
Protective Effects of Kirenol against Lipopolysaccharide-Induced Acute Lung Injury through the Modulation of the Proinflammatory NFκB Pathway and the AMPK2-/Nrf2-Mediated HO-1/AOE Pathway

 , , and
, , and 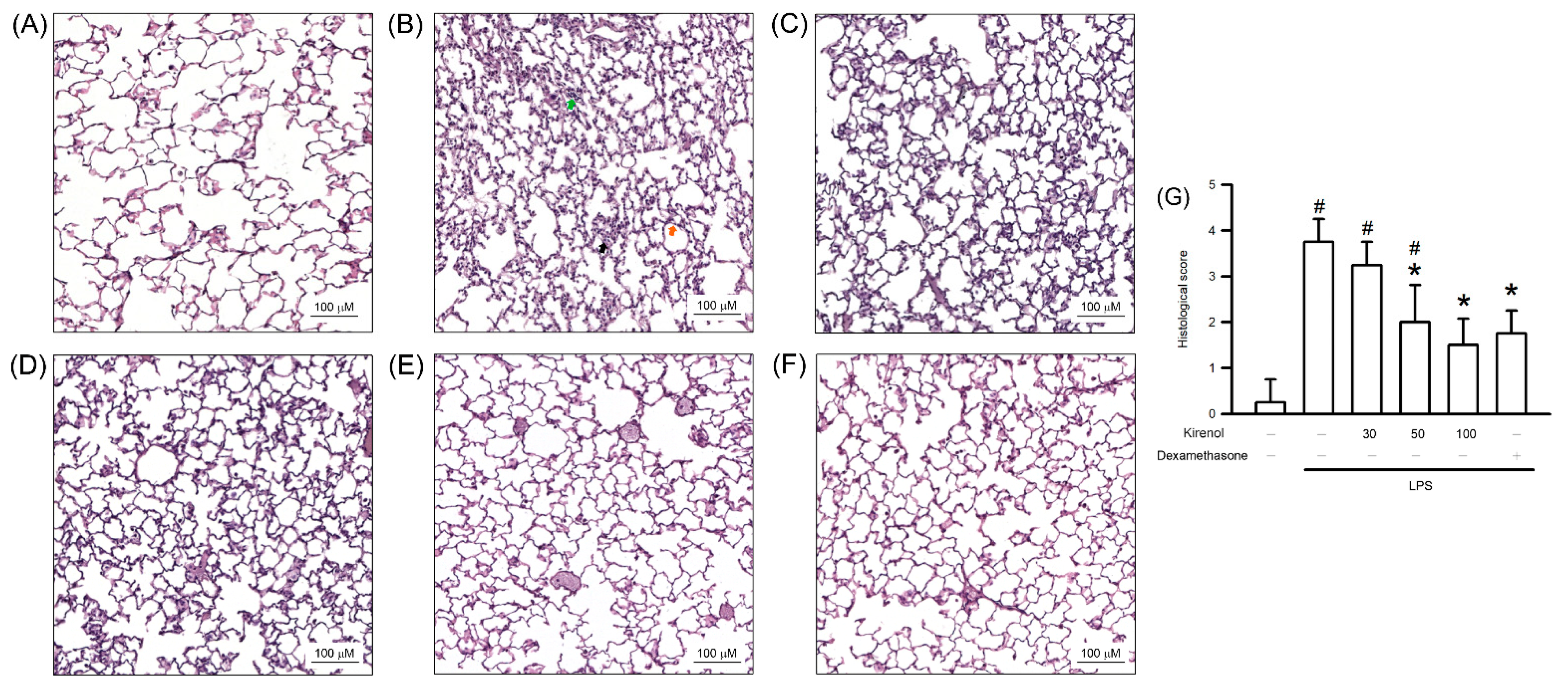{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animal Model of ALI
2.3. Histopathological Study
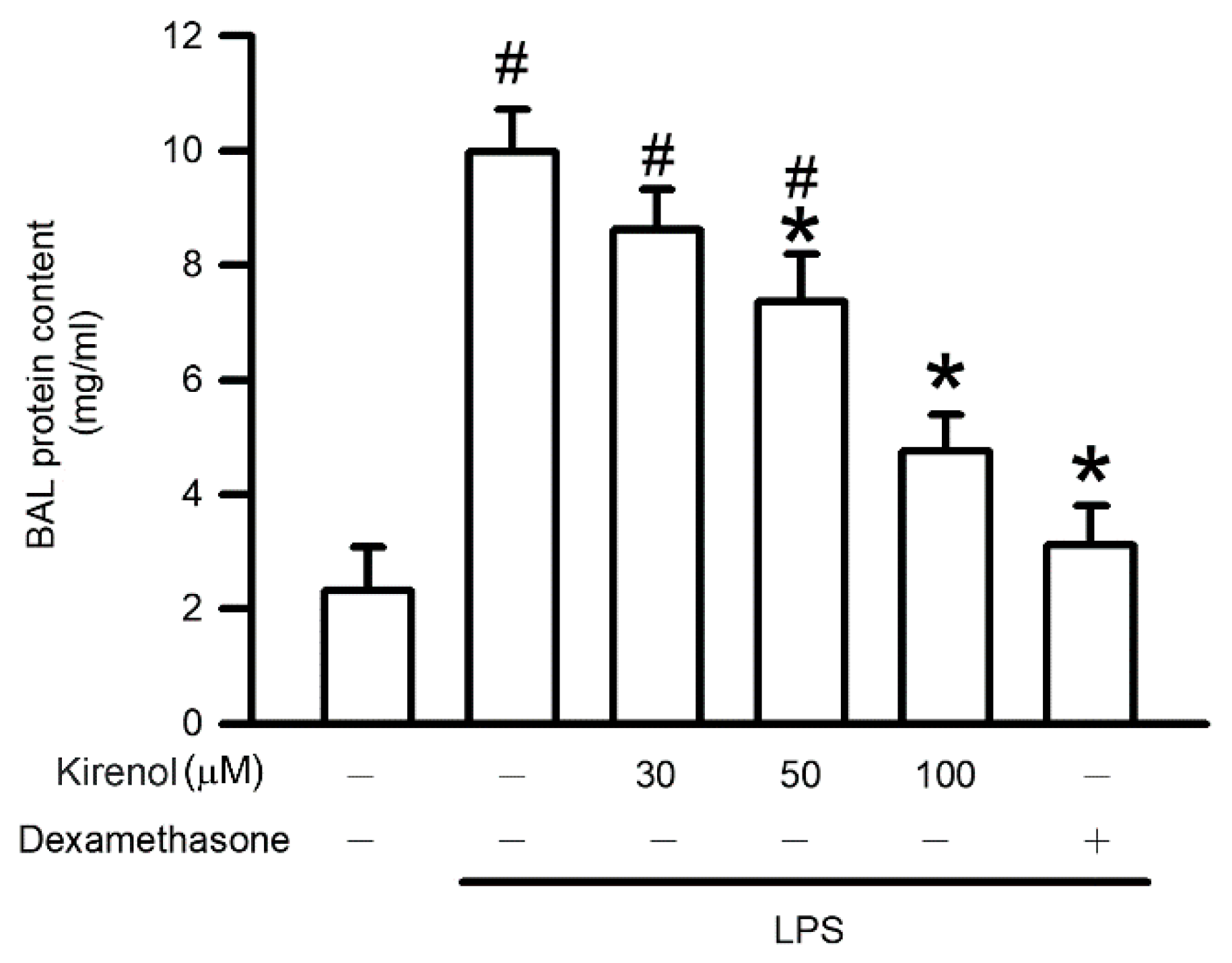
2.4. Bronchoalveolar Lavage Fluid Collection
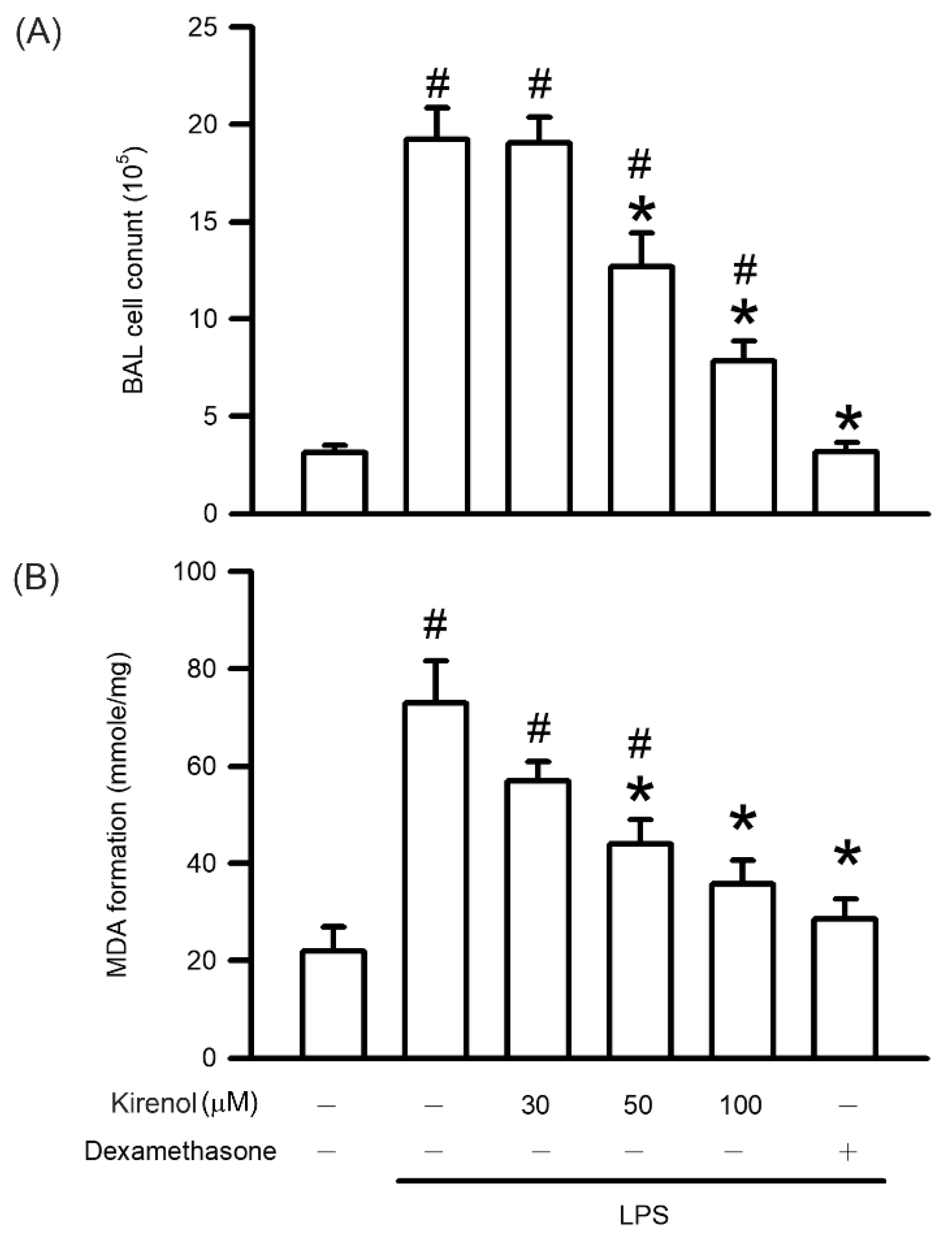
2.5. Thiobarbituric Acid Reactive Substances Assay
2.6. Antioxidant Enzyme Capability and Cytokine Generation Assay
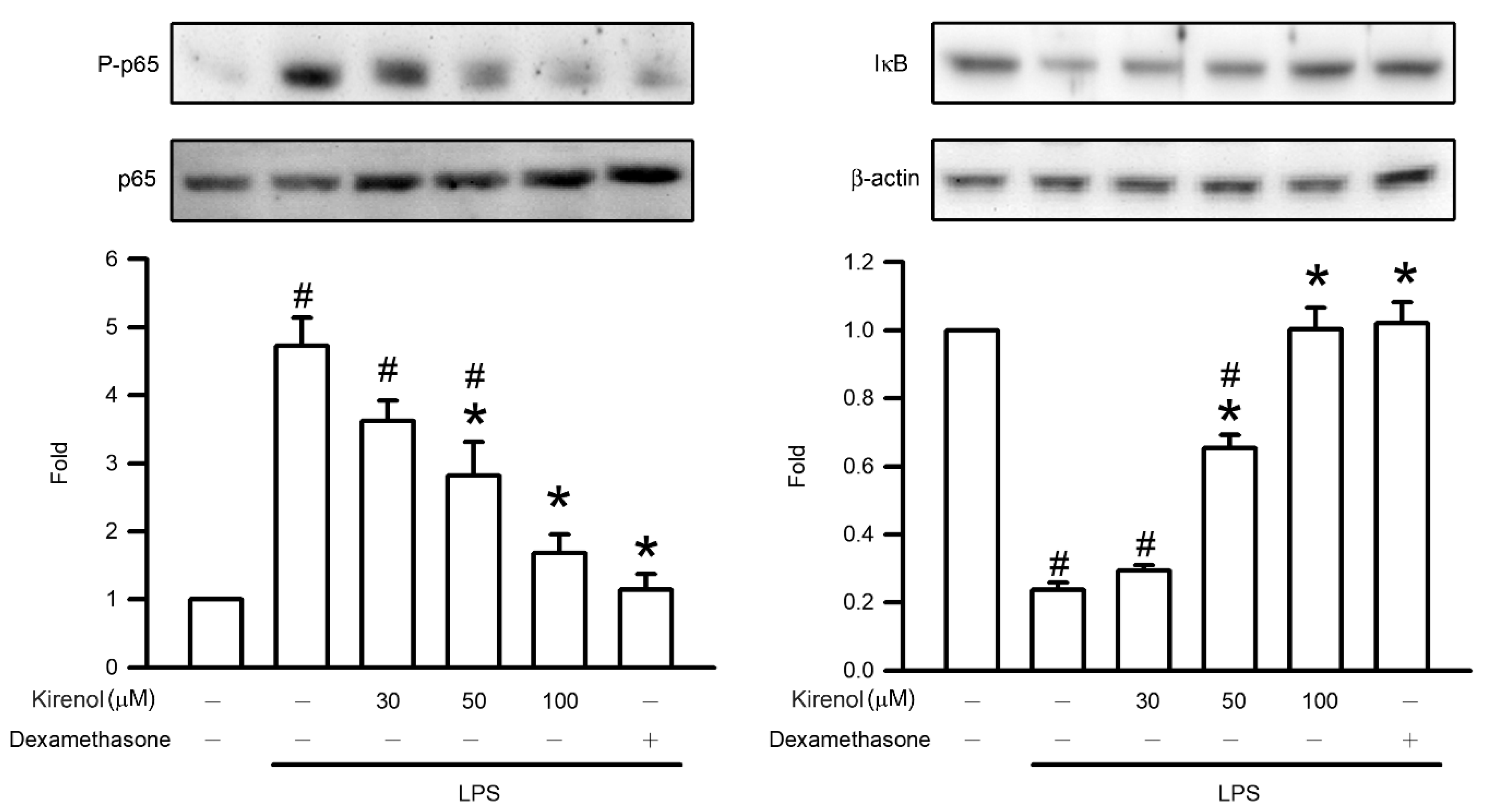
2.7. Western Blot Assay
2.8. Statistical Analysis
3. Results
3.1. Kirenol Protected the Lung Histopathological Changes in Mice with LPS-Induced ALI
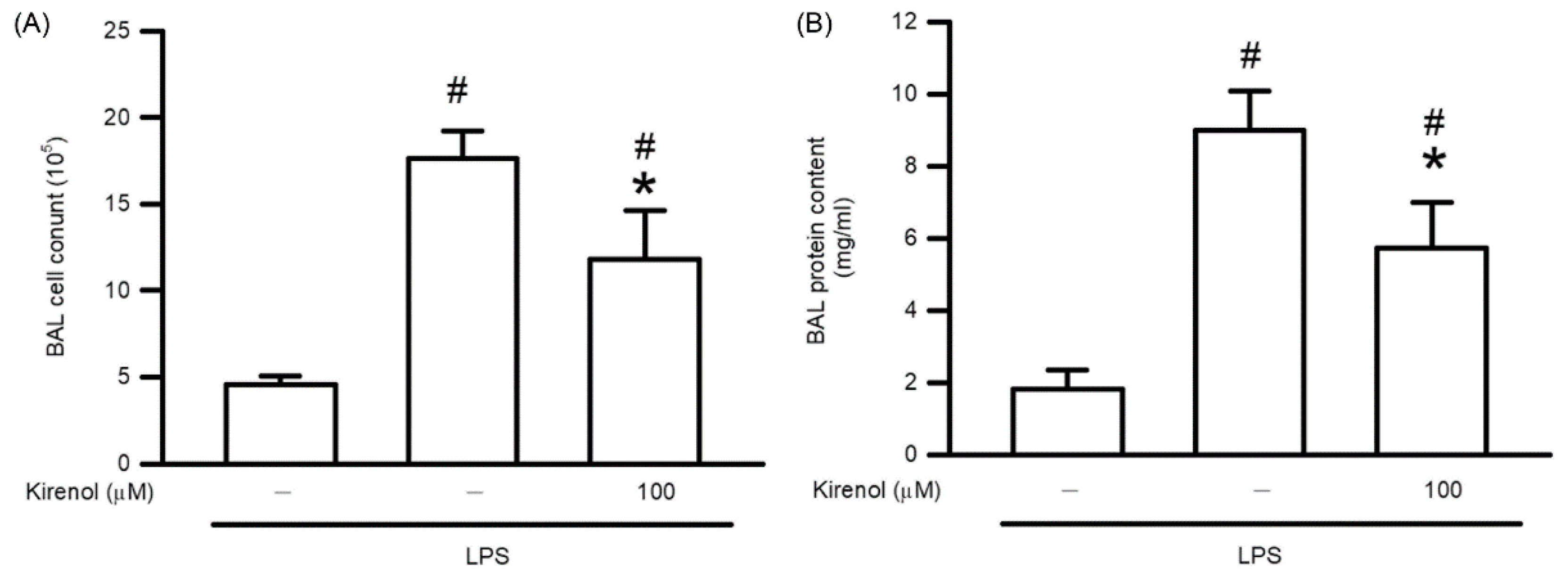
3.2. Kirenol Protected the Alveolar-Capillary Barrier Disruption in Mice with LPS-Induced ALI
3.3. Kirenol Protected the Leukocyte Infiltration and Lipid Peroxidation in Mice with LPS-Induced ALI
3.4. Kirenol Protected the NF-κB p65 Phosphorylation and of IκB Degradation in Mice with LPS-Induced ALI
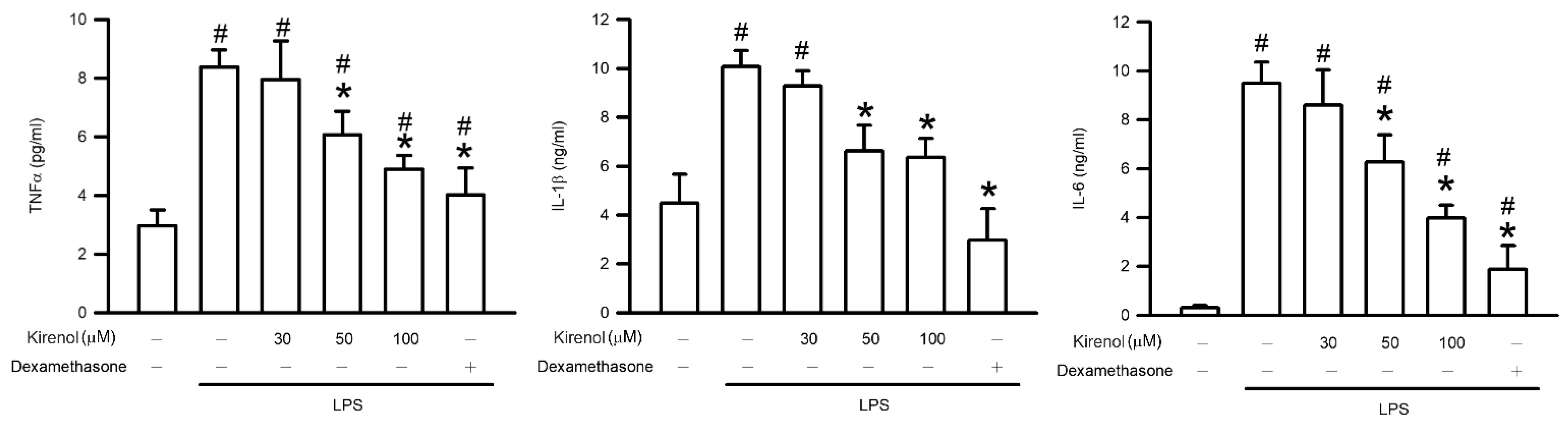
3.5. Kirenol Protected the Production of Proinflammatory Cytokines in Mice with LPS-Induced ALI
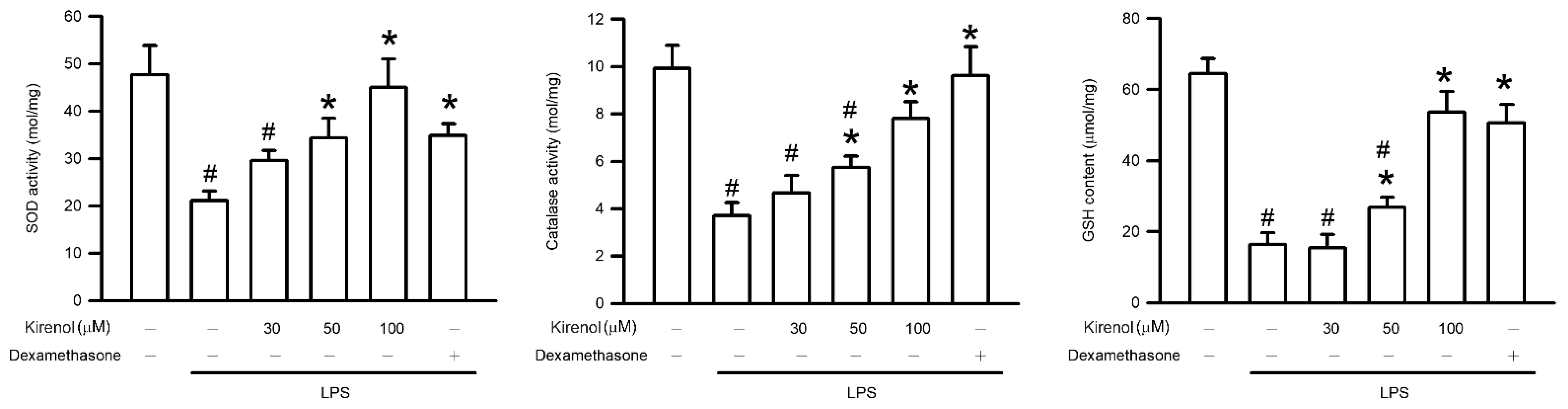
3.6. Kirenol Attenuated the Downregulation of AOEs Activities in Mice with LPS-Induced ALI
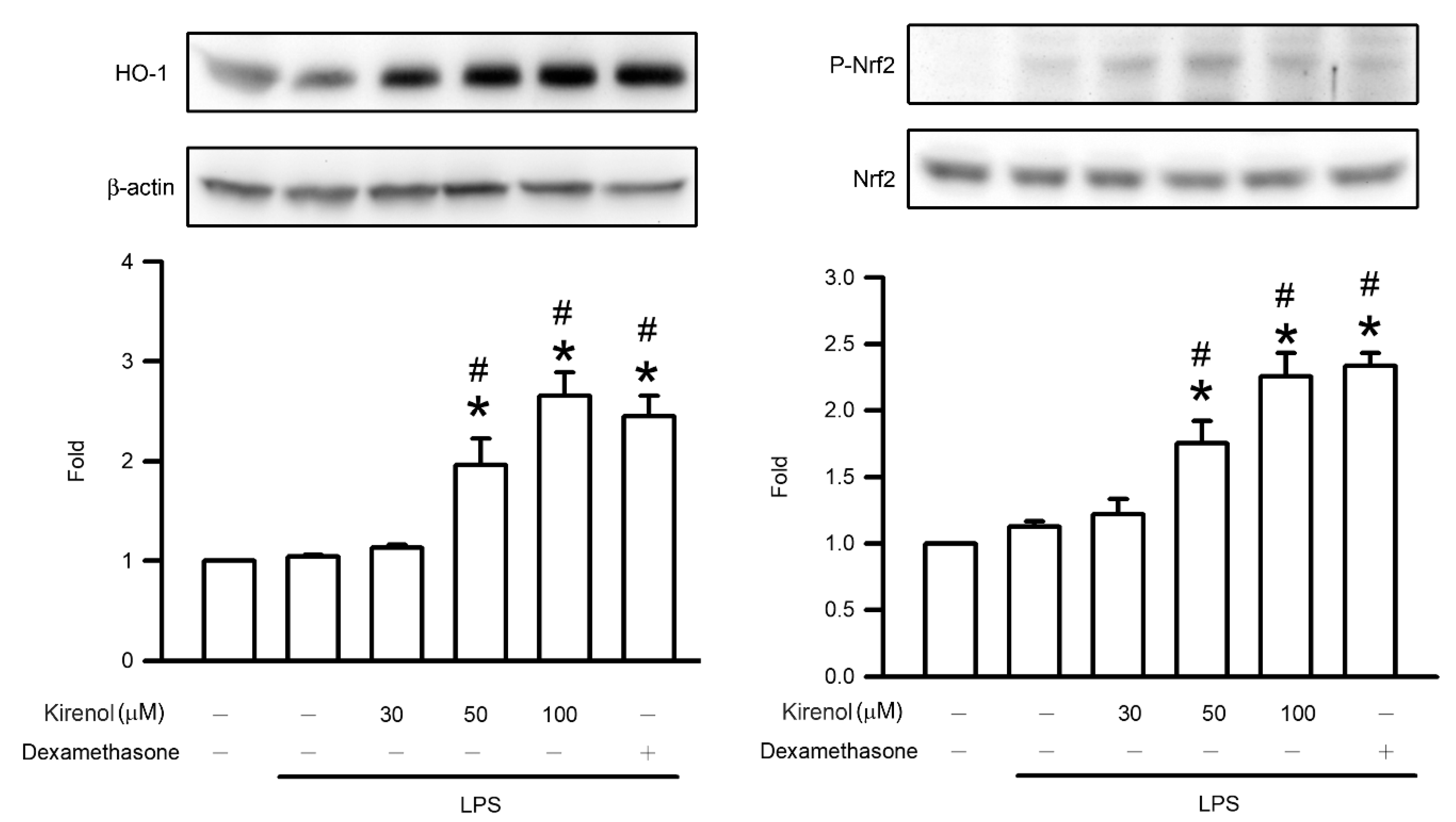
3.7. Kirenol Enhanced the Nrf2 Phosphorylation and HO-1 Expression in Mice with LPS-Induced ALI
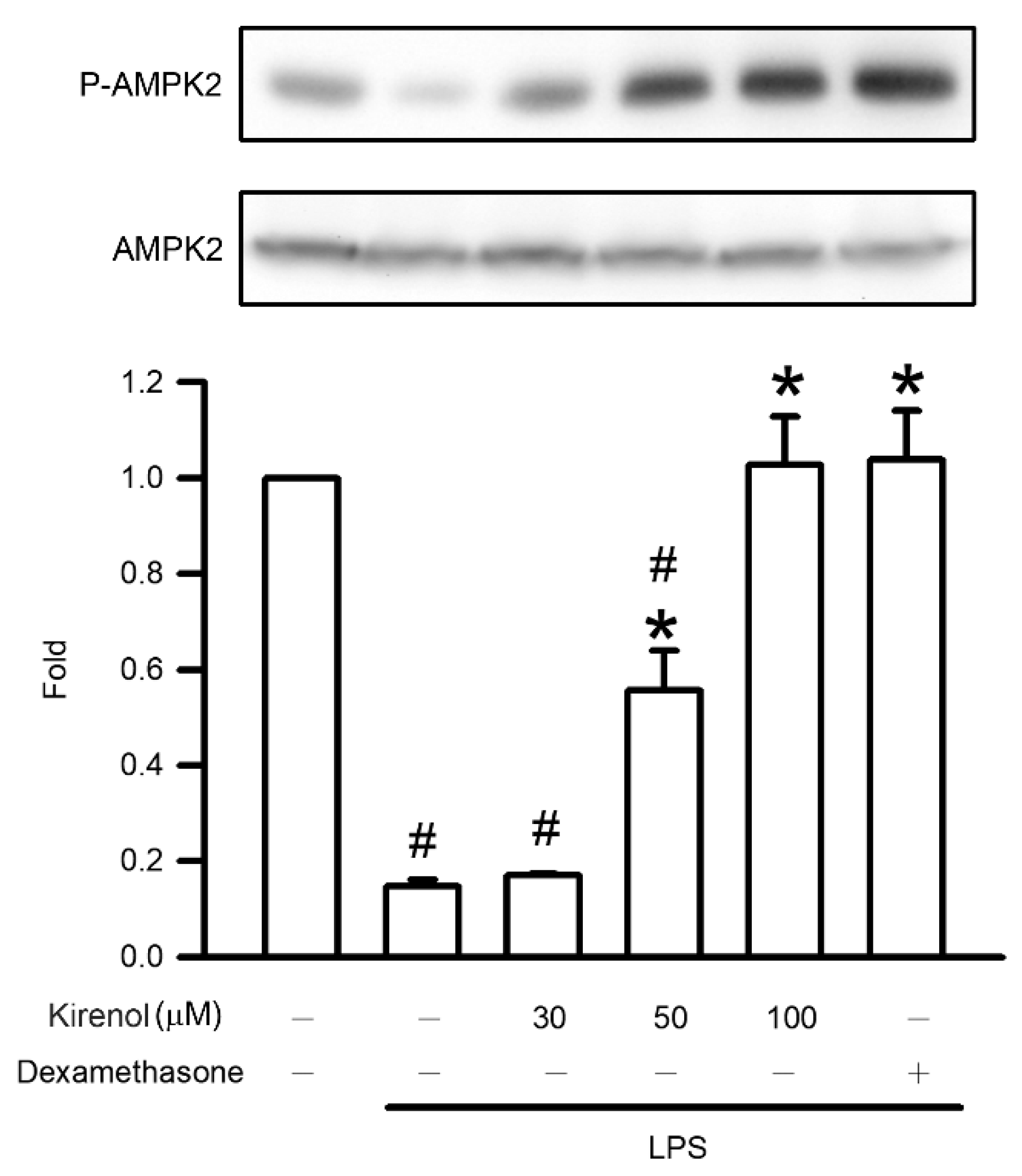
3.8. Kirenol Attenuated the Downregulation of AMPK2 Phosphorylation in Mice with LPS-Induced ALI
3.9. Kirenol Suppressed the Leukocyte Infiltration and Alveolar-Capillary Barrier Disruption in Mice with LPS-Induced ALI
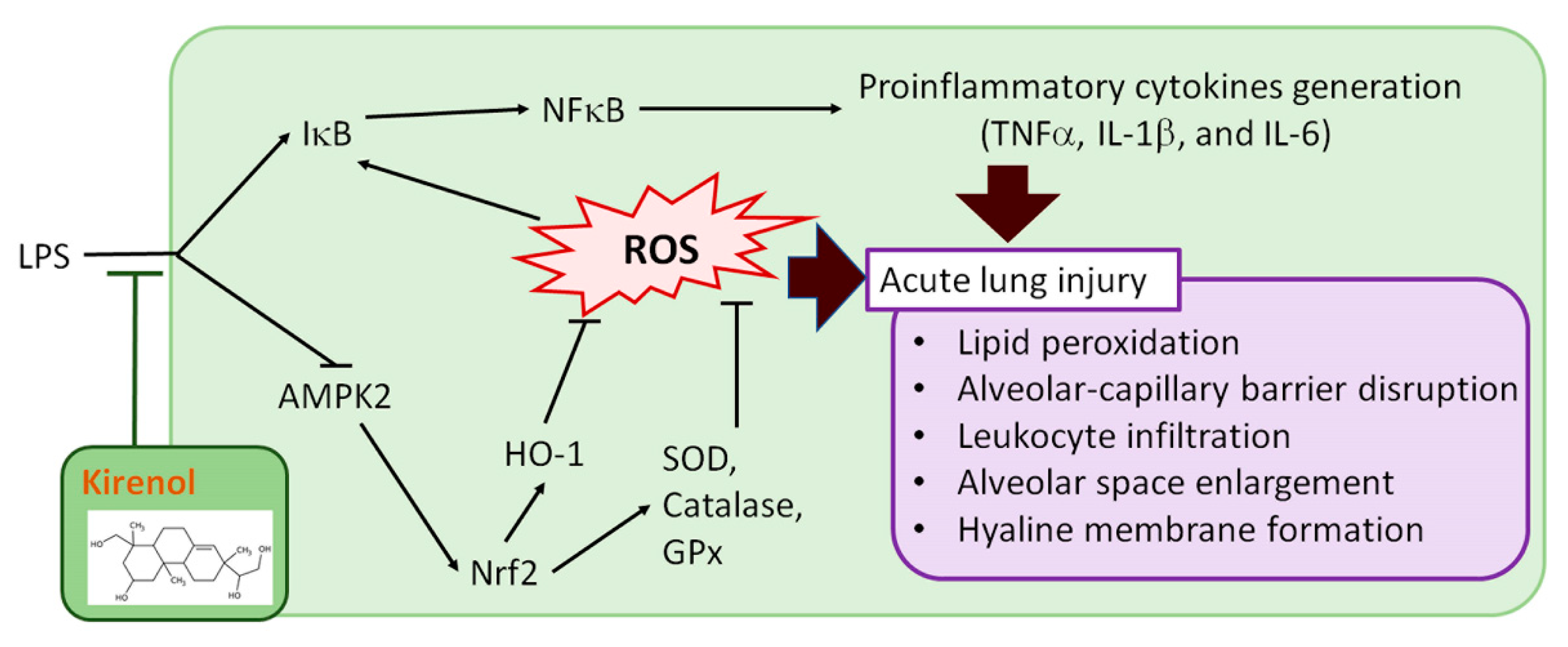
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abedi, F.; Hayes, A.W.; Reiter, R.; Karimi, G. Acute lung injury: The therapeutic role of Rho kinase inhibitors. Pharmacol. Res. 2020, 155, 104736. [Google Scholar] [CrossRef] [PubMed]
- Herrero, R.; Sanchez, G.; Lorente, J.A. New insights into the mechanisms of pulmonary edema in acute lung injury. Ann. Transl. Med. 2018, 6, 32. [Google Scholar] [CrossRef]
- Carnesecchi, S.; Pache, J.C.; Barazzone-Argiroffo, C. NOX enzymes: Potential target for the treatment of acute lung injury. Cell Mol. Life Sci. 2012, 69, 2373–2385. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.L.; Shen, H.T.; Su, C.H.; Chen, W.Y.; Huang-Liu, R.; Chen, C.J.; Chen, S.P.; Kuan, Y.H. Nerolidol Suppresses the Inflammatory Response during Lipopolysaccharide-Induced Acute Lung Injury via the Modulation of Antioxidant Enzymes and the AMPK/Nrf2/HO-1 Pathway. Oxid. Med. Cell Longev. 2019, 2019, 9605980. [Google Scholar] [CrossRef]
- Son, Y.; Lee, J.H.; Chung, H.T.; Pae, H.-O. Therapeutic roles of heme oxygenase-1 in metabolic diseases: Curcumin and resveratrol analogues as possible inducers of heme oxygenase-1. Oxid. Med. Cell Longev. 2013, 2013, 639541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imam, F.; Al-Harbi, N.O.; Al-Harbi, M.M.; Qamar, W.; Aljerian, K.; Belali, O.M.; Alsanea, S.; Alanazi, A.Z.; Alhazzani, K. Apremilast ameliorates carfilzomib-induced pulmonary inflammation and vascular injuries. Int. Immunopharmacol. 2019, 66, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Chen, S.P.; Su, C.H.; Ho, Y.C.; Yang, M.L.; Lee, S.S.; Huang-Liu, R.; Yang, C.P.; Chen, C.J.; Kuan, Y.H. Zerumbone from Zingiber zerumbet Ameliorates Lipopolysaccharide-Induced ICAM-1 and Cytokines Expression via p38 MAPK/JNK-IκB/NF-κB Pathway in Mouse Model of Acute Lung Injury. Chin. J. Physiol. 2018, 61, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.P.; Su, C.H.; Huang-Liu, R.; Lee, M.W.; Chiang, C.Y.; Chen, W.Y.; Chen, C.J.; Wu, S.W.; Kuan, Y.H. Protective effect of nerolidol on lipopolysaccharide-induced acute lung injury through the inhibition of NF-κB activation by the reduction of p38 MAPK and JNK phosphorylation. J. Funct. Food. 2020, 69, 103943. [Google Scholar] [CrossRef]
- Wheeler, A.P.; Bernard, G.R. Acute lung injury and the acute respiratory distress syndrome: A clinical review. Lancet 2007, 369, 1553–1564. [Google Scholar] [CrossRef]
- Jiang, Z.; Yu, Q.H.; Cheng, Y.; Guo, X.J. Simultaneous quantification of eight major constituents in Herba Siegesbeckiae by liquid chromatography coupled with electrospray ionization time-offlight tandem mass spectrometry. J. Pharm. Biomed. Anal. 2011, 55, 452–457. [Google Scholar] [CrossRef]
- Xiong, J.; Ma, Y.; Xu, Y. Diterpenoids from Siegesbeckia pubescens. Phytochemistry 1992, 31, 917–921. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Yang, L.R.; Li, Y.B.; Yan, L.H. Summary on chemical composition and pharmacological action of Herba Siegesbeckiae. Inf. Tradit. Chin. Med. 2006, 23, 15–17. [Google Scholar]
- Nguyen, T.D.; Thuong, P.T.; Hwang, I.H.; Hoang, T.K.; Nguyen, M.K.; Nguyen, H.A.; Na, M. Anti-hyperuricemic, anti-inflammatory and analgesic effects of Siegesbeckia orientalis L. resulting from the fraction with high phenolic contents. BMC Complement. Altern. Med. 2017, 17, 191. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Zhang, H.; Fan, C.Q.; Yue, J.M. Novel diterpenoids and diterpenoid glycosides from Siegesbeckia orientalis. J. Nat. Prod. 2004, 67, 1517–1521. [Google Scholar] [CrossRef]
- Huo, L.; Jiang, Z.; Lei, M.; Wang, X.; Guo, X. Simultaneous quantification of Kirenol and ent-16β,17-dihydroxy-kauran-19-oic acid from Herba Siegesbeckiae in rat plasma by liquid chromatography-tandem mass spectrometry and its application to pharmacokinetic studies. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2013, 937, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Xiao, J.; Wu, Z.W.; Wang, Z.M.; Hu, J.; Fu, H.Z.; Chen, Y.Y.; Qian, R.Q. Kirenol exerts a potent anti-arthritic effect in collagen-induced arthritis by modifying the T cells balance. Phytomedicine 2012, 19, 882–889. [Google Scholar] [CrossRef]
- An, X.; Sun, X.; Hou, Y.; Yang, X.; Chen, H.; Zhang, P.; Wu, J. Protective effect of oxytocin on LPS-induced acute lung injury in mice. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Shastri, M.D.; Shukla, S.D.; Chong, W.C.; Dua, K.; Peterson, G.M.; Patel, R.P.; Hansbro, P.M.; Eri, R.; O’Toole, R.F. Role of Oxidative Stress in the Pathology and Management of Human Tuberculosis. Oxidative Med. Cell. Longev. 2018, 2018, 7695364. [Google Scholar] [CrossRef] [Green Version]
- Barnes, P.J. Oxidative stress-based therapeutics in COPD. Redox Biol. 2020, 33, 101544. [Google Scholar] [CrossRef]
- Li, X.; Wang, W.; Zhao, X.; Zai, J.; Zhao, Q.; Li, Y.; Chaillon, A. Transmission dynamics and evolutionary history of 2019-nCoV. J. Med. Virol. 2020, 92, 501–511. [Google Scholar] [CrossRef]
- Lucas, R.; Hadizamani, Y.; Gonzales, J.N.; Gorshkov, B.; Bodmer, T.; Berthiaume, Y.; Moehrlen, U.; Lode, H.; Huwer, H.; Hudel, M.; et al. Impact of Bacterial Toxins in the Lungs. Toxins 2020, 12, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellani, G.; Laffey, J.G.; Pham, T.; Fan, E.; Brochard, L.; Esteban, A.; Gattinoni, L.; Van Haren, F.; LUNG SAFE Investigators; ESICM Trials Group; et al. Epidemiology, Patterns of Care, and Mortality for Patients with Acute Respiratory Distress Syndrome in Intensive Care Units in 50 Countries. JAMA 2016, 315, 788–800. [Google Scholar] [CrossRef] [PubMed]
- Fan, E.; Brodie, D.; Slutsky, A.S. Acute respiratory distress syndrome: Advances in diagnosis and treatment. JAMA J. Am. Med. Assoc. 2018, 319, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Máca, J.; Jor, O.; Holub, M.; Sklienka, P.; Burša, F.; Burda, M.; Janout, V.; Ševčík, P. Past and present ARDS mortality rates: A systematic review. Respir. Care 2017, 62, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Li, Q.; Jin, L.; Qu, Y.; Liang, B.B.; Zhu, X.T.; Du, H.Y.; Jie, L.G.; Yu, Q.H. Kirenol Inhibits the Function and Inflammation of Fibroblast-like Synoviocytes in Rheumatoid Arthritis in vitro and in vivo. Front. Immunol. 2019, 10, 1304. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Huang, X.Y.; Li, L.; Fan, X.H.; Li, P.C.; Huang, C.Q.; Xiao, J.; Gui, R.; Wang, S. Attenuation of diabetic cardiomyopathy by relying on kirenol to suppress inflammation in a diabetic rat model. J. Cell Mol. Med. 2019, 23, 7651–7663. [Google Scholar] [CrossRef] [Green Version]
- Alastair, G.; Proudfoot, A.G.; McAuley, D.F.; Griffiths, M.J.D.; Hind, M. Human models of acute lung injury. Dis. Model Mech. 2011, 4, 145–153. [Google Scholar]
- Holtzman, M.J.; Byers, D.E.; Alexander-Brett, J.; Wang, X. The role of airway epithelial cells and innate immune cells in chronic respiratory disease. Nat. Rev. Immunol. 2014, 14, 686–698. [Google Scholar] [CrossRef] [Green Version]
- Georas, S.N.; Rezaee, F. Epithelial barrier function: At the front line of asthma immunology and allergic airway inflammation. J. Allergy Clin. Immunol. 2014, 134, 509–520. [Google Scholar] [CrossRef] [Green Version]
- Rackley, C.R.; Stripp, B.R. Building and maintaining the epithelium of the lung. J. Clin. Investig. 2012, 122, 2724–2730. [Google Scholar] [CrossRef] [Green Version]
- Higuita-Castro, N.; Nelson, M.T.; Shukla, V.; Agudelo-Garcia, P.A.; Zhang, W.; Duarte-Sanmiguel, S.M.; Englert, J.A.; Lannutti, J.J.; Hansford, D.J.; Ghadiali, S.N. Using a Novel Microfabricated Model of the Alveolar-Capillary Barrier to Investigate the Effect of Matrix Structure on Atelectrauma. Sci. Rep. 2017, 7, 11623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.P.; Zhou, Y.M.; Ye, Y.J.; Shang, X.M.; Cai, Y.L.; Xiong, C.M.; Wu, Y.X.; Xu, H.X. Topical anti-inflammatory and analgesic activity of kirenol isolated from Siegesbeckia orientalis. J. Ethnopharmacol. 2011, 137, 1089–1094. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Ma, M.; Guo, G.; Tang, Z. Kirenol regulates the cell proliferative and inflammatory markers in DMBA-induced oral squamous cell carcinogenesis in hamster. Environ. Toxicol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.M.; Zhu, S.G.; Wu, Z.W.; Lu, Y.; Fu, H.Z.; Qian, R.Q. Kirenol upregulates nuclear annexin-1 which interacts with NF-kappaB to attenuate synovial inflammation of collagen-induced arthritis in rats. J. Ethnopharmacol. 2011, 137, 774–782. [Google Scholar] [CrossRef]
- Kim, J.; Kim, M.B.; Yun, J.G.; Hwang, J.K. Protective Effects of Standardized Siegesbeckia glabrescens Extract and Its Active Compound Kirenol against UVB-Induced Photoaging through Inhibition of MAPK/NF-kappaB Pathways. J. Microbiol. Biotechnol. 2017, 27, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Wang, X.; Saredy, J.; Yuan, Z.; Yang, X.; Wang, H. Innate-adaptive immunity interplay and redox regulation in immune response. Redox Biol. 2020, 37, 101759. [Google Scholar] [CrossRef]
- Lingappan, K. NF-κB in Oxidative Stress. Curr. Opin. Toxicol. 2017, 7, 81–86. [Google Scholar] [CrossRef]
- Chen, L.; Hu, J.Y.; Wang, S.Q. The role of antioxidants in photoprotection: A critical review. J. Am. Acad. Dermatol. 2012, 67, 1013–1024. [Google Scholar] [CrossRef]
- Ren, J.; Yang, M.; Chen, J.; Ma, S.; Wang, N. Anti-inflammatory and wound healing potential of kirenol in diabetic rats through the suppression of inflammatory markers and matrix metalloproteinase expressions. Biomed. Pharmacother. 2020, 129, 110475. [Google Scholar] [CrossRef]
- Ranman, I.; Biswas, S.K. Oxidants and Antioxidants Antioxidants, Enzymatic. In Encyclopedia of Respiratory Medicine; Academic Press: Cambridge, MA, USA, 2006; pp. 258–266. [Google Scholar]
- Weaver, L.; Hamoud, A.R.; Stec, D.E.; Hinds, T.D., Jr. Biliverdin reductase and bilirubin in hepatic disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 314, G668–G676. [Google Scholar] [CrossRef]
- Thomas, W.S.; Solomon, H.S. Bilirubin Benefits: Cellular Protection by a Biliverdin Reductase Antioxidant cycle. Pediatrics 2004, 113, 1776–1782. [Google Scholar]
- Ahmed, S.M.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Matzinger, M.; Fischhuber, K.; Pölöske, D.; Mechtler, K.; Heiss, E.H. AMPK leads to phosphorylation of the transcription factor Nrf2, tuning transactivation of selected target genes. Redox Biol. 2020, 29, 101393. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, F.C.-F.; Lee, S.-S.; Li, Y.-C.; Ho, Y.-C.; Chen, W.-Y.; Chen, C.-J.; Lee, M.-W.; Yeh, K.-L.; Tsai, S.C.-S.; Kuan, Y.-H. Protective Effects of Kirenol against Lipopolysaccharide-Induced Acute Lung Injury through the Modulation of the Proinflammatory NFκB Pathway and the AMPK2-/Nrf2-Mediated HO-1/AOE Pathway. Antioxidants 2021, 10, 204. https://doi.org/10.3390/antiox10020204
Lin FC-F, Lee S-S, Li Y-C, Ho Y-C, Chen W-Y, Chen C-J, Lee M-W, Yeh K-L, Tsai SC-S, Kuan Y-H. Protective Effects of Kirenol against Lipopolysaccharide-Induced Acute Lung Injury through the Modulation of the Proinflammatory NFκB Pathway and the AMPK2-/Nrf2-Mediated HO-1/AOE Pathway. Antioxidants. 2021; 10(2):204. https://doi.org/10.3390/antiox10020204
Chicago/Turabian StyleLin, Frank Cheau-Feng, Shiuan-Shinn Lee, Yi-Ching Li, Yung-Chuan Ho, Wen-Ying Chen, Chun-Jung Chen, Min-Wei Lee, Kun-Lin Yeh, Stella Chin-Shaw Tsai, and Yu-Hsiang Kuan. 2021. "Protective Effects of Kirenol against Lipopolysaccharide-Induced Acute Lung Injury through the Modulation of the Proinflammatory NFκB Pathway and the AMPK2-/Nrf2-Mediated HO-1/AOE Pathway" Antioxidants 10, no. 2: 204. https://doi.org/10.3390/antiox10020204