
High Therapeutic and Esthetic Properties of Extracellular Vesicles Produced from the Stem Cells and Their Spheroids Cultured from Ocular Surgery-Derived Waste Orbicularis Oculi Muscle Tissues

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
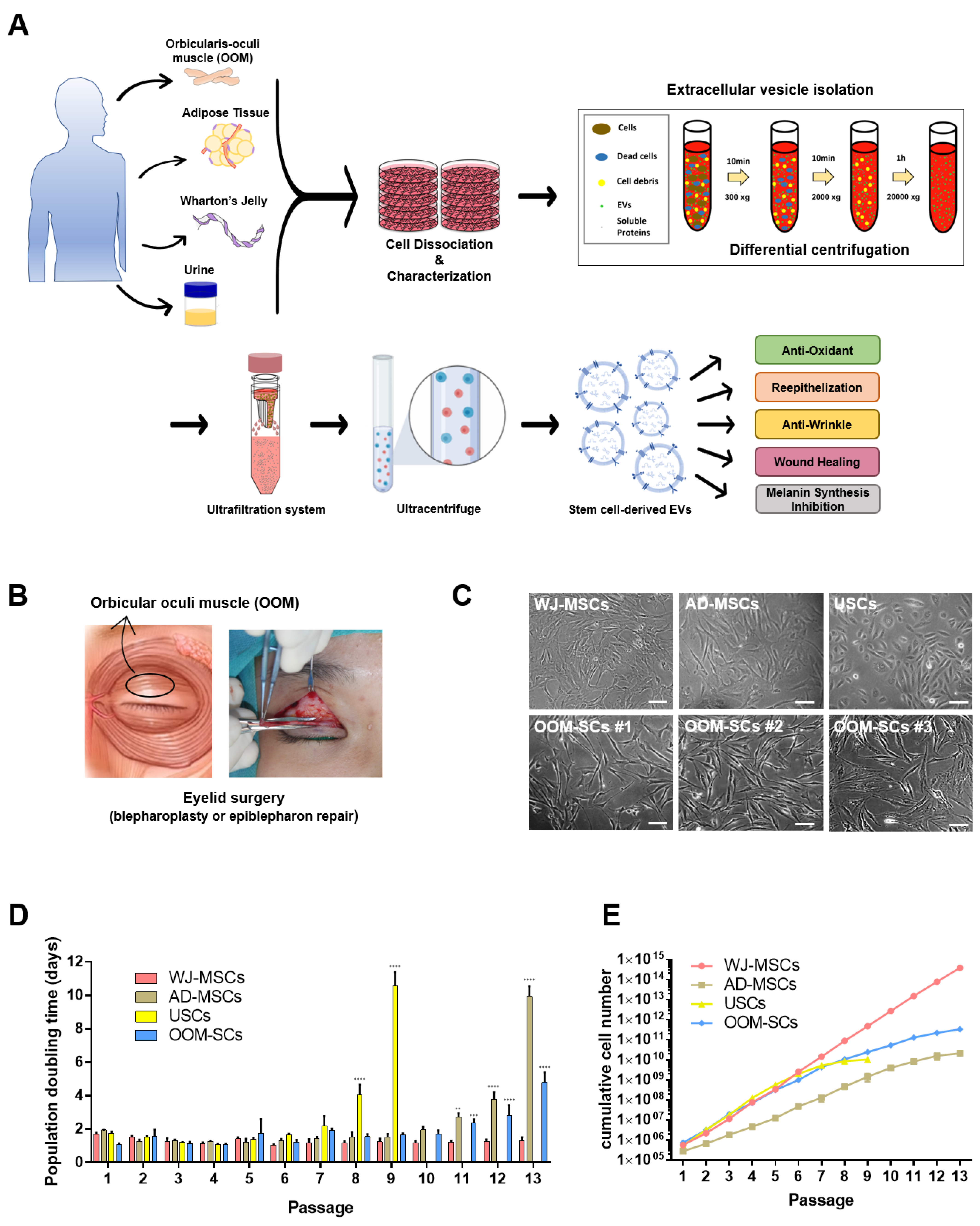
2. Materials and Methods
2.1. Preparation and Culture of OOM-SCs, WJ-MSCs, AD-MSCs, and USCs
2.2. Cell Growth Kinetics, Viability and Colony Formation Assays
2.3. Reverse Transcription Polymerase Chain Reaction (RT-PCR) and Quantitiative PCR (qPCR) Analyses
2.4. Flow Cytometry
2.5. Trilineage Differentiation
2.6. OOM-SC-EV and WJ-MSC-EV Isolation and Characterization
2.7. Western Blotting Assay
2.8. Three-Dimensional (3D) Spheroids Culture and EVs Isolation
2.9. Senescence-Associated β-Galactosidase (SA-βgal) Assay
2.10. Reactive Oxygen Species (ROS) Generation Measurement
2.11. Melanin Content Assay
2.12. Tyrosinase Activity Assay
2.13. In Vitro Cell Migration (Scratch Assay)
2.14. In Vivo Wound Healing Assay
2.15. Statistical Analysis
3. Results
3.1. OOM-SCs Characterization
3.2. OOM-SC-EVs and WJ-MSC-EVs Characterization
3.3. Advanced 3D Culture Approach for Efficient EVs Production
3.4. OOM-SC-EV Treatment Modulated ROS Generation and DELAYED Senescence-Related Changes in NHDFs
3.5. OOM-SC-EV Treatment Modulated the Melanin Synthesis of B16F10 Melanoma Cells
3.6. In Vitro and In Vivo Wound Healing and Anti-Wrinkle Capacities of OOM-SC-EVs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Scuderi, N.; Rubino, C. Island chondro-mucosal flap and skin graft: A new technique in eyelid reconstruction. Br. J. Plast. Surg. 1994, 47, 57–59. [Google Scholar] [CrossRef]
- Zhou, G. Clinical Experience with Orbicularis Oculi Myocutaneous Flaps in the Temporal Area. Plast. Reconstr. Surg. 2003, 112, 1862. [Google Scholar] [CrossRef]
- Yoshimura, Y.; Nakajima, T.; Yoneda, K. Reconstruction of the entire upper eyelid area with a subcutaneous pedicle flap based on the orbicularis oculi muscle. Plast. Reconstr. Surg. 1991, 88, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Kostakoǧlu, N.; Özcan, G. Orbicularis oculi myocutaneous flap in reconstruction of postburn lower eyelid ectropion. Burns 1999, 25, 553–557. [Google Scholar] [CrossRef]
- Mawrie, D.; Kumar, A.; Magdalene, D.; Bhattacharyya, J.; Jaganathan, B.G. Mesenchymal stem cells from human extra ocular muscle harbor neuroectodermal differentiation potential. PloS ONE 2016, 11, e0156697. [Google Scholar] [CrossRef] [PubMed]
- Polisetty, N.; Fatima, A.; Madhira, S.L.; Sangwan, V.S.; Vemuganti, G.K. Mesenchymal cells from limbal stroma of human eye. Mol. Vis. 2008, 14, 431. [Google Scholar]
- Formicola, L.; Marazzi, G.; Sassoon, D.A. The extraocular muscle stem cell niche is resistant to ageing and disease. Front. Aging Neurosci. 2014, 6, 328. [Google Scholar] [CrossRef]
- Pacheco-Pinedo, E.C.; Budak, M.T.; Zeiger, U.; Jørgensen, L.H.; Bogdanovich, S.; Schrøder, H.D.; Rubinstein, N.A.; Khurana, T.S. Transcriptional and functional differences in stem cell populations isolated from extraocular and limb muscles. Physiol. Genom. 2009, 37, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, Y.; Takenaka, N.; Sakurai, H.; Ueno, M.; Kinoshita, S.; Sotozono, C.; Sato, T. Human Skeletal Muscle Cells Derived from the Orbicularis Oculi Have Regenerative Capacity for Duchenne Muscular Dystrophy. Int. J. Mol. Sci. 2019, 20, 3456. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Liao, C.; Chen, X.; Xu, Y.; Tan, J.; Han, F.; Ye, X. Identification and Characterization of Skeletal Muscle Stem Cells from Human Orbicularis Oculi Muscle. Tissue Eng. Part C Methods 2018, 24, 486–493. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 1–18. [Google Scholar] [CrossRef]
- Gowen, A.; Shahjin, F.; Chand, S.; Odegaard, K.E.; Yelamanchili, S.V. Mesenchymal stem cell-derived extracellular vesicles: Challenges in clinical applications. Front. Cell Dev. Biol. 2020, 8, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.-J.; Fang, Q.-H.; Liu, M.-L.; Lin, J.-N. Current understanding of the role of Adipose-derived Extracellular Vesicles in Metabolic Homeostasis and Diseases: Communication from the distance between cells/tissues. Theranostics 2020, 10, 7422. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.; Fan, J.; Lyon, C.; Wan, M.; Hu, Y. Role of extracellular vesicles in viral and bacterial infections: Pathogenesis, diagnostics, and therapeutics. Theranostics 2018, 8, 2709. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.-H.; Kim, J.-H. A comprehensive review on factors influences biogenesis, functions, therapeutic and clinical implications of exosomes. Int. J. Nanomed. 2021, 16, 1281. [Google Scholar] [CrossRef] [PubMed]
- Nassar, W.; El-Ansary, M.; Sabry, D.; Mostafa, M.A.; Fayad, T.; Kotb, E.; Temraz, M.; Saad, A.-N.; Essa, W.; Adel, H. Umbilical cord mesenchymal stem cells derived extracellular vesicles can safely ameliorate the progression of chronic kidney diseases. Biomater. Res. 2016, 20, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kordelas, L.; Rebmann, V.; Ludwig, A.; Radtke, S.; Ruesing, J.; Doeppner, T.; Epple, M.; Horn, P.; Beelen, D.; Giebel, B. MSC-derived exosomes: A novel tool to treat therapy-refractory graft-versus-host disease. Leukemia 2014, 28, 970–973. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Shi, H.; Yin, S.; Ji, C.; Zhang, X.; Zhang, B.; Wu, P.; Shi, Y.; Mao, F.; Yan, Y. Human mesenchymal stem cell derived exosomes alleviate type 2 diabetes mellitus by reversing peripheral insulin resistance and relieving β-cell destruction. ACS Nano 2018, 12, 7613–7628. [Google Scholar] [CrossRef]
- Zhao, M.; Shi, J.; Cai, W.; Liu, K.; Shen, K.; Li, Z.; Wang, Y.; Hu, D. Advances on Graphene-Based Nanomaterials and Mesenchymal Stem Cell-Derived Exosomes Applied in Cutaneous Wound Healing. Int. J. Nanomed. 2021, 16, 2647. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Sun, X.; Liu, L.; Jiang, H.; Shen, Y.; Xu, X.; Li, J.; Zhang, G.; Huang, J.; Lin, Z. Exosomes and their therapeutic potentials of stem cells. Stem Cells Int. 2016, 2016, 7653489. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, F.G.; Carvalho, M.M.; Sousa, N.; Salgado, A.J. Mesenchymal stem cells secretome: A new paradigm for central nervous system regeneration? Cell. Mol. Life Sci. 2013, 70, 3871–3882. [Google Scholar] [CrossRef] [PubMed]
- Keshtkar, S.; Azarpira, N.; Ghahremani, M.H. Mesenchymal stem cell-derived extracellular vesicles: Novel frontiers in regenerative medicine. Stem Cell Res. Ther. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Chen, Z.; Pan, D.; Li, H.; Shen, J. Umbilical cord-derived mesenchymal stem cell-derived exosomes combined pluronic F127 hydrogel promote chronic diabetic wound healing and complete skin regeneration. Int. J. Nanomed. 2020, 15, 5911. [Google Scholar] [CrossRef] [PubMed]
- Basu, J.; Ludlow, J.W. Exosomes for repair, regeneration and rejuvenation. Expert Opin. Biol. Ther. 2016, 16, 489–506. [Google Scholar] [CrossRef]
- Phinney, D.G.; Pittenger, M.F. Concise review: MSC-derived exosomes for cell-free therapy. Stem Cells 2017, 35, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, P.; Larijani, B.; Alavi-Moghadam, S.; Tayanloo-Beik, A.; Mohamadi-Jahani, F.; Ranjbaran, N.; Payab, M.; Falahzadeh, K.; Mousavi, M.; Arjmand, B. Mesenchymal stem cells-derived exosomes for wound regeneration. Cell Biol. Transl. Med. 2018, 4, 119–131. [Google Scholar]
- Zhao, B.; Zhang, Y.; Han, S.; Zhang, W.; Zhou, Q.; Guan, H.; Liu, J.; Shi, J.; Su, L.; Hu, D. Exosomes derived from human amniotic epithelial cells accelerate wound healing and inhibit scar formation. J. Mol. Histol. 2017, 48, 121–132. [Google Scholar] [CrossRef]
- Hu, P.; Yang, Q.; Wang, Q.; Shi, C.; Wang, D.; Armato, U.; Prà, I.D.; Chiarini, A. Mesenchymal stromal cells-exosomes: A promising cell-free therapeutic tool for wound healing and cutaneous regeneration. Burn. Trauma 2019, 7. [Google Scholar] [CrossRef]
- An, T.; Chen, Y.; Tu, Y.; Lin, P. Mesenchymal stromal cell-derived extracellular vesicles in the treatment of diabetic foot ulcers: Application and challenges. Stem Cell Rev. Rep. 2021, 17, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Liu, W.; Li, J.; Lu, J.; Lu, H.; Jia, W.; Liu, F. Exosomes derived from atorvastatin-pretreated MSC accelerate diabetic wound repair by enhancing angiogenesis via AKT/eNOS pathway. Stem Cell Res. Ther. 2020, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lai, P.; Chen, X.; Guo, L.; Wang, Y.; Liu, X.; Liu, Y.; Zhou, T.; Huang, T.; Geng, S.; Luo, C. A potent immunomodulatory role of exosomes derived from mesenchymal stromal cells in preventing cGVHD. J. Hematol. Oncol. 2018, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Quiñones-Vico, M.I.; Sanabria-de la Torre, R.; Sánchez-Díaz, M.; Sierra-Sánchez, Á.; Montero-Vílchez, T.; Fernández-González, A.; Arias-Santiago, S. The Role of Exosomes Derived From Mesenchymal Stromal Cells in Dermatology. Front. Cell Dev. Biol. 2021, 9, 647012. [Google Scholar] [CrossRef]
- Théry, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- György, B.; Hung, M.E.; Breakefield, X.O.; Leonard, J.N. Therapeutic applications of extracellular vesicles: Clinical promise and open questions. Annu. Rev. Pharmacol. Toxicol. 2015, 55, 439–464. [Google Scholar] [CrossRef] [Green Version]
- Henriques-Antunes, H.; Cardoso, R.M.; Zonari, A.; Correia, J.; Leal, E.C.; Jimenez-Balsa, A.; Lino, M.M.; Barradas, A.; Kostic, I.; Gomes, C. The kinetics of small extracellular vesicle delivery impacts skin tissue regeneration. ACS Nano 2019, 13, 8694–8707. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, M.; Gong, A.; Zhang, X.; Wu, X.; Zhu, Y.; Shi, H.; Wu, L.; Zhu, W.; Qian, H. HucMSC-exosome mediated-Wnt4 signaling is required for cutaneous wound healing. Stem Cells 2015, 33, 2158–2168. [Google Scholar] [CrossRef]
- Sicco, C.L.; Reverberi, D.; Balbi, C.; Ulivi, V.; Principi, E.; Pascucci, L.; Becherini, P.; Bosco, M.C.; Varesio, L.; Franzin, C. Mesenchymal stem cell-derived extracellular vesicles as mediators of anti-inflammatory effects: Endorsement of macrophage polarization. Stem Cells Transl. Med. 2017, 6, 1018–1028. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, D.; Das, A.; Roy, S. Wound Healing, Tissue Repair and Regeneration in Diabetes; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Campos, A.M.; Maciel, E.; Moreira, A.S.; Sousa, B.; Melo, T.; Domingues, P.; Curado, L.; Antunes, B.; Domingues, M.R.M.; Santos, F. Lipidomics of Mesenchymal Stromal Cells: Understanding the Adaptation of Phospholipid Profile in Response to Pro-Inflammatory Cytokines. J. Cell. Physiol. 2016, 231, 1024–1032. [Google Scholar] [CrossRef]
- Bjørge, I.; Kim, S.; Mano, J.; Kalionis, B.; Chrzanowski, W. Extracellular vesicles, exosomes and shedding vesicles in regenerative medicine–a new paradigm for tissue repair. Biomater. Sci. 2018, 6, 60–78. [Google Scholar] [CrossRef]
- Silva, A.M.; Teixeira, J.H.; Almeida, M.I.; Goncalves, R.M.; Barbosa, M.A.; Santos, S.G. Extracellular vesicles: Immunomodulatory messengers in the context of tissue repair/regeneration. Eur. J. Pharm. Sci. 2017, 98, 86–95. [Google Scholar] [CrossRef]
- Carrasco, E.; Soto-Heredero, G.; Mittelbrunn, M. The role of extracellular vesicles in cutaneous remodeling and hair follicle dynamics. Int. J. Mol. Sci. 2019, 20, 2758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucharzewski, M.; Rojczyk, E.; Wilemska-Kucharzewska, K.; Wilk, R.; Hudecki, J.; Los, M.J. Novel trends in application of stem cells in skin wound healing. Eur. J. Pharmacol. 2019, 843, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yan, Z.; Yang, F.; Huang, Y.; Yu, Y.; Zhou, L.; Sun, Z.; Cui, D.; Yan, Y. Exosomes derived from human umbilical cord mesenchymal stem cells accelerate cutaneous wound healing by enhancing angiogenesis through delivering angiopoietin-2. Stem Cell Rev. Rep. 2021, 17, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Gil, M.; Dayem, A.A.; Choi, S.; Kang, G.-H.; Yang, G.-M.; Cho, S.; Jeong, Y.; Kim, S.J.; Seok, J. Improved isolation and culture of urine-derived stem cells (USCs) and enhanced production of immune cells from the USC-derived induced pluripotent stem cells. J. Clin. Med. 2020, 9, 827. [Google Scholar] [CrossRef] [Green Version]
- Bruckova, L.; Soukup, T.; Visek, B.; Moos, J.; Moosova, M.; Pavelkova, J.; Rezabek, K.; Kucerova, L.; Micuda, S.; Brcakova, E. Proliferative potential and phenotypic analysis of long-term cultivated human granulosa cells initiated by addition of follicular fluid. J. Assist. Reprod. Genet. 2011, 28, 939–950. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Shelke, G.V.; Lässer, C.; Gho, Y.S.; Lötvall, J. Importance of exosome depletion protocols to eliminate functional and RNA-containing extracellular vesicles from fetal bovine serum. J. Extracell. Vesicles 2014, 3, 24783. [Google Scholar] [CrossRef] [Green Version]
- Debacq-Chainiaux, F.; Erusalimsky, J.D.; Campisi, J.; Toussaint, O. Protocols to detect senescence-associated beta-galactosidase (SA-βgal) activity, a biomarker of senescent cells in culture and in vivo. Nat. Protoc. 2009, 4, 1798. [Google Scholar] [CrossRef]
- Gebäck, T.; Schulz, M.M.P.; Koumoutsakos, P.; Detmar, M. TScratch: A novel and simple software tool for automated analysis of monolayer wound healing assays: Short Technical Reports. Biotechniques 2009, 46, 265–274. [Google Scholar] [CrossRef]
- Benavides, F.; Oberyszyn, T.M.; VanBuskirk, A.M.; Reeve, V.E.; Kusewitt, D.F. The hairless mouse in skin research. J. Dermatol. Sci. 2009, 53, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Liao, C.; Chen, X.; Wu, Y. In Vitro and In Vivo Osteogenesis of Human Orbicularis Oculi Muscle-Derived Stem Cells. Tissue Eng. Regen. Med. 2018, 15, 445–452. [Google Scholar] [CrossRef]
- Haniffa, M.A.; Collin, M.P.; Buckley, C.D.; Dazzi, F. Mesenchymal stem cells: The fibroblasts’ new clothes? Haematologica 2009, 94, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Chang, W.; Wei, H.; Zhang, K. Comparison of the biological characteristics of mesenchymal stem cells derived from bone marrow and skin. Stem Cells Int. 2016, 2016, 3658798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaculik, C.; Schuster, C.; Bauer, W.; Iram, N.; Pfisterer, K.; Kramer, G.; Reinisch, A.; Strunk, D.; Elbe-Bürger, A. Human dermis harbors distinct mesenchymal stromal cell subsets. J. Investig. Dermatol. 2012, 132, 563–574. [Google Scholar] [CrossRef] [Green Version]
- Bae, S.; Shim, S.H.; Park, C.W.; Son, H.K.; Ju Lee, H.; Son, J.Y.; Jeon, C.; Kim, H. Combined omics analysis identifies transmembrane 4 L6 family member 1 as a surface protein marker specific to human mesenchymal stem cells. Stem Cells Dev. 2011, 20, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Lv, F.-J.; Tuan, R.S.; Cheung, K.M.; Leung, V.Y. Concise review: The surface markers and identity of human mesenchymal stem cells. Stem Cells 2014, 32, 1408–1419. [Google Scholar] [CrossRef] [PubMed]
- Battula, V.L.; Treml, S.; Bareiss, P.M.; Gieseke, F.; Roelofs, H.; De Zwart, P.; Müller, I.; Schewe, B.; Skutella, T.; Fibbe, W.E. Isolation of functionally distinct mesenchymal stem cell subsets using antibodies against CD56, CD271, and mesenchymal stem cell antigen-1. Haematologica 2009, 94, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Holley, R.J.; Tai, G.; Williamson, A.J.; Taylor, S.; Cain, S.A.; Richardson, S.M.; Merry, C.L.; Whetton, A.D.; Kielty, C.M.; Canfield, A.E. Comparative quantification of the surfaceome of human multipotent mesenchymal progenitor cells. Stem Cell Rep. 2015, 4, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Hu, L.; Zhou, X.; Xiong, Z.; Zhang, C.; Shehada, H.M.; Hu, B.; Song, J.; Chen, L. Exosomes secreted by human adipose mesenchymal stem cells promote scarless cutaneous repair by regulating extracellular matrix remodelling. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Costin, B.R.; Plesec, T.P.; Kopplin, L.J.; Chundury, R.V.; McBride, J.M.; Levine, M.R.; Perry, J.D. Regional Variations in Orbicularis Oculi Histology. Ophthalmic Plast. Reconstr. Surg. 2015, 31, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Crapnell, K.; Blaesius, R.; Hastings, A.; Lennon, D.P.; Caplan, A.I.; Bruder, S.P. Growth, differentiation capacity, and function of mesenchymal stem cells expanded in serum-free medium developed via combinatorial screening. Exp. Cell Res. 2013, 319, 1409–1418. [Google Scholar] [CrossRef]
- Katsuda, T.; Kosaka, N.; Takeshita, F.; Ochiya, T. The therapeutic potential of mesenchymal stem cell-derived extracellular vesicles. Proteomics 2013, 13, 1637–1653. [Google Scholar] [CrossRef]
- Park, K.-S.; Bandeira, E.; Shelke, G.V.; Lässer, C.; Lötvall, J. Enhancement of therapeutic potential of mesenchymal stem cell-derived extracellular vesicles. Stem Cell Res. Ther. 2019, 10, 288. [Google Scholar] [CrossRef]
- Huang, Y.-C.; Lai, L.-C. The potential roles of stem cell-derived extracellular vesicles as a therapeutic tool. Ann. Transl. Med. 2019, 7, 693. [Google Scholar] [CrossRef]
- Tang, T.-T.; Wang, B.; Lv, L.-L.; Liu, B.-C. Extracellular vesicle-based Nanotherapeutics: Emerging frontiers in anti-inflammatory therapy. Theranostics 2020, 10, 8111. [Google Scholar] [CrossRef]
- Cao, G.; Chen, B.; Zhang, X.; Chen, H. Human Adipose-Derived Mesenchymal Stem Cells-Derived Exosomal microRNA-19b Promotes the Healing of Skin Wounds Through Modulation of the CCL1/TGF-β Signaling Axis. Clin. Cosmet. Investig. Dermatol. 2020, 13, 957–971. [Google Scholar] [CrossRef]
- Hao, Y.; Miao, J.; Liu, W.; Cai, K.; Huang, X.; Peng, L. Mesenchymal Stem Cell-Derived Exosomes Carry MicroRNA-125a to Protect Against Diabetic Nephropathy by Targeting Histone Deacetylase 1 and Downregulating Endothelin-1. Diabetes Metab. Syndr. Obes. Targets Ther. 2021, 14, 1405. [Google Scholar] [CrossRef]
- Vlassov, A.V.; Magdaleno, S.; Setterquist, R.; Conrad, R. Exosomes: Current knowledge of their composition, biological functions, and diagnostic and therapeutic potentials. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2012, 1820, 940–948. [Google Scholar] [CrossRef]
- Ramirez, M.I.; Amorim, M.G.; Gadelha, C.; Milic, I.; Welsh, J.A.; Freitas, V.M.; Nawaz, M.; Akbar, N.; Couch, Y.; Makin, L. Technical challenges of working with extracellular vesicles. Nanoscale 2018, 10, 881–906. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak, J.; Miekus, K.; Kucia, M.; Zhang, J.; Reca, R.; Dvorak, P.; Ratajczak, M. Embryonic stem cell-derived microvesicles reprogram hematopoietic progenitors: Evidence for horizontal transfer of mRNA and protein delivery. Leukemia 2006, 20, 847–856. [Google Scholar] [CrossRef] [Green Version]
- Luga, V.; Zhang, L.; Viloria-Petit, A.M.; Ogunjimi, A.A.; Inanlou, M.R.; Chiu, E.; Buchanan, M.; Hosein, A.N.; Basik, M.; Wrana, J.L. Exosomes mediate stromal mobilization of autocrine Wnt-PCP signaling in breast cancer cell migration. Cell 2012, 151, 1542–1556. [Google Scholar] [CrossRef] [Green Version]
- Gross, J.C.; Chaudhary, V.; Bartscherer, K.; Boutros, M. Active Wnt proteins are secreted on exosomes. Nat. Cell Biol. 2012, 14, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Huang, W.; Meng, W.; Jegga, A.G.; Wang, Y.; Cai, W.; Kim, H.W.; Pasha, Z.; Wen, Z.; Rao, F. Heat shock improves Sca-1+ stem cell survival and directs ischemic cardiomyocytes toward a prosurvival phenotype via exosomal transfer: A critical role for HSF1/miR-34a/HSP70 pathway. Stem Cells 2014, 32, 462–472. [Google Scholar] [CrossRef] [Green Version]
- Ong, S.-G.; Lee, W.H.; Huang, M.; Dey, D.; Kodo, K.; Sanchez-Freire, V.; Gold, J.D.; Wu, J.C. Cross talk of combined gene and cell therapy in ischemic heart disease: Role of exosomal microRNA transfer. Circulation 2014, 130, S60–S69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Ge, X.; Shi, Z.; Yu, C.; Lu, C.; Wei, Y.; Zeng, A.; Wang, X.; Yan, W.; Zhang, J. Extracellular vesicles derived from hypoxic glioma stem-like cells confer temozolomide resistance on glioblastoma by delivering miR-30b-3p. Theranostics 2021, 11, 1763. [Google Scholar] [CrossRef] [PubMed]
- Prattichizzo, F.; Matacchione, G.; Giuliani, A.; Sabbatinelli, J.; Olivieri, F.; de Candia, P.; De Nigris, V.; Ceriello, A. Extracellular vesicle-shuttled miRNAs: A critical appraisal of their potential as nano-diagnostics and nano-therapeutics in type 2 diabetes mellitus and its cardiovascular complications. Theranostics 2021, 11, 1031. [Google Scholar] [CrossRef]
- Xia, B.; Gao, J.; Li, S.; Huang, L.; Zhu, L.; Ma, T.; Zhao, L.; Yang, Y.; Luo, K.; Shi, X. Mechanical stimulation of Schwann cells promote peripheral nerve regeneration via extracellular vesicle-mediated transfer of microRNA 23b-3p. Theranostics 2020, 10, 8974. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Li, L.; Fang, X.; Zang, M. Exosome-Encapsulated microRNA-127-3p Released from Bone Marrow-Derived Mesenchymal Stem Cells Alleviates Osteoarthritis Through Regulating CDH11-Mediated Wnt/β-Catenin Pathway. J. Pain Res. 2021, 14, 297. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhu, Y.; Dong, X.; Cao, G.; Li, Y.; Fan, Y.; Chen, Q.; Cai, H.; Wu, Y. Exosomes Derived from Pancreatic Cancer Cells Induce Osteoclast Differentiation Through the miR125a-5p/TNFRSF1B Pathway. OncoTargets Ther. 2021, 14, 2727. [Google Scholar] [CrossRef]
- Tran, P.H.; Xiang, D.; Nguyen, T.N.; Tran, T.T.; Chen, Q.; Yin, W.; Zhang, Y.; Kong, L.; Duan, A.; Chen, K. Aptamer-guided extracellular vesicle theranostics in oncology. Theranostics 2020, 10, 3849. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Duan, L.; Lu, J.; Xia, J. Engineering exosomes for targeted drug delivery. Theranostics 2021, 11, 3183. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.; Busatto, S.; Pham, A.; Tian, M.; Suh, A.; Carson, K.; Quintero, A.; Lafrence, M.; Malik, H.; Santana, M.X. Extracellular vesicle-based drug delivery systems for cancer treatment. Theranostics 2019, 9, 8001. [Google Scholar] [CrossRef]
- Zhang, K.; Dong, C.; Chen, M.; Yang, T.; Wang, X.; Gao, Y.; Wang, L.; Wen, Y.; Chen, G.; Wang, X. Extracellular vesicle-mediated delivery of miR-101 inhibits lung metastasis in osteosarcoma. Theranostics 2020, 10, 411. [Google Scholar] [CrossRef]
- Chen, P.; Wang, L.; Fan, X.; Ning, X.; Yu, B.; Ou, C.; Chen, M. Targeted delivery of extracellular vesicles in heart injury. Theranostics 2021, 11, 2263. [Google Scholar] [CrossRef]
- Villa, A.; Garofalo, M.; Crescenti, D.; Rizzi, N.; Brunialti, E.; Vingiani, A.; Belotti, P.; Sposito, C.; Franzè, S.; Cilurzo, F. Transplantation of autologous extracellular vesicles for cancer-specific targeting. Theranostics 2021, 11, 2034. [Google Scholar] [CrossRef]
- Mousavinejad, M.; Andrews, P.W.; Shoraki, E.K. Current biosafety considerations in stem cell therapy. Cell J. 2016, 18, 281. [Google Scholar]
- Kishore, R.; Khan, M. More than tiny sacks: Stem cell exosomes as cell-free modality for cardiac repair. Circ. Res. 2016, 118, 330–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Zhao, L.; Wei, Y.; Ea, V.L.; Nian, H.; Wei, R. Recent advances of exosomes in immune-mediated eye diseases. Stem Cell Res. Ther. 2019, 10, 278. [Google Scholar] [CrossRef] [Green Version]
- Klingeborn, M.; Dismuke, W.M.; Rickman, C.B.; Stamer, W.D. Roles of exosomes in the normal and diseased eye. Prog. Retin. Eye Res. 2017, 59, 158–177. [Google Scholar] [CrossRef]
- Gonzalez, E.; Falcón-Pérez, J.M. Cell-derived extracellular vesicles as a platform to identify low-invasive disease biomarkers. Expert Rev. Mol. Diagn. 2015, 15, 907–923. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, Y.; Kong, Y. Exosomes derived from mesenchymal stem cells modulate miR-126 to ameliorate hyperglycemia-induced retinal inflammation via Targeting HMGB1. Investig. Ophthalmol. Vis. Sci. 2019, 60, 294–303. [Google Scholar] [CrossRef] [Green Version]
- Moisseiev, E.; Anderson, J.D.; Oltjen, S.; Goswami, M.; Zawadzki, R.J.; Nolta, J.A.; Park, S.S. Protective effect of intravitreal administration of exosomes derived from mesenchymal stem cells on retinal ischemia. Curr. Eye Res. 2017, 42, 1358–1367. [Google Scholar] [CrossRef]
- Samaeekia, R.; Rabiee, B.; Putra, I.; Shen, X.; Park, Y.J.; Hematti, P.; Eslani, M.; Djalilian, A.R. Effect of human corneal mesenchymal stromal cell-derived exosomes on corneal epithelial wound healing. Investig. Ophthalmol. Vis. Sci. 2018, 59, 5194–5200. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-J.; mi Yoo, S.; Park, H.H.; Lim, H.J.; Kim, Y.-L.; Lee, S.; Seo, K.-W.; Kang, K.-S. Exosomes derived from human umbilical cord blood mesenchymal stem cells stimulates rejuvenation of human skin. Biochem. Biophys. Res. Commun. 2017, 493, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.; Lee, J.; Kim, Y.J.; Rhee, W.J.; Park, J.H. Exosomes Derived from Human Induced Pluripotent Stem Cells Ameliorate the Aging of Skin Fibroblasts. Int. J. Mol. Sci. 2018, 19, 1715. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Wang, J.; Zhou, X.; Xiong, Z.; Zhao, J.; Yu, R.; Huang, F.; Zhang, H.; Chen, L. Exosomes derived from human adipose mensenchymal stem cells accelerates cutaneous wound healing via optimizing the characteristics of fibroblasts. Sci. Rep. 2016, 6, 32993. [Google Scholar] [CrossRef] [PubMed]
- Gallet, R.; Dawkins, J.; Valle, J.; Simsolo, E.; De Couto, G.; Middleton, R.; Tseliou, E.; Luthringer, D.; Kreke, M.; Smith, R.R. Exosomes secreted by cardiosphere-derived cells reduce scarring, attenuate adverse remodelling, and improve function in acute and chronic porcine myocardial infarction. Eur. Heart J. 2017, 38, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Bodega, G.; Alique, M.; Puebla, L.; Carracedo, J.; Ramírez, R.M. Microvesicles: ROS scavengers and ROS producers. J. Extracell. Vesicles 2019, 8, 1626654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waghule, T.; Singhvi, G.; Dubey, S.K.; Pandey, M.M.; Gupta, G.; Singh, M.; Dua, K. Microneedles: A smart approach and increasing potential for transdermal drug delivery system. Biomed. Pharmacother. 2019, 109, 1249–1258. [Google Scholar] [CrossRef] [PubMed]
- Hearing, V.J. Biogenesis of pigment granules: A sensitive way to regulate melanocyte function. J. Dermatol. Sci. 2005, 37, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Briganti, S.; Camera, E.; Picardo, M. Chemical and instrumental approaches to treat hyperpigmentation. Pigment. Cell Res. 2003, 16, 101–110. [Google Scholar] [CrossRef]
- Yasui, H.; Sakurai, H. Age-dependent generation of reactive oxygen species in the skin of live hairless rats exposed to UVA light. Exp. Dermatol. 2003, 12, 655–661. [Google Scholar] [CrossRef]
- Yamakoshi, J.; Otsuka, F.; Sano, A.; Tokutake, S.; Saito, M.; Kikuchi, M.; Kubota, Y. Lightening effect on ultraviolet-induced pigmentation of guinea pig skin by oral administration of a proanthocyanidin-rich extract from grape seeds. Pigment. Cell Res. 2003, 16, 629–638. [Google Scholar] [CrossRef]
- Kao, Y.-Y.; Chuang, T.-F.; Chao, S.-H.; Yang, J.-H.; Lin, Y.-C.; Huang, H.-Y. Evaluation of the antioxidant and melanogenesis inhibitory properties of pracparatum mungo (lu-do huang). J. Tradit. Complementary Med. 2013, 3, 163–170. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Application) | NCBI Accession Number | Forward Primer (5′ to 3′) Reverse Primer (5′ to 3′) | Product Length (bp) |
|---|---|---|---|
| NANOG (Semi-qPCR) | NM_001355281.2 | TCCTGAACCTCAGCTACAAAC GCGTCACACCATTGCTATTC | 108 |
| SOX2 (Semi-qPCR) | NM_003106.4 | CATCACCCACAGCAAATGAC GAAGTCCAGGATCTCTCTCATAAA | 110 |
| REX1 (Semi-qPCR) | NM_001304358.2 | GTTTCGTGTGTCCCTTTCAAG CTGTTATCTGCTTCATCCTGTTG | 141 |
| ITGA6 (Semi-qPCR) | NM_001394928.1 | CGAAACCAAGGTTCTGAGCCCA CTTGGATCTCCACTGAGGCAGT | 151 |
| CD146 (Semi-qPCR) | NM_006500.3 | GTGTTGAATCTGTCTTGTGAA ATGCCTCAGATCGATG | 600, 419 |
| TM4SF1 (Semi-qPCR) | NM_014220.3 | GGCTACTGTGTCATTGTGGCAG ACTCGGACCATGTGGAGGTATC | 133 |
| CD19 (Semi-qPCR) | NM_001385732.1 | CCTGGGGTCCCAGTCCTATG GCTCCAGAGGTTGGCATCAT | 287 |
| CD45 (Semi-qPCR) | NM_080921.4 | TCAGTGGTCCCATTGTTGTG GCATCTCTGTGGCCTTAGCT | 146 |
| CD29 (Semi-qPCR) | NM_002211.4 | GCCGCGCGGAAAAGATG ACATCGTGCAGAAGTAGGCA | 206 |
| CD73 (Semi-qPCR) | NM_002526.4 | TATCCGGTCGCCCATTGATG ACGCTATGCTCAAAGGCCTT | 138 |
| CD105 (Semi-qPCR) | NM_001278138.2 | CCAAGACCGGGTCTCAAGAC TGTACCAGAGTGCAGCAGTG | 174 |
| CD166 (Semi-qPCR) | NM_001627.4 | GAACACGATGAGGCAGACGA CCGAGGTCCTTGTTTACATGTTT | 179 |
| GAPDH (Semi-qPCR) | NM_001357943.2 | AATCCCATCACCATCTTCCAG ATGACCCTTTTGGCTCCC | 146 |
| GPX1 (qPCR) | NM_001329503.2 | CAGTCGGTGTATGCCTTCTCG GAGGGACGCCACATTCTCG | 105 |
| GPX2 (qPCR) | NM_002083.4 | GGTAGATTTCAATACGTTCCGGG TGACAGTTCTCCTGATGTCCAAA | 174 |
| GPX3 (qPCR) | NM_001329790.2 | AGAGCCGGGGACAAGAGAA ATTTGCCAGCATACTGCTTGA | 153 |
| GPX4 (qPCR) | NM_001367832.1 | GAGGCAAGACCGAAGTAAACTAC CCGAACTGGTTACACGGGAA | 100 |
| GPX7 (qPCR) | NM_015696.5 | CCCACCACTTTAACGTGCTC GGCAAAGCTCTCAATCTCCTT | 86 |
| GSR (qPCR) | NM_001195102.3 | TTCCAGAATACCAACGTCAAAGG GTTTTCGGCCAGCAGCTATTG | 94 |
| SOD1 (qPCR) | NM_000454.5 | GGTGGGCCAAAGGATGAAGAG CCACAAGCCAAACGACTTCC | 227 |
| SOD2 (qPCR) | NM_001322816.2 | GCTCCGGTTTTGGGGTATCTG GCGTTGATGTGAGGTTCCAG | 92 |
| SOD3 (qPCR) | NM_003102.4 | ATGCTGGCGCTACTGTGTTC CTCCGCCGAGTCAGAGTTG | 99 |
| Catalase (qPCR) | NM_001752.4 | TGTTGCTGGAGAATCGGGTTC TCCCAGTTACCATCTTCTGTGTA | 87 |
| TMX1 (qPCR) | NM_030755.5 | AGTATGTCAGCACTCTTTCAGC CACACTGGCAATCCAAGGTCT | 83 |
| TXNIP (qPCR) | NM_006472.6 | GGTCTTTAACGACCCTGAAAAGG ACACGAGTAACTTCACACACCT | 87 |
| TXN (qPCR) | NM_001244938.2 | GTGAAGCAGATCGAGAGCAAG CGTGGCTGAGAAGTCAACTACTA | 87 |
| Col1A1 (qPCR) | NM_000088.4 | GATTCCCTGGACCTAAAGGTGC AGCCTCTCCATCTTTGCCAGCA | 107 |
| MCP-1 (qPCR) | NM_002982.4 | AGAATCACCAGCAGCAAGTGTCC TCCTGAACCCACTTCTGCTTGG | 98 |
| MCP-3 (qPCR) | NM_006273.4 | ACAGAAGGACCACCAGTAGCCA GGTGCTTCATAAAGTCCTGGACC | 117 |
| CCL5 (qPCR) | NM_002985.3 | CCTGCTGCTTTGCCTACATTGC ACACACTTGGCGGTTCTTTCGG | 125 |
| PAI-1 (qPCR) | NM_001386456.1 | CTCATCAGCCACTGGAAAGGCA GACTCGTGAAGTCAGCCTGAAAC | 154 |
| TGF-β1 (qPCR) | NM_000660.7 | TACCTGAACCCGTGTTGCTCTC GTTGCTGAGGTATCGCCAGGAA | 122 |
| TGF-β3 (qPCR) | NM_001329939.2 | CTAAGCGGAATGAGCAGAGGATC TCTCAACAGCCACTCACGCACA | 161 |
| GAPDH (qPCR) | NM_001357943.2 | GTCTCCTCTGACTTCAACAGCG ACCACCCTGTTGCTGTAGCCAA | 131 |
| WJ-MSCs | AD-MSCs | USCs | OOM-SCs | |
|---|---|---|---|---|
| Volume/Length | 14 cm | 60 cc | 100 mL | 2 cm |
| Cumulative Cell Number | 3.8 × 1014 | 2.1 × 1010 | 1.0 × 1010 | 3.3 × 1011 |
| Doubling time | 30.53 h | 42.34 h | 44.46 h | 39.58 h |
| WJ-MSCs | USCs | OOM-SCs | |
|---|---|---|---|
| Adipogenic | +++ | ++ | +++ |
| Chondrogenic | +++ | ++ | ++ |
| Osteogenic | +++ | ++ | +++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, K.M.; Dayem, A.A.; Choi, Y.; Lee, Y.; An, J.; Gil, M.; Lee, S.; Kwak, H.J.; Vellingirl, B.; Shin, H.J.; et al. High Therapeutic and Esthetic Properties of Extracellular Vesicles Produced from the Stem Cells and Their Spheroids Cultured from Ocular Surgery-Derived Waste Orbicularis Oculi Muscle Tissues. Antioxidants 2021, 10, 1292. https://doi.org/10.3390/antiox10081292
Lim KM, Dayem AA, Choi Y, Lee Y, An J, Gil M, Lee S, Kwak HJ, Vellingirl B, Shin HJ, et al. High Therapeutic and Esthetic Properties of Extracellular Vesicles Produced from the Stem Cells and Their Spheroids Cultured from Ocular Surgery-Derived Waste Orbicularis Oculi Muscle Tissues. Antioxidants. 2021; 10(8):1292. https://doi.org/10.3390/antiox10081292
Chicago/Turabian StyleLim, Kyung Min, Ahmed Abdal Dayem, Yujin Choi, Yoonjoo Lee, Jongyub An, Minchan Gil, Soobin Lee, Hee Jeong Kwak, Balachandar Vellingirl, Hyun Jin Shin, and et al. 2021. "High Therapeutic and Esthetic Properties of Extracellular Vesicles Produced from the Stem Cells and Their Spheroids Cultured from Ocular Surgery-Derived Waste Orbicularis Oculi Muscle Tissues" Antioxidants 10, no. 8: 1292. https://doi.org/10.3390/antiox10081292