
Salvigenin, a Trimethoxylated Flavone from Achillea Wilhelmsii C. Koch, Exerts Combined Lipid-Lowering and Mitochondrial Stimulatory Effects

 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Procedures
2.2. Plant Collection, Extraction, and Purification
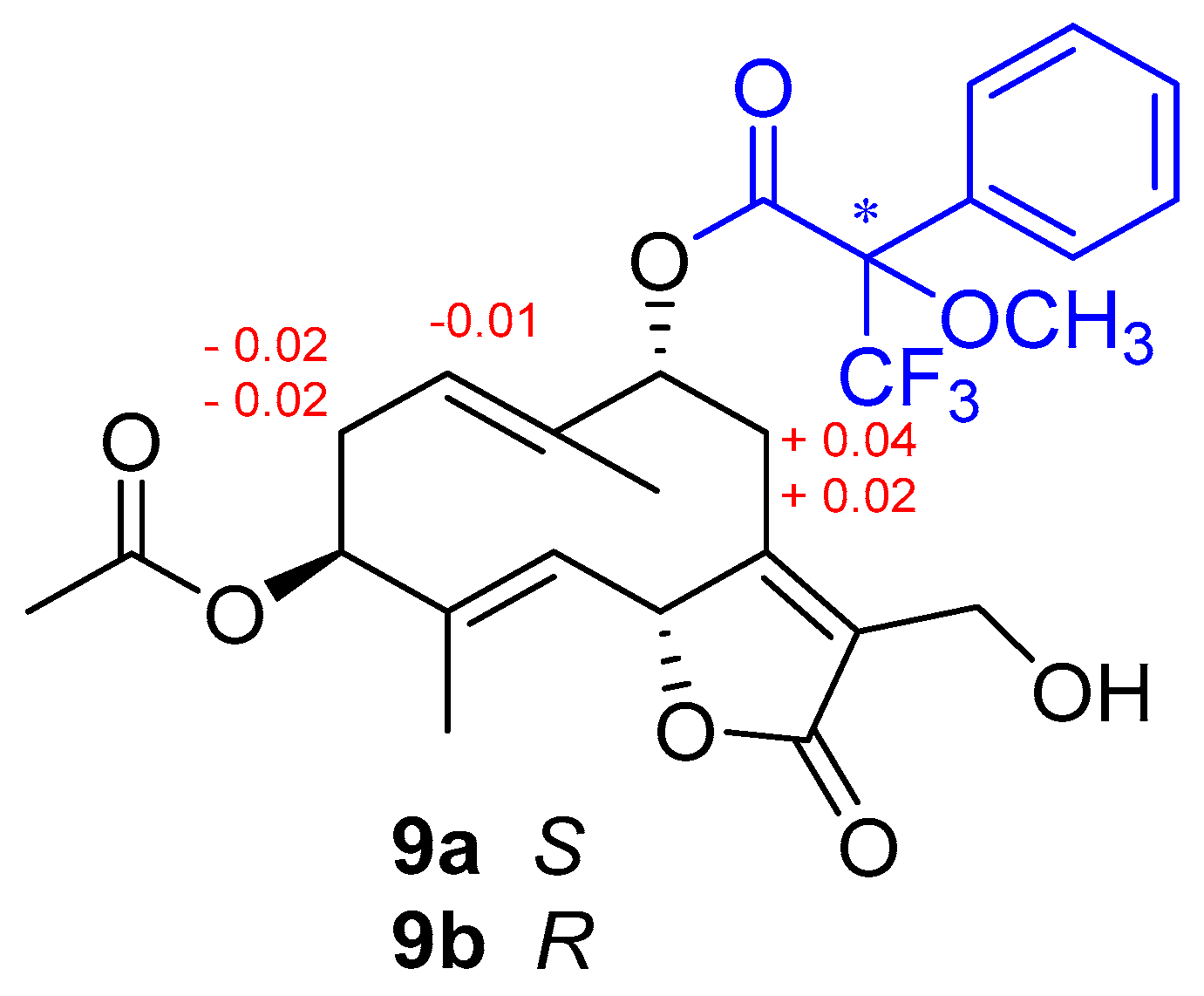
2.3. Mosher Ester Derivatization of Wilhelmsolide
2.4. Reagents for Biological Assays
2.5. Cell Culture and D2O Labeling
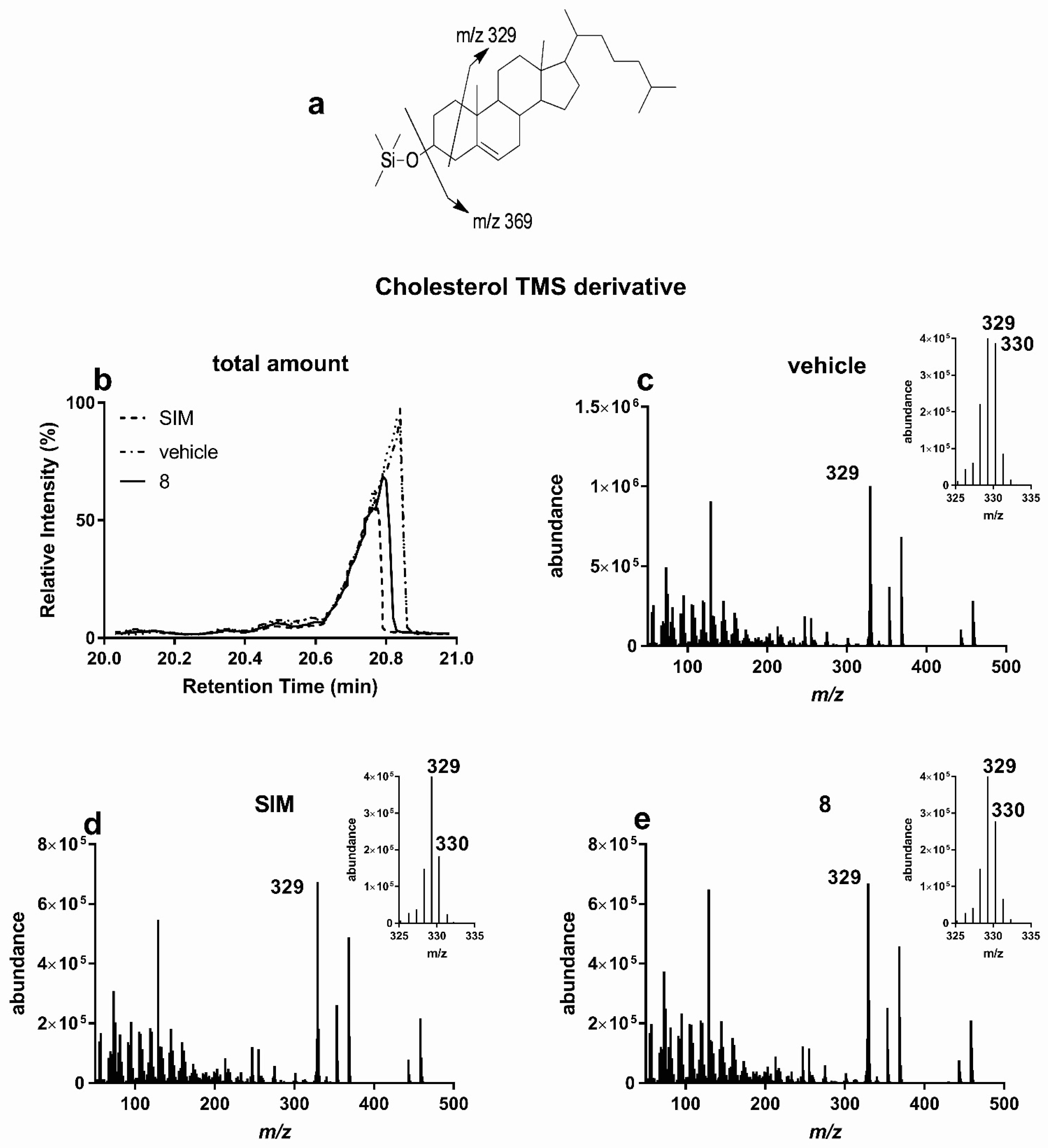
2.6. GC-MS for Biological Tests
2.7. Mitochondrial Activity
2.8. 2-NBDG Glucose Uptake Assay on HuH7 Cells
2.9. Statistical Analysis
3. Results
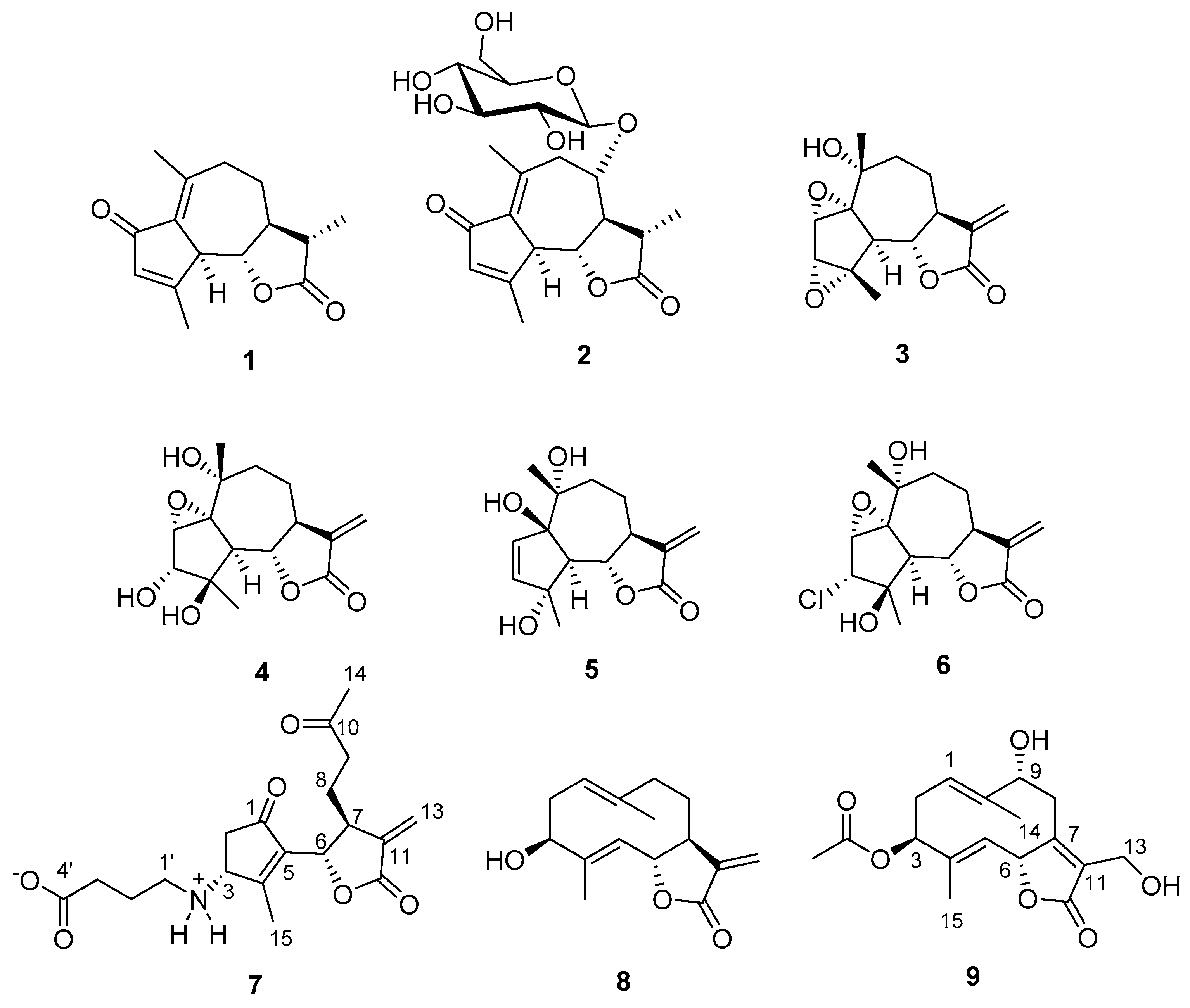
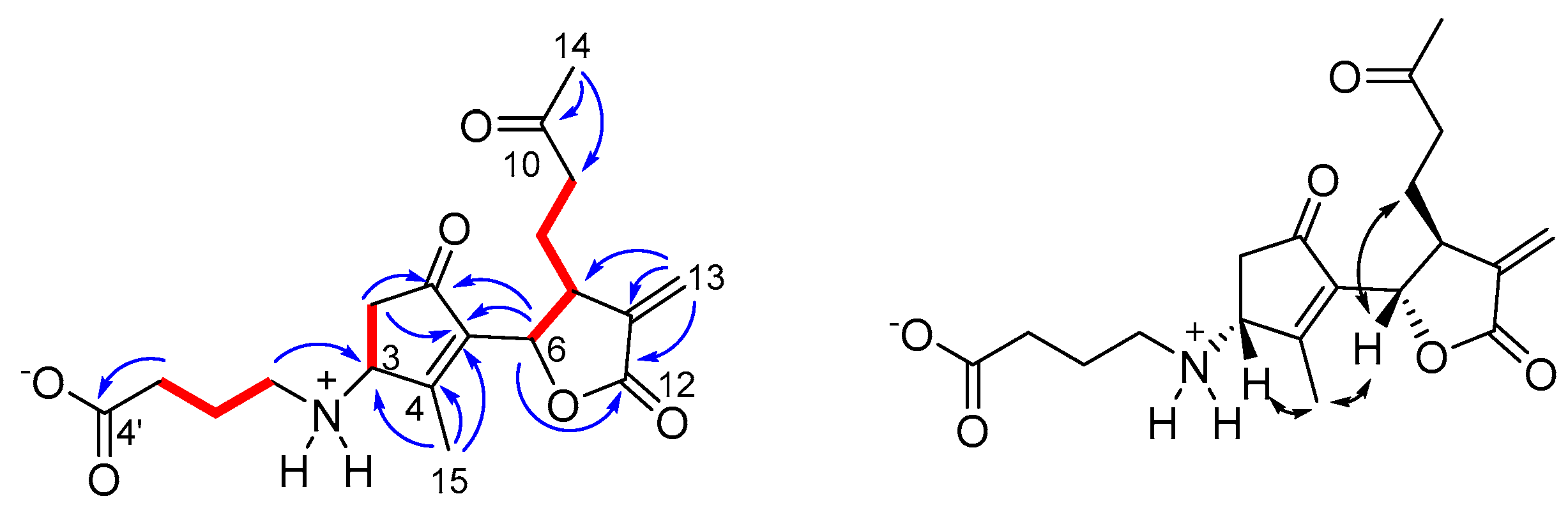
3.1. Chemical Analysis and Structure Elucidation of Wilhelmsin and Wilhelmsolide
3.2. Biological Evaluation
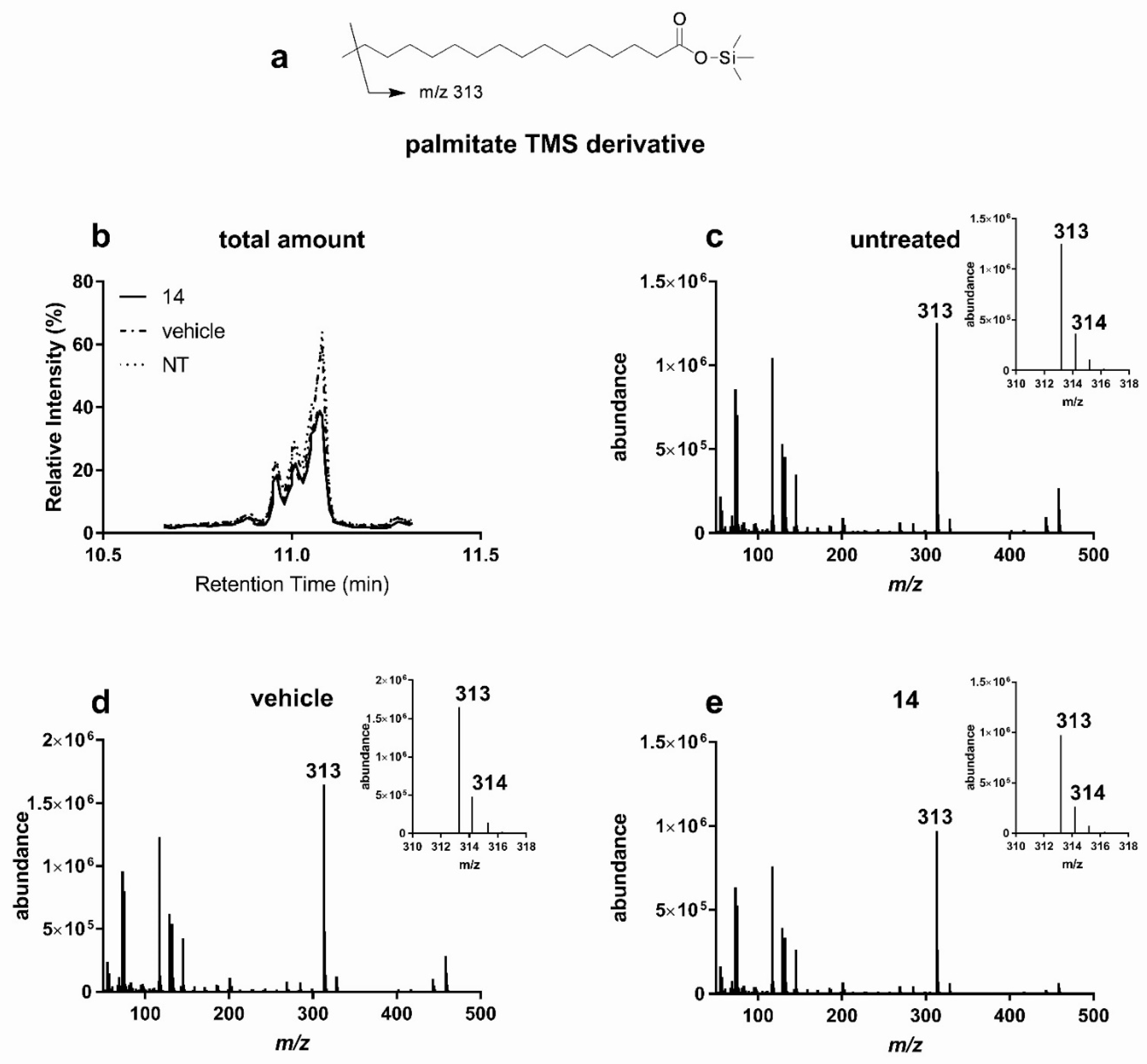
3.2.1. Lipid Lowering Activity
3.2.2. Mitochondrial Stimulatory Activity
3.2.3. Glucose Uptake Modulatory Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shin, J.-A.; Lee, J.-H.; Lim, S.-Y.; Ha, H.-S.; Kwon, H.-S.; Park, Y.-M.; Lee, W.-C.; Kang, M.-I.; Yim, H.-W.; Yoon, K.-H.; et al. Metabolic syndrome as a predictor of type 2 diabetes, and its clinical interpretations and usefulness. J. Diabetes Investig. 2013, 4, 334–343. [Google Scholar] [CrossRef]
- Srikanthan, K.; Feyh, A.; Visweshwar, H.; Shapiro, J.I.; Sodhi, K. Systematic Review of Metabolic Syndrome Biomarkers: A Panel for Early Detection, Management, and Risk Stratification in the West Virginian Population. Int. J. Med. Sci. 2016, 13, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Noce, A.; Di Lauro, M.; Di Daniele, F.; Zaitseva, A.P.; Marrone, G.; Borboni, P.; Di Daniele, N. Natural Bioactive Compounds Useful in Clinical Management of Metabolic Syndrome. Nutrients 2021, 13, 630. [Google Scholar] [CrossRef]
- Nasab, F.K.; Esmailpour, M. Ethno-medicinal survey on weed plants in agro-ecosystems: A case study in Jahrom, Iran. Environ. Dev. Sustain. 2018, 21, 2145–2164. [Google Scholar] [CrossRef]
- Derakhshande, P.; Navabi, S.J.; Shokoohinia, Y.; Rouhi-Broujeni, H.; Deris, F.; Behbood, L.; Shahbazi, F.; Amiri, M.; Farzaei, M.H. Efficacy and safety of Achillea wilhelmsii C. Koch capsules on symptom severity and quality of life in patients with irritable bowel syndrome: A randomized, placebo-controlled clinical trial. J. Complement. Integr. Med. 2019, 16. [Google Scholar] [CrossRef] [PubMed]
- Amiri, M.; Navabi, J.; Shokoohinia, Y.; Heydarpour, F.; Bahrami, G.; Behbood, L.; Derakhshandeh, P.; Momtaz, S.; Farzaei, M.H. Efficacy and safety of a standardized extract from Achillea wilhelmsii C. Koch in patients with ulcerative colitis: A randomized double blind placebo-controlled clinical trial. Complement. Ther. Med. 2019, 45, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudabady, M.; Lashkari, M.; Niazmand, S.; Soukhtanloo, M. Cardioprotective effects of Achillea wilhelmsii on the isolated rat heart in ischemia-reperfusion. J. Tradit. Complement. Med. 2017, 7, 501–507. [Google Scholar] [CrossRef]
- Asgary, S.; Naderi, G.H.; Sarrafzadegan, N.; Mohammadifard, N.; Mostafavi, S.; Vakili, R. Antihypertensive and antihyperlipidemic effects of Achillea wilhelmsii. Drugs Exp. Clin. Res. 2000, 26, 89–93. [Google Scholar]
- Khazneh, E.; Hribová, P.; Hošek, J.; Suchý, P.; Kollár, P.; Pražanová, G.; Muselík, J.; Hanaková, Z.; Václavík, J.; Miłek, M.; et al. The chemical composition of Achillea wilhelmsii C. Koch and Its desirable effects on hyperglycemia, inflammatory mediators and hypercholesterolemia as risk factors for cardiometabolic disease. Molecules 2016, 21, 404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommella, E.; Badolati, N.; Riccio, G.; Salviati, E.; Bottone, S.; Dentice, M.; Campiglia, P.; Tenore, G.C.; Stornaiuolo, M.; Novellino, E. A Boost in Mitochondrial Activity Underpins the Cholesterol-Lowering Effect of Annurca Apple Polyphenols on Hepatic Cells. Nutrients 2019, 11, 163. [Google Scholar] [CrossRef] [Green Version]
- Asariha, M.; Chahardoli, A.; Karimi, N.; Gholamhosseinpour, M.; Khoshroo, A.; Nemati, H.; Shokoohinia, Y.; Fattahi, A. Green synthesis and structural characterization of gold nanoparticles from Achillea wilhelmsii leaf infusion and in vitro evaluation. Bull. Mater. Sci. 2020, 43, 1–10. [Google Scholar] [CrossRef]
- Martinez, V.M.; Muñoz-Zamora, A.; Joseph-Nathan, P. Conformational analysis of achillin and leukodin. J. Nat. Prod. 1988, 51, 221–228. [Google Scholar] [CrossRef]
- Michalska, K.; Kisiel, W. Sesquiterpene lactones from Taraxacum obovatum. Planta Med. 2003, 69, 181–183. [Google Scholar] [CrossRef]
- Bohlmann, F.; Zdero, C. Sesquiterpene lactones and other constituents from Tanacetum parthenium. Phytochemistry 1982, 21, 2543–2549. [Google Scholar] [CrossRef]
- Huneck, S.; Zdero, C.; Bohlmann, F. Seco-guaianolides and other constituents from Artemisia species. Phytochemistry 1986, 25, 883–889. [Google Scholar] [CrossRef]
- Zhang, L.-B.; Lv, J.-L. Sesquiterpenoids from Artemisia argyi and their COXs inhibitory activities. Fitoterapia 2019, 139. [Google Scholar] [CrossRef] [PubMed]
- Mata, R.; Delgado, G.; De Vivar, A.R. Sesquiterpene lactones of artemisia klotzchiana. Phytochemistry 1985, 24, 1515–1519. [Google Scholar] [CrossRef]
- Asakawa, Y.; Toyota, M.; Takemoto, T. Two guaiane-type sesquiterpene lactones and their related sesquiterpene lactones from Porella japonica. Phytochemistry 1981, 20, 257–261. [Google Scholar] [CrossRef]
- Forino, M.; Tenore, G.C.; Tartaglione, L.; Carmela, D.; Novellino, E.; Ciminiello, P. (1S,3R,4S,5R)5-O-Caffeoylquinic acid: Isolation, stereo-structure characterization and biological activity. Food Chem. 2015, 178, 306–310. [Google Scholar] [CrossRef]
- Bai, N.; He, K.; Zhou, Z.; Lai, C.-S.; Zhang, L.; Quan, Z.; Shao, X.; Pan, M.-H.; Ho, C.-T. Flavonoids from Rabdosia rubescens exert anti-inflammatory and growth inhibitory effect against human leukemia HL-60 cells. Food Chem. 2010, 122, 831–835. [Google Scholar] [CrossRef]
- Kontogianni, V.G.; Primikyri, A.; Sakka, M.; Gerothanassis, I.P. Simultaneous determination of artemisinin and its analogs and flavonoids in Artemisia annua crude extracts with the use of NMR spectroscopy. Magn. Reson. Chem. 2019, 58, 232–244. [Google Scholar] [CrossRef]
- Balboul, B.A.; Ahmed, A.A.; Otsuka, H.; Bando, M.; Kido, M.; Takeda, Y. A guaianolide and a germacranolide from Achillea santolina. Phytochemistry 1997, 46, 1045–1049. [Google Scholar] [CrossRef]
- Nair, A.G.R.; Ananda Rao, S.; Voirin, B.; Bonvin, J.F. Polyphenolic compounds from leaves of Acanthospermum hispidum. Fitoterapia 1985, 56, 249–250. [Google Scholar]
- Flores, I.S.; Martinelli, B.C.; Lião, L.M. High-resolution magic angle spinning nuclear magnetic resonance (HR-MAS NMR) as a tool in the determination of biomarkers of Passiflora-based herbal medicines. Fitoterapia 2020, 142. [Google Scholar] [CrossRef]
- Formisano, C.; Rigano, D.; Lopatriello, A.; Sirignano, C.; Ramaschi, G.; Arnoldi, L.; Riva, A.; Sardone, N.; Taglialatela-Scafati, O. Detailed Phytochemical Characterization of Bergamot Polyphenolic Fraction (BPF) by UPLC-DAD-MS and LC-NMR. J. Agric. Food Chem. 2019, 67, 3159–3167. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Veitch, N.C.; Houghton, P.J.; Simmonds, M.S. Flavone C-glycosides from Viola yedoensis Makino. Chem. Pharm. Bull. 2003, 51, 1204–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramesh, S.A.; Tyerman, S.D.; Xu, B.; Bose, J.; Kaur, S.; Conn, V.; Domingos, P.; Ullah, S.; Wege, S.; Shabala, S.; et al. GABA signalling modulates plant growth by directly regulating the activity of plant-specific anion transporters. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Ohtani, I.; Kusumi, T.; Kashman, Y.; Kakisawa, H. High-field FT NMR application of Mosher’s method. The absolute configurations of marine terpenoids. J. Am. Chem. Soc. 1991, 113, 4092–4096. [Google Scholar] [CrossRef]
- Graf, B.L.; Raskin, I.; Cefalu, W.T.; Ribnicky, D.M. Plant-derived therapeutics for the treatment of metabolic syndrome. Curr. Opin. Investig. Drugs 2010, 11, 1107–1115. [Google Scholar] [CrossRef]
- Hall, I.H.; Lee, K.H.; Starnes, C.O.; Muraoka, O.; Sumida, Y.; Waddell, T.G. Antihyperlipidemic activity of sesquiterpene lactones and related compounds. J. Pharm. Sci. 1980, 69, 694–697. [Google Scholar] [CrossRef]
- Bhatia, G.; Khanna, A.K.; Sonkar, R.; Mishra, S.K.; Srivastava, S.; Lakshmi, V. Lipid lowering and antioxidant activity of flavones in triton treated hyperlipidemic rats. Med. Chem. Res. 2010, 20, 1622–1626. [Google Scholar] [CrossRef]
- Babaei, F.; Moafizad, A.; Darvishvand, Z.; Mirzababaei, M.; Hosseinzadeh, H.; Nassiri-As, M. Review of the effects of vitexin in oxidative stress-related diseases. Food Sci. Nutr. 2020, 8, 2569–2580. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pos. | δH, Multiplicity, J in Hz | δC, Type |
|---|---|---|
| 1 | 203.7, C | |
| 2a | 2.69–2.71 | 37.2, CH2 |
| 2b | 2.44–2.45 | |
| 3 | 5.22, br d, 6.5 | 53.5, CH |
| 4 | 171.4, C | |
| 5 | 139.1, C | |
| 6 | 5.08, d, 5.4 | 76.3, CH |
| 7 | 3.19, overlapped | 42.7, CH |
| 8a | 1.94–1.96 | 27.3, CH2 |
| 8b | 1.86–1.88 | |
| 9 | 2.59, t, 7.3 | 39.2, CH2 |
| 10 | 208.9, C | |
| 11 | 139.0, C | |
| 12 | 170.8, C | |
| 13a | 6.26, d, 2.8 | 121.3, CH2 |
| 13b | 5.75, d, 2.8 | |
| 14 | 2.19, s | 28.5, CH3 |
| 15 | 2.05, s | 12.8, CH3 |
| 3.38–3.39 | 42.8, CH2 | |
| 1′ | 3.10, overlapped | |
| 2′ | 2.09, overlapped | 17.5, CH2 |
| 3′ | 2.45, overlapped | 30.7, CH2 |
| 4′ | 177.1, C |
| Pos. | δH, Multiplicity, J in Hz | δC, Type |
|---|---|---|
| 1 | 4.99, dd, 2.5, 12.1 | 125.8, CH |
| 2a | 2.54–2.56 | 30.1, CH2 |
| 2b | 2.35–2.36 | |
| 3 | 5.14, dd, 6.1, 10.3 | 77.8, CH |
| 4 | 137.4, C | |
| 5 | 4.38, d, 10.5 | 122.6, CH |
| 6 | 5.44, d, 10.5 | 80.6, CH |
| 7 | 161.9, CH | |
| 8a | 2.98, dd, 13.8, 1.5 | 35.0, CH2 |
| 8b | 2.60, dd, 10.1, 13.8 | |
| 9a | 4.27, dd, 10.1, 1.5 | 78.1, CH |
| 10 | 139.5, C | |
| 11 | 129.7, C | |
| 12 | 172.9, C | |
| 13a | 4.48, d, 14.1 | 55.2, CH2 |
| 13b | 4.54, d, 14.1 | |
| 14 | 1.64, s | 10.5, CH3 |
| 15 | 1.74, s | 12.0, CH3 |
| AcO- | 2.10, s | 21.1, CH3 170.1, C |
| Compound | Cholesterol Biosynthesis a | Palmitic Acid Biosynthesis b | Mitochondrial Activity c | Glucose Uptake d |
|---|---|---|---|---|
| vehicle | –8.3 ± 0.6 ** | 2.5 ± 0.2 * | –0.3 ± 0.6 n.s | 0.4 ± 0.2 n.s |
| Simvastatin | 14.8 ± 4.0 *** | - | - | - |
| A.P.E. (400 mg/L) | - | –47.6 ± 1.9 *** | - | - |
| A.P.E. (400 mg/L) | - | - | 22.6 ± 0.9 *** | - |
| Insulin (100 nM) | - | - | - | 74.4 ± 3.6 *** |
| 1 | −7.3 ± 0.2 * | 0.7 ± 1.1 n.s. | 2.1 ± 0.2 *** | 0.9 ± 1.2 n.s. |
| 2 | −8.1 ± 0.4 ** | –1.9 ± 0.3 n.s. | 0.3 ± 0.3 n.s. | 1.9 ± 0.4 n.s. |
| 3 | −8.1 ± 2.2 ** | 0.1 ± 1.7 n.s | 2.4 ± 0.2 *** | 0.3 ± 0.5 n.s. |
| 4 | −9.1 ± 3.0 * | 1.2 ± 0.2 n.s. | 3.7 ± 0.6*** | 0.9 ± 0.6 n.s. |
| 5 | −10.1 ± 2.2 * | –1.6 ± 2.1 n.s. | 0.5 ± 0.2 n.s. | 0.4 ± 0.7 n.s. |
| 6 | –17.4 ± 7.4 * | –1.5 ± 0.2 n.s. | 1.9 ± 0.5 *** | 1.7 ± 1.2 n.s. |
| 7 | –19.9 ± 4.1 * | 2.1 ± 1.2 n.s. | 2.6 ± 0.3 *** | 0.8 ± 0.7 n.s. |
| 8 | –12.7 ± 0.1 *** | 2.9 ± 1.7 * | 1.8 ± 0.2 *** | 1.4 ± 0.4 n.s. |
| 12 | -8.6 ± 3.2 * | 0.9 ± 1.2 n.s. | 0.2 ± 0.5 n.s. | 0.7 ± 0.8 n.s |
| 13 | –12.2 ± 2.2 ** | –6.3 ± 0.4 *** | 1.6 ± 0.2 *** | 16.2 ± 0.2 *** |
| 14 | –9.1 ± 1.6 * | –22.5 ± 0.7 *** | 15.4 ± 0.4 *** | 0.7 ± 0.5 n.s |
| 15 | –8.1 ± 0.9 * | 1.0 ± 0.5 n.s | 0.8 ± 0.6 n.s. | 1.4 ± 0.9 n.s |
| 16 | –5.7 ± 2.2 n.s. | 2.3 ± 1.5 n.s. | 2.2 ± 0.3 *** | 0.6 ± 1.9 n.s |
| 17 | –6.9 ± 2.9 * | 1.9 ± 1.3 n.s. | 1.9 ± 0.5 *** | 1.5 ± 0.5 n.s |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serino, E.; Chahardoli, A.; Badolati, N.; Sirignano, C.; Jalilian, F.; Mojarrab, M.; Farhangi, Z.; Rigano, D.; Stornaiuolo, M.; Shokoohinia, Y.; et al. Salvigenin, a Trimethoxylated Flavone from Achillea Wilhelmsii C. Koch, Exerts Combined Lipid-Lowering and Mitochondrial Stimulatory Effects. Antioxidants 2021, 10, 1042. https://doi.org/10.3390/antiox10071042
Serino E, Chahardoli A, Badolati N, Sirignano C, Jalilian F, Mojarrab M, Farhangi Z, Rigano D, Stornaiuolo M, Shokoohinia Y, et al. Salvigenin, a Trimethoxylated Flavone from Achillea Wilhelmsii C. Koch, Exerts Combined Lipid-Lowering and Mitochondrial Stimulatory Effects. Antioxidants. 2021; 10(7):1042. https://doi.org/10.3390/antiox10071042
Chicago/Turabian StyleSerino, Elena, Azam Chahardoli, Nadia Badolati, Carmina Sirignano, Fereshteh Jalilian, Mahdi Mojarrab, Zahra Farhangi, Daniela Rigano, Mariano Stornaiuolo, Yalda Shokoohinia, and et al. 2021. "Salvigenin, a Trimethoxylated Flavone from Achillea Wilhelmsii C. Koch, Exerts Combined Lipid-Lowering and Mitochondrial Stimulatory Effects" Antioxidants 10, no. 7: 1042. https://doi.org/10.3390/antiox10071042