
Human Placental NADPH Oxidase Mediates sFlt-1 and PlGF Secretion in Early Pregnancy: Exploration of the TGF-β1/p38 MAPK Pathways

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Materials
2.3. Placenta Retrieval, Chorionic Villi Storage and Culture
2.4. TGF-β1 and SB203580 Treatments
2.5. Immunostaining
2.6. Western Blot Analysis
2.7. Measurement of NADPH Oxidase Activity
2.8. Measurement of sFlt-1 Secretion
2.9. Statistical Analysis
3. Results
3.1. Studied Population
3.2. NADPH Oxidase Activity Modulation Using TGF-β1
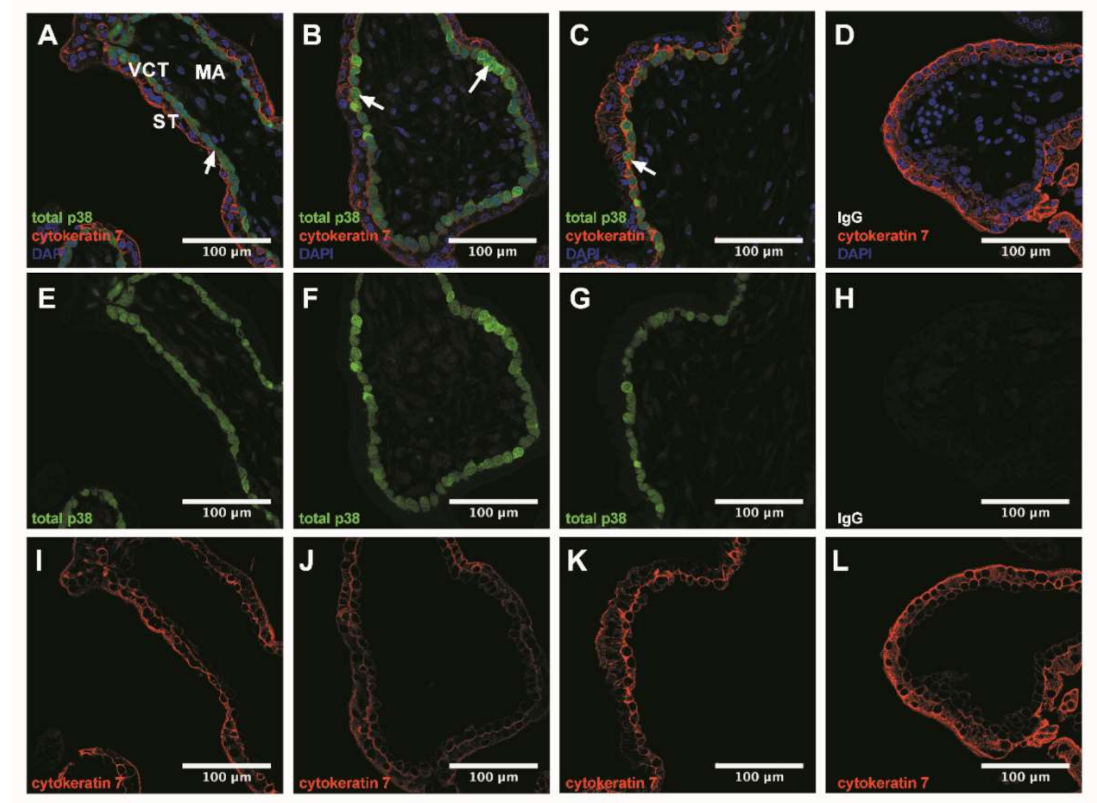
3.3. p38 Protein Is Mostly Expressed in Villous Cytotrophoblast
3.4. TGF-β1-Mediated p38 MAPK Activation Can Be Prompt and Long-Term Phenomenon
3.5. TGF-β1 Canonical Pathway Study: Involvement of SMAD2
3.6. Implication of p38 MAPK Pathway and NADPH Oxidase on sFlt-1 Secretion and Angiogenic Balance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Barker, D.J.P. The developmental origins of well–being. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 1359–1366. [Google Scholar] [CrossRef]
- Barker, D.J.P. Adult consequences of fetal growth restriction. Clin. Obstet. Gynecol. 2006, 49, 270–283. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W. Latest advances in understanding preeclampsia. Science 2005, 308, 1592–1594. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Lemoine, E.; Granger, J.P.; Karumanchi, S.A. Preeclampsia. Circ. Res. 2019, 124, 1094–1112. [Google Scholar] [CrossRef]
- US Preventive Services Task Force; Bibbins-Domingo, K.; Grossman, D.C.; Curry, S.J.; Barry, M.J.; Davidson, K.W.; Doubeni, C.A.; Epling, J.W.; Kemper, A.R.; Krist, A.H.; et al. Screening for preeclampsia. JAMA 2017, 317, 1661–1667. [Google Scholar] [CrossRef]
- Sovio, U.; Gaccioli, F.; Cook, E.; Hund, M.; Charnock-Jones, D.S.; Smith, G.C. Prediction of preeclampsia using the soluble fms-like tyrosine kinase 1 to placental growth factor ratio. Hypertension 2017, 69, 731–738. [Google Scholar] [CrossRef]
- Stepan, H.; Hund, M.; Andraczek, T. Combining biomarkers to predict pregnancy complications and redefine preeclampsia. Hypertension 2020, 75, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Dröge, L.A.; Perschel, F.H.; Stütz, N.; Gafron, A.; Frank, L.; Busjahn, A.; Henrich, W.; Verlohren, S. Prediction of preeclampsia-related adverse outcomes with the sflt-1 (Soluble fms-Like Tyrosine Kinase 1)/PlGF (placental growth factor)-ratio in the clinical routine. Hypertension 2021, 77, 461–471. [Google Scholar] [CrossRef]
- Maynard, S.; Epstein, F.H.; Karumanchi, S.A. Preeclampsia and Angiogenic imbalance. Annu. Rev. Med. 2008, 59, 61–78. [Google Scholar] [CrossRef]
- Tsatsaris, V.; Goffin, F.; Munaut, C.; Brichant, J.-F.; Pignon, M.-R.; Noel, A.; Schaaps, J.-P.; Cabrol, D.; Frankenne, F.; Foidart, J.-M. Overexpression of the soluble vascular endothelial growth factor receptor in preeclamptic patients: Pathophysiological consequences. J. Clin. Endocrinol. Metab. 2003, 88, 5555–5563. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Zhu, F.; Ding, Y. Serum screening in first trimester to predict pre-eclampsia, small for gestational age and preterm delivery: Systematic review and meta-analysis. BMC Pregnancy Childbirth 2015, 15, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, F.P.; Ryan, R.M.; Chappell, L.C. Prospective biomarkers in preterm preeclampsia: A review. Pregnancy Hypertens. 2018, 14, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Zeisler, H.; Llurba, E.; Chantraine, F.; Vatish, M.; Staff, A.C.; Sennström, M.; Olovsson, M.; Brennecke, S.P.; Stepan, H.; Allegranza, D.; et al. Predictive Value of the sFlt-1:PlGF ratio in women with suspected preeclampsia. N. Engl. J. Med. 2016, 374, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Djurovic, S.; Schjetlein, R.; Wisløff, F.; Haugen, G.; Husby, H.; Berg, K. Plasma concentrations of Lp(a) lipoprotein and TGF-β1 are altered in preeclampsia. Clin. Genet. 2008, 52, 371–376. [Google Scholar] [CrossRef]
- Chaudhury, A.; Howe, P.H. The tale of transforming growth factor-beta (TGFβ) signaling: A soigné enigma. IUBMB Life 2009, 61, 929–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massagué, J. TGF-β signal transduction. Annu. Rev. Biochem. 1998, 67, 753–791. [Google Scholar] [CrossRef] [PubMed]
- Lash, G.E.; Otun, H.A.; Innes, B.A.; Bulmer, J.N.; Searle, R.F.; Robson, S.C. Inhibition of trophoblast cell invasion by TGFB1, 2, and 3 is associated with a decrease in active proteases. Biol. Reprod. 2005, 73, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Sivasubramaniyam, T.; Yinon, Y.; Tagliaferro, A.; Ray, J.; Nevo, O.; Post, M.; Caniggia, I. Aberrant TGFβ Signaling contributes to altered trophoblast differentiation in preeclampsia. Endocrinology 2016, 157, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Thannickal, V.J.; Fanburg, B.L. Activation of an H2O2-generating NADH oxidase in human lung fibroblasts by transforming growth factor β1. J. Biol. Chem. 1995, 270, 30334–30338. [Google Scholar] [CrossRef] [Green Version]
- Herrera, B.; Murillo, M.M.; Álvarez-Barrientos, A.; Beltrán, J.; Fernández, M.; Fabregat, I. Source of early reactive oxygen species in the apoptosis induced by transforming growth factor-β in fetal rat hepatocytes. Free. Radic. Biol. Med. 2004, 36, 16–26. [Google Scholar] [CrossRef]
- Hernandez, I.; Fournier, T.; Chissey, A.; Therond, P.; Slama, A.; Beaudeux, J.-L.; Zerrad-Saadi, A. NADPH oxidase is the major source of placental superoxide in early pregnancy: Association with MAPK pathway activation. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Son, Y.; Cheong, Y.-K.; Kim, N.-H.; Chung, H.-T.; Kang, D.G.; Pae, H.-O. Mitogen-activated protein kinases and reactive oxygen species: How can ros activate mapk pathways? J. Signal Transduct. 2011, 2011, 1–6. [Google Scholar] [CrossRef]
- Yurtsever, Z.; Scheaffer, S.M.; Romero, A.G.; Holtzman, M.J.; Brett, T.J. The crystal structure of phosphorylated MAPK13 reveals common structural features and differences in p38 MAPK family activation. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 790–799. [Google Scholar] [CrossRef] [PubMed]
- Brancho, D.M.; Tanaka, N.; Jaeschke, A.; Ventura, J.-J.; Kelkar, N.; Tanaka, Y.; Kyuuma, M.; Takeshita, T.; Flavell, R.A.; Davis, R.J. Mechanism of p38 MAP kinase activation in vivo. Genes Dev. 2003, 17, 1969–1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuenda, A.; Rousseau, S. P38 MAP-Kinases pathway regulation, function and role in human diseases. Biochim. Biophys. Acta Bioenerg. 2007, 1773, 1358–1375. [Google Scholar] [CrossRef] [Green Version]
- Gong, X.; Ming, X.; Deng, P.; Jiang, Y. Mechanisms regulating the nuclear translocation of p38 MAP kinase. J. Cell. Biochem. 2010, 110, 1420–1429. [Google Scholar] [CrossRef]
- Han, J.; Lee, J.D.; Bibbs, L.; Ulevitch, R.J. A map kinase targeted by endotoxin and hyperosmolarity in mammalian cells. Science 1994, 265, 808–811. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Deng, Q.; Luo, X.; Chen, Y.; Shan, N.; QinYin, D. Oxidative stress-induced Gadd45α inhibits trophoblast invasion and increases sFlt1/sEng secretions via p38 MAPK involving in the pathology of pre-eclampsia. J. Matern. Neonatal Med. 2016, 29, 3776–3785. [Google Scholar] [CrossRef]
- Szabó, S.C.; Mody, M.; Romero, R.; Xu, Y.; Karaszi, K.; Mihalik, N.; Xu, Z.; Bhatti, G.; Fule, T.; Hupuczi, P.; et al. Activation of villous trophoblastic p38 and ERK1/2 signaling pathways in preterm preeclampsia and HELLP syndrome. Pathol. Oncol. Res. 2015, 21, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Daoud, G.; Amyot, M.; Rassart, E.; Masse, A.; Simoneau, L.; Lafond, J. ERK1/2 and p38 regulate trophoblasts differentiation in human term placenta. J. Physiol. 2005, 566, 409–423. [Google Scholar] [CrossRef]
- Raijmakers, M.; Burton, G.; Jauniaux, E.; Seed, P.; Peters, W.; Steegers, E.; Poston, L. Placental NAD(P)H oxidase mediated superoxide generation in early pregnancy. Placenta 2006, 27, 158–163. [Google Scholar] [CrossRef]
- Dechend, R.; Viedt, C.; Müller, D.N.; Ugele, B.; Brandes, R.P.; Wallukat, G.; Park, J.-K.; Janke, J.; Barta, P.; Theuer, J.; et al. AT 1 receptor agonistic antibodies from preeclamptic patients stimulate NADPH oxidase. Circulation 2003, 107, 1632–1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raijmakers, M.; Peters, W.; Steegers, E.; Poston, L. NAD(P)H oxidase associated superoxide production in human placenta from normotensive and pre-eclamptic women. Placenta 2004, 25, S85–S89. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Wei, S.-J.; Zhang, D.; Hu, X.; Xu, Z.; Wilson, B.; El-Benna, J.; Hong, J.-S.; Flood, P.M. Potent anti-inflammatory and neuroprotective effects of TGF-β1 are mediated through the inhibition of ERK and p47phox-Ser345 Phosphorylation and translocation in microglia. J. Immunol. 2008, 181, 660–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchmaier, C.M.; Busse, R.; Schini-Kerth, V.R.B. Oxidative stress and expression of P22phox are involved in the up-regulation of tissue factor in vascular smooth muscle cells in response to activated platelets. FASEB J. 2000, 14, 1518–1528. [Google Scholar]
- Peraçoli, M.T.S.; Menegon, F.T.F.; Borges, V.T.M.; de Araujo Costa, R.A.; Thomazini-Santos, I.A.; Peraçoli, J.C. Platelet aggregation and TGF-beta1 plasma levels in pregnant women with preeclampsia. J. Reprod. Immunol. 2008, 79, 79–84. [Google Scholar] [CrossRef]
- Olieslagers, S.; Pardali, E.; Tchaikovski, V.; Dijke, P.T.; Waltenberger, J. TGF-β1/ALK5-induced monocyte migration involves PI3K and p38 pathways and is not negatively affected by diabetes mellitus. Cardiovasc. Res. 2011, 91, 510–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashdan, N.A.; Lloyd, P.G. Fluid shear stress upregulates placental growth factor in the vessel wall via NADPH oxidase 4. Am. J. Physiol. Circ. Physiol. 2015, 309, H1655–H1666. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Chen, L.; Liu, B.; Vialli, C.; Stone, P.; Ching, L.-M.; Chamley, L. The role of autocrine TGFβ1 in endothelial cell activation induced by phagocytosis of necrotic trophoblasts: A possible role in the pathogenesis of pre-eclampsia. J. Pathol. 2010, 221, 87–95. [Google Scholar] [CrossRef]
- Oh, M.-K.; Kim, I.-S. Involvement of placental growth factor upregulated via TGF-β1-ALK1-Smad1/5 signaling in prohaptoglobin-induced angiogenesis. PLoS ONE 2019, 14, e0216289. [Google Scholar] [CrossRef] [Green Version]
- Haider, S.; Kunihs, V.; Fiala, C.; Pollheimer, J.; Knöfler, M. Expression pattern and phosphorylation status of Smad2/3 in different subtypes of human first trimester trophoblast. Placenta 2017, 57, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Nevo, O.; Soleymanlou, N.; Wu, Y.; Xu, J.; Kingdom, J.; Many, A.; Zamudio, S.; Caniggia, I. Increased expression of sFlt-1 in in vivo and in vitro models of human placental hypoxia is mediated by HIF-1. Am. J. Physiol. Integr. Comp. Physiol. 2006, 291, R1085–R1093. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Shu, C.; Liu, Z.; Tong, W.; Cui, M.; Wei, C.; Tang, J.J.; Liu, X.; Hu, J.; Jiang, J.; et al. The expression of serum sEGFR, sFlt-1, sEndoglin and PLGF in preeclampsia. Pregnancy Hypertens. 2018, 13, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Fiers, W.; Beyaert, R.; Boone, E.; Cornelis, S.; Declercq, W.; DeCoster, E.; Denecker, G.; Depuydt, B.; De Valck, D.; De Wilde, G.; et al. TNF-induced intracellular signaling leading to gene induction or to cytotoxicity by necrosis or by apoptosis. J. Inflamm. 1995, 47, 67–75. [Google Scholar] [PubMed]
- Nakashima, A.; Yamanaka-Tatematsu, M.; Fujita, N.; Koizumi, K.; Shima, T.; Yoshida, T.; Nikaido, T.; Okamoto, A.; Yoshimori, T.; Saito, S. Impaired autophagy by soluble endoglin, under physiological hypoxia in early pregnant period, is involved in poor placentation in preeclampsia. Autophagy 2013, 9, 303–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehr, R.N.; Kheirollah, A.; Seif, F.; Dayati, P.; Babaahmadi-Rezaei, H. Reactive oxygen species and p38MAPK have a role in the smad2 linker region phosphorylation induced by TGF-β. Iran. J. Med Sci. 2018, 43, 401–408. [Google Scholar]
- Matsuzaki, K. Smad phospho-isoforms direct context-dependent TGF-β signaling. Cytokine Growth Factor Rev. 2013, 24, 385–399. [Google Scholar] [CrossRef]
- Sorrentino, A.; Thakur, N.; Grimsby, S.; Marcusson, A.; Von Bulow, V.; Schuster, N.; Zhang, S.; Heldin, C.-H.; Landström, M. The type I TGF-β receptor engages TRAF6 to activate TAK1 in a receptor kinase-independent manner. Nat. Cell Biol. 2008, 10, 1199–1207. [Google Scholar] [CrossRef]
- Yamashita, M.; Fatyol, K.; Jin, C.; Wang, X.; Liu, Z.; Zhang, Y.E. TRAF6 mediates smad-independent activation of JNK and p38 by TGF-β. Mol. Cell 2008, 31, 918–924. [Google Scholar] [CrossRef] [Green Version]
- Ge, B.; Gram, H.; Di Padova, F.; Huang, B.; New, L.; Ulevitch, R.J.; Luo, Y.; Han, J. MAPKK-independent activation of p38alpha mediated by TAB1-dependent autophosphorylation of p38alpha. Science 2002, 295, 1291–1294. [Google Scholar] [CrossRef] [PubMed]
- Ota, T.; Fujii, M.; Sugizaki, T.; Ishii, M.; Miyazawa, K.; Aburatani, H.; Miyazono, K. Targets of transcriptional regulation by two distinct type I receptors for transforming growth factor-? in human umbilical vein endothelial cells. J. Cell. Physiol. 2002, 193, 299–318. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernandez, I.; Chissey, A.; Guibourdenche, J.; Atasoy, R.; Coumoul, X.; Fournier, T.; Beaudeux, J.-L.; Zerrad-Saadi, A. Human Placental NADPH Oxidase Mediates sFlt-1 and PlGF Secretion in Early Pregnancy: Exploration of the TGF-β1/p38 MAPK Pathways. Antioxidants 2021, 10, 281. https://doi.org/10.3390/antiox10020281
Hernandez I, Chissey A, Guibourdenche J, Atasoy R, Coumoul X, Fournier T, Beaudeux J-L, Zerrad-Saadi A. Human Placental NADPH Oxidase Mediates sFlt-1 and PlGF Secretion in Early Pregnancy: Exploration of the TGF-β1/p38 MAPK Pathways. Antioxidants. 2021; 10(2):281. https://doi.org/10.3390/antiox10020281
Chicago/Turabian StyleHernandez, Isabelle, Audrey Chissey, Jean Guibourdenche, Roger Atasoy, Xavier Coumoul, Thierry Fournier, Jean-Louis Beaudeux, and Amal Zerrad-Saadi. 2021. "Human Placental NADPH Oxidase Mediates sFlt-1 and PlGF Secretion in Early Pregnancy: Exploration of the TGF-β1/p38 MAPK Pathways" Antioxidants 10, no. 2: 281. https://doi.org/10.3390/antiox10020281