
Alpha-Ketoglutarate: A Potential Inner Mitochondrial and Cytosolic Protector against Peroxynitrite and Peroxynitrite-Induced Nitration?

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Nuclear Magnetic Resonance Spectroscopy
2.3. Preparation of ONOO−
2.4. Enzymatically Colorimetric αKG Quantification Assay Kit
2.5. ONOO−—Luminol Measurements Using αKG
2.6. Nitration of BSA with ONOO−
2.7. Estimation of Nitrated Tyrosine BSA by an ELISA Technique
2.8. Statistical Analysis
3. Results
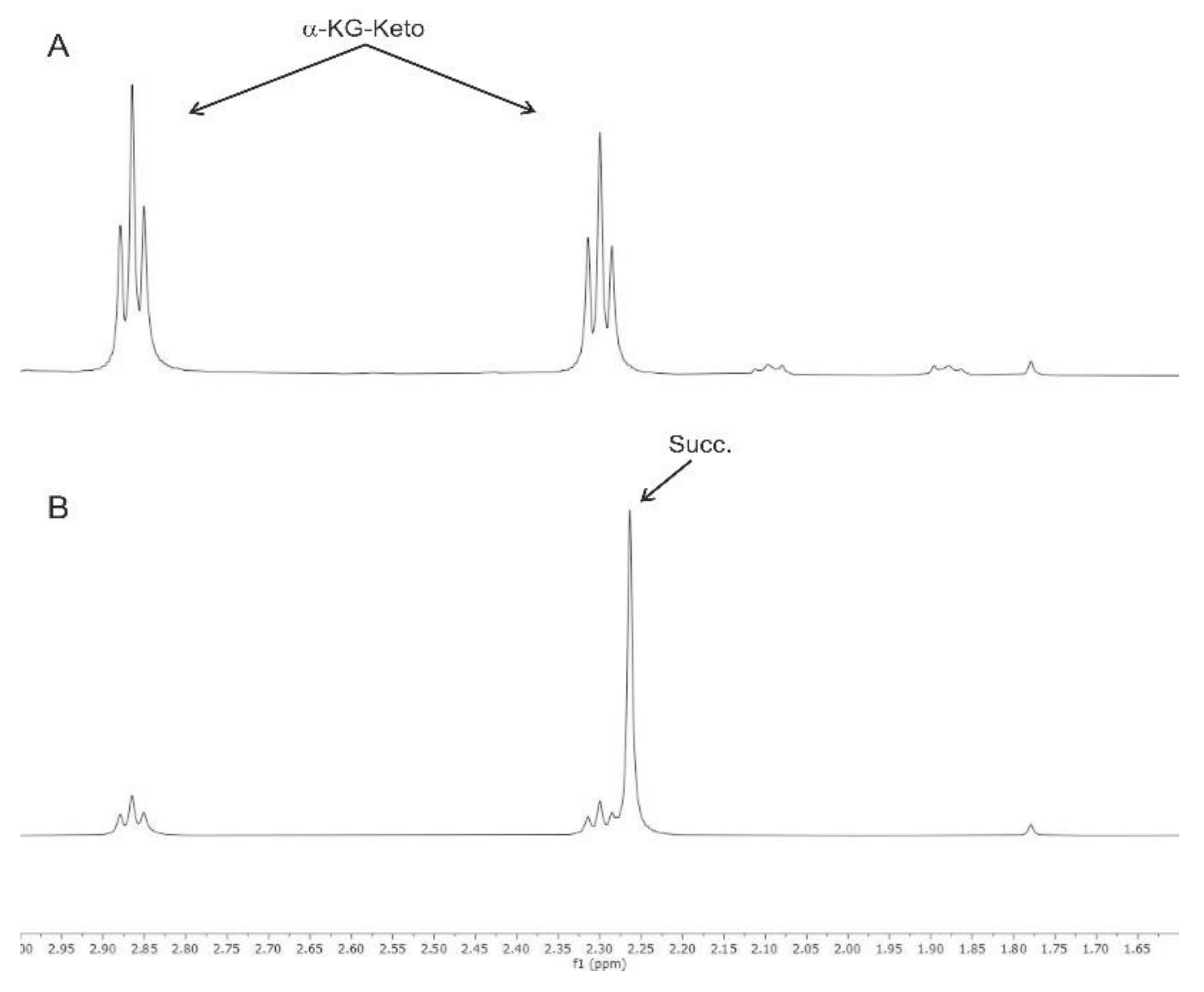
3.1. Nuclear Magnetic Resonance Spectroscopy
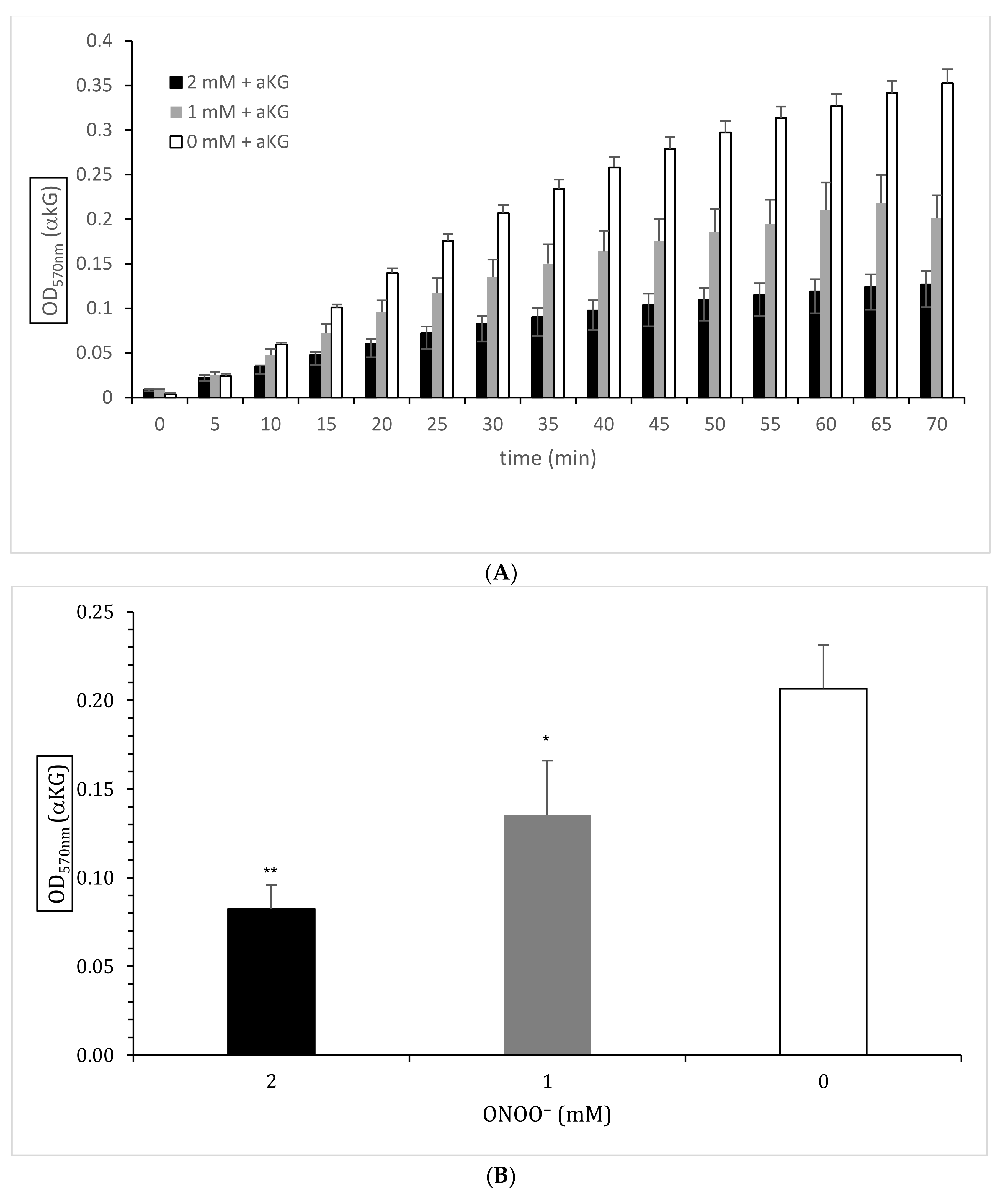
3.2. Enzymatically Colorimetric Assay Kit for αKG Quantification
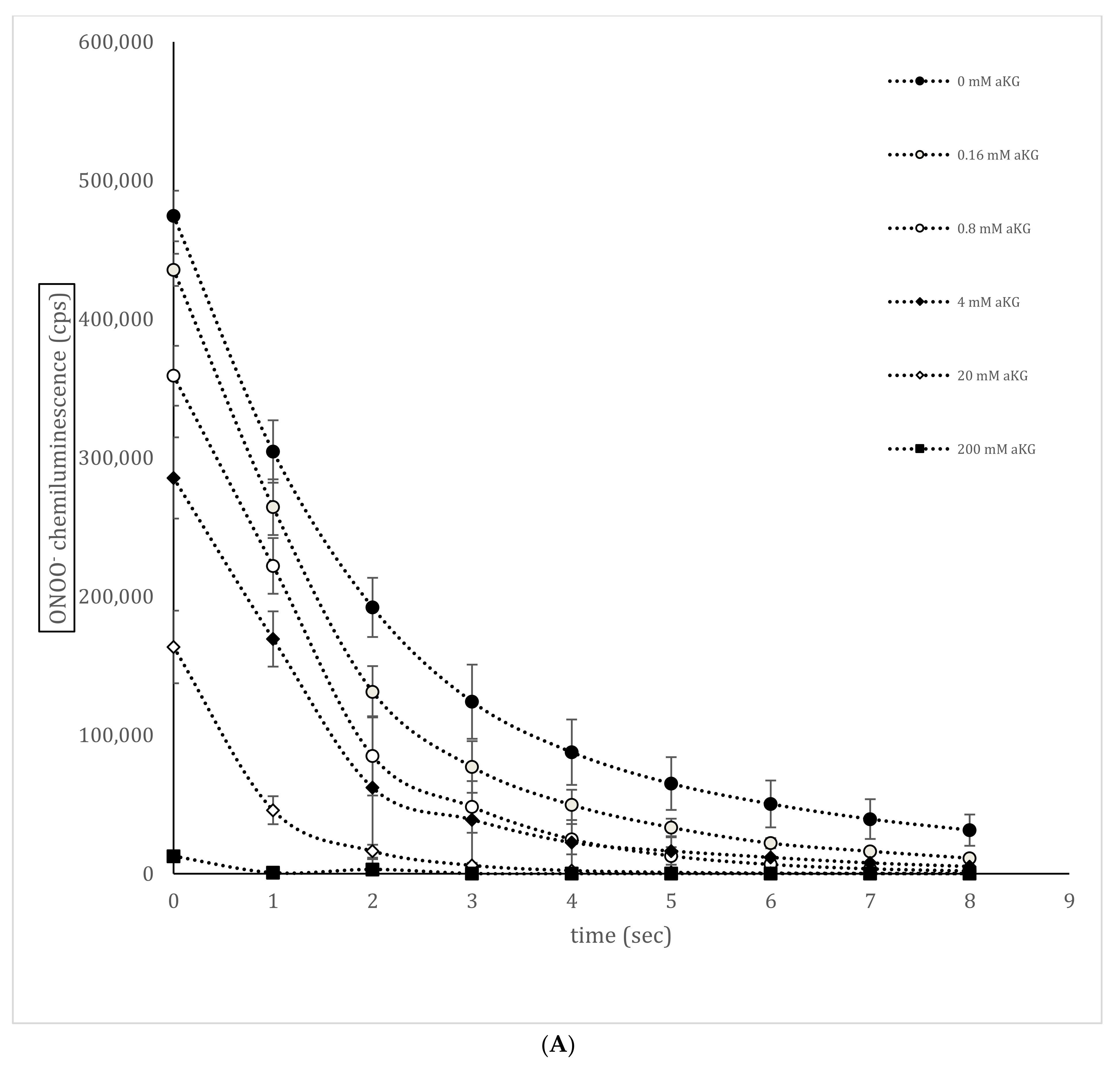
3.3. ONOO−—Luminol-Induced Chemiluminescence in the Presence or Absence of αKG
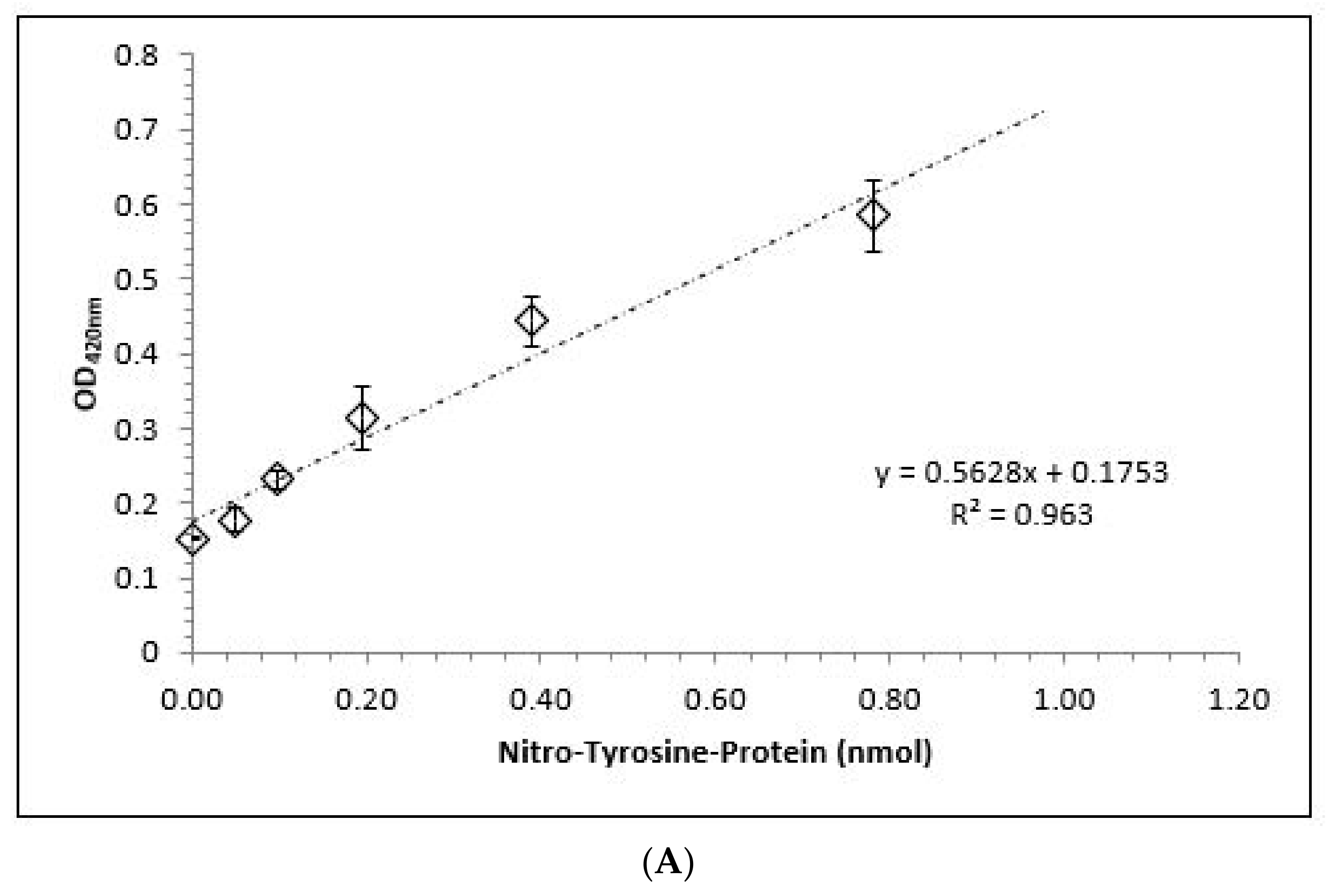
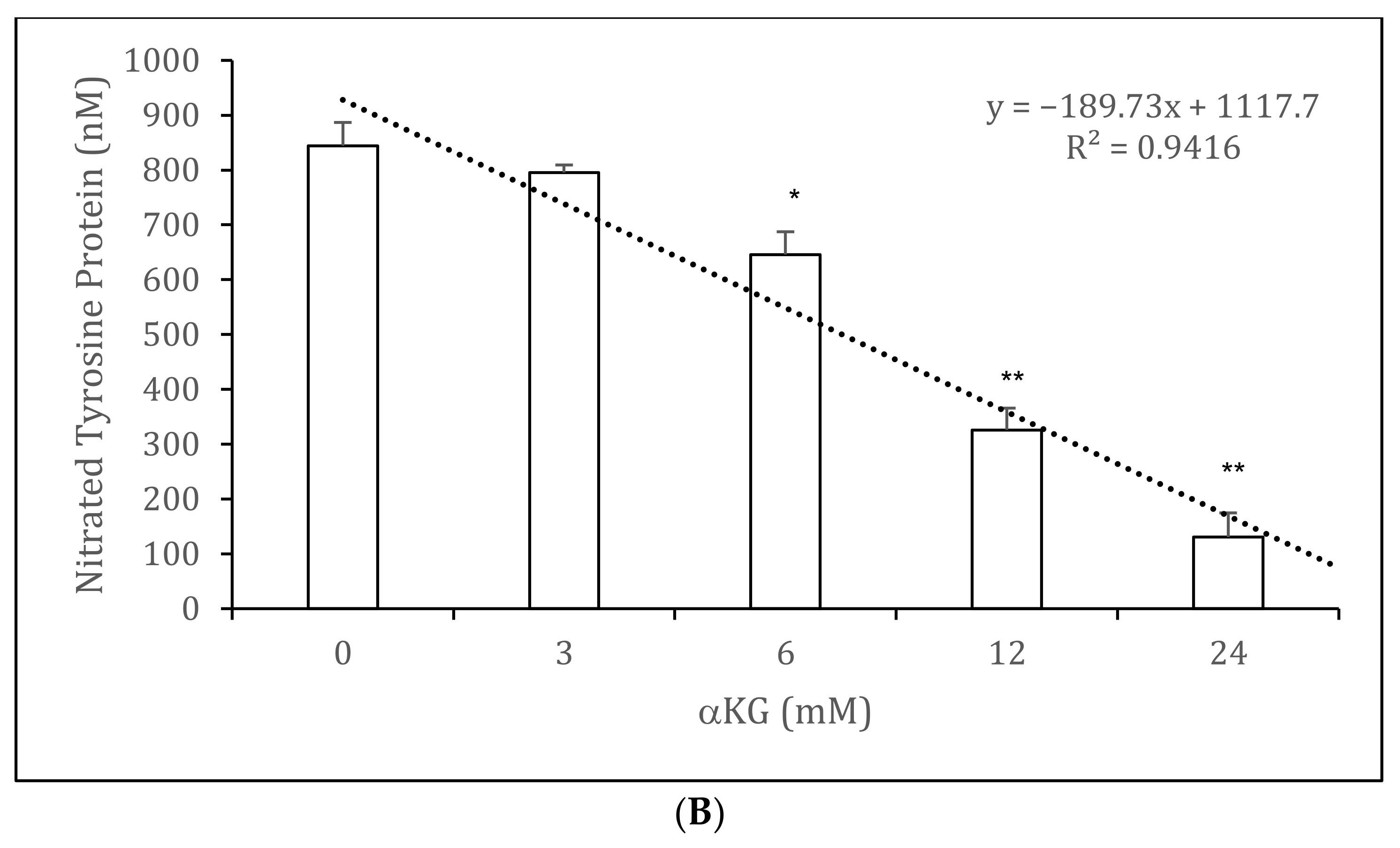
3.4. Nitrated Tyrosine Estimation on BSA in the Presence and Absence of αKG
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matzi, V.; Lindenmann, J.; Muench, A.; Greilberger, J.; Juan, H.; Wintersteiger, R.; Maier, A.; Smolle-Juettner, F.M. The impact of preoperative micronutrient supplementation in lung surgery. A prospective randomized trial of oral supplementation of combined alpha-ketoglutaric acid and 5-hydroxymethylfurfural. Eur. J. Cardiothorac. Surg. 2007, 32, 776–782. [Google Scholar] [CrossRef] [Green Version]
- Andrä, M.; Russ, M.; Jauk, S.; Lamacie, M.; Lang, I.; Arnold, R.; Brcic, I.; Santos, R.; Wintersteiger, R.; Ortner, A. Antioxidant Solution in Combination with Angiotensin-(1-7) Provides Myocardial Protection in Langendorff-Perfused Rat Hearts. Oxid. Med. Cell. Longev. 2020. [Google Scholar] [CrossRef] [PubMed]
- Nath, K.A.; Ngo, E.O.; Hebbel, R.P.; Croatt, A.J.; Zhou, B.; Nutter, L.M. alpha-Ketoacids scavenge H2O2 in vitro and in vivo and reduce menadione-induced DNA injury and cytotoxicity. Am. J. Physiol. 1995, 268, C227–C236. [Google Scholar] [CrossRef] [PubMed]
- Long, L.H.; Halliwell, B. Artefacts in cell culture: α-Ketoglutarate can scavenge hydrogen peroxide generated by ascorbate and epigallocatechin gallate in cell culture media. Biochem. Biophys. Res. Commun. 2011, 406, 20–24. [Google Scholar] [CrossRef]
- Greilberger, J.F.; Wintersteiger, R.; Astrid, O.; Greilberger, M.; Herwig, R. Combination of 2-oxoglutarate/ascorbic acid/5-hydroxy-methyl-furfur-aldehyde/carnosine inhibits protein oxidation during radical exposure of cigarette smoke. Proteins 2018, 5, 10–12. [Google Scholar] [CrossRef]
- Blough, N.V.; Zafiriou, O.C. Reaction of superoxide with nitric oxide to form peroxonitrite in alkaline aqueous solution. Inorg. Chem. 1985, 24, 3502–3504. [Google Scholar] [CrossRef]
- Viner, R.I.; Ferrington, D.A.; Williams, T.D.; Bigelow, D.J.; Schöneich, C. Protein modification during biological aging: Selective tyrosine nitration of the SERCA2a isoform of the sarcoplasmic reticulum Ca2+-ATPase in skeletal muscle. Biochem. J. 1999, 340, 657–669. [Google Scholar] [CrossRef]
- Ferrer-Sueta, G.; Campolo, N.; Trujillo, M.; Bartesaghi, S.; Carballal, S.; Romero, N.; Alvarez, B.; Radi, R. Biochemistry of peroxynitrite and protein tyrosine nitration. Chem. Rev. 2018, 118, 1338–1408. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [Green Version]
- Estévez, A.G.; Spear, N.; Manuel, S.M.; Radi, R.; Henderson, C.E.; Barbeito, L.; Beckman, J.S. Nitric oxide and superoxide contribute to motor neuron apoptosis induced by trophic factor deprivation. J. Neurosci. 1998, 18, 923–931. [Google Scholar] [CrossRef]
- Cassina, P.; Cassina, A.; Pehar, M.; Castellanos, R.; Gandelman, M.; de León, A.; Robinson, K.M.; Mason, R.P.; Beckman, J.S.; Barbeito, L.; et al. Mitochondrial dysfunction in SOD1G93A-bearing astrocytes promotes motor neuron degeneration: Prevention by mitochondrial-targeted antioxidants. J. Neurosci. 2008, 28, 4115–4122. [Google Scholar] [CrossRef] [Green Version]
- Fiuza, B.; Subelzú, N.; Calcerrada, P.; Straliotto, M.R.; Piacenza, L.; Cassina, A.; da Rocha, J.B.T.; Radi, R.; de Bem, A.F.; Peluffo, G. Impact of SIN-1-derived peroxynitrite flux on endothelial cell redox homeostasis and bioenergetics: Protective role of diphenyl diselenide via induction of peroxiredoxins. Free. Radic. Res. 2015, 49, 122–132. [Google Scholar] [CrossRef]
- Hooper, D.; Spitsin, S.; Kean, R.B.; Champion, J.M.; Dickson, G.M.; Chaudhry, I.; Koprowski, H. Uric acid, a natural scavenger of peroxynitrite, in experimental allergic encephalomyelitis and multiple sclerosis. Proc. Natl. Acad. Sci. USA 1998, 95, 675–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 5th ed.; Oxford University Press: New York, NY, USA, 2015. [Google Scholar]
- Hughes, M.N.; Nicklin, H.G. The chemistry of pernitrites. Part I. Kinetics of decomposition of pernitrous acid. J. Chem. Soc. A 1968, 450–452. [Google Scholar] [CrossRef]
- Radi, R.; Cosgrove, T.P.; Beckman, J.S.; Freeman, B.A. Peroxynitrite-induced luminol chemiluminescence. Biochem. J. 1993, 290, 51–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatassery, G.T.; Lai, J.C.; DeMaster, E.G.; Smith, W.E.; Quach, H.T. Oxidation of vitamin E and vitamin C and inhibition of brain mitochondrial oxidative phosphorylation by peroxynitrite. J. Neurosci. Res. 2004, 75, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Zdzisińska, B.; Żurek, A.; Kandefer-Szerszeń, M. Alpha-ketoglutarate as a molecule with pleiotropic activity: Well-known and novel possibilities of therapeutic use. Arch. Immunol. Ther. Exp. 2017, 65, 21–36. [Google Scholar] [CrossRef] [Green Version]
- Van der Vliet, A.; Hristova, M.; Cross, C.E.; Eiserich, J.P.; Goldkorn, T. Peroxynitrite induces covalent dimerization of epidermal growth factor receptors in A431 epidermoid carcinoma cells. J. Biol. Chem. 1998, 273, 31860–31866. [Google Scholar] [CrossRef] [Green Version]
- Souza, J.M.; Peluffo, G.; Radi, R. Protein tyrosine nitration—Functional alteration or just a biomarker. Free. Radic. Biol. Med. 2008, 45, 357–366. [Google Scholar] [CrossRef]
- Cobbs, C.S.; Whisenhunt, T.R.; Wesemann, D.R.; Harkins, L.E.; Van Meir, E.G.; Samanta, M. Inactivation of wild-type p53 protein function by reactive oxygen and nitrogen species in malignant glioma cells. Cancer Res. 2003, 63, 8670–8673. [Google Scholar]
- Li, X.; De Sarno, P.; Song, L.; Beckman, J.S.; Jope, R.S. Peroxynitrite modulates tyrosine phosphorylation and phosphoinositide signalling in human neuroblastoma SH-SY5Y cells: Attenuated effects in human 1321N1 astrocytoma cells. Biochem. J. 1998, 331, 599–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greilberger, J. Preclinical and clinical testing of a alpha-ketoglutarate/5-HMF/N-actyl-selenomethionine and N-acteyl-methionine for treating tumors. J. Cancer Sci. Ther. 2013, 5, 76. [Google Scholar]
- Herwig, R.; Horninger, W.; Pinggera, G.M.; Rehder, P.; Frauscher, F.; Konwalinka, G.; Bartsch, G. Metabolomics therapy with 2-oxo-glutaric acid solution (Karal solution) in patients with hormone and chemotherapy insensitive metastatic prostate cancer leads to an increase of PSA doubling time and decrease of blood supply in tumour lesions. Eur. Urol. Suppl. 2005, 4, 146. [Google Scholar] [CrossRef]
- Tennant, D.A.; Gottlieb, E. HIF prolyl hydroxylase-3 mediates alpha-ketoglutarate-induced apoptosis and tumor suppression. J. Mol. Med. 2010, 88, 839–849. [Google Scholar] [CrossRef] [PubMed]
- Herwig, R.; Greilberger, J. Medication against male infertility (Arzneimittel gegen männliche Sterilität). HG Pharma. Austria 2016, 3, 1–8. [Google Scholar]
- Mariacher, C.; Gatterer, H.; Greilberger, J.; Djukic, R.; Greilberger, M.; Philippe, M.; Burtscher, M. Effects of antioxidant supplementation on exercise performance in acute normobaric hypoxia. Int. J. Sport Nutr. Exerc. Metab. 2014, 24, 227–235. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greilberger, J.; Greilberger, M.; Wintersteiger, R.; Zangger, K.; Herwig, R. Alpha-Ketoglutarate: A Potential Inner Mitochondrial and Cytosolic Protector against Peroxynitrite and Peroxynitrite-Induced Nitration? Antioxidants 2021, 10, 1501. https://doi.org/10.3390/antiox10091501
Greilberger J, Greilberger M, Wintersteiger R, Zangger K, Herwig R. Alpha-Ketoglutarate: A Potential Inner Mitochondrial and Cytosolic Protector against Peroxynitrite and Peroxynitrite-Induced Nitration? Antioxidants. 2021; 10(9):1501. https://doi.org/10.3390/antiox10091501
Chicago/Turabian StyleGreilberger, Joachim, Michaela Greilberger, Reinhold Wintersteiger, Klaus Zangger, and Ralf Herwig. 2021. "Alpha-Ketoglutarate: A Potential Inner Mitochondrial and Cytosolic Protector against Peroxynitrite and Peroxynitrite-Induced Nitration?" Antioxidants 10, no. 9: 1501. https://doi.org/10.3390/antiox10091501