
Protective Effects of 6,7,4′-Trihydroxyflavanone on Hypoxia-Induced Neurotoxicity by Enhancement of HO-1 through Nrf2 Signaling Pathway

Abstract
:
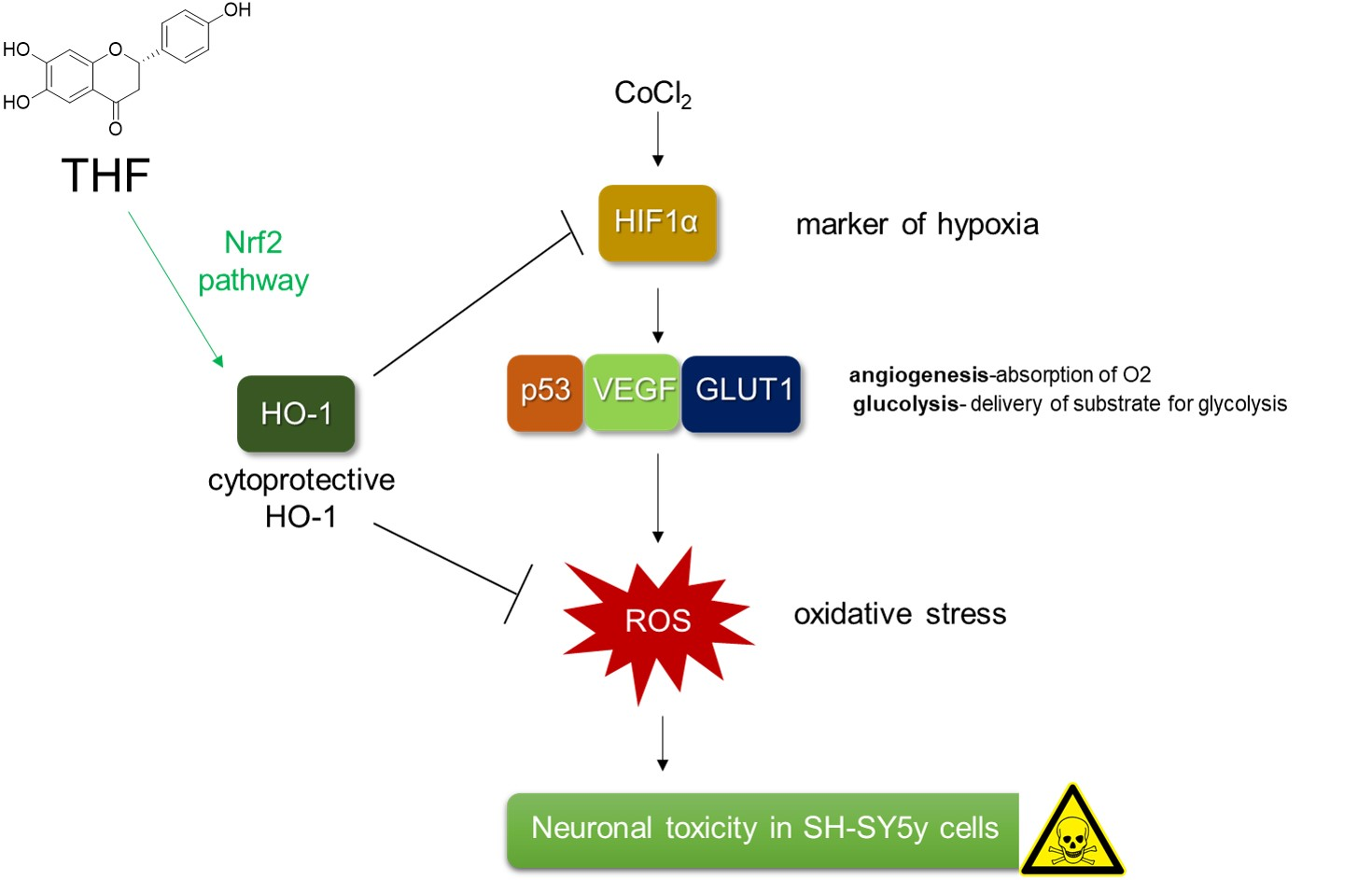{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Isolation of 6,7,4’-Tryhydroxyflavanone from Dalbergia Odorifera
2.3. Reagents and Antibodies
2.4. MTT Assay
2.5. Determination of Dead Cell Population by AnnexinV Staining
2.6. Western Blot Analysis
2.7. Apoptosis Assay
2.8. RT-PCR and Realtime Quantitative RT-PCR
2.9. Reactive Oxygen Species (ROS) Measurement
2.10. Detection of Nrf2 Nuclear Translocation
2.11. Statistics
3. Results
3.1. THF Does Not Induce Cell Death and Apoptosis in SH-SY5y Cells
3.2. THF Protects SH-SY5y Cells from CoCl2-Induced Cytotoxicity in Hypoxic Condition
3.3. THF Blocks the Cleavage of Caspase Family in CoCl2-Induced Hypoxia Condition
3.4. THF Inhibits CoCl2-Induced Hypoxia-Related Genes in SH-SY5y Cells
3.5. THF Attenuates the CoCl2-Induced Oxidative Stress in SH-SY5y Cells
3.6. THF Promotes HO-1 Expression by Leading Nrf2 Translocation in SH-SY5y Cells
3.7. Enhancement of HO-1 by THF Pre-treatment Protects SH-SY5y Cells from CoCl2-Induced Neurotoxicity in Hypoxic Condition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martini, S.; Austin, T.; Aceti, A.; Faldella, G.; Corvaglia, L. Free radicals and neonatal encephalopathy: Mechanisms of injury, biomarkers, and antioxidant treatment perspectives. Pediatr. Res. 2020, 87, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Ratan, R.R.; Siddiq, A.; Smirnova, N.; Karpisheva, K.; Haskew-Layton, R.; McConoughey, S.; Langley, B.; Estevez, A.; Huerta, P.T.; Volpe, B.; et al. Harnessing hypoxic adaptation to prevent, treat, and repair stroke. J. Mol. Med. 2007, 85, 1331–1338. [Google Scholar] [CrossRef] [Green Version]
- Yagishita, S.; Hirasawa, A. Intermittent hypoxia produces Alzheimer disease? Oncotarget 2017, 8, 41786–41787. [Google Scholar] [CrossRef]
- Snyder, B.; Shell, B.; Cunningham, J.T.; Cunningham, R.L. Chronic intermittent hypoxia induces oxidative stress and inflammation in brain regions associated with early-stage neurodegeneration. Physiol. Rep. 2017, 5. [Google Scholar] [CrossRef]
- Wiener, C.M.; Booth, G.; Semenza, G.L. In vivo expression of mRNAs encoding hypoxia-inducible factor 1. Biochem. Biophys. Res. Commun. 1996, 225, 485–488. [Google Scholar] [CrossRef]
- Jin, K.L.; Mao, X.O.; Nagayama, T.; Goldsmith, P.C.; Greenberg, D.A. Induction of vascular endothelial growth factor and hypoxia-inducible factor-1α by global ischemia in rat brain. Neuroscience 2000, 99, 577–585. [Google Scholar] [CrossRef]
- Koumenis, C.; Alarcon, R.; Hammond, E.; Sutphin, P.; Hoffman, W.; Murphy, M.; Derr, J.; Taya, Y.; Lowe, S.W.; Kastan, M.; et al. Regulation of p53 by Hypoxia: Dissociation of Transcriptional Repression and Apoptosis from p53-Dependent Transactivation. Mol. Cell. Biol. 2001, 21, 1297–1310. [Google Scholar] [CrossRef] [Green Version]
- Krock, B.L.; Skuli, N.; Simon, M.C. Hypoxia-Induced Angiogenesis: Good and Evil. Genes Cancer 2011, 2, 1117–1133. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, M.; Sakata, M.; Takeda, T.; Yamamoto, T.; Okamoto, Y.; Sawada, K.; Kimura, A.; Minekawa, R.; Tahara, M.; Tasaka, K.; et al. Induction of glucose transporter 1 expression through hypoxia-inducible factor 1α under hypoxic conditions in trophoblast-derived cells. J. Endocrinol. 2004, 183, 145–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndisang, J.F. Synergistic Interaction Between Heme Oxygenase (HO) and Nuclear-Factor E2- Related Factor-2 (Nrf2) against Oxidative Stress in Cardiovascular Related Diseases. Curr. Pharm. Des. 2017, 23, 1465–1470. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Park, J.S.; Lee, Y.S.; Sung, S.H.; Lee, Y.H.; Bae, S.H. The hypertension drug, verapamil, activates Nrf2 by promoting p62-dependent autophagic Keap1 degradation and prevents acetaminophen-induced cytotoxicity. BMB Rep. 2017, 50, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Cheng, H.; Wang, L.; Fu, J.; Zhang, G.; Guan, D.; Qi, R.; Gao, X.; Zhao, R. Protective roles of NRF2 signaling pathway in cobalt chloride-induced hypoxic cytotoxicity in human HaCaT keratinocytes. Toxicol. Appl. Pharmacol. 2018, 355, 189–197. [Google Scholar] [CrossRef]
- Wang, W.; Weng, X.; Cheng, D. Antioxidant activities of natural phenolic components from Dalbergia odorifera T. Chen. Food Chem. 2000, 71, 45–49. [Google Scholar] [CrossRef]
- Lee, D.S.; Kim, K.S.; Ko, W.; Li, B.; Keo, S.; Jeong, G.S.; Oh, H.; Kim, Y.C. The neoflavonoid latifolin isolated from meoh extract of Dalbergia odorifera attenuates inflammatory responses by inhibiting NF-κB activation via Nrf2-mediated heme oxygenase-1 expression. Phyther. Res. 2014, 28, 1216–1223. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.N.; Kim, Y.G.; Lee, J.H.; Min, B.S.; Jeong, G.S. 6,7,4′-Trihydroxyflavone inhibits osteoclast formation and bone resorption in vitro and in vivo. Phyther. Res. 2019, 33, 2948–2959. [Google Scholar] [CrossRef]
- Lee, H.S.; Jeong, G.S. 6,7,4′ -Trihydroxyflavanone Prevents Methamphetamine-Induced T Cell Deactivation by Protecting the Activated T Cells from Apoptosis. Am. J. Chin. Med. 2021. [Google Scholar] [CrossRef] [PubMed]
- Cavia-Saiz, M.; Busto, M.D.; Pilar-Izquierdo, M.C.; Ortega, N.; Perez-Mateos, M.; Muñiz, P. Antioxidant properties, radical scavenging activity and biomolecule protection capacity of flavonoid naringenin and its glycoside naringin: A comparative study. J. Sci. Food Agric. 2010, 90, 1238–1244. [Google Scholar] [CrossRef] [PubMed]
- Rashmi, R.; Bojan Magesh, S.; Mohanram Ramkumar, K.; Suryanarayanan, S.; Venkata SubbaRao, M. Antioxidant Potential of Naringenin Helps to Protect Liver Tissue from Streptozotocin-Induced Damage. Rep. Biochem. Mol. Biol. 2018, 7, 76–84. [Google Scholar] [PubMed]
- Shen, C.C.; Cheng, J.J.; Lay, H.L.; Wu, S.Y.; Ni, C.L.; Teng, C.M.; Chen, C.C. Cytotoxic apigenin derivatives from Chrysopogon aciculatis. J. Nat. Prod. 2012, 75, 198–201. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Li, W.; Yin, H.; Cheong, Y.K.; Ren, G.; Yang, Z. Pretreatment-Etidronate Alleviates CoCl 2 Induced-SH-SY5Y Cell Apoptosis via Decreased HIF-1α and TRPC5 Channel Proteins. Neurochem. Res. 2019, 44, 428–440. [Google Scholar] [CrossRef]
- Kurokawa, M.; Kornbluth, S. Caspases and kinases in a death grip. Cell 2009, 138, 838–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGarry, T.; Biniecka, M.; Veale, D.J.; Fearon, U. Hypoxia, oxidative stress and inflammation. Free Radic. Biol. Med. 2018, 125, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Lu, F.; Latham, C.; Zander, D.S.; Visner, G.A. Heme oxygenase-1 expression in human lungs with cystic fibrosis and cytoprotective effects against Pseudomonas aeruginosa in vitro. Am. J. Respir. Crit. Care Med. 2004, 170, 633–640. [Google Scholar] [CrossRef] [Green Version]
- Yuan, F.; Chen, Y.; Dellian, M.; Safabakhsh, N.; Ferrara, N.; Jain, R.K. Time-dependent vascular regression and permeability changes in established human tumor xenografts induced by an anti-vascular endothelial growth factor/vascular permeability factor antibody. Proc. Natl. Acad. Sci. USA 1996, 93, 14765–14770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plate, K.H.; Breier, G.; Weich, H.A.; Risau, W. Vascular endothelial growth factor is a potential tumour angiogenesis factor in human gliomas in vivo. Nature 1992, 359, 845–848. [Google Scholar] [CrossRef]
- Iyer, N.V.; Kotch, L.E.; Agani, F.; Leung, S.W.; Laughner, E.; Wenger, R.H.; Gassmann, M.; Gearhart, J.D.; Lawler, A.M.; Yu, A.Y.; et al. Cellular and developmental control of O2 homeostasis by hypoxia- inducible factor 1α. Genes Dev. 1998, 12, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, Y.; Kitamura, M. Anti-apoptotic effect of quercetin: Intervention in the JNK- and ERK-mediated apoptotic pathways. Kidney Int. 2000, 58, 1078–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vauzour, D.; Vafeiadou, K.; Rice-Evans, C.; Williams, R.J.; Spencer, J.P.E. Activation of pro-survival Akt and ERK1/2 signalling pathways underlie the anti-apoptotic effects of flavanones in cortical neurons. J. Neurochem. 2007, 103, 1355–1367. [Google Scholar] [CrossRef]
- Wang, K.; Chen, Z.; Huang, J.; Huang, L.; Luo, N.; Liang, X.; Liang, M.; Xie, W. Naringenin prevents ischaemic stroke damage via anti-apoptotic and anti-oxidant effects. Clin. Exp. Pharmacol. Physiol. 2017, 44, 862–871. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-S.; Jeong, G.-S. Protective Effects of 6,7,4′-Trihydroxyflavanone on Hypoxia-Induced Neurotoxicity by Enhancement of HO-1 through Nrf2 Signaling Pathway. Antioxidants 2021, 10, 341. https://doi.org/10.3390/antiox10030341
Lee H-S, Jeong G-S. Protective Effects of 6,7,4′-Trihydroxyflavanone on Hypoxia-Induced Neurotoxicity by Enhancement of HO-1 through Nrf2 Signaling Pathway. Antioxidants. 2021; 10(3):341. https://doi.org/10.3390/antiox10030341
Chicago/Turabian StyleLee, Hyun-Su, and Gil-Saeng Jeong. 2021. "Protective Effects of 6,7,4′-Trihydroxyflavanone on Hypoxia-Induced Neurotoxicity by Enhancement of HO-1 through Nrf2 Signaling Pathway" Antioxidants 10, no. 3: 341. https://doi.org/10.3390/antiox10030341