
The Mechanisms of Thiosulfate Toxicity against Saccharomyces cerevisiae


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Strains, Mutants, and Plasmids
2.3. Measurement of Growth Curves of S. cerevisiae in the Presence of Thiosulfate or Sulfite
2.4. Detection of Killing Effect of Thiosulfate at Low pH
2.5. Rhodanese Assay
2.6. Determination of Cellular Thiosulfate Concentration
2.7. Measurements of Mitochondrial Membrane Potential and Cellular ATP Concentration
2.8. Assaying GAPDH Activity
2.9. Measuring Oxygen Consumption
3. Results
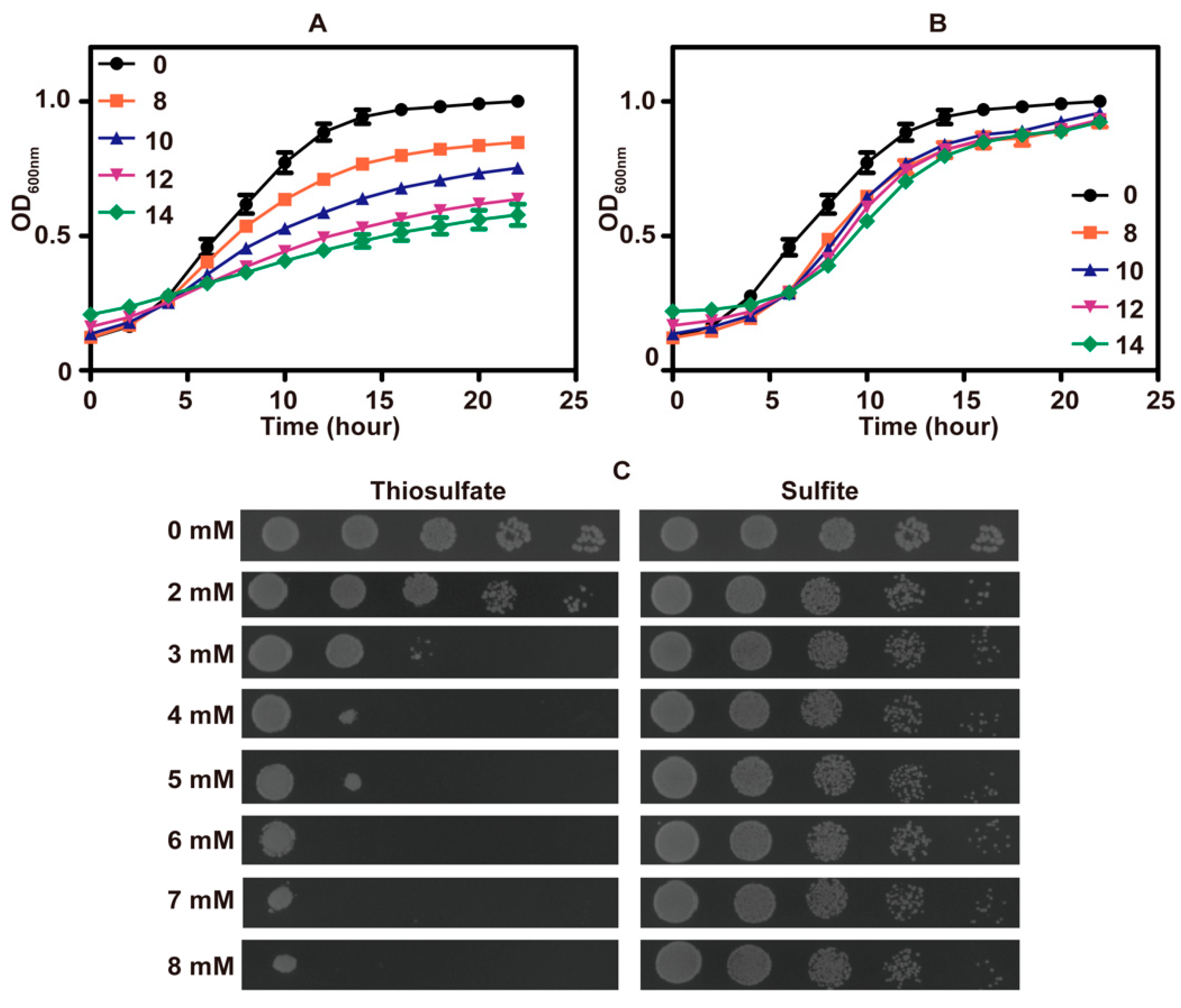
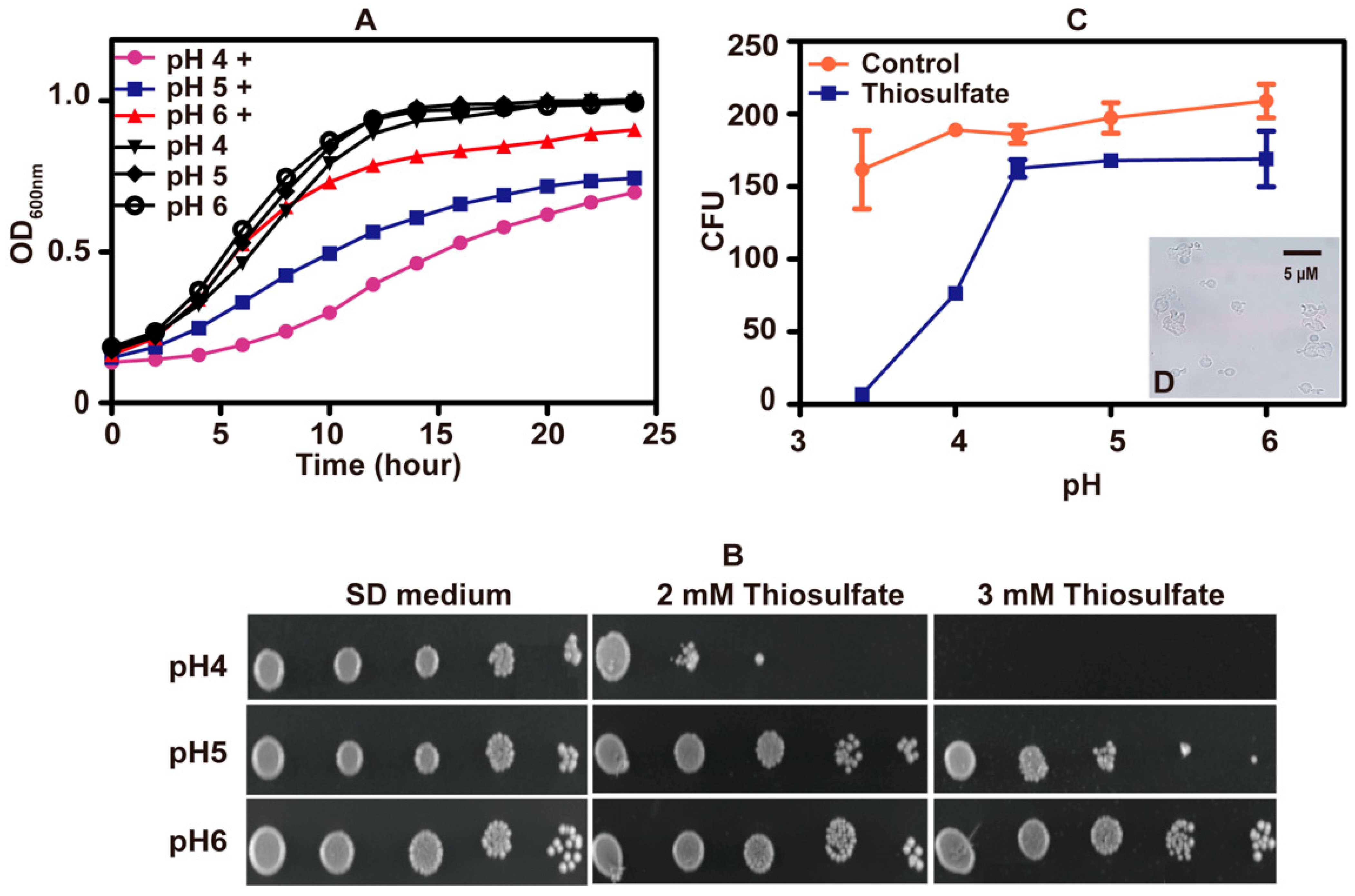
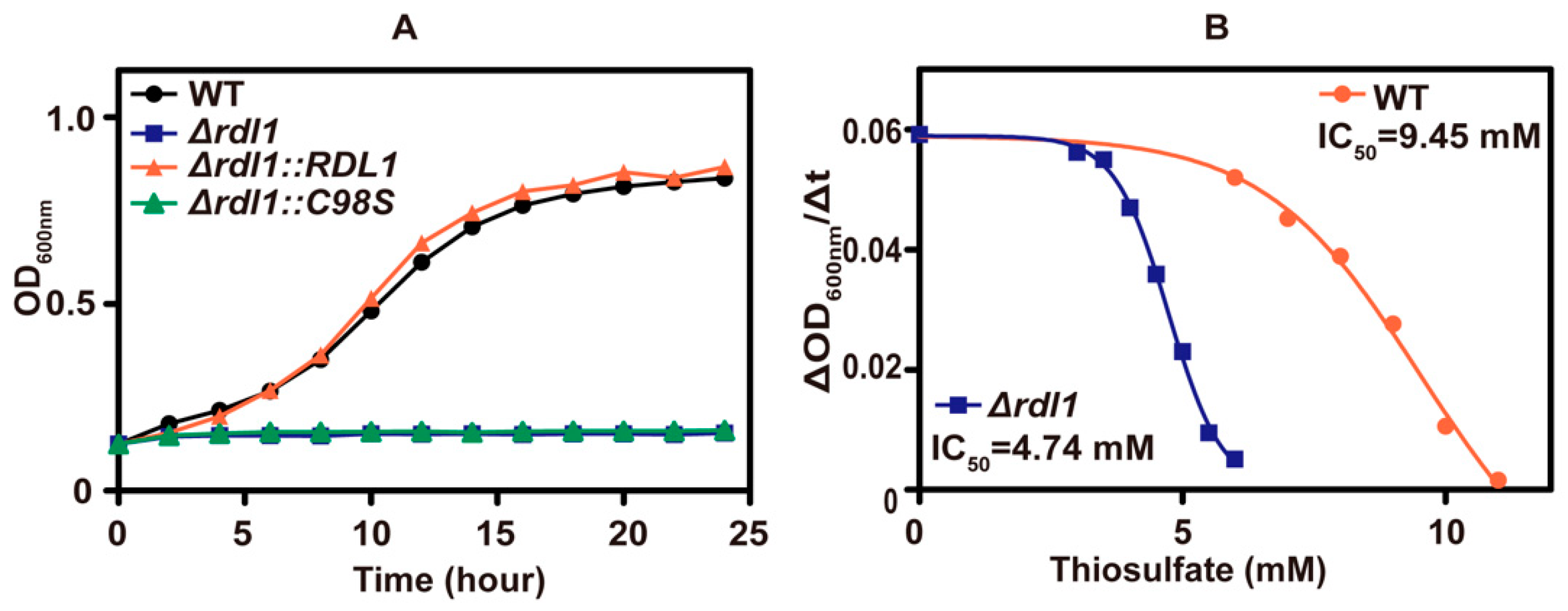
3.1. Thiosulfate Inhibited the Growth of S. cerevisiae at pH 6 and Killed It at pH 3.4
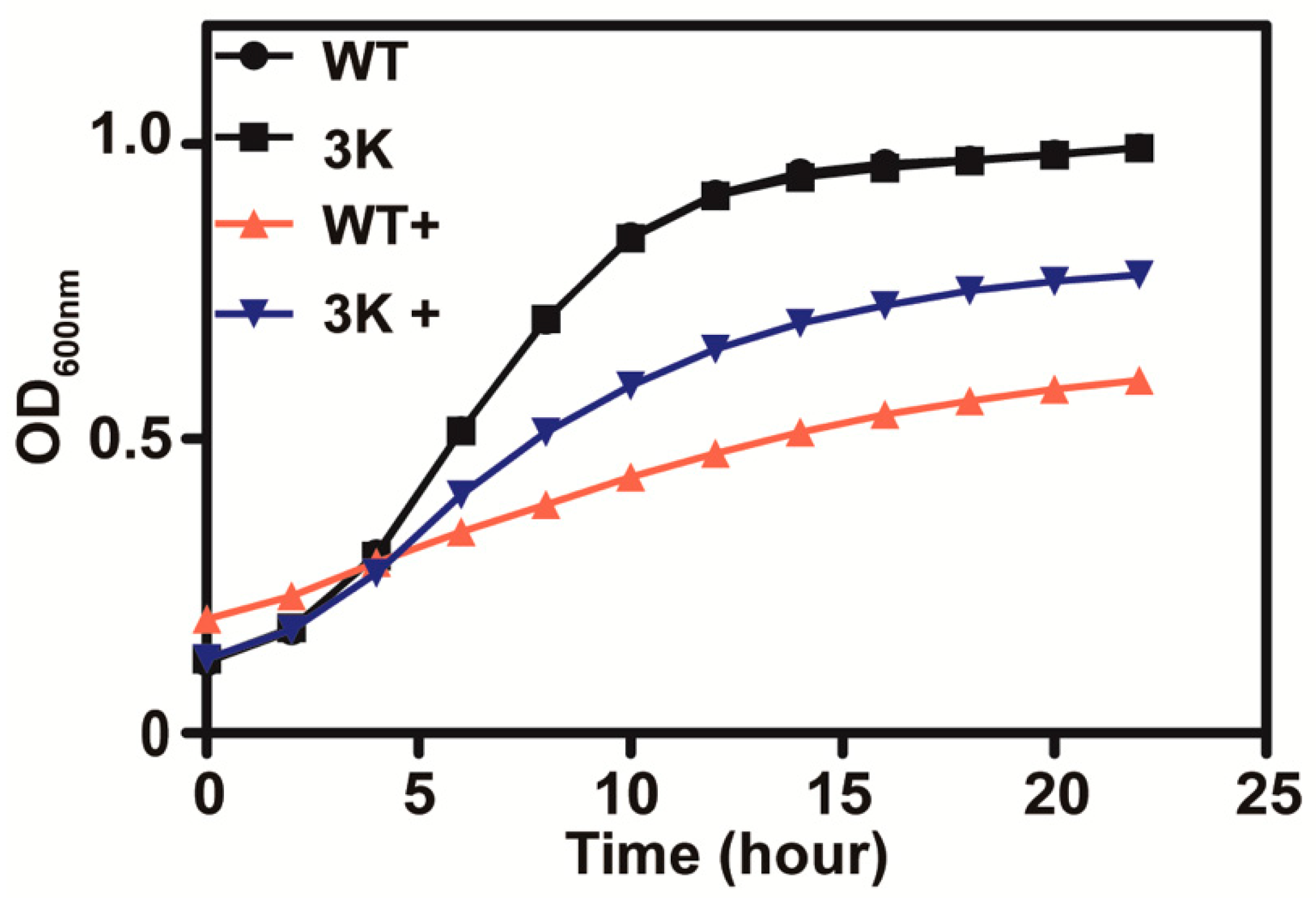
3.2. Thiosulfate Is Actively Transported into the Cells for Its Inhibition
3.3. Thiosulfate Itself Is Inhibitory to the Yeast at pH 6
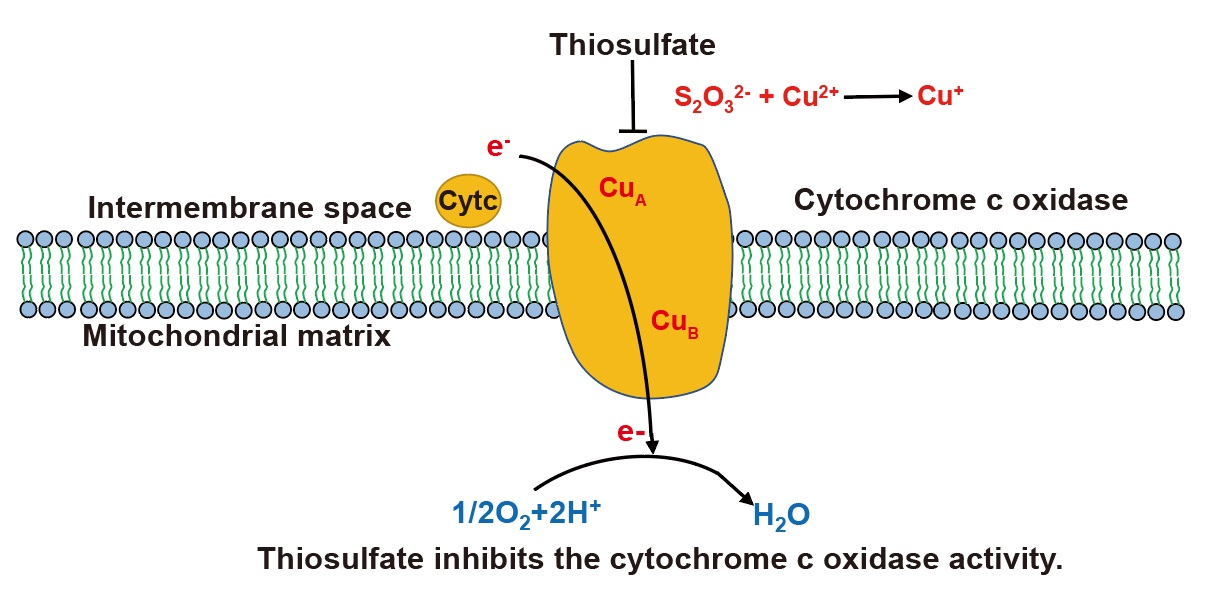
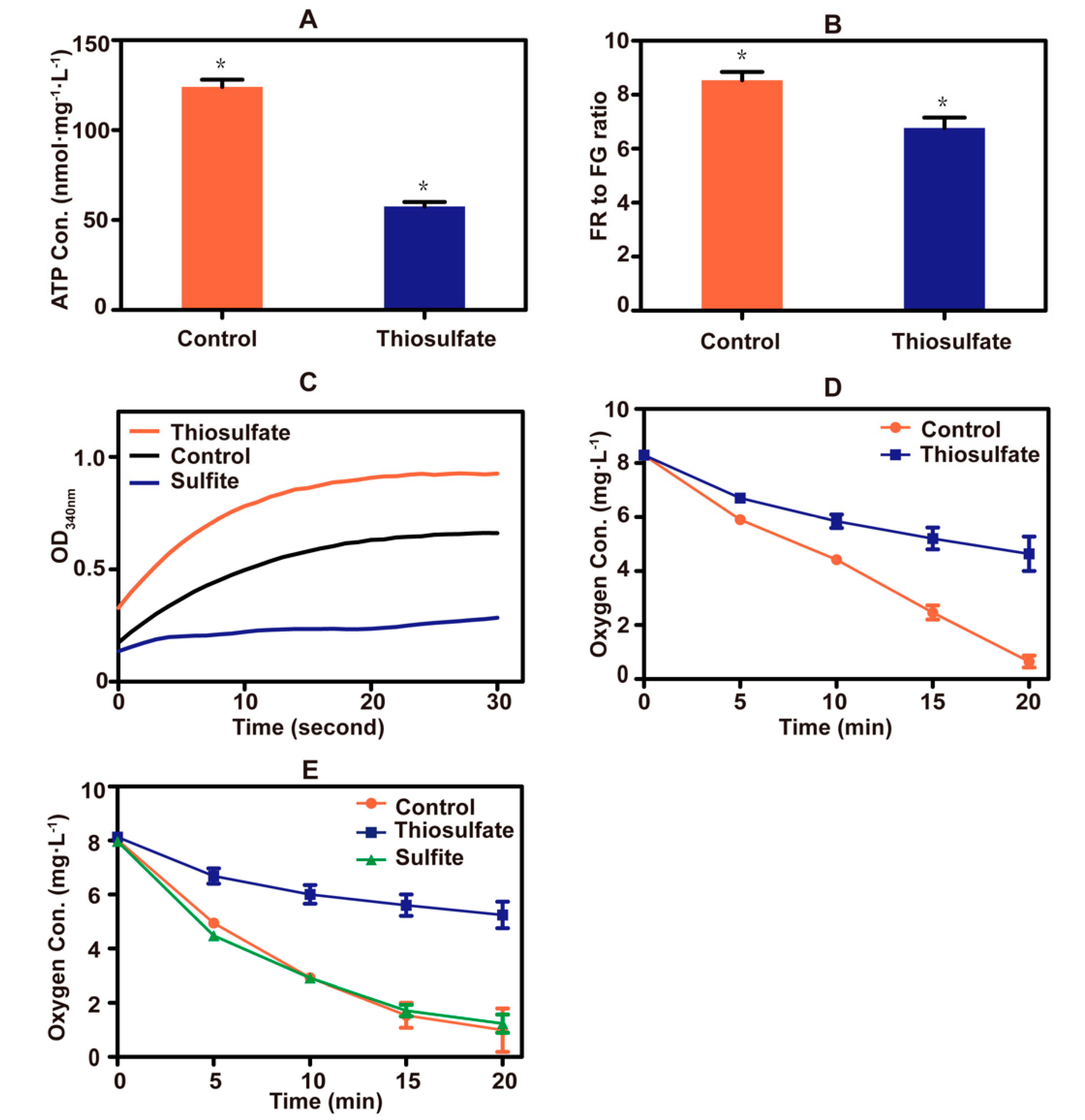
3.4. Thiosulfate Perturbs the Mitochondrial Bioenergetics in S. cerevisiae
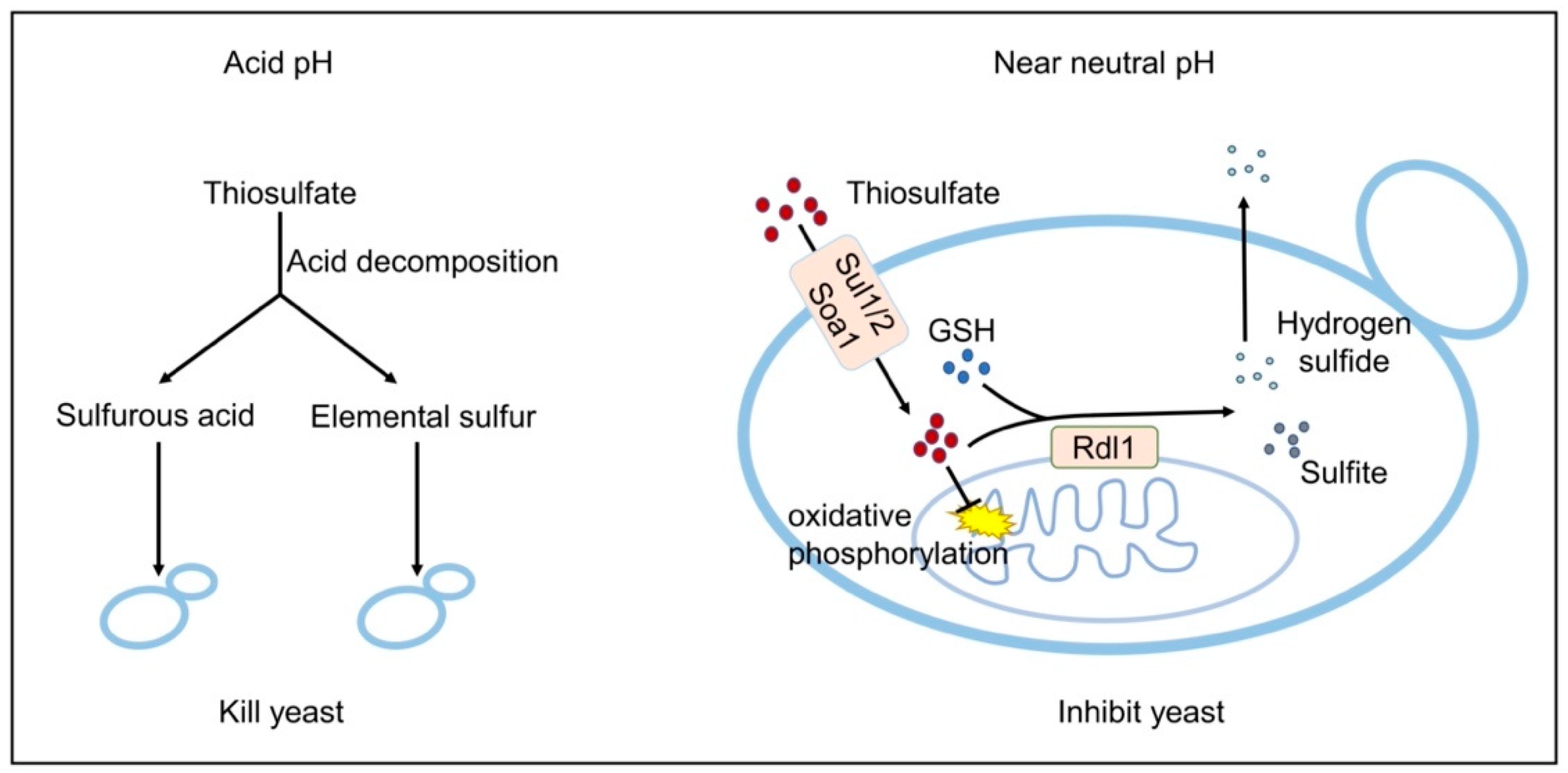
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meyer, B. Elemental sulfur. Chem. Rev. 1976, 76, 367–388. [Google Scholar] [CrossRef]
- Jorgensen, B.B. A Thiosulfate Shunt in the Sulfur Cycle of Marine Sediments. Science 1990, 249, 152–154. [Google Scholar] [CrossRef]
- Xin, Y.; Liu, H.; Cui, F.; Xun, L. Recombinant Escherichia coli with sulfide: Quinone oxidoreductase and persulfide dioxygenase rapidly oxidises sulfide to sulfite and thiosulfate via a new pathway. Environ. Microbiol. 2016, 18, 5123–5136. [Google Scholar] [CrossRef]
- Xin, Y.; Gao, R.; Cui, F.; Lu, C.; Liu, H.; Liu, H.; Xia, Y.; Xun, L. The heterotrophic bacterium Cupriavidus pinatubonensis JMP134 oxidizes sulfide to sulfate with thiosulfate as a key intermediate. Appl. Environ. Microbiol. 2020, 86, e01835-20. [Google Scholar] [CrossRef]
- Xia, Y.; Lu, C.; Hou, N.; Xin, Y.; Liu, J.; Liu, H.; Xun, L. Sulfide production and oxidation by heterotrophic bacteria under aerobic conditions. ISME J. 2017, 11, 2754–2766. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liu, R.; Xi, S.; Cai, R.; Sun, C. A novel bacterial thiosulfate oxidation pathway provides a new clue about the formation of zero-valent sulfur in deep sea. ISME J. 2020, 14, 2261–2274. [Google Scholar] [CrossRef] [PubMed]
- Rameez, M.J.; Pyne, P.; Mandal, S.; Chatterjee, S.; Ghosh, W.J.M.R. Two pathways for thiosulfate oxidation in the alphaproteobacterial chemolithotroph Paracoccus thiocyanatus SST. Microbiol. Res. 2019, 230, 126345. [Google Scholar] [CrossRef]
- Houghton, J.L.; Foustoukos, D.I.; Flynn, T.M.; Vetriani, C.; Bradley, A.S.; Fike, D.A. Thiosulfate oxidation by Thiomicrospira thermophila: Metabolic flexibility in response to ambient geochemistry. Environ. Microbiol. 2016, 18, 3057–3072. [Google Scholar] [CrossRef] [Green Version]
- Funahashi, E.; Saiki, K.; Honda, K.; Sugiura, Y.; Kawano, Y.; Ohtsu, I.; Watanabe, D.; Wakabayashi, Y.; Abe, T.; Nakanishi, T.; et al. Finding of thiosulfate pathway for synthesis of organic sulfur compounds in Saccharomyces cerevisiae and improvement of ethanol production. J. Biosci. Bioeng. 2015, 120, 666–669. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, X.; Li, H.; Liu, H.; Xia, Y.; Xun, L. The complete pathway for thiosulfate utilization in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2018, 84, e01241-18. [Google Scholar] [CrossRef] [Green Version]
- Kawano, Y.; Onishi, F.; Shiroyama, M.; Miura, M.; Tanaka, N.; Oshiro, S.; Nonaka, G.; Nakanishi, T.; Ohtsu, I. Improved fermentative L-cysteine overproduction by enhancing a newly identified thiosulfate assimilation pathway in Escherichia coli. Appl. Microbiol. Biotechnol. 2017, 101, 6879–6889. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Yamagata, S.; Masui, R.; Inoue, Y.; Shibata, T.; Yokoyama, S.; Kuramitsu, S.; Iwama, T. Cloning and overexpression of the oah1 gene encoding O-acetyl-L-homoserine sulfhydrylase of Thermus thermophilus HB8 and characterization of the gene product. Biochim. Biophys. Acta 2001, 1549, 61–72. [Google Scholar] [CrossRef]
- Melideo, S.L.; Jackson, M.R.; Jorns, M.S. Biosynthesis of a central intermediate in hydrogen sulfide metabolism by a novel human sulfurtransferase and its yeast ortholog. Biochemistry 2014, 53, 4739–4753. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.W.; Walker, M.E.; Fedrizzi, B.; Gardner, R.C.; Jiranek, V. Hydrogen sulfide and its roles in Saccharomyces cerevisiae in a winemaking context. FEMS Yeast Res. 2017, 17, fox058. [Google Scholar] [CrossRef] [Green Version]
- Dinegar, R.H.; Smellie, R.H.; Mer, V.K.L. Kinetics of the acid decomposition of sodium thiosulfate in dilute solutions. J. Am. Chem. Soc. 1951, 73, 2050–2054. [Google Scholar] [CrossRef]
- Taylor, S.L.; Higley, N.A.; Bush, R.K. Sulfites in foods: Uses, analytical methods, residues, fate, exposure assessment, metabolism, toxicity, and hypersensitivity. Adv. Food Res. 1986, 30, 1–76. [Google Scholar]
- Liu, X.; Sang, M.; Zhang, X.; Zhang, T.; Zhang, H.; He, X.; Li, S.; Sun, X.; Zhang, Z. Enhancing expression of SSU1 genes in Saccharomyces uvarum leads to an increase in sulfite tolerance and transcriptome profiles change. FEMS Yeast Res. 2017, 17, fox023. [Google Scholar] [CrossRef]
- Chang, I.S.; Kim, B.H.; Shin, P.K.; Microbiology, A.E. Use of sulfite and hydrogen peroxide to control bacterial contamination in ethanol fermentation. Appl. Environ. Microbiol. 1997, 63, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Nardi, T.; Corich, V.; Giacomini, A.; Blondin, B. A sulphite-inducible form of the sulphite efflux gene SSU1 in a Saccharomyces cerevisiae wine yeast. Microbiology 2010, 156, 1686–1696. [Google Scholar] [CrossRef] [Green Version]
- King, A.D., Jr.; Ponting, J.D.; Sanshuck, D.W.; Jackson, R.; Mihara, K. Factors Affecting Death of Yeast by Sulfur Dioxide. J. Food Prot. 1981, 44, 92–97. [Google Scholar] [CrossRef]
- Schimz, K.L.; Holzer, H. Rapid decrease of ATP content in intact cells of Saccharomyces cerevisiae after incubation with low concentrations of sulfite. Arch. Microbiol. 1979, 121, 225–229. [Google Scholar] [CrossRef]
- Hinze, H.; Holzer, H. Analysis of the energy metabolism after incubation of Saccharomyces cerevisiae with sulfite or nitrite. Arch. Microbiol. 1986, 145, 27–31. [Google Scholar] [CrossRef]
- Steinacker, A. A suggested mechanism of action for sulfite sensitivity. J. Allergy Clin. Immunol. 1986, 77, 116–117. [Google Scholar] [CrossRef]
- Neta, P.; Huie, R.E. Free-radical chemistry of sulfite. Environ. Health Perspect. 1985, 64, 209–217. [Google Scholar] [CrossRef] [PubMed]
- McCord, J.M.; Fridovich, I. The utility of superoxide dismutase in studying free radical reactions. I. Radicals generated by the interaction of sulfite, dimethyl sulfoxide, and oxygen. J. Biol. Chem. 1969, 244, 6056–6063. [Google Scholar] [CrossRef]
- Peiser, G.D.; Yang, S.F. Sulfite-mediated destruction of β-carotene. J. Agric. Food Chem. 1979, 27, 446–449. [Google Scholar] [CrossRef]
- Yang, S.F. Destruction of tryptophan during the aerobic oxidation of sulfite ions. Environ. Res. 1973, 6, 395–402. [Google Scholar] [CrossRef]
- Williams, J.S.; Cooper, R.M. The oldest fungicide and newest phytoalexin—A reappraisal of the fungitoxicity of elemental sulphur. Plant Pathol. 2010, 53, 263–279. [Google Scholar] [CrossRef]
- Rai, M.; Ingle, A.P.; Paralikar, P. Sulfur and sulfur nanoparticles as potential antimicrobials: From traditional medicine to nanomedicine. Expert Rev. Anti Infect. Ther. 2016, 14, 969–978. [Google Scholar] [CrossRef]
- Sato, I.; Shimatani, K.; Fujita, K.; Abe, T.; Shimizu, M.; Fujii, T.; Hoshino, T. Glutathione reductase/glutathione is responsible for cytotoxic elemental sulfur tolerance via polysulfide shuttle in fungi. J. Biol. Chem. 2011, 286, 20283–20291. [Google Scholar] [CrossRef] [Green Version]
- Islamov, R.A.; Bishimova, I.; Sabitov, A.N.; Ilin, A.I.; Burkitbaev, M.M. Lack of Mutagenic Activity of Sulfur Nanoparticles in Micronucleus Test on L5178Y Cell Culture. Cell Tissue Biol. 2018, 12, 27–32. [Google Scholar] [CrossRef]
- Choudhury, S.R.; Mandal, A.; Ghosh, M.; Basu, S.; Chakravorty, D.; Goswami, A. Investigation of antimicrobial physiology of orthorhombic and monoclinic nanoallotropes of sulfur at the interface of transcriptome and metabolome. Appl. Microbiol. Biotechnol. 2013, 97, 5965–5978. [Google Scholar] [CrossRef]
- Cetkauskaite, A.; Pessala, P.; Sodergren, A. Elemental sulfur: Toxicity in vivo and in vitro to bacterial luciferase, in vitro yeast alcohol dehydrogenase, and bovine liver catalase. Environ. Toxicol. 2004, 19, 372–386. [Google Scholar] [CrossRef]
- De La Calzada-Jeanlouie, M.; Coombs, J.; Shaukat, N.; Olsen, D. Utility of sodium thiosulfate in acute cyanide toxicity. Ann. Emerg. Med. 2013, 61, 124–125. [Google Scholar] [CrossRef]
- Gueldener, U.; Heinisch, J.; Koehler, G.J.; Voss, D.; Hegemann, J.H. A second set of loxP marker cassettes for Cre-mediated multiple gene knockouts in budding yeast. Nucleic Acids Res. 2002, 30, e23–e30. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Chu, W.; Qi, Q.; Xun, L. New insights into the QuikChange™ process guide the use of Phusion DNA polymerase for site-directed mutagenesis. Nucleic Acids Res. 2015, 43, e12. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Li, K.; Li, J.; Wang, T.; Gu, L.; Xun, L. T5 exonuclease-dependent assembly offers a low-cost method for efficient cloning and site-directed mutagenesis. Nucleic Acids Res. 2019, 47, e15. [Google Scholar] [CrossRef] [Green Version]
- Togawa, T.; Ogawa, M.; Nawata, M.; Ogasawara, Y.; Kawanabe, K.; Tanabe, S. High performance liquid chromatographic determination of bound sulfide and sulfite and thiosulfate at their low levels in human serum by pre-column fluorescence derivatization with monobromobimane. Chem. Pharm. Bull. 1992, 40, 3000–3004. [Google Scholar] [CrossRef] [PubMed]
- Yaglom, J.; Linskens, M.; Sadis, S.; Rubin, D.M.; Futcher, B.; Finley, D. p34Cdc28-mediated control of Cln3 cyclin degradation. Mol. Cell. Biol. 1995, 15, 731–741. [Google Scholar] [CrossRef] [Green Version]
- Nash, R.; Tokiwa, G.; Anand, S.; Erickson, K.; Futcher, A. The WHI1+ gene of Saccharomyces cerevisiae tethers cell division to cell size and is a cyclin homolog. EMBO J. 1988, 7, 4335–4346. [Google Scholar] [CrossRef]
- Reers, M.; Smith, T.W.; Chen, L.B. J-aggregate formation of a carbocyanine as a quantitative fluorescent indicator of membrane potential. Biochemistry 1991, 30, 4480–4486. [Google Scholar] [CrossRef]
- Jiang, H.; Yang, Y.; Zhang, Y.; Xie, Z.; Zhao, X.; Sun, Y.; Kong, W. The dual role of poly (ADP-ribose) polymerase-1 in modulating parthanatos and autophagy under oxidative stress in rat cochlear marginal cells of the stria vascularis. Redox Biol. 2018, 14, 361–370. [Google Scholar] [CrossRef]
- Su, B.; Ji, Y.; Sun, X.; Liu, X.; Chen, Z. Brain-derived neurotrophic factor (BDNF)-induced mitochondrial motility arrest and presynaptic docking contribute to BDNF-enhanced synaptic transmission. J. Biol. Chem. 2014, 289, 1213–1226. [Google Scholar] [CrossRef] [Green Version]
- Ferdinand, W. The isolation and specific activity of rabbit-muscle glyceraldehyde phosphate dehydrogenase. Biochem. J. 1964, 92, 578–585. [Google Scholar] [CrossRef] [Green Version]
- Rolia, E.; Chakrabarti, C.L. Kinetics of decomposition of tetrathionate, trithionate, and thiosulfate in alkaline media. Environ. Sci. Technol. 1982, 16, 852–857. [Google Scholar] [CrossRef]
- Ghezzi, D.; Zeviani, M. Assembly factors of human mitochondrial respiratory chain complexes: Physiology and pathophysiology. Adv. Exp. Med. Biol. 2012, 748, 65–106. [Google Scholar]
- Breuer, P.L.; Jeffrey, M.I. The reduction of copper (II) and the oxidation of thiosulfate and oxysulfur anions in gold leaching solutions. Hydrometallurgy 2003, 70, 163–173. [Google Scholar] [CrossRef]
- Packer, L.; Mustafa, M.G. Pathways of electron flow established by tetramethyl-phenylenediamine in mitochondria and ascites tumor cells. BBA-Enzymol. Biol. Oxid. 1966, 113, 1–12. [Google Scholar] [CrossRef]
- Valli, M.; Sauer, M.; Branduardi, P. Intracellular pH Distribution in Saccharomyces cerevisiae Cell Populations, Analyzed by Flow Cytometry. Appl. Environ. Microbiol. 2005, 71, 1515–1521. [Google Scholar] [CrossRef] [Green Version]
- Mansilla, N.; Racca, S.; Gras, D.E.; Gonzalez, D.H.; Welchen, E. The Complexity of Mitochondrial Complex IV: An Update of Cytochrome c Oxidase Biogenesis in Plants. Int. J. Mol. ENCES 2018, 19, 662–695. [Google Scholar] [CrossRef] [Green Version]
- Paul, B.D.; Snyder, S.H.; Kashfi, K. Effects of hydrogen sulfide on mitochondrial function and cellular bioenergetics. Redox. Biol. 2021, 38, 101772. [Google Scholar] [CrossRef] [PubMed]
- Schimz, K.L. The effect of sulfite on the yeast Saccharomyces cerevisiae. Arch. Microbiol. 1980, 125, 89–95. [Google Scholar] [CrossRef]
- Nadai, C.; Treu, L.; Campanaro, S.; Giacomini, A.; Corich, V. Different mechanisms of resistance modulate sulfite tolerance in wine yeasts. Appl. Microbiol. Biotechnol. 2016, 100, 797–813. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.M.; Williams, J.S. Elemental sulphur as an induced antifungal substance in plant defence. J. Exp. Bot. 2004, 55, 1947–1953. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.R.; Ghosh, M.; Goswami, A.J.C.M. Inhibitory effects of sulfur nanoparticles on membrane lipids of Aspergillus niger: A novel route of fungistasis. Curr. Microbiol. 2012, 65, 91–97. [Google Scholar] [CrossRef]
- Suleiman, M.; Al-Masri, M.; Ali, A.A.; Aref, D.; Saadeddin, A.H.I.; Warad, I. Synthesis of Nano-sized Sulfur Nanoparticles and their Antibacterial Activities. J. Mater. Environ. Sci. 2015, 6, 513–518. [Google Scholar]
- Xu, Z.; Qiu, Z.; Liu, Q.; Huang, Y.; Li, D.; Shen, X.; Fan, K.; Xi, J.; Gu, Y.; Tang, Y.; et al. Converting organosulfur compounds to inorganic polysulfides against resistant bacterial infections. Nat. Commun. 2018, 9, 3713–3725. [Google Scholar] [CrossRef] [Green Version]
- Nivoliez, A. Use of Thiosulfate to Potentiate the Anti-Pathogenic Effect of Lactobacilli. U.S. Patent 15/899,273, 21 May 2015. [Google Scholar]
- Hall, A.H.; Rumack, B.H. Hydroxycobalamin/sodium thiosulfate as a cyanide antidote. J. Emerg. Med. 1987, 5, 115–121. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Relevant Characteristics |
|---|---|
| S. cerevisiae BY4742 | MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0 |
| E. coli DH5a | supE44, AlacU169, hsdR17, recA1, endA1, gyrA96, thi-1, relA1 |
| Δsul1 Δsul2 Δsoa1 | SUL1::loxP, SUL2::loxP, SOA1::BLE |
| Δrdl1 | RDL1::loxP |
| Δrdl2 | RDL2::loxP |
| Δrdl1 Δrdl2 | RDL1::loxP, RDL2::BLE |
| Δrdl1::RDL1 | RDL1::loxP, YEplac195-RDL1 |
| Δrdl1::RDL1 C98S | RDL1::loxP, YEplac195-RDL1 C98S |
| Δrdl1::RDL2 | RDL1::loxP, YEplac195-RDL2 |
| Plasmids | |
| RDL1-Yeplac195 | RDL1 in YEplac195, control by own promoter |
| RDL1-Yeplac195 C98S | RDL1 C98S in YEplac195, control by own promoter |
| RDL2-Yeplac195 | RDL2 in YEplac195, control by own promoter |
| Names | Sequences | Purpose |
|---|---|---|
| R1 ko F | ATTCTTTCTCGTTTATTTTCAGGGTTTGTGACTAAGAAACGATATTAAAGCTTCGTACGCTGCAGGTC | Knock out RDL1 |
| R1 ko R | TACTAGCTTACGAAAATACACAGGGTACATACCTAGAGTATACAAGGCCAATACGCAAACCGCCTCT | |
| R2 ko F | GCGATAACTCTCAACAAATGGAAGCGAGACAGAAGAAAAAGACCAACGCTTCGTACGCTGCAGGTC | Knock out RDL2 |
| R2 ko R | AAGGTTGTCTATATACAGGATATATCGATTATACTTGTTTCTTTTTGGCCCAATACGCAAACCGCCTCT | |
| R1 F | TATGACCATGATTACGCCATTTTATTGGCGCATAGACAAG | Overexpression of Rdl1 |
| R1 R | GTCGACCTGCAGGCATGCATGGGGTGTTCGACTAGGTT | |
| R2 F | TATGACCATGATTACGCCAGAACCATCTGAGTACTCGATT | Overexpression of Rdl2 |
| R2 R | GTCGACCTGCAGGCATGCAGAAAAAGTCTGAGAAACGTAAAGT |
| Strains | Rhodanese Activity |
|---|---|
| Wt | 100% |
| Δrdl1 | 46.6% |
| Δrdl2 | 70.3% |
| Δrdl1Δrdl2 | 26.3% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Xia, Y.; Liu, H.; Liu, H.; Xun, L. The Mechanisms of Thiosulfate Toxicity against Saccharomyces cerevisiae. Antioxidants 2021, 10, 646. https://doi.org/10.3390/antiox10050646
Chen Z, Xia Y, Liu H, Liu H, Xun L. The Mechanisms of Thiosulfate Toxicity against Saccharomyces cerevisiae. Antioxidants. 2021; 10(5):646. https://doi.org/10.3390/antiox10050646
Chicago/Turabian StyleChen, Zhigang, Yongzhen Xia, Huaiwei Liu, Honglei Liu, and Luying Xun. 2021. "The Mechanisms of Thiosulfate Toxicity against Saccharomyces cerevisiae" Antioxidants 10, no. 5: 646. https://doi.org/10.3390/antiox10050646