. Pept. Sci. 2020, 26, e3238. [Google Scholar]
- Wang, J.; Chen, C.; Xu, Y.; Jia, C.; Zhang, B.; Niu, M.; Zhao, S.; Xiong, S. Selection of Antioxidant Peptides from Gastrointestinal Hydrolysates of Fermented Rice Cake by Combining Peptidomics and Bioinformatics. ACS Food Sci. Technol. 2021, 1, 443–452. [Google Scholar]
- Pihlanto, A. Antioxidative peptides derived from milk proteins. Int. Dairy J. 2006, 16, 1306–1314. [Google Scholar]
- Wu, H. Structure-Activity Relationship of Antioxidative Peptides Derived from Whey Protien. Master’s Thesis, Jiangnan University, Wuxi, China, 2011. [Google Scholar]
- Berton-Carabin, C.C.; Ropers, M.H.; Genot, C. Lipid Oxidation in Oil-in-Water Emulsions: Involvement of the Interfacial Layer. Compr. Rev. Food Sci. F 2014, 13, 945–977. [Google Scholar]
- Chen, H.; Muramoto, K.; Yamauchi, F. Structural analysis of antioxidative peptides from soybean beta-conglycinin. J. Agric. Food. Chem. 1995, 43, 574–578. [Google Scholar]
- Suetsuna, K. Antioxidant Peptides from the Protease Digest of Prawn (Penaeus japonicus) Muscle. Mar. Biotechnol. 2000, 2, 5–10. [Google Scholar]
- Raja, M.; Raj Muhamed, R.; Muthu, S.; Suresh, M.; Muthu, K. Synthesis, spectroscopic (FT-IR, FT-Raman, NMR, UV–Visible), Fukui function, antimicrobial and molecular docking study of (E)-1-(3-bromobenzylidene)semicarbazide by DFT method. J. Mol. Struct. 2017, 1130, 374–384. [Google Scholar]
- Shen, S.; Chahal, B.; Majumder, K.; You, S.J.; Wu, J. Identification of Novel Antioxidative Peptides Derived from a Thermolytic Hydrolysate of Ovotransferrin by LC-MS/MS. J. Agric. Food. Chem. 2010, 58, 7664–7672. [Google Scholar]
- Guo, H.; Kouzuma, Y.; Yonekura, M. Structures and properties of antioxidative peptides derived from royal jelly protein. Food Chem. 2009, 113, 238–245. [Google Scholar]
- Torkova, A.; Koroleva, O.; Khrameeva, E.; Fedorova, T.; Tsentalovich, M. Structure-Functional Study of Tyrosine and Methionine Dipeptides: An Approach to Antioxidant Activity Prediction. Int. J. Mol. Sci. 2015, 16, 25353–25376. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EC Number | Name | Preferential Cleavage Sites | Inhibition Rate (IR, %) |
|---|---|---|---|
| 3.4.21.1 | Chymotrypsin | Cleaves, Tyr-|-Xaa, Trp-|-Xaa, Phe-|-Xaa, and Leu-|-Xaa | —1 |
| 3.4.21.62 | Alcalase Novo/ Bacillus subtilis alkaline proteinase | Hydrolysis of proteins with broad specificity for peptide bonds, especially aromatic or hydrophobic amino acids. Cleaves, Glu-|-Xaa, Met-|-Xaa, Leu-|-Xaa, Tyr-|-Xaa, Lys-|-Xaa, Trp-|-Xaa, and Gln-|-Xaa | 95.11 ± 0.71 |
| 3.4.23.1 | Pepsin A (pH = 1.3) | Preferential cleavage, hydrophobic, preferably aromatic. Cleaves, Phe-|-Val, Gln-|-His, Glu-|-Ala, Ala-|-Leu, Leu-|-Tyr, Tyr-|-Leu, Gly-|-Phe, Phe-|-Phe, and Phe-|-Tyr | —1 |
| 3.4.24.27 | Bacillolysin/ Bacillus subtilis neutral proteinase | Cleaves, Xaa-|-Leu> Xaa-|-Phe | 81.44 ± 1.94 |
| Number | Sequence | Length | Mw (Da) | GRAVY | Features |
|---|---|---|---|---|---|
| 1 | FSEY | 4 | 544.55 | −0.70 | With Tyr, Trp, or Met residue at the C-terminal. |
| 2 | QIESW | 5 | 661.70 | −0.84 | |
| 3 | SEGFEW | 6 | 753.75 | −1.05 | |
| 4 | IDLGTTY | 7 | 781.85 | 0.24 | |
| 5 | GEGFFEM | 7 | 815.88 | −0.04 | |
| 6 | AHSVVYAIR | 9 | 1015.16 | 0.74 | With Tyr residue in the sequence. |
| 7 | NLNQCQRYM | 9 | 1169.33 | −1.29 | With Cys, Tyr, and Met residues in the sequence. |
| 8 | HLHSAT | 6 | 664.71 | −0.38 | With ABTS and DPPH radical scavenging ability. |
| 9 | ADGF | 4 | 408.40 | 0.18 |
| Dipeptide | EHOMO | ELUMO | E-Gap | GE Value | |
|---|---|---|---|---|---|
| With GE value higher than 1 | WC | −5.61 | −0.73 | 4.87 | 1.13 ± 0.04 |
| WY | −5.59 | −0.61 | 4.98 | 80.28 ± 0.68 | |
| MW | −5.91 | −0.75 | 5.15 | 31.59 ± 1.13 | |
| MC | −6.21 | −0.88 | 5.33 | 2.24 ± 0.03 | |
| MY | −6.08 | −0.59 | 5.49 | 56.17 ± 6.44 | |
| MH | −5.91 | −0.52 | 5.39 | 2.56 ± 0.68 | |
| YH | −5.99 | −0.34 | 5.65 | 16.06 ± 1.09 | |
| With GE value between 0.5~1 | WD | −5.75 | −0.61 | 5.13 | 0.70 ± 0.02 |
| MD | −6.17 | −0.73 | 5.45 | 0.75 ± 0.21 | |
| YC | −6.29 | −0.63 | 5.66 | 0.85 ± 0.04 | |
| YD | −6.26 | −0.60 | 5.66 | 0.64 ± 0.08 | |
| CH | −6.17 | −0.47 | 5.70 | 0.57 ± 0.04 | |
| With GE value less than 0.5 | WH | −4.77 | −0.29 | 4.48 | 0.33 ± 0.03 |
| WP | −5.59 | −0.52 | 5.07 | 0.20 ± 0.00 | |
| YP | −6.14 | −0.47 | 5.67 | 0.36 ± 0.12 | |
| KD | −6.42 | −0.68 | 5.75 | 0.22 ± 0.03 | |
| CD | −6.63 | −0.76 | 5.88 | 0.46 ± 0.01 | |
| IR | −6.27 | −0.16 | 5.94 | 0.00 ± 0.00 | |
| AH | −6.54 | −0.34 | 6.00 | 0.12 ± 0.04 | |
| MP | −6.16 | −0.14 | 6.02 | 0.35 ± 0.04 | |
| KP | −6.34 | −0.05 | 6.29 | 0.00 ± 0.00 | |
| CP | −6.57 | −0.18 | 6.39 | 0.70 ± 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, C.; Liu, M.; Zhao, H.; Lv, Z.; Liang, L.; Zhang, B. A Novel Insight into Screening for Antioxidant Peptides from Hazelnut Protein: Based on the Properties of Amino Acid Residues. Antioxidants 2022, 11, 127. https://doi.org/10.3390/antiox11010127
Shi C, Liu M, Zhao H, Lv Z, Liang L, Zhang B. A Novel Insight into Screening for Antioxidant Peptides from Hazelnut Protein: Based on the Properties of Amino Acid Residues. Antioxidants. 2022; 11(1):127. https://doi.org/10.3390/antiox11010127
Chicago/Turabian StyleShi, Chenshan, Miaomiao Liu, Hongfei Zhao, Zhaolin Lv, Lisong Liang, and Bolin Zhang. 2022. "A Novel Insight into Screening for Antioxidant Peptides from Hazelnut Protein: Based on the Properties of Amino Acid Residues" Antioxidants 11, no. 1: 127. https://doi.org/10.3390/antiox11010127