
Essential Oils from Zingiber striolatum Diels Attenuate Inflammatory Response and Oxidative Stress through Regulation of MAPK and NF-κB Signaling Pathways

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Extraction of EOZS
2.2. Chemicals and Reagents
2.3. Analysis of Chemical Compounds of EOZS
2.4. Antioxidant Assay
2.5. Cell Culture and Detected of Cytotoxicity
2.6. Determination of NO Production
2.7. Determination of Intracellular ROS
2.8. ELISA Assay
2.9. Analysis of Cell Morphology and Size
2.10. Cell Migration Assay
2.11. Determination of Phagocytosis
2.12. RNA Sequencing
2.13. Western Blotting
2.14. Immunofluorescence Staining
2.15. TPA-Induced Mouse Ear Edema Model
2.16. Hematoxylin–Eosin Staining
2.17. Immunohistochemistry Staining
2.18. Statistical Analysis
3. Results
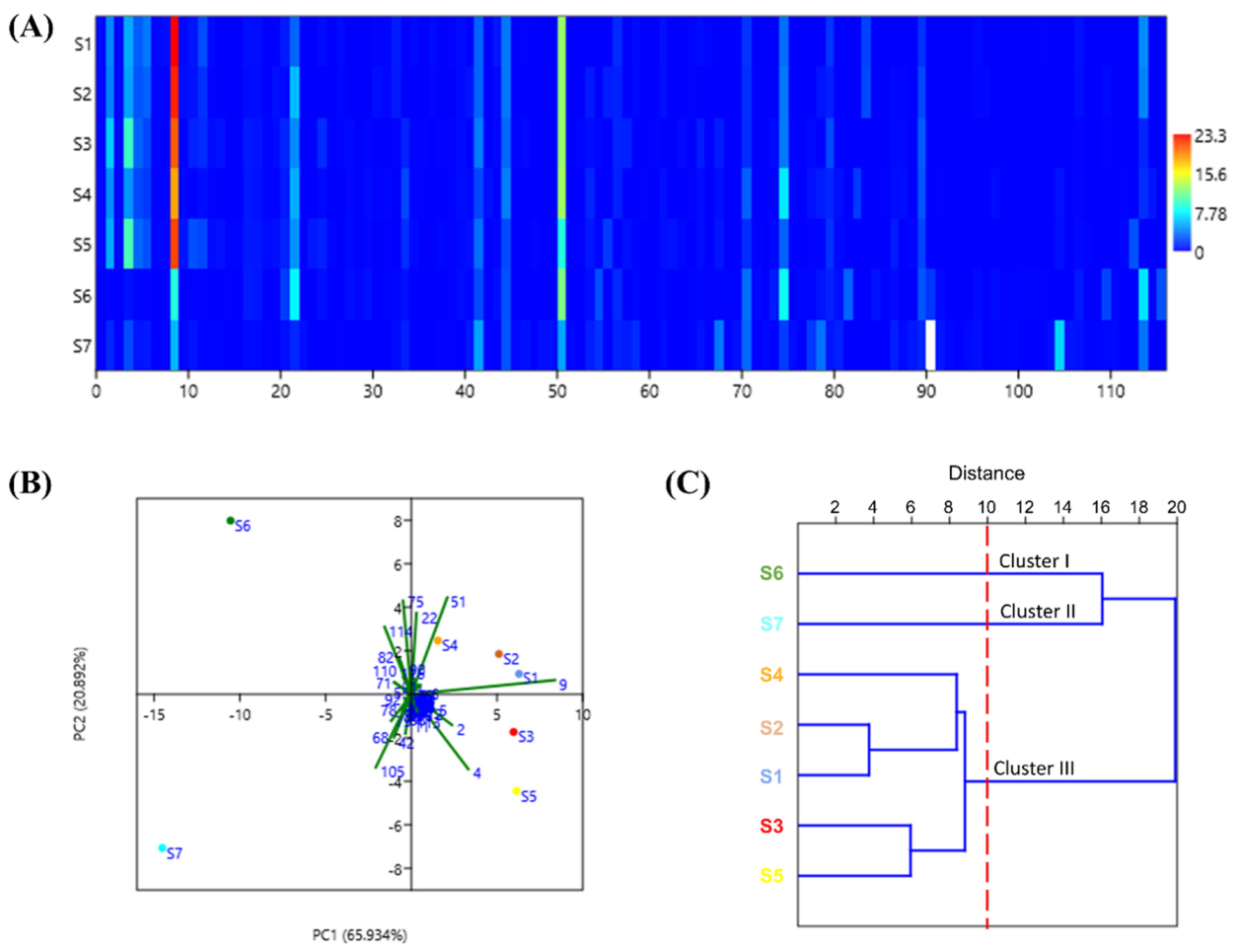
3.1. The Compositions, PCA and HCA of EOZS from Different Regions
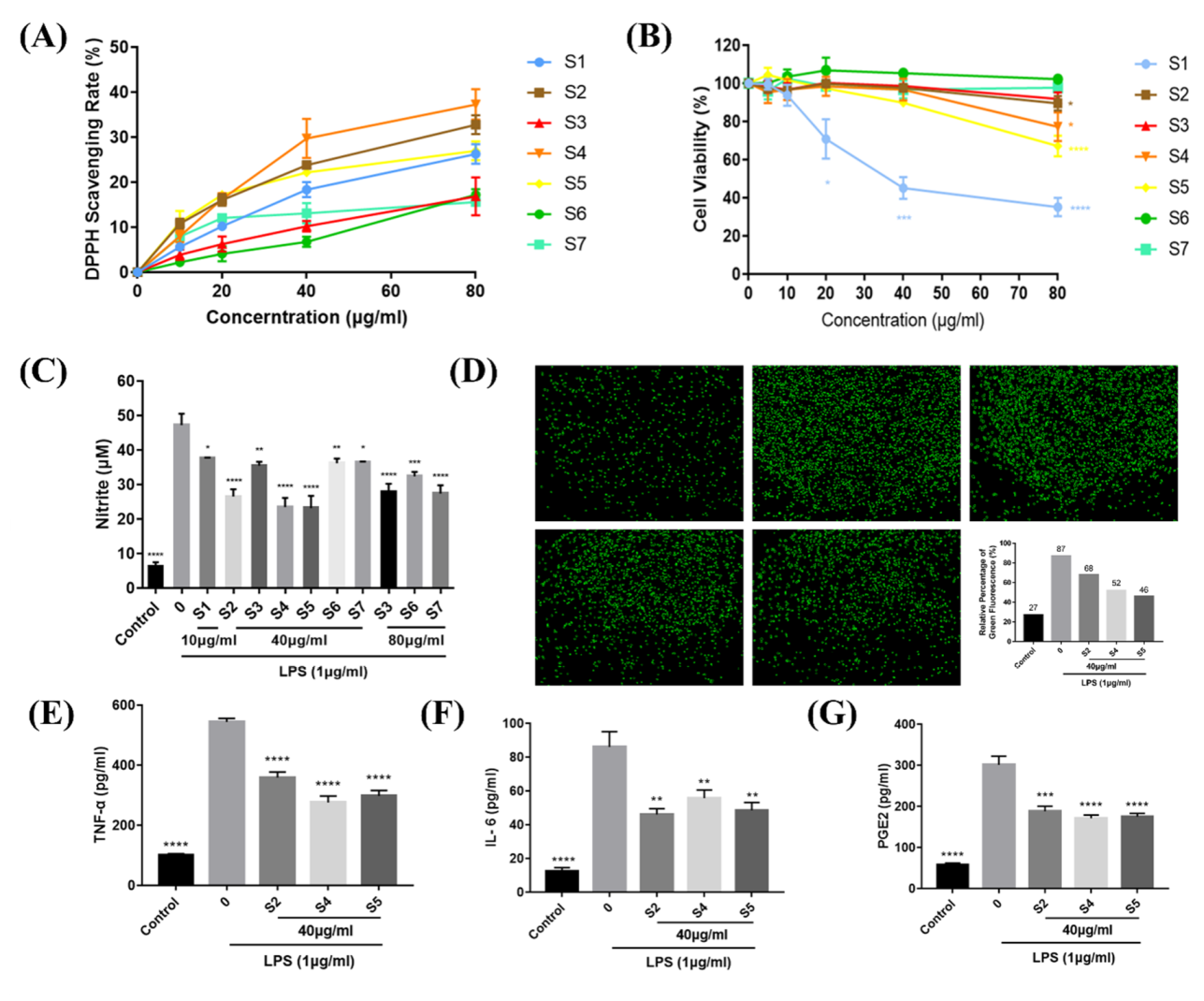
3.2. Evaluation of the Antioxidant Activity of EOZS
3.2.1. DPPH Scavenging Activity
3.2.2. Effects of EOZS on NO and ROS in LPS-Induced RAW 264.7 Cells
3.3. EOZS Attenuated Production of Pro-Inflammatory Mediators
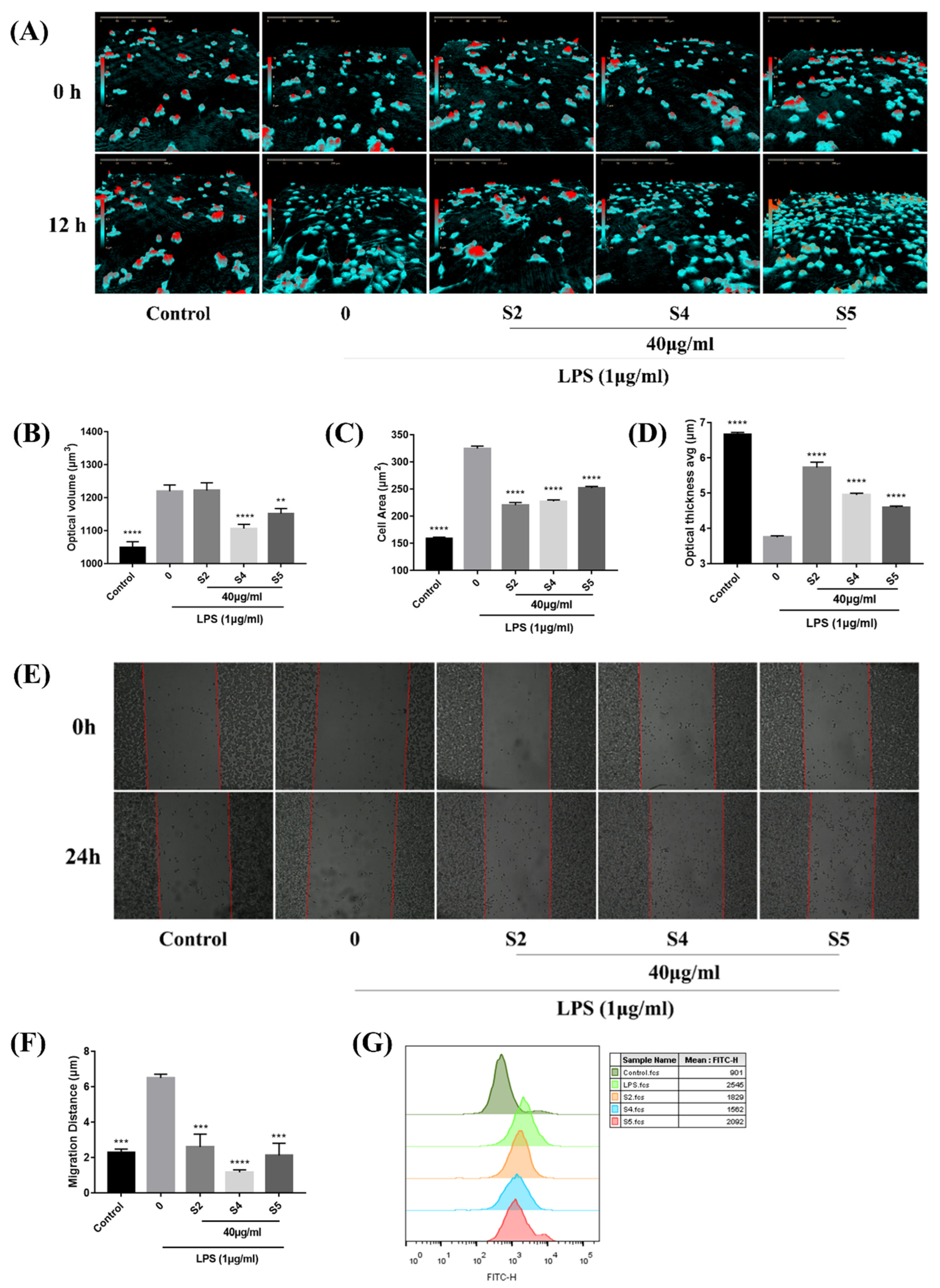
3.4. EOZS Reduced Cellular Morphological Changes, Migration and Phagocytic Capability in LPS-Induced RAW 264.7 Cells
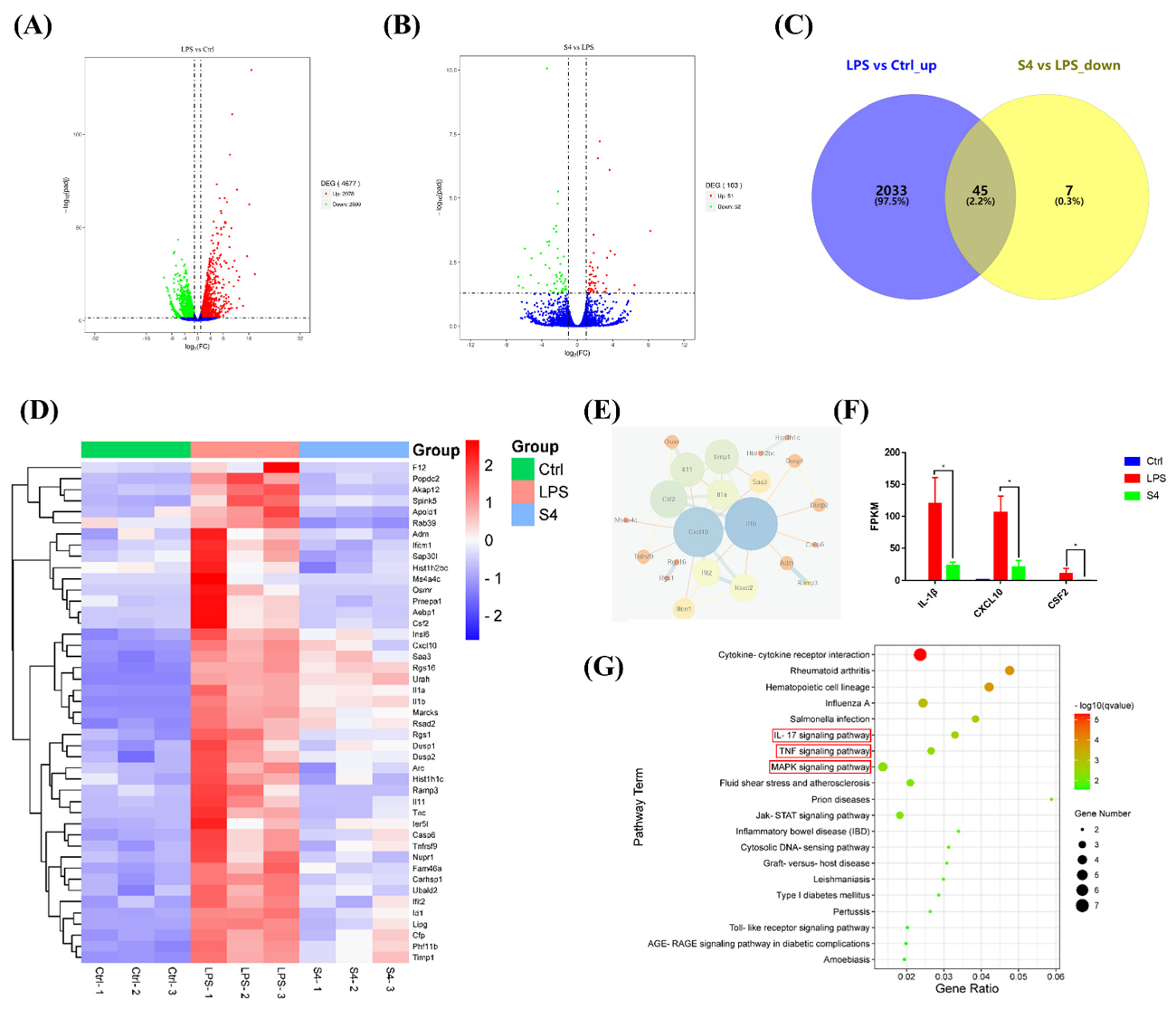
3.5. Bioinformatics Analysis of RNA Sequencing Data
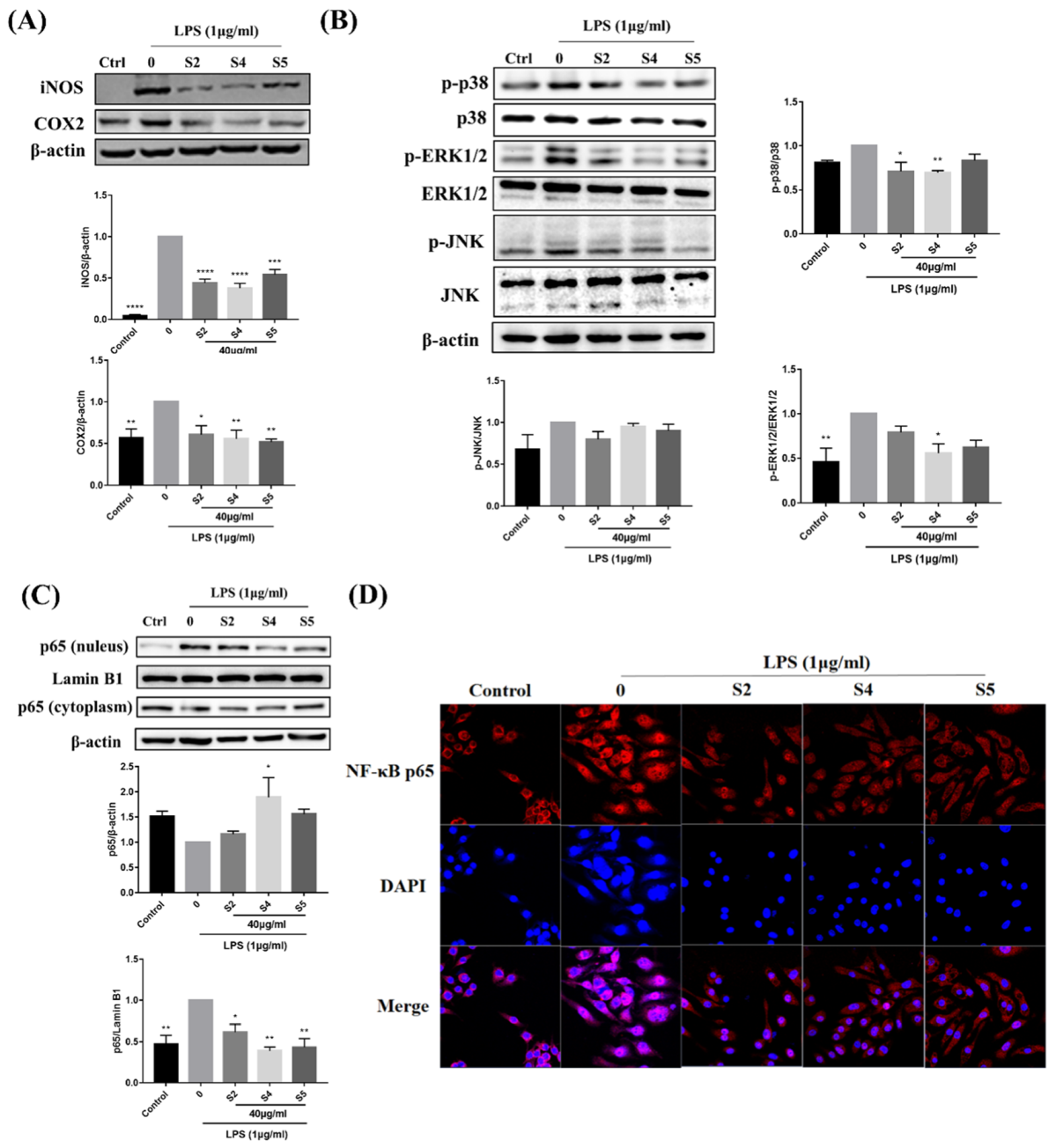
3.6. EOZS Inhibited Expression Level of iNOS, COX2 and Activation of MAPK and NF-κB Pathways in LPS-Induced RAW 264.7 Cells
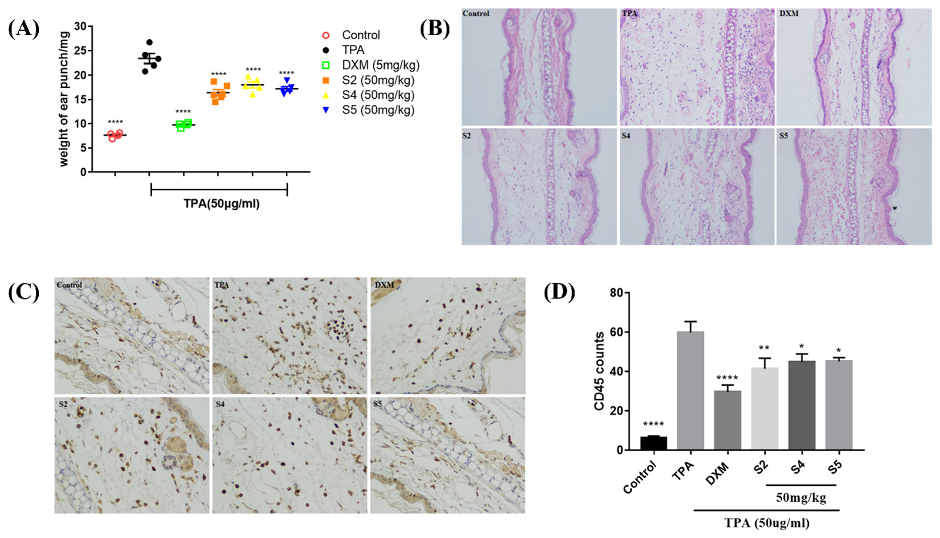
3.7. EOZS Ameliorated Ear Edema and Infiltration of Immune Cells in TPA-Induced Mouse Model
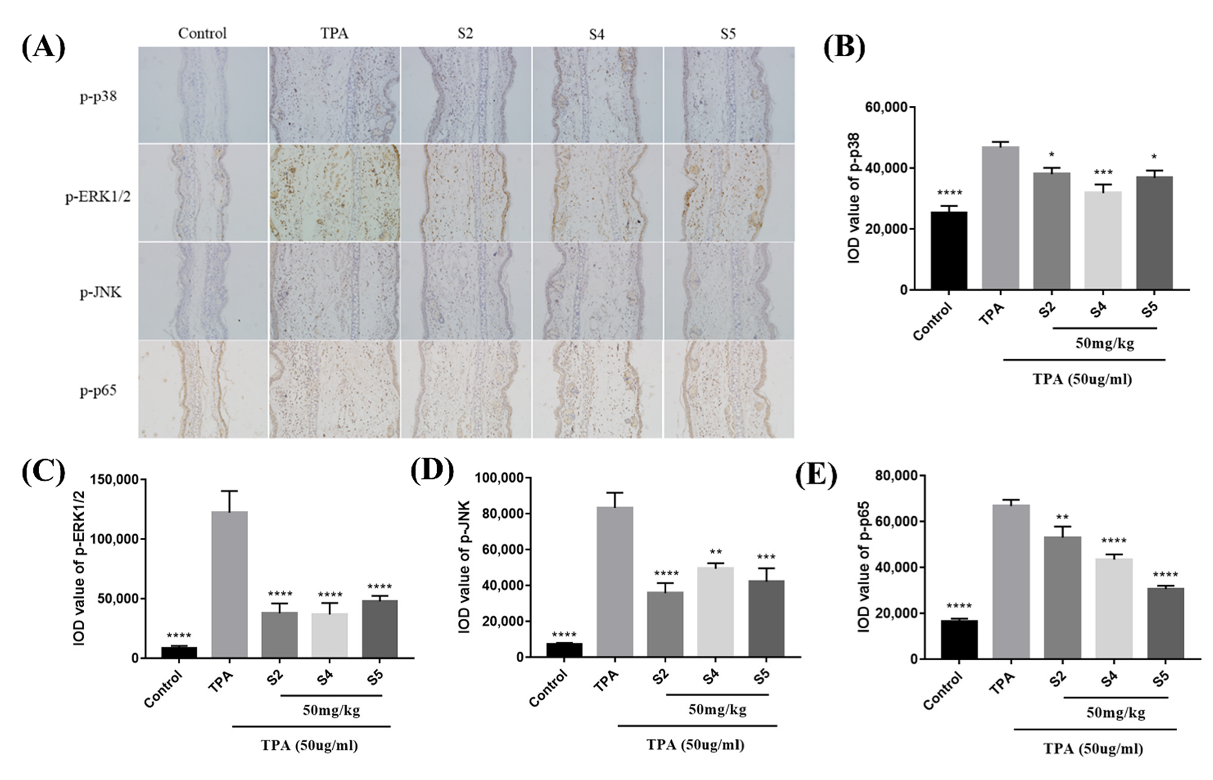
3.8. EOZS Suppressed MAPK and NF-κB Pathways in TPA-Induced Mouse Model
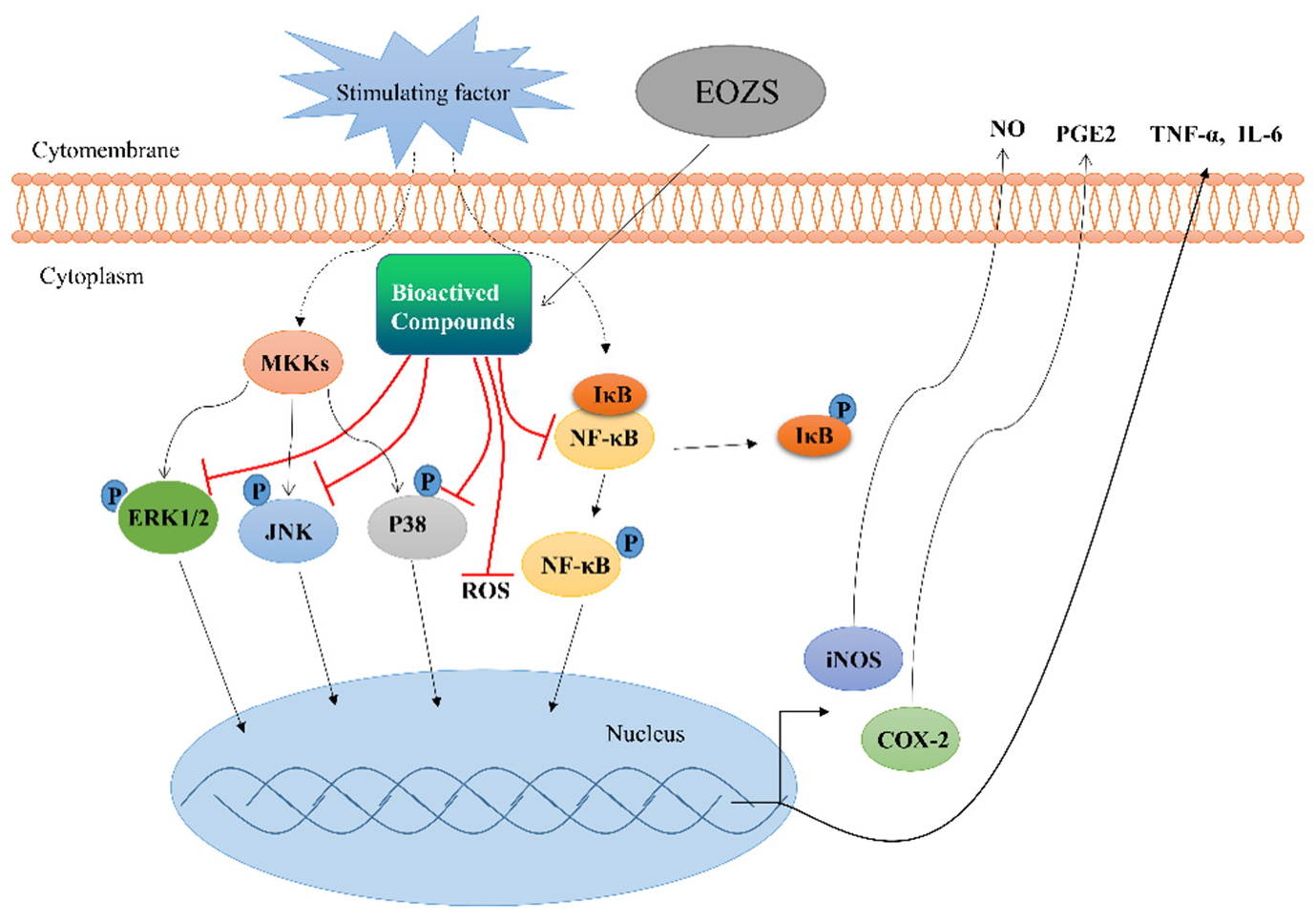
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Nasef, N.A.; Mehta, S.; Ferguson, L.R. Susceptibility to chronic inflammation: An update. Arch. Toxicol. 2017, 91, 1131–1141. [Google Scholar] [CrossRef]
- Duran, W.N.; Breslin, J.W.; Sanchez, F.A. The NO cascade, eNOS location, and microvascular permeability. Cardiovasc. Res. 2010, 87, 254–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fajgenbaum, D.C.; June, C.H. Cytokine storm. New Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Bozec, A.; Ramming, A.; Schett, G. Anti-inflammatory and immune-regulatory cytokines in rheumatoid arthritis. Nat. Rev. Rheumatol. 2019, 15, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Sorci-Thomas, M.G.; Thomas, M.J. Microdomains, inflammation, and atherosclerosis. Circ. Res. 2016, 118, 679–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdulkhaleq, L.A.; Assi, M.A.; Abdullah, R.; Zamri-Saad, M.; Taufiq-Yap, Y.H.; Hezmee, M.N.M. The crucial roles of inflammatory mediators in inflammation: A review. Vet. World 2018, 11, 627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamidzadeh, K.; Christensen, S.M.; Dalby, E.; Chandrasekaran, P.; Mosser, D.M. Macrophages and the recovery from acute and chronic inflammation. Annu. Rev. Physiol. 2017, 79, 567–592. [Google Scholar] [CrossRef] [Green Version]
- Lv, H.; Yang, H.; Wang, Z.; Feng, H.; Deng, X.; Cheng, G.; Ci, X. Nrf2 signaling and autophagy are complementary in protecting lipopolysaccharide/d-galactosamine-induced acute liver injury by licochalcone A. Cell Death Dis. 2019, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Beltrán-García, J.; Osca-Verdegal, R.; Pallardó, F.V.; Ferreres, J.; Rodríguez, M.; Mulet, S.; Sanchis-Gomar, F.; Carbonell, N.; García-Giménez, J.L. Oxidative stress and inflammation in COVID-19-associated sepsis: The potential role of anti-oxidant therapy in avoiding disease progression. Antioxidants 2020, 9, 936. [Google Scholar] [CrossRef]
- Deng, Y.; Matsui, Y.; Pan, W.; Li, Q.; Lai, Z. Yap1 plays a protective role in suppressing free fatty acid-induced apoptosis and promoting beta-cell survival. Protein Cell 2016, 7, 362–372. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, A.; Tsuge, M.; Miyahara, N.; Tsukahara, H. Reactive oxygen species and antioxidative defense in chronic obstructive pulmonary disease. Antioxidants 2021, 10, 1537. [Google Scholar] [CrossRef]
- Miguel, M.G. Antioxidant and anti-inflammatory activities of essential oils: A short review. Molecules 2010, 15, 9252–9287. [Google Scholar] [CrossRef] [Green Version]
- Freires, I.A.; Denny, C.; Benso, B.; De Alencar, S.M.; Rosalen, P.L. Antibacterial activity of essential oils and their isolated constituents against cariogenic bacteria: A systematic review. Molecules 2015, 20, 7329–7358. [Google Scholar] [CrossRef]
- Bhalla, Y.; Gupta, V.K.; Jaitak, V. Anticancer activity of essential oils: A review. J. Sci. Food Agr. 2013, 93, 3643–3653. [Google Scholar] [CrossRef]
- Huang, Z.; Xie, L.; Wang, H.; Zhong, J.; Li, Y.; Liu, J.; Ou, Z.; Liang, X.; Li, Y.; Huang, H.; et al. Geographic distribution and impacts of climate change on the suitable habitats of Zingiber species in China. Ind. Crop Prod. 2019, 138, 111429. [Google Scholar] [CrossRef]
- Chen, T.; Cai, J.; Ni, J.; Yang, F. An UPLC-MS/MS application to investigate chemical compositions in the ethanol extract with hypoglycemic activity from Zingiber striolatum Diels. J. Chin. Pharm. Sci. 2016, 25, 116–121. [Google Scholar]
- Zhang, C.; Zhang, X.; Jiang, N.; Wang, J.; Li, W.; Zhou, D.; Zheng, X. Analysis of amino acid composition and evaluation of nutritional value of Zingiber strioatum Diels. J. Hubei Univ. Natl.-Nat. Sci. Ed. 2014, 32, 380–383. [Google Scholar]
- Tian, M.; Liu, T.; Wu, X.; Hong, Y.; Zhou, Y. Chemical composition, antioxidant, antimicrobial and anticancer activities of the essential oil from the rhizomes of Zingiber striolatum Diels. Nat. Prod. Res. 2020, 34, 2621–2625. [Google Scholar] [CrossRef]
- Tian, M.; Hong, Y.; Wu, X.; Zhang, M.; Lin, B.; Zho, Y. Chemical constituents and cytotoxic activities of essential oils from the flowers, leaves and stems of Zingiber striolatum diels. Rec. Nat. Prod. 2020, 14, 144–149. [Google Scholar] [CrossRef]
- Raposo, J.D.A.; Figueiredo, P.L.B.; Santana, R.L.; Junior, A.Q.D.; Suemitsu, C.; Silva, R.D.; Mourão, R.H.V.; Maia, J.G.S. Seasonal and circadian study of the essential oil of Myrcia sylvatica (G. Mey) DC., a valuable aromatic species occurring in the Lower Amazon River region. Biochem. Syst. Ecol. 2018, 79, 21–29. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; Volume 456. [Google Scholar]
- Berwal, R.; Vasudeva, N.; Sharma, S.; Das, S. Investigation on biomolecules in ethanol extract of fruits of Prosopis Juliflora (Sw.) DC. using GC-MS. J. Herbs Spices Med. Plants 2019, 25, 172–180. [Google Scholar] [CrossRef]
- Dötterl, S.; Jürgens, A. Spatial fragrance patterns in flowers of Silene latifolia: Lilac compounds as olfactory nectar guides? Plant Syst. Evol. 2005, 255, 99–109. [Google Scholar] [CrossRef]
- Hajlaoui, H.; Mighri, H.; Noumi, E.; Snoussi, M.; Trabelsi, N.; Ksouri, R.; Bakhrouf, A. Chemical composition and biological activities of Tunisian Cuminum cyminum L. essential oil: A high effectiveness against Vibrio spp. strains. Food Chem. Toxicol. 2010, 48, 2186–2192. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y.; Wei, S.; Yan, Z. Comparative seasonal variation and chemical composition of essential oils from the leaves and stems of Schefflera heptaphylla using microwave-assisted and conventional hydrodistillation. Ind. Crop Prod. 2012, 36, 229–237. [Google Scholar] [CrossRef]
- Lawal, O.A.; Ogunwande, I.A.; Opoku, A.R. Chemical composition of essential oils of Plumeria rubra L. grown in Nigeria. Eur. J. Med. Plants 2015, 6, 55–61. [Google Scholar] [CrossRef]
- Rahman, M.M.; Sultana, T.; Ali, M.Y.; Rahman, M.M.; Al-Reza, S.M.; Rahman, A. Chemical composition and antibacterial activity of the essential oil and various extracts from Cassia sophera L. against Bacillus sp. from soil. Arab. J. Chem. 2017, 10, S2132–S2137. [Google Scholar] [CrossRef] [Green Version]
- Ospina, L.M.P.; Muñoz, P.B.; Matulevich, J.; Teherán, A.A.; Villamizar, L.B. Composition and antimicrobial activity of the essential oils of three plant species from the Sabana of Bogota (Colombia): Myrcianthes leucoxyla, Vallea stipularis and Phyllanthus salviifolius. Nat. Prod. Commun. 2016, 11, 1913–1918. [Google Scholar] [CrossRef] [Green Version]
- Lazari, D.M.; Skaltsa, H.D.; Constantinidis, T. Volatile constituents of Centaurea pelia DC.; C. thessala Hausskn. subsp. drakiensis (Freyn & Sint.) Georg. and C. zuccariniana DC. from Greece. Flavour Fragr. J. 2000, 15, 7–11. [Google Scholar]
- Cui, J.; Guo, S.; Fu, S.; Xiao, P.; Wang, M. Effects of inoculating fungi on agilawood formation in Aquilaria sinensis. Chin. Sci. Bull. 2013, 58, 3280–3287. [Google Scholar] [CrossRef] [Green Version]
- Yao, X.T.; Ling, P.X.; Jiang, S.; Lai, P.X.; Zhu, C.G. Analysis of the essential oil from Gaillardia pulchella Foug. and its antioxidant activity. J. Oleo Sci. 2013, 62, 329–333. [Google Scholar] [CrossRef] [Green Version]
- Zahra, G.; Khadijeh, B.; Hossein, D.; Ali, S. Essential oil composition of two Scutellaria species from Iran. J. Tradit. Chin. Med. Sci. 2019, 6, 244–253. [Google Scholar] [CrossRef]
- Tian, H.; Zhao, H.; Zhou, H.; Zhang, Y. Chemical composition and antimicrobial activity of the essential oil from the aerial part of Dictamnus dasycarpus Turcz. Ind. Crop. Prod. 2019, 140, 111713. [Google Scholar] [CrossRef]
- Khaledian, Y.; Pajohi-Alamoti, M.; Bazargani-Gilani, B. Development of cellulose nanofibers coating incorporated with ginger essential oil and citric acid to extend the shelf life of ready-to-cook barbecue chicken. J. Food Process. Preserv. 2019, 43, e14114. [Google Scholar] [CrossRef]
- Radulović, N.S.; Ðorđević, M.R. Chemical composition of the tuber essential oil from Helianthus tuberosus L.(Asteraceae). Chem. Biodivers. 2014, 11, 427–437. [Google Scholar] [CrossRef]
- Elamrani, A.; Benaissa, M. Chemical composition and antibacterial activity of the essential oil of Ononis natrix from Morocco. J. Essent. Oil Bear. Plants 2010, 13, 477–488. [Google Scholar] [CrossRef]
- Liao, S.; Han, C.; Xu, D.; Fu, X.; Wang, J.; Kong, L. 4-Octyl itaconate inhibits aerobic glycolysis by targeting GAPDH to exert anti-inflammatory effects. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cui, S.; Wu, Q.; Wang, J.; Li, M.; Qian, J.; Li, S. Quercetin inhibits LPS-induced macrophage migration by suppressing the iNOS/FAK/paxillin pathway and modulating the cytoskeleton. Cell Adh. Migr. 2019, 13, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Jiang, Y.; Chu, Q.; Zheng, X. Radix Tetrastigma extract from different origins protect RAW264. 7 macrophages against LPS-induced inflammation. J. Food Sci. 2020, 85, 1586–1595. [Google Scholar] [CrossRef]
- Yang, W.S.; Yi, Y.S.; Kim, D.; Kim, M.H.; Park, J.G.; Kim, E.; Lee, S.Y.; Yoon, K.; Kim, J.H.; Park, J.; et al. Nuclear factor kappa-B and activator protein-1-mediated immunostimulatory activity of compound K in monocytes and macrophages. J. Ginseng Res. 2017, 41, 298–306. [Google Scholar] [CrossRef]
- Zeng, W.J.; Tan, Z.; Lai, X.F.; Xu, Y.N.; Mai, C.L.; Zhang, J.; Lin, Z.J.; Liu, X.G.; Sun, S.L.; Zhou, L.J. Topical delivery of L-theanine ameliorates TPA-induced acute skin inflammation via downregulating endothelial PECAM-1 and neutrophil infiltration and activation. Chem. Biol. Interact. 2018, 8, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.; Choudhary, V.; Seremwe, M.; Edwards, J.G.; Wang, A.; Emmons, A.C.; Bollag, K.A.; Johnson, M.H.; Bollag, W.B. Soy phosphatidylglycerol reduces inflammation in a contact irritant ear edema mouse model in vivo. J. Pharmacol. Exp. Ther. 2018, 366, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, R.; Wang, Y.; Zhang, J.; Wang, Q.; Cheng, A.; Guo, X.; Wang, X.; Sun, J. Supercritical CO2 fluid extraction of croton crassifolius Geisel root: Chemical composition and anti-proliferative, autophagic, apoptosis-inducing, and related molecular effects on A549 tumour cells. Phytomedicine 2019, 61, 152846. [Google Scholar] [CrossRef] [PubMed]
- Leite, L.H.I.; Leite, G.O.; Silva, B.A.F.; Santos, S.A.A.R.; Magalhães, F.E.A.; Menezes, P.P.; Serafini, M.R.; Teixeira, C.S.; Brito, R.G.; Santos, P.L.; et al. Molecular mechanism underlying orofacial antinociceptive activity of Vanillosmopsis arborea Baker (Asteraceae) essential oil complexed with β-cyclodextrin. Phytomedicine 2019, 55, 293–301. [Google Scholar] [CrossRef]
- Lee, J.; Chang, K.; Kim, G. Composition and anti-inflammatory activities of Zanthoxylum schinifolium essential oil: Suppression of inducible nitric oxide synthase, cyclooxygenase-2, cytokines and cellular adhesion. J. Sci. Food Agric. 2009, 89, 1762–1769. [Google Scholar] [CrossRef]
- Rufino, A.; Ribeiro, M.; Judas, F.; Salgueiro, L.; Lopes, M.; Cavaleiro, C.; Mendes, A. Anti-inflammatory and chondroprotective activity of (+)-α-pinene: Structural and enantiomeric selectivity. J. Nat. Prod. 2014, 77, 264–269. [Google Scholar] [CrossRef]
- Mazutti da Silva, S.M.; Rezende Costa, C.R.; Martins Gelfuso, G.; Silva Guerra, E.N. Wound healing effect of essential oil extracted from Eugenia dysenterica DC (Myrtaceae) leaves. Molecules 2019, 24, 2. [Google Scholar] [CrossRef] [Green Version]
- Woguem, V.; Fogang, H.; Maggi, F.; Tapondjou, L.; Womeni, H.; Quassinti, L.; Bramucci, M.; Vitali, L.; Petrelli, D.; Lupidi, G.; et al. Volatile oil from striped African pepper (Xylopia parviflora, Annonaceae) possesses notable chemopreventive, anti-inflammatory and antimicrobial potential. Food Chem. 2014, 149, 183–189. [Google Scholar] [CrossRef]
- Karkossa, I.; Raps, S.; Bergen, V.M.; Schubert, K. Systematic review of multi-omics approaches to investigate toxicological effects in macrophages. Int. J. Mol. Sci. 2020, 21, 9371. [Google Scholar] [CrossRef]
- Li, P.; Wang, Z.; Lam, S.M.; Shui, G. Rebaudioside A enhances resistance to oxidative stress and extends lifespan and healthspan in Caenorhabditis elegans. Antioxidants 2021, 10, 262. [Google Scholar] [CrossRef]
- Hotamisligil, G.S.; Davis, R.J. Cell signaling and stress responses. Cold Spring Harb. Perspect. Biol. 2016, 8, a006072. [Google Scholar] [CrossRef] [Green Version]
- Xing, B.; Bachstetter, A.D.; Van Eldik, L.J. Microglial p38α MAPK is critical for LPS-induced neuron degeneration, through a mechanism involving TNFα. Mol. Neurodegener. 2011, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Akira, S. Signaling to NF-κB by Toll-like receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [Green Version]
- Polednik, K.M.; Koch, A.C.; Felzien, L.K. Effects of essential oil from Thymus vulgaris on viability and inflammation in zebrafish embryos. Zebrafish 2018, 15, 361–371. [Google Scholar] [CrossRef]
- Rao, Z.; Xu, F.; Wen, T.; Wang, F.; Sang, W.; Zeng, N. Protective effects of essential oils from Rimulus cinnamon on endotoxin poisoning mice. Biomed. Pharmacother. 2018, 101, 304–310. [Google Scholar] [CrossRef]
- Wang, H.; Song, L.; Ju, W.; Wang, X.; Dong, L.; Zhang, Y.; Ya, P.; Yang, C.; Li, F. The acute airway inflammation induced by PM 2.5 exposure and the treatment of essential oils in Balb/c mice. Sci. Rep. 2017, 7, 44256. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Compounds 1 | Cal. RI 2 | Lit. RI 3 | Ref. | Relative Percentage (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| S1 | S2 | S3 | S4 | S5 | S6 | S7 | |||||
| 1 | α-Thujene | 920 | 924 | [22] | 0.28 | 0.23 | 0.17 | 0.24 | 0.24 | - | 0.04 |
| 2 | α-Pinene | 924 | 932 | [22] | 4.16 | 3.93 | 6.26 | 4.35 | 5.21 | 0.10 | 1.18 |
| 3 | Camphene | 941 | 946 | [22] | 0.15 | 0.11 | 0.21 | 0.14 | 0.24 | - | 0.03 |
| 4 | β-Pinene | 961 | 974 | [22] | 5.20 | 4.97 | 9.53 | 4.53 | 10.01 | 0.46 | 2.24 |
| 5 | β-Myrcene | 979 | 988 | [22] | 2.80 | 2.54 | 3.02 | 2.18 | 3.15 | 0.15 | 0.63 |
| 6 | α-Phellandrene | 994 | 1002 | [22] | 3.69 | 3.00 | 2.05 | 1.72 | 2.43 | 0.11 | 0.73 |
| 7 | 3-Carene | 1000 | 1008 | [22] | 0.21 | 0.18 | 0.16 | 0.14 | 0.17 | - | 0.03 |
| 8 | α-Terpinene | 1004 | 1014 | [22] | 0.35 | 0.12 | - | 0.05 | 0.19 | - | 0.04 |
| 9 | β-Phellandrene | 1015 | 1025 | [22] | 23.34 | 22.27 | 20.57 | 18.38 | 21.27 | 8.81 | 5.60 |
| 10 | (Z)-β-Ocimene | 1029 | 1032 | [22] | 0.25 | 0.39 | 0.09 | 0.23 | 0.59 | - | 0.18 |
| 11 | (E)-β-Ocimene | 1037 | 1044 | [22] | 0.61 | 0.52 | 0.99 | - | 2.40 | 0.04 | 0.52 |
| 12 | γ-Terpinene | 1044 | 1054 | [22] | 2.31 | 1.48 | 1.28 | 0.39 | 2.10 | 0.09 | 0.12 |
| 13 | Terpinolene | 1080 | 1086 | [22] | 0.29 | 0.14 | 0.32 | 0.13 | 0.35 | - | 0.11 |
| 14 | Linalool | 1095 | 1095 | [22] | - | - | 0.53 | - | 0.51 | - | 0.09 |
| 15 | Limonene oxide | 1106 | 1132 | [22] | - | - | - | - | 0.15 | 0.05 | - |
| 16 | (−)-endo-Fenchol | 1109 | 1114 | [22] | - | - | 0.06 | - | 0.11 | - | - |
| 17 | cis-p-Menth-2-en-1-ol | 1116 | 1118 | [22] | 0.44 | 0.31 | 0.73 | 0.67 | 0.73 | 0.80 | 0.29 |
| 18 | trans-p-Menth-2-en-1-ol | 1137 | 1140 | [22] | 0.37 | 0.36 | 0.56 | 0.52 | 0.69 | 0.66 | 0.26 |
| 19 | Pinocarvone | 1154 | 1160 | [22] | - | - | 0.15 | 0.11 | 0.31 | - | 0.05 |
| 20 | β-Pinene oxide | 1164 | 1154 | [22] | - | 0.50 | 0.33 | 0.05 | 0.47 | - | - |
| 21 | (−)-Terpinen-4-ol | 1170 | 1174 | [22] | 0.82 | 0.26 | 1.10 | 0.80 | 1.06 | 1.83 | 0.43 |
| 22 | Cryptone | 1176 | 1183 | [22] | 3.08 | 5.74 | 4.21 | 5.34 | 4.47 | 7.57 | 1.13 |
| 23 | α-Terpineol | 1185 | 1186 | [22] | - | - | - | - | - | - | 0.43 |
| 24 | (−)-Myrtenal | 1187 | 1195 | [22] | - | - | - | - | 0.70 | - | - |
| 25 | cis-Piperitol | 1192 | 1195 | [22] | 0.33 | - | 1.33 | 0.27 | 0.99 | 0.44 | - |
| 26 | (−)-Myrtenol | 1201 | 1194 | [22] | - | - | - | - | 0.17 | - | - |
| 27 | trans-Piperitol | 1206 | 1207 | [22] | 0.28 | 0.24 | 0.13 | - | - | - | 0.07 |
| 28 | 2-Pentylcyclopentanone | 1210 | 1208 | [23] | - | - | 0.27 | 0.20 | 0.26 | 0.36 | - |
| 29 | Lilac alcohol D | 1231 | 1232 | [24] | - | - | - | 0.07 | - | 0.19 | - |
| 30 | Cumin aldehyde | 1240 | 1238 | [22] | - | - | 0.18 | 0.31 | 0.23 | 0.47 | 0.05 |
| 31 | Carvone | 1244 | 1239 | [22] | 0.14 | 0.37 | - | 0.10 | - | - | - |
| 32 | Piperitone | 1255 | 1249 | [22] | 0.18 | 0.21 | - | 0.19 | 0.09 | 0.08 | - |
| 33 | Phellandral | 1264 | 1273 | [22] | 0.86 | 0.77 | 0.11 | - | - | 0.22 | 0.34 |
| 34 | p-Menth-1-en-7-al | 1267 | 1273 | [22] | - | - | 1.05 | 2.15 | 1.35 | 0.48 | 0.95 |
| 35 | Isobornyl acetate | 1279 | 1283 | [22] | 0.23 | - | 0.13 | 0.09 | 0.27 | 0.08 | 0.05 |
| 36 | 2-Caren-10-al | 1284 | 1289 | [25] | - | - | 0.12 | 0.08 | - | - | - |
| 37 | 2-Undecanone | 1291 | 1293 | [22] | 0.29 | 0.20 | - | - | - | - | - |
| 38 | δ-Elemene | 1327 | 1335 | [22] | - | - | 0.10 | - | 0.21 | - | 0.10 |
| 39 | α-Cubebene | 1337 | 1345 | [22] | 0.11 | - | 0.14 | 0.09 | 0.08 | 0.07 | 0.08 |
| 40 | α-Copaene | 1363 | 1374 | [22] | 0.34 | 0.29 | 0.35 | 0.39 | 0.26 | 0.36 | - |
| 41 | β-cubebene | 1376 | 1387 | [22] | - | 0.59 | - | 0.27 | 0.46 | 0.16 | 0.66 |
| 42 | β-Elemene | 1384 | 1389 | [22] | 2.99 | 3.01 | 3.76 | 2.12 | 3.33 | 2.21 | 4.97 |
| 43 | α-Cedrene | 1395 | 1410 | [22] | - | - | - | 0.42 | - | - | 0.11 |
| 44 | α-Gurjunene | 1396 | 1409 | [22] | 0.72 | 0.30 | 0.26 | - | 0.21 | 0.44 | - |
| 45 | β-Caryophyllene | 1407 | 1417 | [22] | 3.60 | 3.60 | 3.11 | 3.76 | 2.19 | 2.45 | 2.50 |
| 46 | β-Humulene | 1412 | 1436 | [22] | - | - | 0.08 | - | - | - | 0.28 |
| 47 | γ-Elemene | 1421 | 1434 | [22] | 0.07 | - | 0.27 | - | - | - | 0.51 |
| 48 | α-Bergamotene | 1425 | 1432 | [22] | - | - | - | - | 0.16 | - | - |
| 49 | α-guaiene | 1427 | 1437 | [22] | - | - | - | - | - | - | 0.44 |
| 50 | Tetradecane | 1430 | 1400 | [22] | - | - | - | - | - | - | 0.45 |
| 51 | α-Humulene | 1442 | 1452 | [22] | 12.38 | 12.19 | 12.36 | 12.66 | 8.77 | 11.58 | 5.03 |
| 52 | Aromadendrene | 1447 | 1441 | [22] | - | - | - | - | - | - | 0.16 |
| 53 | (E)-β-Farnesene | 1454 | 1454 | [22] | - | - | 0.16 | - | 0.08 | - | - |
| 54 | allo-Aromadendrene | 1467 | 1458 | [22] | - | 0.77 | 0.36 | 1.08 | 0.66 | 0.36 | 0.88 |
| 55 | Germacrene D | 1473 | 1484 | [22] | - | - | 0.23 | - | 0.17 | 2.40 | 1.46 |
| 56 | Pentadecane | 1478 | 1500 | [22] | - | 0.86 | - | - | 1.75 | - | 0.09 |
| 57 | β-selinene | 1482 | 1489 | [22] | 1.29 | 1.01 | 1.09 | 0.99 | - | 1.22 | 1.00 |
| 58 | Valencene | 1488 | 1496 | [22] | - | - | 0.95 | - | 0.46 | - | 0.82 |
| 59 | Elixene | 1490 | 1446 | [26] | 0.29 | 0.25 | - | - | 0.11 | 0.24 | 1.32 |
| 60 | 2-Tridecanone | 1492 | 1495 | [22] | - | - | - | 0.14 | - | - | - |
| 61 | (Z,E)-α-Farnesene | 1493 | 1500 | [27] | - | - | 0.15 | - | - | - | - |
| 62 | α-Selinene | 1498 | 1498 | [22] | 0.44 | 0.47 | - | 0.40 | 0.40 | 0.23 | 0.39 |
| 63 | β-Bisabolene | 1500 | 1505 | [22] | - | - | - | - | 0.52 | - | - |
| 64 | (E,E)-α-Farnesene | 1503 | 1505 | [22] | - | - | 0.25 | - | 0.21 | - | - |
| 65 | α-Panasinsen | 1513 | 1518 | [26] | 0.16 | 0.15 | - | 0.14 | - | 0.14 | 0.07 |
| 66 | δ-Cadinene | 1517 | 1522 | [22] | 0.33 | - | 0.83 | 0.35 | 0.38 | 0.18 | 0.54 |
| 67 | (Z)-Nerolidol | 1540 | 1531 | [22] | 0.14 | - | - | - | - | 0.04 | 0.15 |
| 68 | Germacrene B | 1557 | 1559 | [22] | - | - | 0.80 | 0.39 | 0.63 | 0.14 | 3.61 |
| 69 | Epiglobulol | 1562 | 1532 | [26] | - | - | - | - | - | 0.09 | - |
| 70 | (E)-Nerolidol | 1567 | 1561 | [22] | 0.09 | - | - | - | - | 0.07 | 0.49 |
| 71 | Caryophyllene oxide | 1582 | 1582 | [22] | 1.58 | 1.42 | 1.35 | 2.86 | 1.30 | 3.33 | 3.12 |
| 72 | Viridiflorol | 1591 | 1592 | [22] | - | - | - | - | 0.13 | 0.28 | - |
| 73 | Globulol | 1595 | 1590 | [22] | - | - | - | - | - | - | 0.27 |
| 74 | Tetradecanal | 1603 | 1612 | [22] | - | - | 0.33 | - | 0.26 | 0.79 | 0.09 |
| 75 | 2,5,9-Trimethylcycloundeca-4,8-dienone | 1609 | - | - | 3.31 | 3.61 | 3.64 | 7.12 | 2.95 | 8.09 | 1.80 |
| 76 | selina-6-en-4-ol | 1622 | 1624 | [26] | - | - | 0.10 | 0.12 | - | - | 0.36 |
| 77 | Diethyl phthalate | 1625 | 1639 | [28] | - | - | - | - | - | 0.24 | - |
| 78 | Cubenol | 1645 | 1645 | [22] | - | - | 0.25 | 0.46 | 0.10 | 0.18 | 1.71 |
| 79 | 1-Tetradecanol | 1662 | 1671 | [22] | - | 0.60 | - | 0.84 | 0.10 | 0.59 | 3.46 |
| 80 | α-Cadinol | 1668 | 1652 | [22] | 1.58 | 0.96 | 2.26 | 1.15 | 1.49 | 1.01 | 0.65 |
| 81 | β-Bisabolol | 1676 | 1674 | [22] | - | - | - | 0.42 | 0.23 | 0.43 | 0.99 |
| 82 | α-Eudesmol | 1678 | 1652 | [22] | - | - | - | - | - | 2.96 | - |
| 83 | 2-Hexyl-1-decanol | 1679 | 1673 | [29] | - | - | - | 1.72 | - | - | - |
| 84 | Heptadecane | 1680 | 1700 | [22] | 2.03 | 2.16 | - | - | - | - | - |
| 85 | α-Bisabolol | 1694 | 1685 | [22] | - | - | - | - | - | 0.49 | - |
| 86 | 2-pentadecanone | 1698 | 1697 | [22] | - | - | - | 0.06 | - | 0.08 | - |
| 87 | Pentadecanal | 1710 | 1714 | [30] | - | 0.22 | 0.22 | 0.36 | 0.79 | 0.29 | 0.97 |
| 88 | 6-Isopropenyl-4,8a-dimethyl-1,2,3,5,6,7,8,8a-octahydro-naphthalen-2-ol | 1714 | 1714 | [31] | - | 0.12 | 0.22 | 0.39 | - | - | 0.29 |
| 89 | 7R,8R-8-Hydroxy-4-isopropylidene-7-methylbicyclo[5.3.1]undec-1-ene | 1725 | 1726 | [32] | - | - | - | - | 0.20 | - | 0.67 |
| 90 | Isolongifolol | 1755 | 1729 | [22] | 1.45 | 2.08 | 2.68 | 2.55 | 2.03 | 2.56 | 1.18 |
| 91 | 1-Octadecene | 1777 | 1789 | [22] | - | - | - | - | - | 1.64 | - |
| 92 | (Z)-7-hexadecenal | 1790 | 1802 | [33] | - | - | - | 0.07 | - | 0.25 | 0.68 |
| 93 | Hexadecanal | 1813 | 1812 | [27] | - | - | - | - | - | - | 0.21 |
| 94 | 6,10,14-Trimethyl-2-pentadecanone | 1844 | 1846 | [30] | - | - | - | - | - | - | 0.04 |
| 95 | E-11(12-Cyclopropyl)dodecen-1-ol acetate | 1876 | - | - | - | - | - | - | - | - | 0.26 |
| 96 | Nonadecane | 1887 | 1900 | [22] | 0.25 | 0.24 | - | - | - | 0.29 | 0.45 |
| 97 | Linolenyl alcohol | 1897 | 1901 | [34] | - | 0.19 | - | - | - | - | - |
| 98 | (E,E)-farnesyl acetate | 1928 | 1919 | [30] | - | - | - | 0.33 | - | 0.32 | 0.14 |
| 99 | (Z,Z)-Geranyl linalool | 1958 | 1960 | [22] | - | - | - | - | - | 0.07 | 0.10 |
| 100 | α-Springene | 1974 | 1986 | [35] | - | - | - | - | - | 0.08 | 0.19 |
| 101 | Biformene | 1975 | 1987 | [36] | - | - | - | 0.09 | 0.07 | 0.11 | - |
| 102 | 1-Heptadecanol | 1977 | 1986 | [37] | - | - | - | - | - | 0.14 | - |
| 103 | Eicosane | 1982 | 2000 | [30] | 0.06 | - | - | - | - | 0.11 | 0.08 |
| 104 | Dibutyl phthalate | 1993 | 1962 | [31] | - | - | - | - | - | - | 0.43 |
| 105 | Heptadecanoic acid | 2022 | 2045 | [36] | - | - | - | 0.50 | 0.45 | - | 6.51 |
| 106 | (E,E)-Geranyl linalool | 2031 | 2026 | [22] | - | - | - | - | - | - | 0.58 |
| 107 | Henicosane | 2088 | 2100 | [22] | 0.48 | 0.38 | 0.10 | 0.14 | 0.22 | 0.69 | 0.50 |
| 108 | 13-Epimanool | 2099 | 2036 | [36] | - | - | - | - | 0.11 | - | 0.14 |
| 109 | Phytol | 2135 | 2156 | [32] | - | - | - | - | - | - | 0.11 |
| 110 | Docosane | 2197 | 2200 | [22] | - | 0.26 | 0.12 | - | - | 2.04 | 0.23 |
| 111 | Octadecanol acetate | 2250 | 2209 | [22] | - | - | - | - | - | 0.19 | 0.27 |
| 112 | cis-9-Tricosene | 2289 | 2298 | [27] | - | - | - | 0.08 | 0.19 | - | 0.03 |
| 113 | Tricosane | 2331 | 2300 | [22] | - | - | - | - | 2.48 | - | - |
| 114 | Tetracosane | 2420 | 2400 | [22] | 4.25 | 4.01 | 1.15 | 1.38 | - | 6.94 | 3.55 |
| 115 | Trans-squalene | 2806 | 2810 | [36] | - | - | - | 0.64 | - | - | - |
| 116 | 1-Hexacosanol | 2840 | 2852 | [30] | - | - | - | - | - | 2.49 | - |
| Total (%) | 88.60 | 88.58 | 93.31 | 87.91 | 95.05 | 82.05 | 71.76 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Z.; Xie, L.; Xu, Y.; Zhao, K.; Li, X.; Zhong, J.; Lu, Y.; Xu, X.; Goodin, S.; Zhang, K.; et al. Essential Oils from Zingiber striolatum Diels Attenuate Inflammatory Response and Oxidative Stress through Regulation of MAPK and NF-κB Signaling Pathways. Antioxidants 2021, 10, 2019. https://doi.org/10.3390/antiox10122019
Huang Z, Xie L, Xu Y, Zhao K, Li X, Zhong J, Lu Y, Xu X, Goodin S, Zhang K, et al. Essential Oils from Zingiber striolatum Diels Attenuate Inflammatory Response and Oxidative Stress through Regulation of MAPK and NF-κB Signaling Pathways. Antioxidants. 2021; 10(12):2019. https://doi.org/10.3390/antiox10122019
Chicago/Turabian StyleHuang, Zebin, Lingna Xie, Yongyu Xu, Kai Zhao, Xuetong Li, Jiaben Zhong, Yujing Lu, Xuetao Xu, Susan Goodin, Kun Zhang, and et al. 2021. "Essential Oils from Zingiber striolatum Diels Attenuate Inflammatory Response and Oxidative Stress through Regulation of MAPK and NF-κB Signaling Pathways" Antioxidants 10, no. 12: 2019. https://doi.org/10.3390/antiox10122019